Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea

 ,
, 

 and
and
Abstract
1. Introduction
2. Materials and Methods
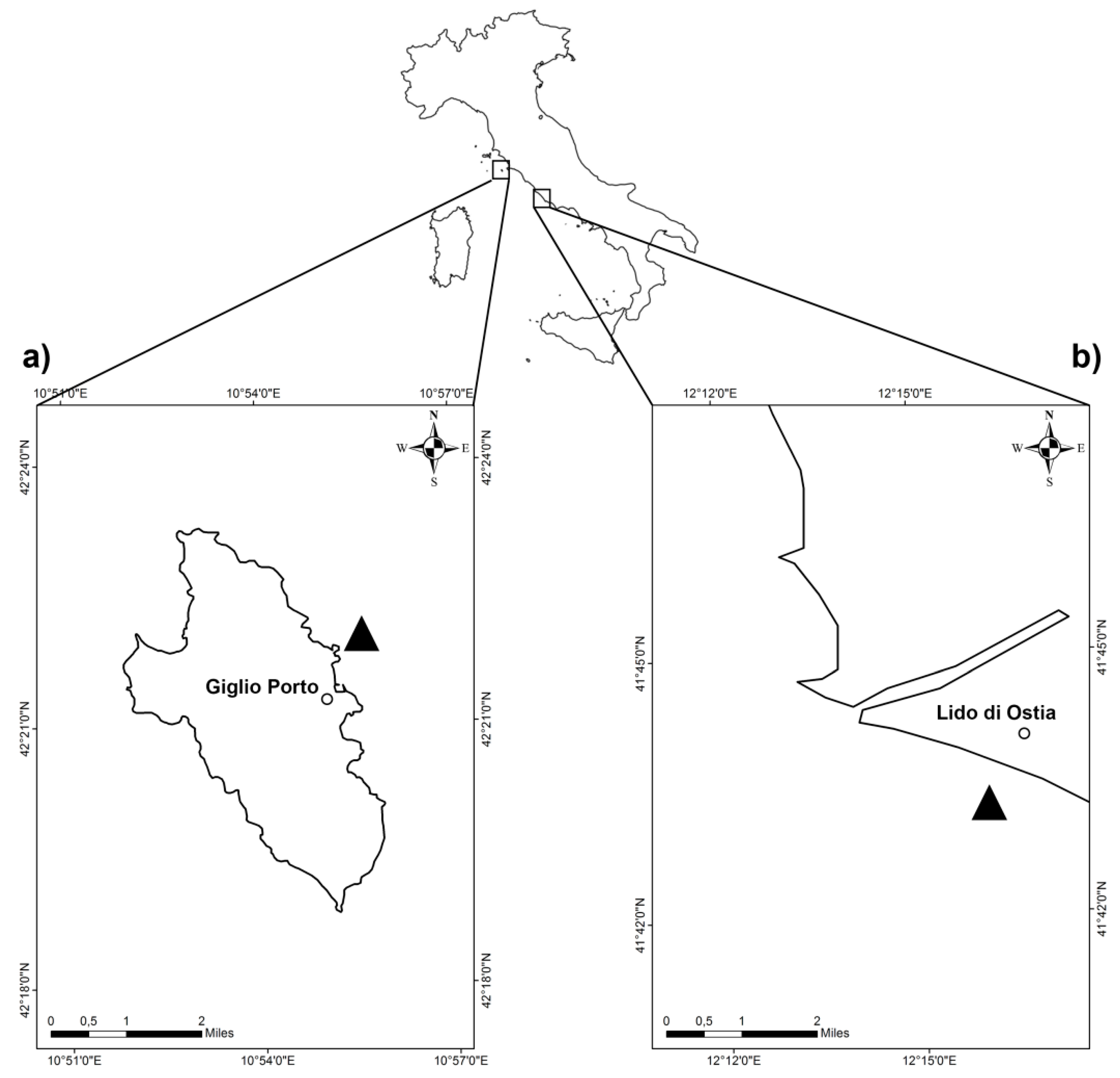
2.1. Study Area and Sampling Methods
2.2. Relation to the Substrate and Trophic Groups
2.3. Data Analysis
3. Results
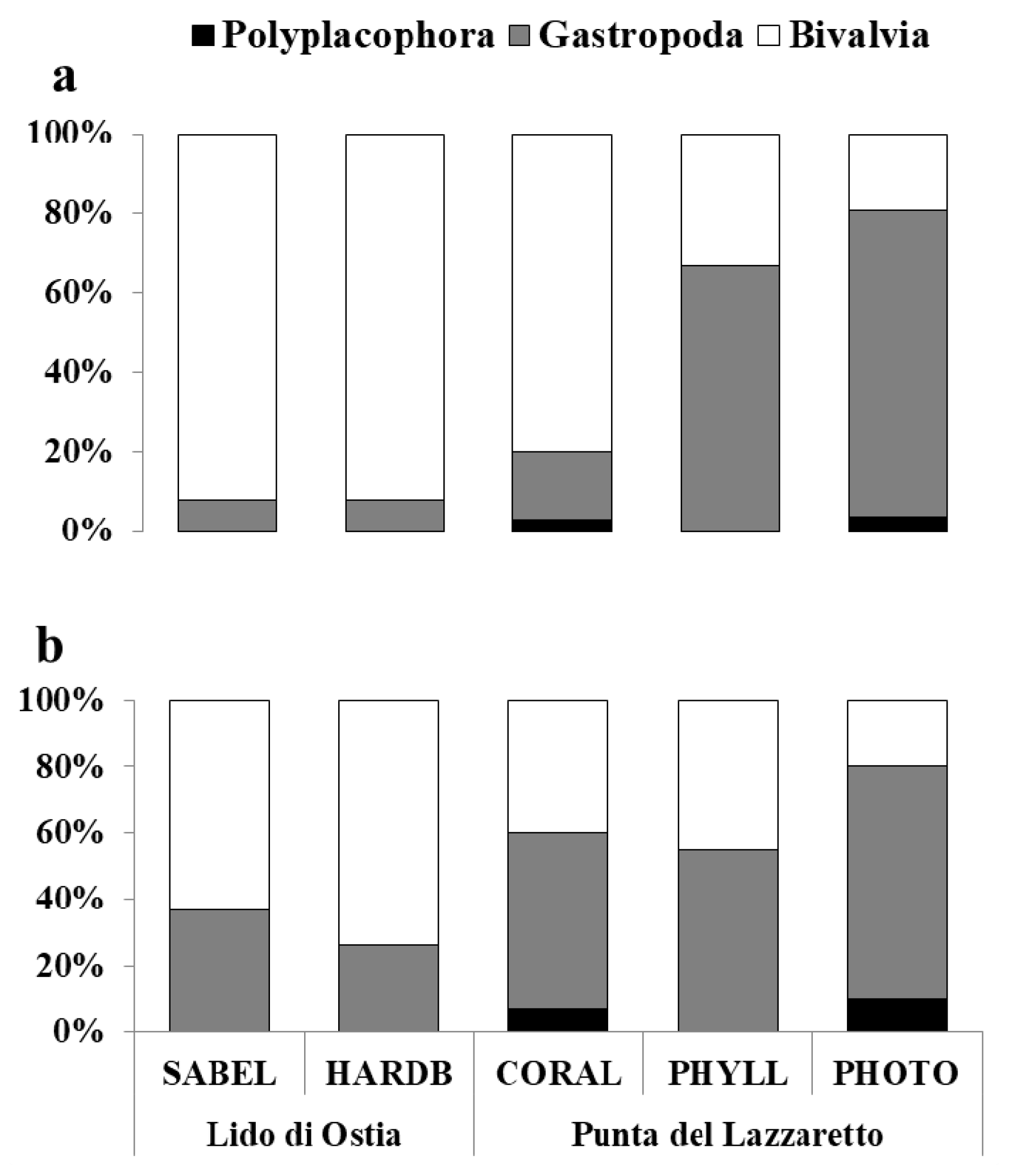
3.1. Composition of Mollusc Assemblages
3.2. Species-Substrate Relationship
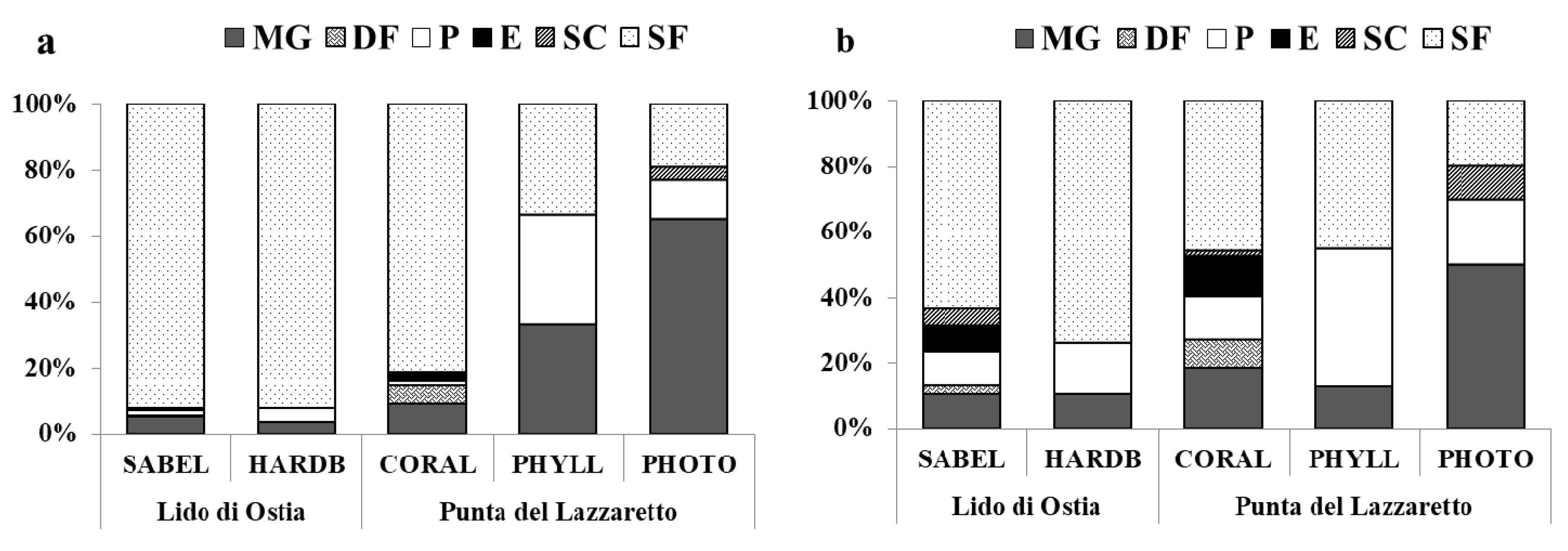
3.3. Trophic Analysis
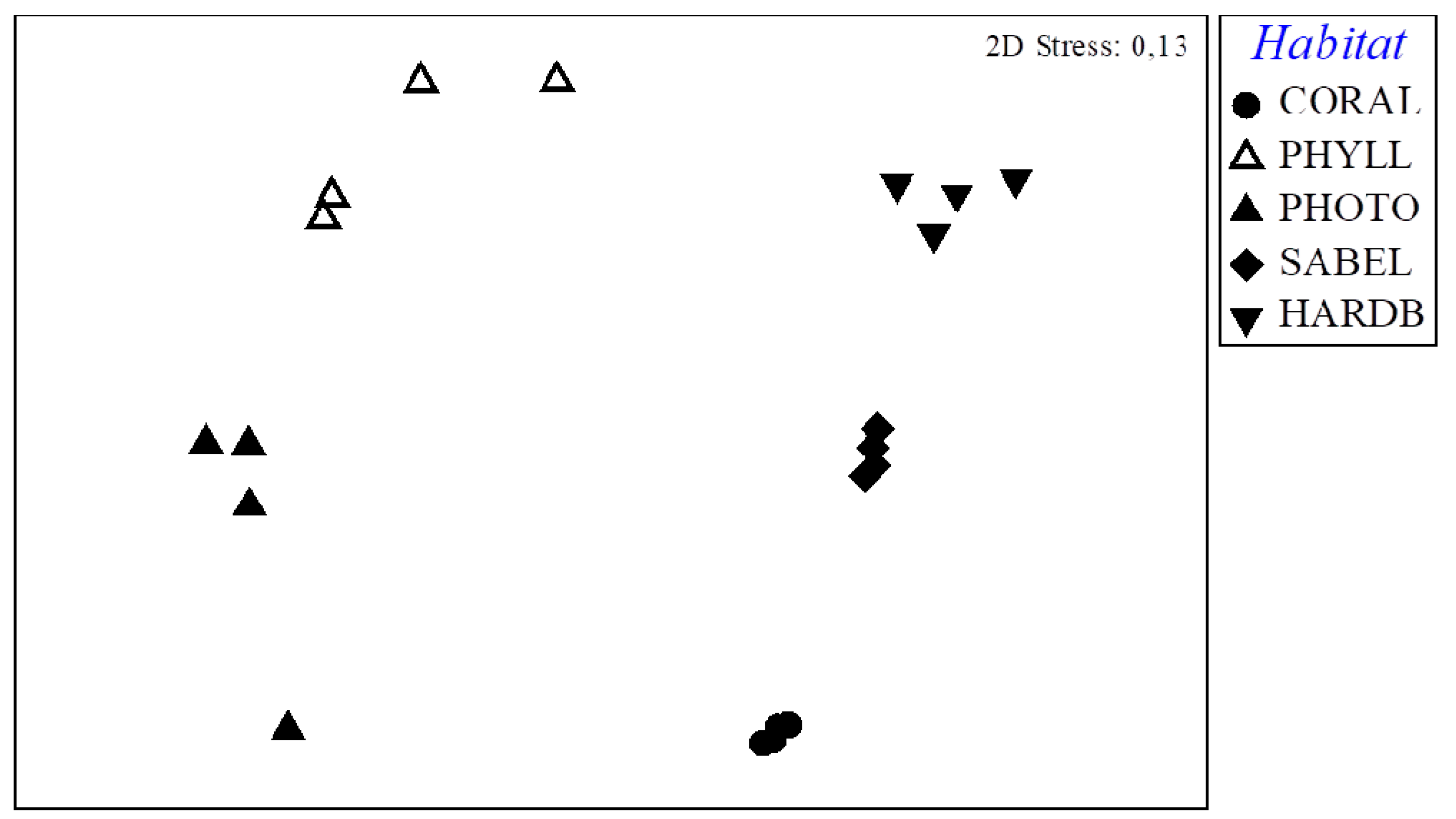
3.4. Inter-Habitat Comparison of the Molluscan Assemblages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Bianchi, C.N. La biocostruzione negli ecosistemi marini e la biologia marina italiana. Biol. Mar. Mediterr. 2001, 8, 112–130. [Google Scholar]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean bioconstructions along the Italian coast. In Advances in Marine Biology, 1st ed.; Sheppard, C., Fuiman, L.A., Young, C.M., Eds.; Zoe Kruze: London, UK, 2018; Volume 79, pp. 61–136. [Google Scholar] [CrossRef]
- Bonifazi, A.; Ventura, D.; Gravina, M.F.; Lasinio, G.J.; Belluscio, A.; Ardizzone, G.D. Unusual algal turfs associated with the rhodophyta Phyllophora crispa: Benthic assemblages along a depth gradient in the Central Mediterranean Sea. Estuar. Coast. Shelf. Sci. 2017, 185, 77–93. [Google Scholar] [CrossRef]
