The Influence of Canopy Cover on the Ecological Function of A Key Autogenic Ecosystem Engineer

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Investigation of Species Associations with Natural Variations in Canopy Cover of Hormosira banksii
2.3. Experimental Investigation of the Effect of Variations in Canopy Cover of Hormosira banksii on the Physical Environment and Associated Species Assemblages
2.4. Data Analysis
3. Results
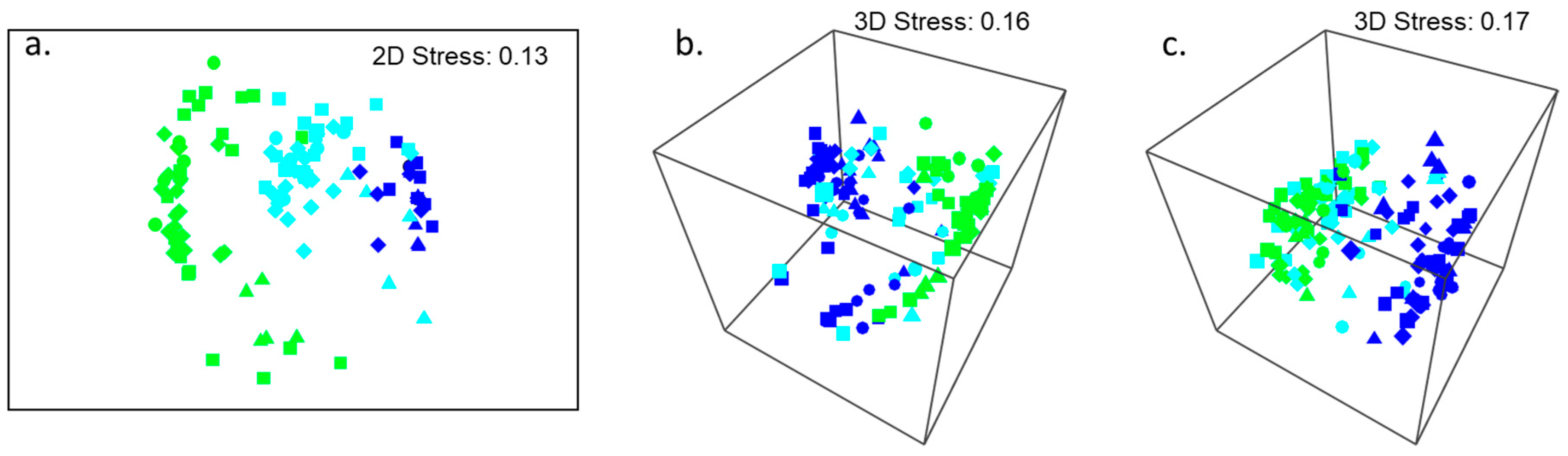
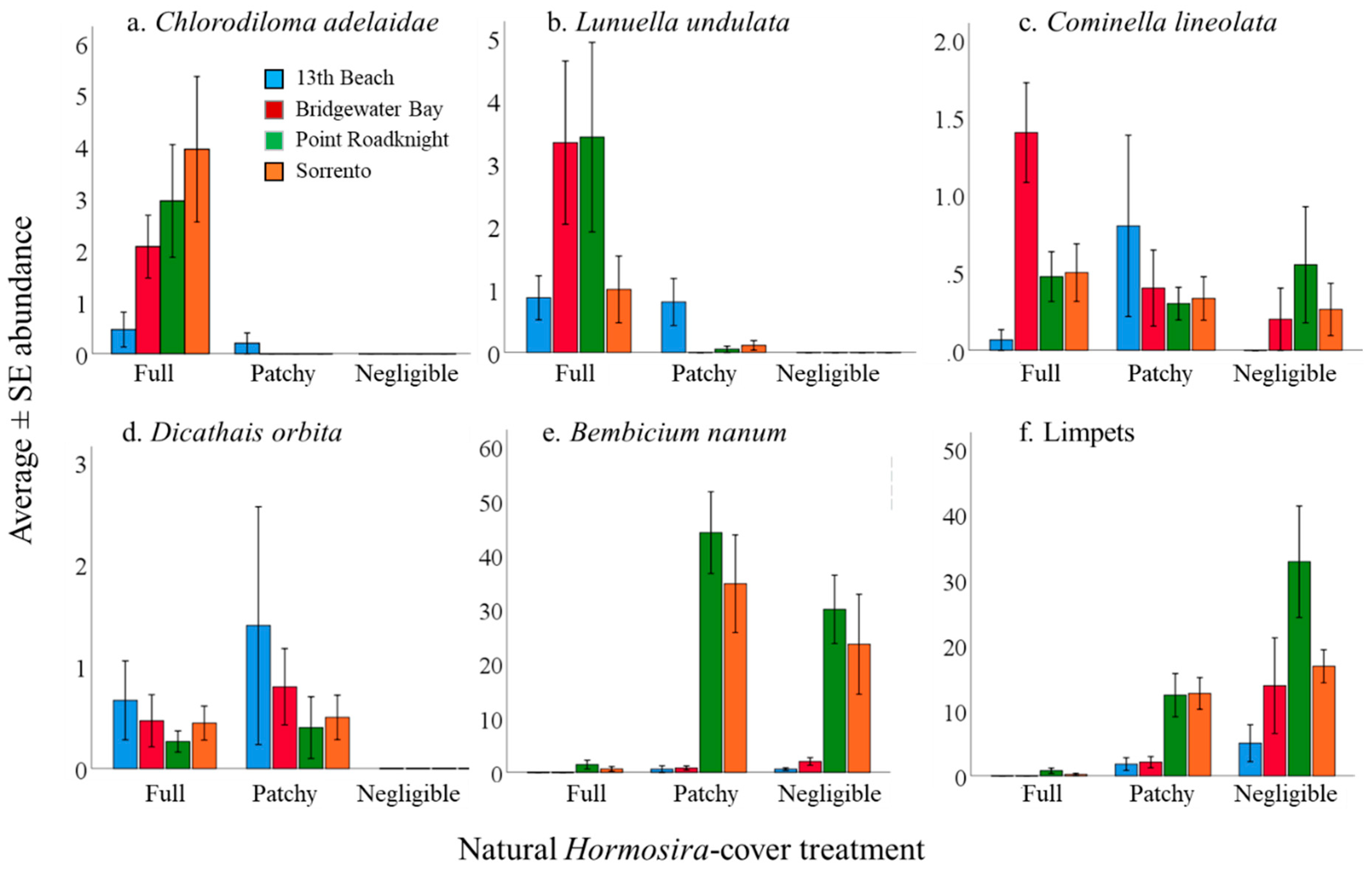
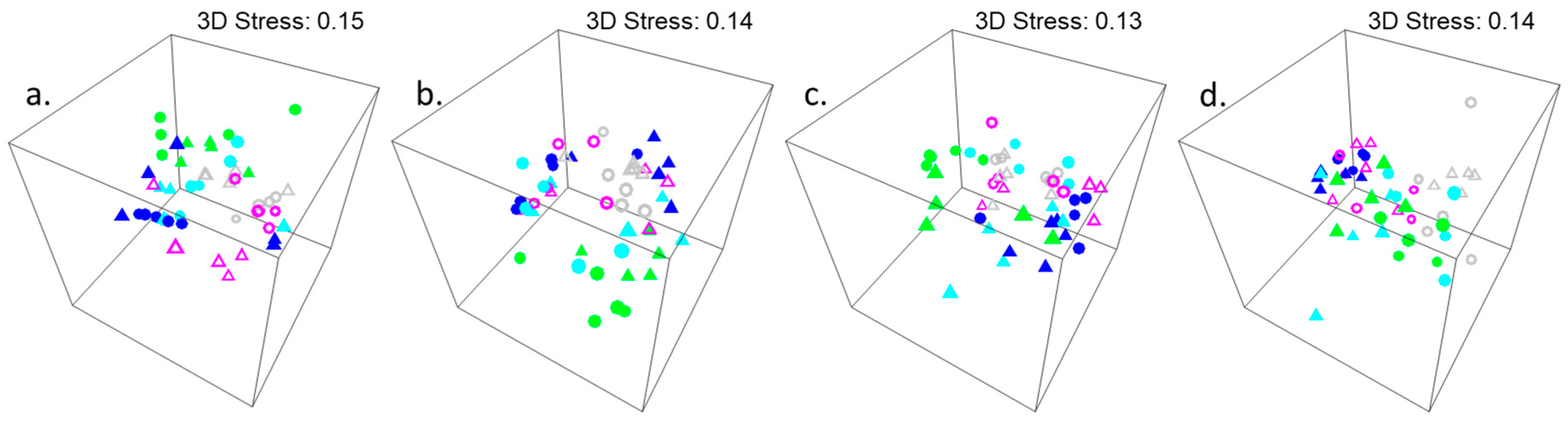
3.1. Investigation of Species Associations with Natural Variations in Canopy Cover of Hormosira banksii
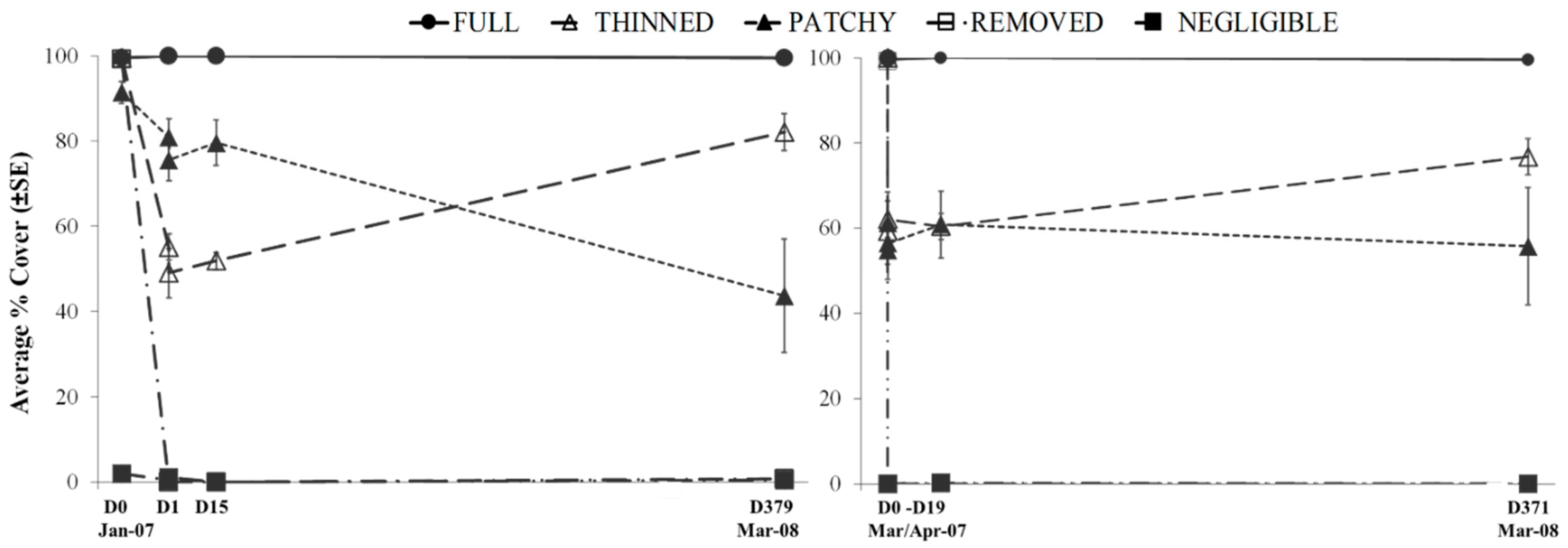
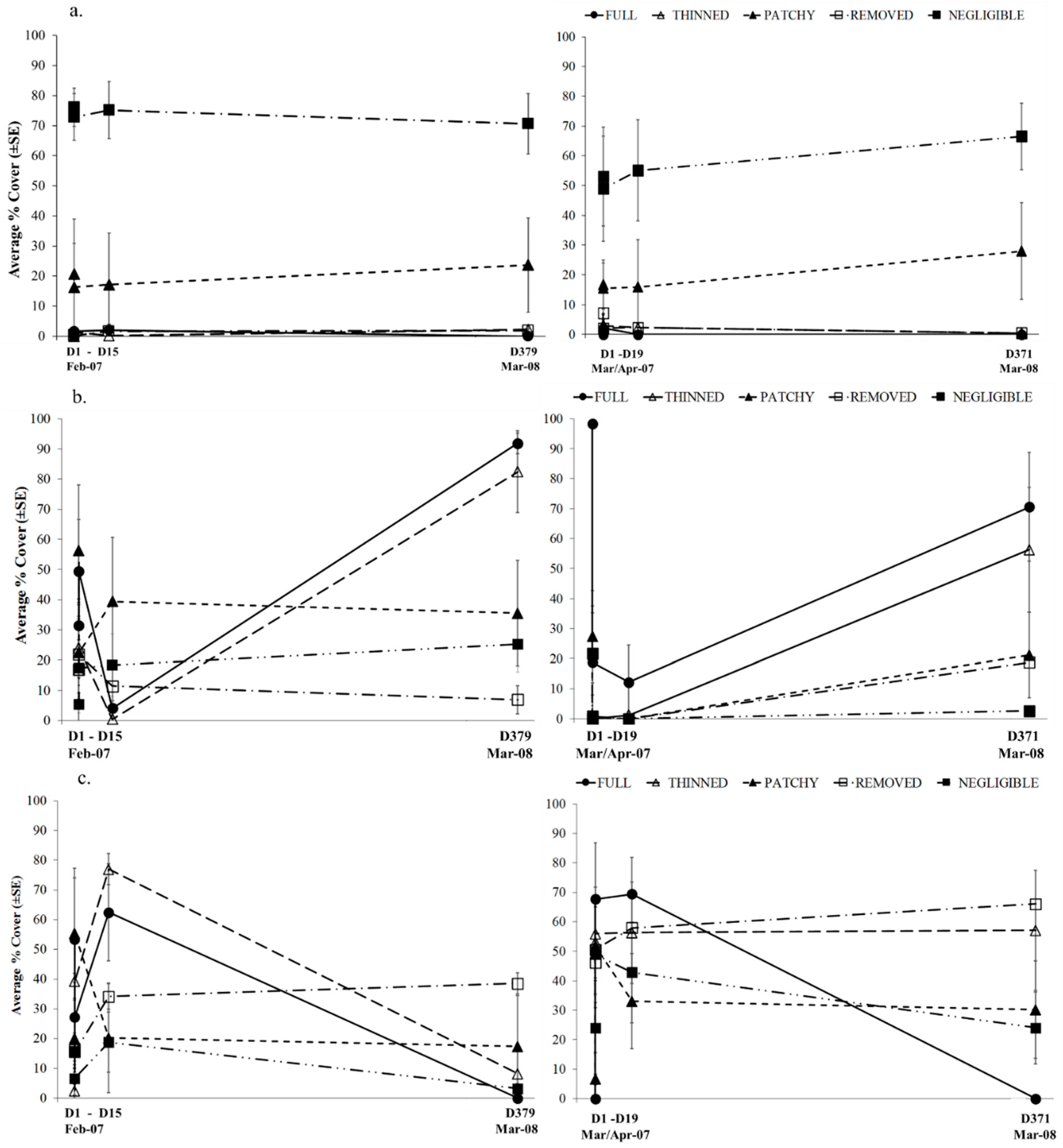
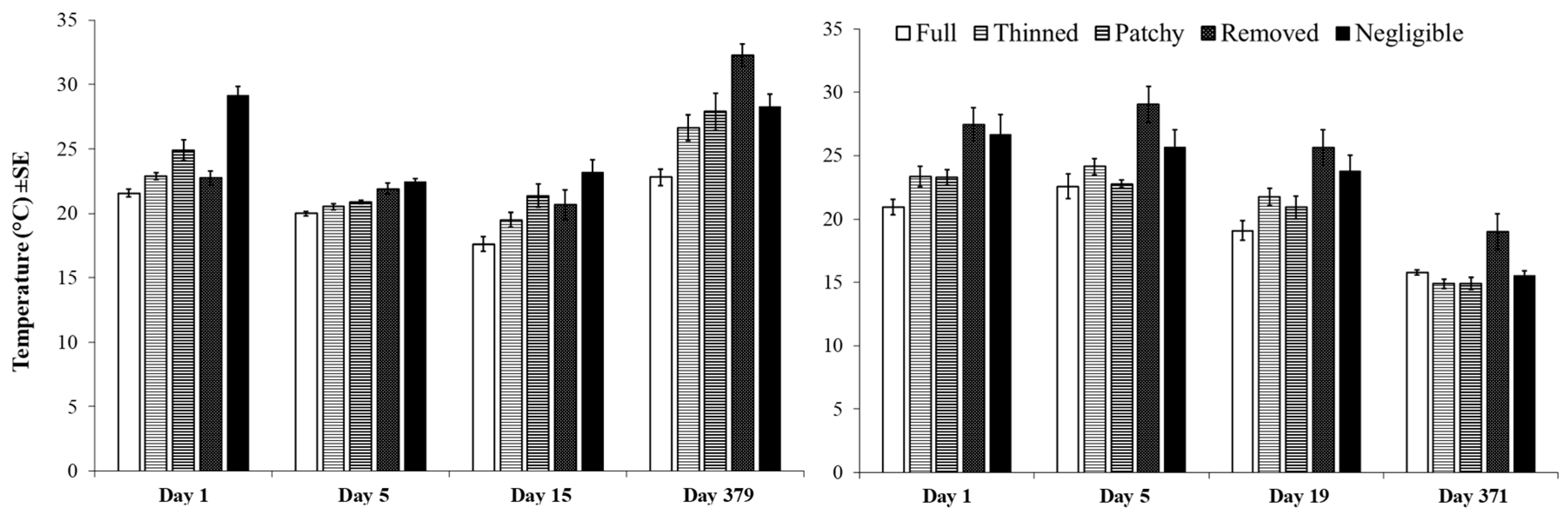
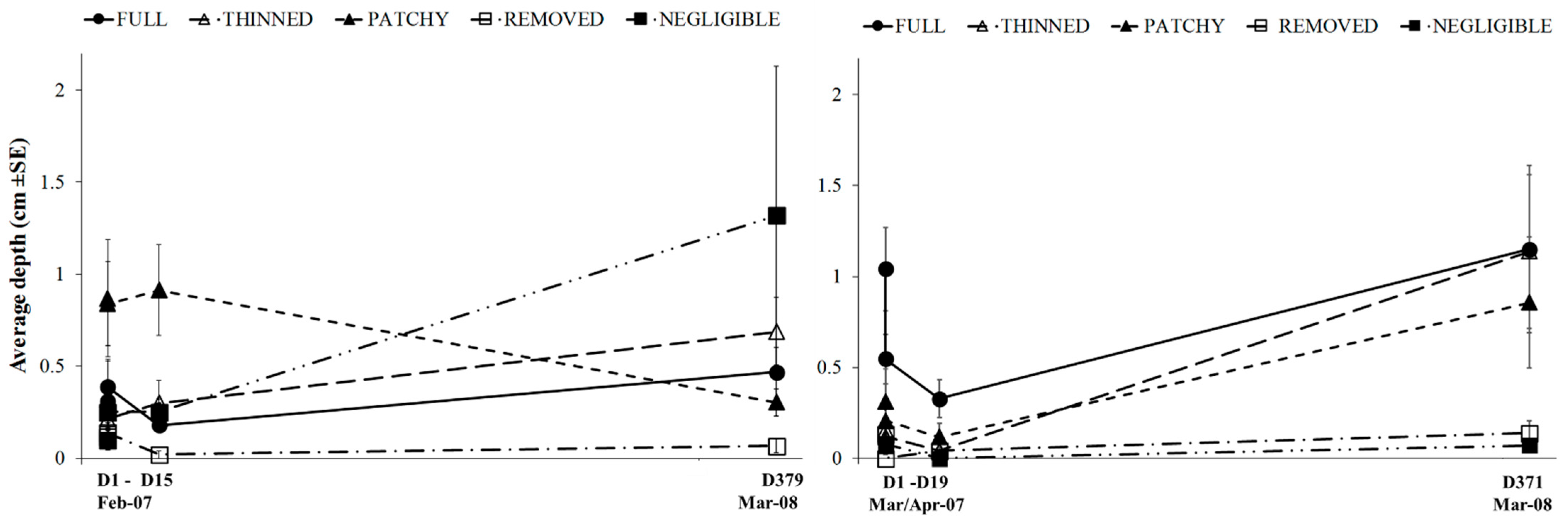
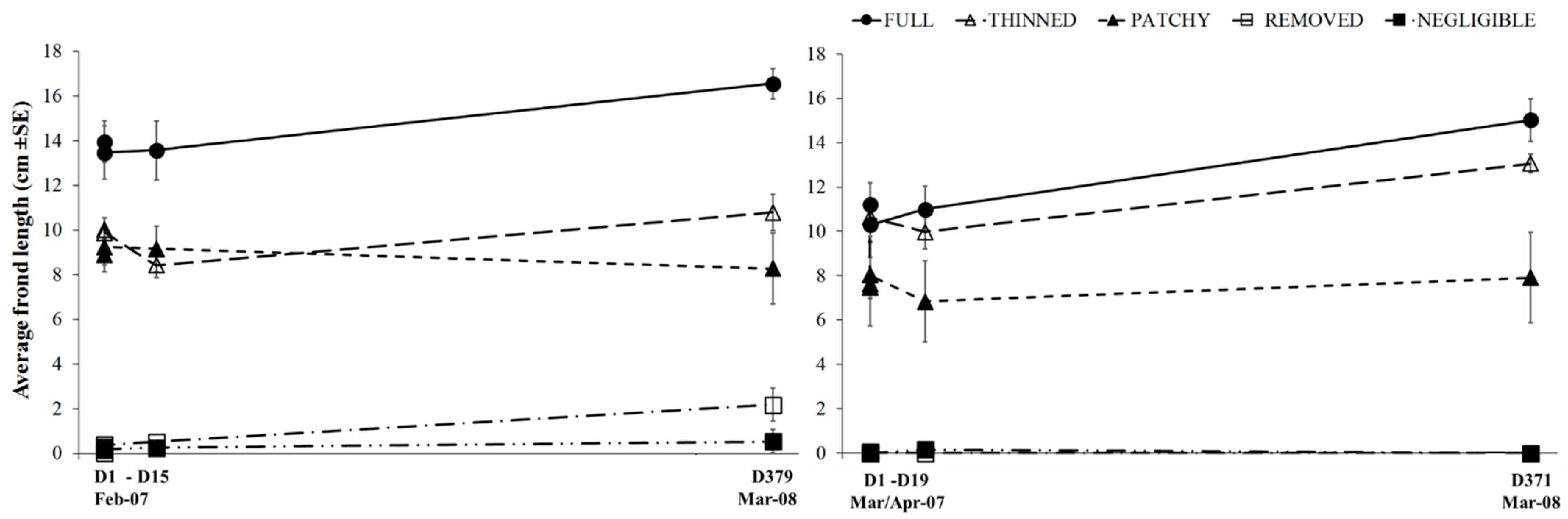
3.2. Experimental Investigation of the Effect of Variations in Canopy Cover of Hormosira banksii on the Physical Environment and Associated Species Assemblages
4. Discussion
4.1. Hormosira banksii as An Ecosystem Engineer
4.2. Species Associations with Cover
4.3. Alternate Habitat Types and Stable States
4.4. The Influence of Sand
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bridgewater Bay | 13th Beach | Sorrento Back Beach | Point Roadknight | |
|---|---|---|---|---|
| Primary Algae | Capreolia implexa Cladophora sp. Coralline Turf Hormosira banksii Scytosiphon lomentaria Ulva sp. | Capreolia implexa Ceramium sp. Cladophora sp. Colpomenia sinuosa Coralline Turf Encrusting Coralline Hormosira banksii Sargassum sp. Laurencia filiformis Laurencia spp. Ulva sp. Unidentified Reds | Capreolia implexa Cladophora sp. Coralline Turf Hormosira banksii Laurencia sp. Rivularia sp. Ulva sp. | Capreolia implexa Coralline turf Diplocladia patersonis Encrusting Coralline Hormosira banksii Notheia anomala Scytosiphon lomentaria Ulva sp. |
| Understorey Algae ‘Coralline Turf’ is likely Corallina officinalis but due to compact form was not able to be positively identified in the field. | Caulocystis cephalornithos Ceramium sp. Chaetomorpha sp. Cladophora sp. Colpomenia sinuosa Coralline Turf Diplocladia patersonis Encrusting Coralline Gelidium australe Laurencia botryoides Laurencia filiformis Laurencia spp. Notheia anomala Scytosiphon lomentaria Sargassum sp. Ulva sp. Unidentified Encrusting Black Unidentified Epiphytes Unidentified Reds | Encrusting green Capreolia implexa Caulerpa brownii Ceramium sp. Chaetomorpha sp. Cladophora sp. Colpomenia sinuosa Coralline Encrusting Coralline Turf Cystophora spp. Laurencia botryoides Laurencia filiformis Laurencia spp. Notheia anomala Pachydictyon sp. Sargassum sp. Ulva sp. Unidentified Reds | Capreolia implexa Ceramium sp. Cladophora sp. Cystophora sp. Coralline Turf Diplocladia patersonis Encrusting Coralline Laurencia sp. Rivularia sp. Ulva sp. | Capreolia implexa Ceramium sp. Cladophora sp. Coralline Turf Diplocladia patersonis Encrusting Coralline Laurencia sp. Rivularia sp. Splachnidium rugosum Unidentified Reds |
| Invertebrates | ||||
| Bivalves | Brachidontes rostratus Xenostrobus pulex Unidentified Bivalve | Xenostrobus pulex Unidentified Bivalve | Xenostrobus pulex | Xenostrobus pulex |
| Polycheates | Galeolaria caespitosa | Galeolaria caespitosa | Galeolaria caespitosa | Galeolaria caespitosa |
| Limpets (and false limpets) | Cellana tramoserica Lottia mixta Montfortula rugosa Notoacmea alta Notoacmea flammea Onchidella nigricans Patelloida alticostata Patelloida latistrigata Siphonaria diemenensis Siphonaira zelandica Juvenile limpets (unidentified) | Cellana tramoserica Lottia mixta Notoacmea flammea Notoacmea sp. Onchidella nigricans Patelloida alticostata Siphonaria diemenensis Siphonaria zelandica Juvenile limpets (unidentified) | Cellana tramoserica Lottia mixta Notoacmea alta Notoacmea flammea Notoacmea petterdi Notoacmea sp. Patelloida alticostata Patelloida latistrigata Siphonaria diemenensis Siphonaria funiculata Siphonaria zelandica Juvenile limpets (unidentified) | Cellana tramoserica Montfortula rugosa Notoacmea sp. Notoacmea flammea Notoacmea petterdi Patella peroni Patelloida alticostata Patelloida latistrigata Siphonaria diemenensis Siphonaria zelandica Siphonaria funiculata Juvenile limpets (unidentified) |
| Gastropods | Austrocochlea constricta Austrocochlea porcata Austrolittornia unifasciata Bembicium nanum Cantharidius pulcherimus Chlorodiloma adelaidea Chlorodiloma odontis Cominella lineolata Dicathais orbita Littorinid juvenile (unidentified) Mitre sp. Phasianotrochus eximus Thalotia conica Lunella undulata | Austrocochlea constricta Bembicium nanum Cantharidius pulcherimus Chlorodiloma adelaidea Chlorodiloma odontis Cominella lineolata Cystophora sp. Dentrimitrella pulla Dicathais orbita Lepsellia vinosa Littorinid juvenile (unidentified) Mitre cabonaria Phasianotrochus eximus Thalotia conica Lunella undulata Waimatea obscura | Austrocochlea constricta Austrolittorina unifasciata Bembicium nanum Chlorodiloma adelaidae Cominella lineolata Dicathais orbita Littorinid juvenile (unidentified) Nerita atramentosa Lunella undulata | Austrocochlea constricta Austrocochlea juvenile Bembicium nanum Chlorodiloma adelaidae Cominella lineolata Dicathis orbita Lepsiella vinosa Littorinid juvenile (unidentified) Mitre sp. Nerita atramentosa Lunella undulata |
| Echinoderms | Meridiastra calcar Parvulastra exigua | Meridiastra calcar | Meridiastra calcar Parvulastra exigua | |
| Crustaceans | Chthalaumus antennatus Paragrapsus quadradentatus Guinusia chabrus Naxia aurita Unidentified Amphipods Unidentified Copepods Unidentified Harpactocoids Unidentified Isopods | Chthalaumus antennatus Naxia aurita Nectocarcinus turbculosa Paragrapsus quadradentatus Guinusia chabrus Unidentified Amphipods Unidentified Copepods Unidentified Harpactocoids Unidentified Isopods | Cthalaumus antennatus Paragrapsus quadradentatus Paragrapsus sp. Unidentified Isopods Unidentified Amphipods | Chthalaumus antennatus Paragrapsus quadradentatus Unidentified Isopods |
| Anemones | Actinia tenebrosa Anthothoe albocinta Aulactinia verartra Oulactis mucosa Oulactis sp. Unidentified anemones | Anthothoe albocinta Oulactis mucosa Oulactis sp. | Oulactis mucosa Oulactis sp. Unidentified Anemone | Actinia tenebrosa Aulactinia verartra |
| Chitons | Unidentified Chiton | Unidentified Chiton | Unidentified Chiton |


| Negligible, Full | Negligible, Patchy | Negligible, Removed | Negligible, Thinned | Full, Patchy | Full, Removed | Full, Thinned | Patchy, Removed | Patchy, Thinned | Removed, Thinned | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 1 % contribution | Average dissimilarity | 85.69 | 86.79 | 92.19 | 95.80 | 71.50 | 93.79 | 86.46 | 89.44 | 86.46 | 79.29 |
| Sand | 31.06 | 24.75 | 11.39 | 41.64 | 34.32 | 35.03 | 23.11 | 35.03 | 34.13 | ||
| Bare rock | 24.28 | 25.10 | 14.49 | 16.62 | 23.92 | 13.05 | 23.92 | ||||