- Dubois, S.; Retière, C.; Olivier, F. Biodiversity associated with Sabellaria alveolata (Polychaeta: Sabellariidae) reefs: Effects of human disturbances. J. Mar. Biol. Assoc. UK 2002, 82, 817–826. [Google Scholar] [CrossRef]
- La Porta, B.; Nicoletti, L. Sabellaria alveolata (Linnaeus) reefs in the central Tyrrhenian Sea (Italy) and associated polychaete fauna. Zoosymposia 2009, 2, 527–536. [Google Scholar]
- Bonifazi, A.; Ventura, D.; Mancini, E. Sabellaria reefs as reservoirs of rare species: The case of Eulalia ornata Saint-Joseph, 1888 (Annelida: Phyllodocidae). Mar. Freshw. Res. 2018, 69, 1635–1640. [Google Scholar] [CrossRef]
- Gruet, Y. Recherches sur l’ écologie des “recifs” d’Hermelles édifiés par l’annélide polychéte Sabellaria alveolata (Linné). Ph.D. Thesis, Universite de Nantes, Nantes, France, 1982. [Google Scholar]
- Dias, A.S.; Paula, J. Associated fauna of Sabellaria alveolata colonies on the central coast of Portugal. J. Mar. Biol. Assoc. UK 2001, 81, 169–170. [Google Scholar] [CrossRef]
- Salomidi, M.; Katsanevakis, S.; Borja, Á.; Braeckman, U.; Damalas, D.; Galparsoro, I.; Mifsud, R.; Mirto, S.; Pascual, M.; Pipitone, C.; et al. Assessment of goods and services, vulnerability, and conservation status of European seabed biotopes: A stepping stone towards ecosystem-based marine spatial management. Mediterr. Mar. Sci. 2012, 13, 49–88. [Google Scholar] [CrossRef]