| Xenostrobus pulex | 22.07 | 21.22 | 20.56 | 33.77 | |||||||
| Capreolia implexa | 10.43 | 12.93 | 12.93 | ||||||||
| Capreolia implexa and Xenostrobus pulex complex | 15.06 | 13.71 | 14.11 | 13.99 | |||||||
| Hormosira holdfasts | 18.10 | 17.87 | 18.77 | 34.13 | |||||||
| Day 5 % contribution | Average dissimilarity | 83.06 | 68.07 | 76.89 | 83.50 | 67.48 | 65.42 | 56.82 | 63.20 | 58.44 | 58.43 |
| Sand | 10.01 | 12.00 | |||||||||
| Bare rock | 24.26 | 26.68 | 22.74 | 23.80 | 26.54 | 14.69 | 23.86 | 29.54 | 27.27 | ||
| Xenostrobus pulex | 11.70 | 10.59 | |||||||||
| Capreolia implexa | 10.21 | ||||||||||
| Capreolia implexa and Xenostrobus pulex complex | 11.00 | 10.24 | |||||||||
| Hormosira holdfasts | 14.32 | 14.17 | 14.59 | 15.47 | |||||||
| Encrusting Coralline | 10.68 | 11.11 | |||||||||
| Day 15/19 % contribution | Average dissimilarity | 89.42 | 89.29 | 89.11 | 87.75 | 77.87 | 76.06 | 63.50 | 82.10 | 79.80 | 70.73 |
| Sand | 10.92 | 16.44 | 12.35 | 13.83 | |||||||
| Bare rock | 44.76 | 28.51 | 33.73 | 43.19 | 37.61 | 52.35 | 55.86 | 35.59 | 49.55 | 55.25 | |
| Xenostrobus pulex | 20.14 | 24.31 | 20.61 | 23.00 | |||||||
| Capreolia implexa | |||||||||||
| Capreolia implexa and Xenostrobus pulex complex | 15.31 | 14.15 | 15.42 | 14.66 | |||||||
| Hormosira holdfasts | 15.20 | 17.96 | 17.70 | 15.68 | |||||||
| Encrusting Coralline | 10.99 | 10.70 | 13.59 | ||||||||
| Day 371/379 % contribution | Average dissimilarity | 74.77 | 65.21 | 74.84 | 70.44 | 66.23 | 82.67 | 43.72 | 73.20 | 61.17 | 68.96 |
| Sand | 15.79 | 13.54 | 13.56 | 23.91 | 16.27 | 11.50 | 21.08 | 16.26 | |||
| Bare rock | 15.20 | 10.61 | 11.50 | 16.20 | 15.49 | 10.70 | 13.56 | 13.36 | 13.76 | ||
| Xenostrobus pulex | 10.58 | 12.59 | |||||||||
| Capreolia implexa | 12.29 | 10.50 | 13.04 | ||||||||
| Capreolia implexa and Xenostrobus pulex complex | 18.14 | 20.89 | 12.81 | 17.40 | |||||||
| Hormosira holdfasts | 10.21 | 11.71 | 17.40 | 12.54 | |||||||
| Encrusting Coralline | 11.18 | 15.70 | 11.15 | ||||||||
| Cladophora sp. | 10.37 | 13.85 | |||||||||
| Galeolaria caespitosa | 13.07 | ||||||||||
| Coralline Turf | 11.72 |
| Negligible, Full | Negligible, Patchy | Negligible, Removed | Negligible, Thinned | Full, Patchy | Full, Removed | Full, Thinned | Patchy, Removed | Patchy, Thinned | Removed, Thinned | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 1 % contribution | Average dissimilarity | 100 | 87.08 | 95.15 | 100 | 87.82 | 97.60 | 84.51 | 94.22 | 78.41 | 92.31 |
| Bembicium nanum | 13.37 | 14.73 | 24.13 | 17.73 | |||||||
| Juvenile littorinid | 13.57 | 19.84 | 14.48 | ||||||||
| Chlorodiloma adelaidae | 11.98 | 13.35 | 21.69 | 15.77 | 13.32 | 10.33 | |||||
| Dicathais orbita | 10.37 | 14.64 | 19.22 | 15.71 | 35.69 | ||||||
| Lunella undulata | 15.06 | 14.35 | 12.40 | ||||||||
| Siphonaria diemenensis | 13.24 | ||||||||||
| Cominella lineolata | 12.95 | ||||||||||
| Day 5 % contribution | Average dissimilarity | 97.22 | 87.20 | 96.04 | 100 | 86.40 | 99.46 | 90.93 | 97.98 | 96.60 | 100 |
| Bembicium nanum | 14.76 | 16.95 | 30.93 | 25.31 | 12.16 | 10.15 | |||||
| Juvenile littorinid | 15.52 | 18.26 | 35.55 | 27.24 | |||||||
| Chlorodiloma adelaidae | 12.33 | 10.02 | |||||||||
| Dicathais orbita | 12.99 | 12.01 | 12.33 | 10.50 | 23.19 | 19.25 | |||||
| Lunella undulata | 10.98 | 12.40 | 11.00 | 13.02 | |||||||
| Oulactis mucosa | 15.90 | 10.34 | 14.59 | ||||||||
| Amphipoda | 23.80 | ||||||||||
| Anthothoe albocinta | 11.90 | ||||||||||
| Day 15/19 % contribution | Average dissimilarity | 99.51 | 89.83 | 97.03 | 94.90 | 81.65 | 96.39 | 85.72 | 95.73 | 86.69 | 95.48 |
| Bembicium nanum | 10.09 | 19.65 | 13.43 | 16.17 | |||||||
| Juvenile littorinid | 14.06 | 14.74 | 24.87 | 19.41 | |||||||
| Chlorodiloma adelaidae | 11.82 | 18.18 | 16.94 | 10.86 | |||||||
| Dicathais orbita | 11.30 | 15.35 | 15.35 | 15.90 | |||||||
| Lunella undulata | 10.41 | 14.44 | 15.99 | 14.96 | 20.61 | 16.87 | |||||
| Juvenile limpet | 18.09 | 13.77 | |||||||||
| Cominella lineolata | 12.45 | 13.20 | 11.47 | 18.27 | |||||||
| Aulactinia veratra | 10.20 | 10.45 | |||||||||
| Day 371/379 % contribution | Average dissimilarity | 100 | 84.84 | 83.78 | 97.99 | 87.81 | 97.15 | 79.51 | 88.77 | 85.52 | 89.96 |
| Bembicium nanum | 14.24 | 15.96 | 13.47 | 15.11 | |||||||
| Juvenile littorinid | 21.25 | 25.36 | 21.07 | 21.59 | |||||||
| Chlorodiloma adelaidae | 11.43 | 17.01 | 12.97 | 23.57 | 15.62 | 10.04 | |||||
| Dicathais orbita | 11.19 | 11.56 | 12.29 | 16.75 | 12.73 | 14.11 | |||||
| Lunella undulata | 12.95 | 18.66 | 14.55 | 20.70 | |||||||
| Siphonaria diemenensis | 11.00 | 10.40 | 11.55 | 11.67 | |||||||
| Cominella lineolata | 11.70 | 13.89 | 15.47 | ||||||||
| Juvenile limpet | 14.67 | 11.15 | 14.71 | 12.26 | |||||||
| Thalotia conica | 10.95 |


References
- Menge, B.A. Predation Intensity in a Rocky Intertidal Community: Effect of an algal canopy, wave action and desiccation on predator feeding rates. Oecologia 1978, 34, 17–35. [Google Scholar] [CrossRef]
- Povey, A.; Keough, M.J. Effects of trampling on plant and animal populations on rocky shores. Oikos 1991, 61, 355–368. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Alper, J. Ecosystem ‘Engineers’ Shape Habitats for Other Species. Science 1998, 280, 1195–1196. [Google Scholar] [CrossRef]
- Keough, M.J.; Quinn, G.P. Effects of periodic disturbances from trampling on rocky intertidal algal beds. Ecol. Appl. 1998, 8, 141–161. [Google Scholar]
- Schiel, D.R.; Taylor, D.I. Effects of trampling on a rocky intertidal algal assemblage in southern New Zealand. J. Exp. Mar. Biol. Ecol. 1999, 235, 213–235. [Google Scholar]
- Jenkins, S.R.; Norton, T.A.; Hawkins, S.J. Long term effects of Ascophyllum nodosum canopy removal on mid shore community structure. J. Mar. Biol. Assoc. UK 2004, 84, 327–329. [Google Scholar] [CrossRef]
- Dudgeon, S.; Petraitis, P.S. First year demography of the foundation species, Ascophyllum nodosum, and its community implications. Oikos 2005, 109, 405–415. [Google Scholar] [CrossRef]
- Lilley, S.A.; Schiel, D.R. Community effects following the deletion of a habitat-forming alga from rocky marine shores. Oecologia 2006, 148, 672–681. [Google Scholar] [CrossRef]
- Schiel, D.R. Rivets or bolts? When single species count in the function of temperate rocky reef communities. J. Exp. Mar. Biol. Ecol. 2006, 338, 233–252. [Google Scholar] [CrossRef]
- Hastings, A.; Byers, J.E.; Crooks, J.A.; Cuddington, K.; Jones, C.G.; Lambrinos, J.G.; Talley, T.S.; Wilson, W.G. Ecosystem engineering in space and time. Ecol. Lett. 2007, 10, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Schiel, D.; Lilley, S. Gradients of disturbance to an algal canopy and the modification of an intertidal community. Mar. Ecol. Prog. Ser. 2007, 339, 1–11. [Google Scholar] [CrossRef]
- Benedetti Cecchi, L.; Cinelli, F. Canopy removal experiments in Cystoseira-dominated rockpools from the Western coast of the Mediterranean (Ligurian Sea). J. Exp. Mar. Biol. Ecol. 1992, 155, 69–83. [Google Scholar] [CrossRef]