- Dubois, S.; Commito, J.A.; Olivier, F.; Retière, C. Effects of epibionts on Sabellaria alveolata (L.) biogenic reefs and their associated fauna in the Bay of Mont Saint-Michel. Estuar. Coast. Shelf Sci. 2006, 68, 635–646. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C.; Navone, A. I popolamenti delle scogliere rocciose sommerse dell’Area Marina Protetta di Tavolara Punta Coda Cavallo (Sardegna nord-orientale). Sci. Rep. Port-Cros Natl. Park. 2010, 24, 39–85. [Google Scholar]
- Kostylev, E.F.; Tkachenko, F.P.; Tretiak, I.P. Establishment of “Zernov’s Phyllophora field” marine reserve: Protection and restoration of a unique ecosystem. Ocean Coast. Manag. 2010, 53, 203–208. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F. How algae influence sessile marine organisms: The tube worms case of study. Estuar. Coast. Shelf Sci. 2016, 178, 12–20. [Google Scholar] [CrossRef]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. In Oceanography and Marine Biology: An Annual Review, 1st ed.; Gibson, R.N., Atkinson, J.A., Gordon, J.D.M., Eds.; Taylor & Francis: Broken Sound Parkway NW, FL, USA, 2006; Volume 44, pp. 123–195. [Google Scholar]
- Tribot, A.S.; Mouquet, N.; Villéger, S.; Raymond, M.; Hoff, F.; Boissery, P.; Holon, F.; Deter, J. Taxonomic and functional diversity increase the aesthetic value of coralligenous reefs. Sci. Rep. 2016, 6, 34229. [Google Scholar] [CrossRef]
- Parravicini, V.; Micheli, F.; Montefalcone, M.; Villa, E.; Morri, C.; Bianchi, C.N. Rapid assessment of epibenthic communities: A comparison between two visual sampling techniques. J. Exp. Mar. Bio. Ecol. 2010, 395, 21–29. [Google Scholar] [CrossRef]
- Kipson, S.; Fourt, M.; Teixidó, N.; Cebrian, E.; Casas, E.; Ballesteros, E.; Zabala, M.; Garrabou, J. Rapid biodiversity assessment and monitoring method for highly diverse benthic communities: A case study of Mediterranean coralligenous outcrops. PLoS ONE 2011, 6, 11–13. [Google Scholar] [CrossRef]
- Gatti, G.; Montefalcone, M.; Rovere, A.; Parravicini, V.; Morri, C.; Albertelli, G.; Bianchi, C.N. Seafloor integrity down the harbor waterfront: The coralligenous shoals off Vado Ligure (NW Mediterranean). Adv. Oceanogr. Limnol. 2012, 3, 51–67. [Google Scholar] [CrossRef]
- Teixidó, N.; Casas, E.; Cebrián, E.; Linares, C.; Garrabou, J. Impacts on Coralligenous Outcrop Biodiversity of a Dramatic Coastal Storm. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Doxa, A.; Holon, F.; Deter, J.; Villéger, S.; Boissery, P.; Mouquet, N. Mapping biodiversity in three-dimensions challenges marine conservation strategies: The example of coralligenous assemblages in north-western Mediterranean Sea. Ecol. Indic. 2016, 61, 1042–1054. [Google Scholar] [CrossRef]
- Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Available online: https://eur-lex.europa.eu/eli/dir/1992/43/oj (accessed on 14 March 2019).