- Cervin, G.; Åberg, P. Do littorinids affect the survival of Ascophyllum nodosum germlings? J. Exp. Mar. Biol. Ecol. 1997, 218, 35–47. [Google Scholar] [CrossRef]
- Pavia, H.; Carr, H.; Åberg, P. Habitat and feeding preferences of crustacean mesoherbivores inhabiting the brown seaweed Ascophyllum nodosum (L.) Le Jol. and its epiphytic macroalgae. J. Exp. Mar. Biol. Ecol. 1999, 236, 15–32. [Google Scholar] [CrossRef]
- Coleman, M.A.; Clark, J.S.; Doblin, M.A.; Bishop, M.J.; Kelaher, B.P. Genetic differentiation between estuarine and open coast ecotypes of a dominant ecosystem engineer. Mar. Freshw. Res. 2018. [Google Scholar] [CrossRef]
- Wootton, H.F.; Keough, M.J. Disturbance type and intensity combine to affect resilience of an intertidal community. Mar. Ecol. Prog. Ser. 2016, 560, 121–133. [Google Scholar] [CrossRef]
- Tait, L.W.; Schiel, D.R. Legacy Effects of Canopy Disturbance on Ecosystem Functioning in Macroalgal Assemblages. PLoS ONE 2011, 6, e26986. [Google Scholar] [CrossRef] [PubMed]
- Raffaelli, D. The grazer-algae interaction in the intertidal zone of New Zealand rocky shores. J. Exp. Mar. Biol. Ecol. 1979, 38, 81–100. [Google Scholar] [CrossRef]
- Underwood, A.J. The vertical distribution and seasonal abundance of intertidal macroalgae on a rocky shore in New South Wales. J. Exp. Mar. Biol. Ecol. 1984, 78, 199–220. [Google Scholar] [CrossRef]
- King, A. Monitoring and Management of Human Activity on Rocky Shores. Ph.D. Thesis, Melbourne University, Melbourne, Australia, 1992. [Google Scholar]
- Bellgrove, A. Recruitment of Intertidal Macroalgae on a Wave-Exposed Rocky Coast; Monash Clayton: Melbourne, Australia, 1998. [Google Scholar]
- Ralph, P.J.; Morrison, D.A.; Addison, A. A quantitative study of the patterns of morphological variation within Hormosira banksii (Turner) Decaisne (Fucales: Phaeophyta) in south-eastern Australia. J. Exp. Mar. Biol. Ecol. 1998, 225, 285–300. [Google Scholar] [CrossRef]
- Underwood, A.J. Grazing and disturbance: An experimental analysis of patchiness in recovery from a severe storm by the intertidal alga Hormosira banksii on rocky shores in New South Wales. J. Exp. Mar. Biol. Ecol. 1998, 231, 291–306. [Google Scholar] [CrossRef]
- Underwood, A.J. Physical disturbances and their direct effect on an indirect effect: Responses of an intertidal assemblage to a severe storm. J. Exp. Mar. Biol. Ecol. 1999, 232, 125–140. [Google Scholar] [CrossRef]
- Pocklington, J.B. Positive Interactions on Rocky Seashores. Are They Important, or Can They Be Ignored? Honours Thesis, Flinders University of South Australia, Adelaide, Australia, 2003. [Google Scholar]
- Lilley, S.A. Removal of Habitat-Forming Species and the Consequences on Community Biodiversity in New Zealand Rocky Shore Ecosystems. Master’s Thesis, Canterbury University, Christchurch, New Zealand, 2004. [Google Scholar]
- Macinnis-Ng, C.M.O.; Morrison, D.A.; Ralph, P.J. Temporal and spatial variation in the morphology of the brown macroalga Hormosira banksii (Fucales, Phaeophyta). Bot. Mar. 2005, 48, 198–207. [Google Scholar] [CrossRef]
- Addison, P.F.E.; Koss, R.S.; O’Hara, T.D. Recreational use of a rocky intertidal reef in Victoria: Implications for ecological research and management. Aust. J. Environ. Manag. 2008, 15, 169–179. [Google Scholar] [CrossRef]
- Schoenwaelder, M.E.A. Physode distribution and the effect of ‘Thallus sunburn’ in Hormosira banksii (Fucales, Phaeophyceae). Bot. Mar. 2002, 45, 262–266. [Google Scholar] [CrossRef]
- Bishop, M.J.; Morgan, T.; Coleman, M.A.; Kelaher, B.P.; Hardstaff, L.K.; Evenden, R.W. Facilitation of molluscan assemblages in mangroves by the fucalean alga Hormosira banksii. Mar. Ecol. Prog. Ser. 2009, 392, 111–122. [Google Scholar] [CrossRef]
- Short, A.D. Beaches of the Victorian Coast & Port Phillip Bay: A Guide to Their Nature, Characteristics, Surf and Safety; University of Sydney & Surf Life Saving Victoria: Sydney, Australia, 2006. [Google Scholar]
- Bird, E. The Coast of Victoria: The Shaping of Scenery; Melbourne University Press: Carlton, Australia, 1993. [Google Scholar]
- Kruskal, J.B. Non-parametric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E Limited: Plymouth, UK, 2008. [Google Scholar]
- Bellgrove, A.; Clayton, M.N.; Quinn, G.P. An integrated study of the temporal and spatial variation in the supply of propagules, recruitment and assemblages of intertidal macroalgae on a wave-exposed rocky coast, Victoria, Australia. J. Exp. Mar. Biol. Ecol. 2004, 310, 207–225. [Google Scholar] [CrossRef]
- Gemelli, F.; Johnson, C.R.; Wright, J.T. Gastropod communities associated with different morphologies of the intertidal seaweed Hormosira banksii. Mar. Freshw. Res. 2019, 70, 280–291. [Google Scholar] [CrossRef]
- Bellgrove, A.; McKenzie, P.; McKenzie, J.; Sfiligoj, B. Restoration of the habitat-forming fucoid alga Hormosira banksii at effluent-affected sites: Competitive exclusion by coralline turfs. Mar. Ecol. Prog. Ser. 2010, 419, 47–56. [Google Scholar] [CrossRef]
- Underwood, A.J. A refutation of critical tidal levels as determinates of the structure of intertidal communities on British shores. J. Exp. Mar. Biol. Ecol. 1978, 33, 185–202. [Google Scholar] [CrossRef]
- Petraitis, P.S.; Latham, R.E. The Importance of Scale in Testing the Origins of Alternative Community States. Ecology 1999, 80, 429–442. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Bagur, M.; Palomo, M. Algal Epibionts as Co-Engineers in Mussel Beds: Effects on Abiotic Conditions and Mobile Interstitial Invertebrates. Diversity 2019, 11, 17. [Google Scholar] [CrossRef]
- Airoldi, L.; Hawkins, S.J. Negative effects of sediment deposition on grazing activity and survival of the limpet Patella vulgata. Mar. Ecol. Prog. Ser. 2007, 332, 235–240. [Google Scholar] [CrossRef]
- McKenzie, P.F.; Bellgrove, A. Dislodgment and attachment strength of the intertidal macroalga Hormosira banksii (Fucales, Phaeophyceae). Phycologia 2009, 48, 335–343. [Google Scholar] [CrossRef]
- Schiel, D.R.; Wood, S.A.; Dunmore, R.A.; Taylor, D.I. Sediment on rocky intertidal reefs: Effects on early post-settlement stages of habitat-forming seaweeds. J. Exp. Mar. Biol. Ecol. 2006, 331, 158–172. [Google Scholar] [CrossRef]
- Pocklington, J.B.; Jenkins, S.R.; Bellgrove, A.; Keough, M.J.; O’Hara, T.D.; Masterson-Algar, P.E.; Hawkins, S.J. Disturbance alters ecosystem engineering by a canopy-forming alga. J. Mar. Biol. Assoc. UK 2018, 98, 687–698. [Google Scholar] [CrossRef]
- Airoldi, L. The effects of sedimentation on rocky coast assemblages. In Oceanography and Marine Biology, An Annual Review; Gibson, R.N., Atkinson, R.J.A., Eds.; Aberdeen University Press/Allen & Unwin: London, UK, 2003; Volume 41, pp. 161–236. [Google Scholar]









| Factor | Time × Site × Treatment df = 12 | Time × Treatment df = 12 | Time × Site df = 12 | Time df = 3 | Site × Treatment df = 4 | Treatment df = 4 | Site df = 1 |