- Pérès, J.M.; Picard, J. Nouveau Manuel de Bionome benthique de la Mer Mediterranee. Recl. des Trav. la Stn. Mar. l’Endoume. 1964, 47, 5–137. [Google Scholar]
- Lassau, S.A.; Hochuli, D.F. Effects of habitat complexity on ant assemblages. Ecography 2004, 27, 157–164. [Google Scholar] [CrossRef]
- Menge, B.A.; Lubchenco, J.; Ashkenas, L.R. Diversity, heterogeneity and consumer pressure in a tropical rocky intertidal community. Oecologia 1985, 65, 394–405. [Google Scholar] [CrossRef]
- Hull, S.L. Seasonal changes in diversity and abundance of ostracods on four species of intertidal algae with differing structural complexity. Mar. Ecol. Prog. Ser. 1997, 161, 71–82. [Google Scholar] [CrossRef]
- Bertness, M.D.; Leonard, G.H.; Levine, J.M.; Schmidt, P.R.; Ingraham, A.O. Testing the relative contribution of positive and negative interactions in rocky intertidal communities. Ecology 1999, 80, 2711–2726. [Google Scholar] [CrossRef]
- Abbiati, M.; Bianchi, C.N.; Castelli, A. Polychaete vertical zonation along a littoral cliff in the west Mediterranean. Mar. Ecol. 1987, 8, 33–48. [Google Scholar] [CrossRef]
- Giangrande, A. Polychaete zonation and its relation to algal distribution down a vertical cliff in the western Mediterranean (Italy): A structural analysis. J. Exp. Mar. Bio. Ecol. 1988, 120, 263–276. [Google Scholar] [CrossRef]
- Chemello, R.; Milazzo, M. Effect of algal architecture on associated fauna: Some evidence from phytal molluscs. Mar. Biol. 2002, 140, 981–990. [Google Scholar] [CrossRef]
- Giangrande, A.; Delos, A.L.; Musco, L.; Licciano, M.; Pierri, C. Polychaete assemblages along a rocky shore on the South Adriatic coast (Mediterranean Sea). Cah. Biol. Mar. 2004, 45, 85–95. [Google Scholar]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Milazzo, M.; Chemello, R.; Badalamenti, F.; Riggio, S. Molluscan assemblages associated with photophilic algae in the Marine Reserve of Ustica Island (Lower Tyrrhenian Sea, Italy). Ital. J. Zool. 2000, 67, 287–295. [Google Scholar] [CrossRef]
- Russo, G.F.; Patti, F.P. Early life history of two closely related gastropods, Rissoa auriscalpium and Rissoa italiensis (Caenogastropoda: Rissoidae). Mar. Boil. 2005, 147, 429–437. [Google Scholar] [CrossRef]
- Donnarumma, L.; Sandulli, R.; Apolloni, L.; Sánchez-Lizaso, J.L.; Russo, G.F. Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions. Diversity 2018, 10, 96. [Google Scholar] [CrossRef]
- Lolas, A.; Antoniadou, C.; Vafidis, D. Spatial variation of molluscan fauna associated with Cystoseira assemblages from a semi-enclosed gulf in the Aegean Sea. Reg. Stud. Mar. Sci. 2018, 19, 17–24. [Google Scholar] [CrossRef]
- Antoniadou, C.; Koutsoubas, D.; Chintiroglou, C.C. Mollusca fauna from infralittoral hard substrate assemblages in the North Aegean Sea. Belgian J. Zool. 2005, 135, 119–126. [Google Scholar]
- Urra, J.; Rueda, J.L.; Gofas, S.; Marina, P.; Salas, C. A species-rich molluscan assemblage in a coralligenous bottom of the Alboran Sea (south-western Mediterranean): Intra-annual changes and ecological considerations. J. Mar. Biol. Assoc. UK 2012, 92, 665–677. [Google Scholar] [CrossRef]
- Poursanidis, D.; Koutsoubas, D. A computerized database (CorMol) on the molluscan fauna from the Mediterranean reef ecosystems: Part I, the coralligenous formations. Quat. Int. 2015, 390, 29–43. [Google Scholar] [CrossRef]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Russo, G.F. Assessing molluscs functional diversity within different coastal habitats of Mediterranean marine protected areas. Ecol. Quest. 2018, 29, 35–51. [Google Scholar]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Appolloni, L.; Bevilacqua, S.; Sbrescia, L.; Sandulli, R.; Terlizzi, A.; Russo, G.F. Does full protection count for the maintenance of β-diversity patterns in marine communities? Evidence from Mediterranean fish assemblages. Aquat. Conserv. 2017, 27, 828–838. [Google Scholar] [CrossRef]
- Buonocore, E.; Picone, F.; Donnarumma, L.; Russo, G.F.; Franzese, P.P. Modeling matter and energy flows in marine ecosystems using emergy and eco-exergy methods to account for natural capital value. Ecol. Modell. 2019, 392, 137–146. [Google Scholar] [CrossRef]
- Bonifazi, A.; Lezzi, M.; Ventura, D.; Lisco, S.; Cardone, F.; Gravina, M.F. Macrofaunal biodiversity associated with different developmental phases of a threatened Mediterranean Sabellaria alveolata (Linnaeus, 1767) reef. Mar. Environ. Res. 2019. [Google Scholar] [CrossRef]
- Piazzi, L.; Gennaro, P.; Montefalcone, M.; Bianchi, C.N.; Cecchi, E.; Morri, C.; Serena, F. STAR: An integrated and standardized procedure to evaluate the ecological status of coralligenous reefs. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 1–13. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Li, S.Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2010. [Google Scholar]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; p. 283. [Google Scholar]
- Purchon, R.D. The Biology of the Mollusca, 2nd ed.; Pergamon Press Ltd.: Oxford, UK, 1977; p. 587. [Google Scholar]
- Gravina, M.F.; Cardone, F.; Bonifazi, A.; Bertrandino, M.S.; Chimienti, G.; Longo, C.; Marzano, C.N.; Moretti, M.; Lisco, S.; Moretti, V.; et al. Sabellaria spinulosa (Polychaeta, Annelida) reefs in the Mediterranean Sea: Habitat mapping, dynamics and associated fauna for conservation management. Estuar. Coast. Shelf Sci. 2018, 200, 248–257. [Google Scholar] [CrossRef]
- Lezzi, M.; Cardone, F.; Mikac, B.; Giangrande, A. Variation and ontogenetic changes of opercular paleae in a population of Sabellaria spinulosa (Polychaeta: Sabellaridae) from the South Adriatic Sea, with remarks on larval development. Sci. Mar. 2015, 79, 1–14. [Google Scholar]
- Lisco, S.; Moretti, M.; Moretti, V.; Cardone, F.; Corriero, G.; Longo, C. Sedimentological features of Sabellaria spinulosa bioconstructions. Mar. Pet. Geol. 2017, 87, 203–212. [Google Scholar] [CrossRef]
- La Porta, B.; Targusi, M.; Lattanzi, L.; La Valle, P.; Nicoletti, L. Analisi Della Fauna Associata Alle Biocostruzioni a Sabellaria alveolata (L.) in Relazione Al Loro Stato Di Conservazione. Biol. Mar. Mediterr. 2009, 16, 36–38. [Google Scholar]
- Ardizzone, G.D.; Gravina, M.F.; Belluscio, A. Temporal development of epibenthic communities on artificial reefs in the central Mediterranean Sea. Bullletin. Mar. Sci. 1989, 44, 592–608. [Google Scholar]
- Thiel, M.; Ullrich, N. Hard rock versus soft bottom: The fauna associated with intertidal mussel beds on hard bottoms along the coast of Chile, and considerations on the functional role of mussel beds. Helgol. Mar. Res. 2002, 56, 21–30. [Google Scholar] [CrossRef]
- Maggi, E.; Bertocci, I.; Vaselli, S.; Benedetti-Cecchi, L. Effects of changes in number, identity and abundance of habitat-forming species on assemblages of rocky seashores. Mar. Ecol. Prog. Ser. 2009, 381, 39–49. [Google Scholar] [CrossRef]
- Arribas, L.P.; Donnarumma, L.; Palomo, M.G.; Scrosati, R.A. Intertidal mussels as ecosystem engineers: Their associated invertebrate biodiversity under contrasting wave exposures. Mar. Biodivers. 2014, 44, 203–211. [Google Scholar] [CrossRef]
- Tongiorgi, P.; Nardi, P.; Galleni, L.; Nigro, M.; Salghetti, U. Feeding habits of Ocinebrina edwardsi (Mollusca: Prosobranchia) a common mussel drill of the Italian coasts. Mar. Ecol. 1981, 2, 169–180. [Google Scholar] [CrossRef]
- Huston, M. A general hypothesis of species diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Gili, J.M.; Coma, R. Benthic suspension feeders: Their paramount role in littoral marine food webs. Trends Ecol. Evol. 1998, 13, 316–321. [Google Scholar] [CrossRef]
- Hrs-Brenko, M.; Legac, M. Inter-and intra-species relationships of sessile bivalves on the eastern coast of the Adriatic. Sea Nat. Croat. 2006, 15, 203–230. [Google Scholar]
- Schiaparelli, S.; Franci, G.; Albertelli, G.; Cattaneo-Vietti, R. A non destructive method to evaluate population stucture and bioerosion activity of the boring bivalve Gastrochaena dubia. J. Coast. Res. 2005, 21, 383–386. [Google Scholar] [CrossRef]
- Casoli, E.; Ricci, S.; Antonelli, F.; Perasso, S.C.; Belluscio, A.; Ardizzone, G.D. Impact and colonization dynamics of the bivalve Rocellaria dubia on limestone experimental panels in the submerged Roman city of Baiae (Naples, Italy). Int. Biodeter. Biodegr. 2016, 108, 9–15. [Google Scholar] [CrossRef]
- Fava, F.; Ponti, M.; Abbiati, M. Role of recruitment processes in structuring coralligenous benthic assemblages in the northern Adriatic continental shelf. PLoS ONE 2016, 11, e0163494. [Google Scholar] [CrossRef]