|---|---|---|---|---|---|---|---|
| Hormosira banksii canopy cover | MSR = 179.473 p = 0.011 | MSR = 588.142 p = 0.0001 * | n.s | n.s | n.s | MSR = 76,844.5 p = 0.0001 | n.s |
| Species Richness | n.s | MSR = 12.151 p = 0.0001 * | n.s | MSR = 63.880 p = 0.0001 * | n.s | n.s | n.s |
| UNDERSTOREY COVER | |||||||
| Xenostrobus pulex & Capriola implexa | n.s | n.s | n.s | n.s | n.s | MSR = 30,174.2 p = 0.0001 | n.s |
| Bare Rock | MSR = 1563.12 p = 0.002 * | MSR = 2127.67 p = 0.0001 * | n.s | MSR = 7219.06 p = 0.0001 * | n.s | n.s | MSR = 12,059.0 p = 0.009 |
| Sand | MSR = 1275.09 p = 0.037 * | MSR = 2983.68 p = 0.0001 * | n.s | MSR = 10,801.3 p = 0.0001 * | n.s | MSR = 10,821.3 p = 0.0001 | n.s |
| MOBILE INVERTEBRATES | |||||||
| Lunella undulata† | MSR = 0.611 p = 0.0001 * | MSR = 0.445 p = 0.003 * | MSR = 0.657 p = 0.009 * | n.s | MSR = 4.386 p = 0.001 | MSR = 8.106 p = 0.001 | MSR = 2.948 p = 0.001 |
| Chlorodiloma adelaidae† | n.s | n.s | n.s | n.s | MSR = 5.445 p = 0.002 | MSR = 15.755 p = 0.0001 | n.s |
| Dicathais orbita† | n.s | MSR = 0.596 p = 0.004 * | n.s | n.s | n.s | MSR = 3.880 p = 0.0001 | MSR = 1.348 p = 0.035 |
| Bembicium nanum† | n.s | MSR = 0.866 p = 0.004 * | n.s | n.s | n.s | MSR = 8.752 p = 0.0001 | n.s |
| PHYSICAL | |||||||
| Temperature | MSR = 9.834 p = 0.0001 * | MSR = 11.296 p = 0.0001 * | MSR = 589.470 p = 0.0001 * | MSR = 87.398 p = 0.0001 * | MSR = 27.602 p = 0.021 | MSR = 156.467 p = 0.0001 | MSR = 112.350 p = 0.001 |
| Light intensity | n.s | n.s | MSR = 5111.26 p = 0.0001 * | MSR = 9461.34 p = 0.0001 * | n.s | MSR = 40,416.1 p = 0.0001 | MSR = 10,353.6 p = 0.0001 |
| COVARIATES | |||||||
| Frond length | n.s | MSR = 6.528 p = 0.015 * | n.s | MSR = 22.376 p = 0.0001 * | n.s | MSR = 1371.70 p = 0.0001 | MSR = 31.113 p = 0.037 |
| Sand depth | MSR = 0.512 p = 0.02 * | n.s | n.s | MSR = 1.377 p = 0.004 * | MSR = 1.149 p = 0.016 | MSR = 1.653 p = 0.002 | n.s |
| a. Understorey Cover | ||||||||
| 13th Beach | Day 1 | Day 5 | Day 15 | Day 379 | ||||
| Groups | t | P(MC) | t | P(MC) | t | P(MC) | t | P(MC) |
| Negligible, Full | 2.7355 | 0.005 | 3.1076 | 0.001 | 3.4761 | 0.003 | 3.0716 | 0.002 |
| Negligible, Patchy | 2.261 | 0.011 | 2.0689 | 0.016 | 2.556 | 0.007 | 1.6355 | 0.048 |
| Negligible, Removed | 3.7294 | 0.001 | 4.1516 | 0.002 | 3.8215 | 0.002 | 4.1695 | 0.001 |
| Negligible, Thinned | 2.999 | 0.002 | 2.3286 | 0.004 | 2.7993 | 0.005 | 2.7044 | 0.002 |
| Full, Patchy | 1.32 | 0.175 | 1.7143 | 0.061 | 1.5254 | 0.099 | 1.4554 | 0.112 |
| Full, Removed | 1.7988 | 0.037 | 2.4151 | 0.007 | 2.684 | 0.005 | 5.3762 | 0.001 |
| Full, Thinned | 1.4372 | 0.124 | 1.2142 | 0.23 | 1.4212 | 0.144 | 1.3216 | 0.178 |
| Patchy, Removed | 2.2744 | 0.01 | 1.954 | 0.026 | 1.974 | 0.017 | 2.3982 | 0.007 |
| Patchy, Thinned | 1.5898 | 0.094 | 0.28847 | 0.956 | 1.4853 | 0.108 | 1.0035 | 0.4 |
| Removed, Thinned | 2.1327 | 0.02 | 1.6114 | 0.082 | 2.0228 | 0.035 | 3.4919 | 0.001 |
| Bridgewater Bay | Day 1 | Day 5 | Day 19 | Day 371 | ||||
| Groups | t | P(MC) | t | P(MC) | t | P(MC) | t | P(MC) |
| Negligible, Full | 3.9928 | 0.001 | 3.472 | 0.003 | 4.3075 | 0.001 | 3.3129 | 0.003 |
| Negligible, Patchy | 2.2683 | 0.01 | 1.791 | 0.055 | 2.9745 | 0.007 | 0.99608 | 0.416 |
| Negligible, Removed | 2.6292 | 0.002 | 2.8406 | 0.005 | 5.0578 | 0.002 | 1.8123 | 0.029 |
| Negligible, Thinned | 3.6341 | 0.003 | 3.4366 | 0.003 | 3.5378 | 0.001 | 2.0145 | 0.017 |
| Full, Patchy | 3.0028 | 0.004 | 1.9309 | 0.043 | 1.9964 | 0.031 | 2.4889 | 0.009 |