- Safriel, U.N.; Ben Eliahu, M.N. The influence of habitat structure and environmental stability on the species diversity of polychaetes in vermetid reefs. In Habitat Structure. The Physical Arrangement of Objects in Space; Bell, S.S., McCoy, E.D., Mushinsky, H.R., Eds.; Chapman and Hall, Ltd.: London, UK, 1991; pp. 349–369. [Google Scholar] [CrossRef]
- Cocito, S. Bioconstruction and biodiversity: Their mutual influence. Sci. Mar. 2004, 68, 137–144. [Google Scholar] [CrossRef]
- Kelly, N.; Metaxas, A. Diversity of invertebrate colonists on simple and complex substrates at hydrothermal vents on the Juan de Fuca Ridge. Aquat. Biol. 2008, 3, 271–281. [Google Scholar] [CrossRef]
- Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Guidetti, P.; Boero, F. Molluscs on subtidal cliffs: Patterns of spatial distribution. J. Mar. Biol. Assoc. UK 2003, 83, 165–172. [Google Scholar] [CrossRef]
- Bode, A. Reproduction of the intertidal chiton Acantochitona crinita within a community of Corallina elongata (Rhodophyta). J. Moll. Stud. 1989, 55, 37–44. [Google Scholar] [CrossRef]
- Öztürk, B.; Ergen, Z.; Önen, M. Polyplacophora (Mollusca) from the Aegean coast of Turkey. Zool. Middle East. 2000, 20, 69–76. [Google Scholar] [CrossRef]
- Hong, J.S. Contribution à l’étude des peuplements d’un fond de concrétionnement coralligène dans la région marseillaise en Mediterranée Nord-occidentale. Bull. KORDI 1982, 4, 27–51. [Google Scholar]
- Boudouresque, C.F. Marine biodiversity in the Mediterranean: Status of species, populations and communities. Trav. Sci. Parc. Natl. Port-Cros. 2004, 20, 97–146. [Google Scholar]
- Zaitsev, Y. An Introduction to the Black Sea Ecology; Odessa: Smil Editing & Publishing Agency Ltd.: Istanbul, Turkey, 2008; p. 228. [Google Scholar]
- Relini, G.; Giaccone, G. Gli habitat prioritari del protocollo SPA/BIO (Convenzione di Barcellona) presenti in Italia. Schede descrittive per l’identificazione. Biol. Mar. Mediterr. 2009, 16, 1–372. [Google Scholar]
- Chintiroglou, C.; Koukouras, A. A populatrion of the Sea Anemone Anemonia viridis (Forskal, 1775) and its associated flora and fauna, in north Aegean Sea. Int. Rev. Ges. Hydrobiol. 1992, 77, 483–495. [Google Scholar] [CrossRef]
- Cosentino, A.; Giacobbe, S. Mollusc assemblages of hard bottom subtidal fringe: A comparison between two coastal typologies. Biodivers. J. 2015, 6, 353–364. [Google Scholar] [CrossRef]
- Chemello, R.; Russo, G.F. The molluscan taxocoene of photophilic algae from the Island of Lampedusa (Strait of Sicily, southern Mediterranean). Boll. Malacol. 1997, 33, 95–104. [Google Scholar]
- Torres, A.C.; Veiga, P.; Rubal, M.; Sousa-Pinto, I. The role of annual macroalgal morphology in driving its epifaunal assemblages. J. Exp. Mar. Bio. Ecol. 2015, 464, 96–106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RS | FG | Species | Lido di Ostia | Punta del Lazzaretto | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SABEL | HARDB | CORAL | PHYLL | PHOTO | ||||||||
| N | %DI | N | %DI | N | %DI | N | %DI | N | %DI | |||
| Polyplacophora | ||||||||||||
| Sd | MG | Acanthochitona crinita (Pennant, 1777) | - | - | - | - | - | - | - | - | 1 | 3.85 |
| Sd | MG | Acanthochitona fascicularis (Linnaeus, 1767) | - | - | - | - | 7 | 0.95 | - | - | - | - |
| Sd | MG | Callochitonseptemvalvis (Montagu, 1803) | - | - | - | - | 3 | 0.41 | - | - | - | - |
| Sd | MG | Chiton corallinus (Spengler, 1797) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| Sd | MG | Lepidopleurus cf cimicoides (Monterosato, 1879) | - | - | - | - | 12 | 1.64 | - | - | - | - |
| Gastropoda | ||||||||||||
| V | DF | Alvania spinosa (Monterosato, 1890) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | DF | Alvania tenera (Philippi, 1844) | - | - | - | - | 2 | 0.27 | - | - | - | - |
| V | P | Aplus scaber (Locard, 1892) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | MG | Bittium latreillii (Payraudeau, 1826) | 2 | 0.17 | - | - | 25 | 3.41 | 18 | 25.00 | 8 | 30.77 |
| V | MG | Bittium reticulatum (Costa, 1778) | 59 | 4.96 | 33 | 2.96 | 4 | 0.55 | 1 | 1.39 | 6 | 23.08 |
| V | MG | Bolma rugosa (Linnaeus, 1767) | - | - | - | - | - | - | 3 | 4.17 | - | - |
| V | DF | Caecum auriculatum (de Folin, 1868) | - | - | - | - | 3 | 0.41 | - | - | - | - |
| V | P | Calliostoma conulus (Linnaeus, 1758) | - | - | - | - | - | - | - | - | 2 | 7.69 |
| V | P | Calliostoma laugieri (Payraudeau, 1826) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| V | P | Cerithiopsis barleei (Jeffreys, 1867) | - | - | - | - | 4 | 0.55 | - | - | - | - |