| Full, Removed | 3.1564 | 0.002 | 2.5595 | 0.007 | 2.0738 | 0.04 | 2.8531 | 0.005 |
| Full, Thinned | 4.3495 | 0.001 | 1.3148 | 0.199 | 1.857 | 0.08 | 1.0017 | 0.399 |
| Patchy, Removed | 1.4045 | 0.14 | 1.5003 | 0.111 | 1.7736 | 0.053 | 1.3559 | 0.127 |
| Patchy, Thinned | 2.3606 | 0.013 | 1.2967 | 0.187 | 1.7255 | 0.072 | 1.3792 | 0.15 |
| Removed, Thinned | 1.0678 | 0.347 | 1.5106 | 0.099 | 0.47141 | 0.848 | 1.7502 | 0.033 |
| b. Mobile Invertebrates | ||||||||
| 13th Beach | Day 1 | Day 5 | Day 15 | Day 379 | ||||
| Groups | t | P(MC) | t | P(MC) | t | P(MC) | t | P(MC) |
| Negligible, Full | 2.1068 | 0.004 | 2.4702 | 0.002 | 2.35 | 0.003 | 2.7648 | 0.004 |
| Negligible, Patchy | 2.3273 | 0.003 | 2.3418 | 0.005 | 2.7097 | 0.001 | 2.5555 | 0.001 |
| Negligible, Removed | 3.0379 | 0.002 | 6.046 | 0.001 | 3.7486 | 0.001 | 2.8849 | 0.002 |
| Negligible, Thinned | 3.6249 | 0.001 | 2.7132 | 0.003 | 2.4678 | 0.005 | 2.7919 | 0.005 |
| Full, Patchy | 1.1599 | 0.263 | 1.3268 | 0.128 | 0.80682 | 0.659 | 0.87774 | 0.495 |
| Full, Removed | 1.993 | 0.015 | 3.0928 | 0.003 | 1.9719 | 0.008 | 2.0366 | 0.015 |
| Full, Thinned | 1.982 | 0.016 | 1.6499 | 0.058 | 0.60027 | 0.817 | 0.95089 | 0.452 |
| Patchy, Removed | 2.0558 | 0.02 | 3.2691 | 0.002 | 2.005 | 0.041 | 1.7523 | 0.04 |
| Patchy, Thinned | 1.3835 | 0.141 | 1.8072 | 0.03 | 1.0276 | 0.407 | 1.1334 | 0.315 |
| Removed, Thinned | 3.0643 | 0.006 | 1.512 | 0.154 | 1.6727 | 0.074 | 1.6586 | 0.076 |
| Bridgewater Bay | Day 1 | Day 5 | Day 19 | Day 371 | ||||
| Groups | t | P(MC) | t | P(MC) | t | P(MC) | t | P(MC) |
| Negligible, Full | 3.8373 | 0.002 | 3.3296 | 0.002 | 2.8513 | 0.002 | 2.8735 | 0.001 |
| Negligible, Patchy | 1.7435 | 0.028 | 2.0275 | 0.016 | 1.5883 | 0.052 | 0.84361 | 0.554 |
| Negligible, Removed | 2.2146 | 0.02 | 2.2748 | 0.01 | 1.8333 | 0.028 | 1.0569 | 0.366 |
| Negligible, Thinned | 2.2084 | 0.003 | 2.1693 | 0.005 | 1.3721 | 0.111 | 1.7979 | 0.02 |
| Full, Patchy | 2.5234 | 0.003 | 1.9401 | 0.021 | 2.0477 | 0.013 | 1.9566 | 0.016 |
| Full, Removed | 2.4472 | 0.006 | 2.3379 | 0.003 | 2.291 | 0.019 | 1.9659 | 0.017 |
| Full, Thinned | 1.436 | 0.111 | 1.6196 | 0.069 | 1.8522 | 0.015 | 1.3402 | 0.174 |
| Patchy, Removed | 1.8608 | 0.026 | 1.5259 | 0.107 | 1.7104 | 0.03 | 0.92669 | 0.524 |
| Patchy, Thinned | 1.3634 | 0.132 | 1.4108 | 0.123 | 1.1969 | 0.231 | 0.98985 | 0.419 |
| Removed, Thinned | 1.5459 | 0.048 | 1.2201 | 0.226 | 1.1947 | 0.24 | 1.2061 | 0.209 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pocklington, J.B.; Keough, M.J.; O’Hara, T.D.; Bellgrove, A. The Influence of Canopy Cover on the Ecological Function of A Key Autogenic Ecosystem Engineer. Diversity 2019, 11, 79. https://doi.org/10.3390/d11050079
Pocklington JB, Keough MJ, O’Hara TD, Bellgrove A. The Influence of Canopy Cover on the Ecological Function of A Key Autogenic Ecosystem Engineer. Diversity. 2019; 11(5):79. https://doi.org/10.3390/d11050079
Chicago/Turabian StylePocklington, Jacqueline B., Michael J. Keough, Tim D. O’Hara, and Alecia Bellgrove. 2019. "The Influence of Canopy Cover on the Ecological Function of A Key Autogenic Ecosystem Engineer" Diversity 11, no. 5: 79. https://doi.org/10.3390/d11050079
APA StylePocklington, J. B., Keough, M. J., O’Hara, T. D., & Bellgrove, A. (2019). The Influence of Canopy Cover on the Ecological Function of A Key Autogenic Ecosystem Engineer. Diversity, 11(5), 79. https://doi.org/10.3390/d11050079