| V | P | Cerithiopsis minima (Brusina, 1865) | 1 | 0.08 | - | - | - | - | - | - | - | - |
| V | P | Chauvetia mamillata (Risso, 1826) | - | - | - | - | - | - | 4 | 5.56 | - | - |
| V | MG | Clanculus cruciatus (Linnaeus, 1758) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | DF | Crisilla semistriata (Montagu, 1808) | 4 | 0.34 | - | - | 21 | 2.86 | - | - | - | - |
| V | P | Diodora graeca (Linnaeus, 1758) | 1 | 0.08 | 5 | 0.45 | - | - | - | - | - | - |
| V | P | Doris verrucosa (Linnaeus, 1758) | 1 | 0.08 | - | - | - | - | - | - | - | - |
| V | P | Enginella leucozona (Philippi, 1844) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| V | P | Felimare tricolor (Cantraine, 1835) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| V | P | Felimida purpurea (Risso, 1831) | - | - | - | - | - | - | 2 | 2.78 | - | - |
| V | MG | Fissurella nubecula (Linnaeus, 1758) | - | - | 9 | 0.81 | - | - | - | - | - | - |
| V | E | Folinella excavata (Philippi, 1836) | - | - | - | - | 5 | 0.68 | - | - | - | - |
| V | P | Fusinus dimassai (Buzzurro & Russo, 2007) | - | - | - | - | 1 | 0.14 | 3 | 4.17 | - | - |
| V | MG | Jujubinus exasperatus (Pennant, 1777) | - | - | - | - | 1 | 0.14 | 2 | 2.78 | - | - |
| V | MG | Manzonia crassa (Kanmacher, 1798) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | P | Marshallora adversa (Montagu, 1803) | - | - | 1 | 0.09 | 1 | 0.14 | 1 | 1.39 | - | - |
| V | P | Metaxia metaxa (Delle Chiaje, 1828) | - | - | - | - | 3 | 0.41 | - | - | - | - |
| V | P | Mitrella scripta (Linnaeus, 1758) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| V | P | Murexsul aradasii (Monterosato in Poirier, 1883) | - | - | - | - | - | - | 2 | 2.78 | - | - |
| V | P | Muricopsis cristata (Brocchi, 1814) | - | - | - | - | 2 | 0.27 | - | - | - | - |
| V | P | Ocenebra edwardsii (Payraudeau, 1826) | 16 | 1.35 | 40 | 3.59 | - | - | - | - | - | - |
| V | P | Ocinebrina aciculata (Lamarck, 1822) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| V | E | Odostomella bicincta (Tiberi, 1868) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | E | Odostomella doliolum (Philippi, 1844) | 2 | 0.17 | - | - | 1 | 0.14 | - | - | - | - |
| V | E | Odostomia cf plicata (Montagu, 1803) | - | - | - | - | 4 | 0.55 | - | - | - | - |
| V | E | Odostomia sp. | 1 | 0.08 | - | - | - | - | - | - | - | - |
| V | E | Parthenina emaciata (Brusina, 1866) | - | - | - | - | 2 | 0.27 | - | - | - | - |
| V | P | Pusia savignyi (Payraudeau, 1826) | - | - | - | - | - | - | 5 | 6.94 | - | - |
| V | P | Pusia tricolor (Gmelin, 1791) | - | - | - | - | 1 | 0.14 | 1 | 1.39 | - | - |
| V | MG | Pusillina philippi (Aradas & Maggiore, 1844) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | MG | Pusillina sp. | 1 | 0.08 | - | - | - | - | - | - | - | - |
| V | P | Raphitoma linearis (Montagu, 1803) | - | - | - | - | 1 | 0.14 | 1 | 1.39 | - | - |
| V | P | Retusa crossei (Bucquoy, Dautzenberg & Dollfus, 1886) | - | - | - | - | - | - | - | - | 1 | 3.85 |
| V | MG | Rissoa sp. | 1 | 0.08 | - | - | - | - | - | - | - | - |
| V | MG | Rissoa variabilis (Megerle von Mühlfeld, 1824) | - | - | - | - | - | - | - | - | 1 | 3.85 |
| V | MG | Rissoina bruguieri (Payraudeau, 1826) | - | - | - | - | 3 | 0.41 | - | - | - | - |
| V | DF | Scissurella costata (d’Orbigny, 1824) | - | - | - | - | 13 | 1.77 | - | - | - | - |
| V | E | Spiralinella incerta (Milaschewich, 1916) | 5 | 0.42 | - | - | 2 | 0.27 | - | - | - | - |
| V | MG | Tecturea virginea (O.F. Müller, 1776) | - | - | - | - | 8 | 1.09 | - | - | 1 | 3.85 |
| Ss | SF | Thylaeodus semisurrectus (Bivona-Bernardi, 1832) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| V | SC | Tritia incrassata (Strøm, 1768) | 1 | 0.08 | - | - | - | - | - | - | 1 | 3.85 |
| V | SC | Tritia pygmaea (Lamarck, 1822) | 1 | 0.08 | - | - | 1 | 0.14 | - | - | - | - |
| Ss | SF | Vermetus granulatus (Gravenhorst, 1831) | - | - | - | - | 8 | 1.09 | - | - | - | - |
| V | E | Vitreolina philippi (de Rayneval & Ponzi, 1854) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| Bivalvia | ||||||||||||
| Ss | SF | Anomia ephippium (Linnaeus, 1758) | 6 | 0.50 | 1 | 0.09 | - | - | 1 | 1.39 | - | - |
| Ss | SF | Arca noae (Linnaeus, 1758) | 3 | 0.25 | - | - | - | - | 2 | 2.78 | - | - |
| Ss | SF | Arca tetragona (Poli, 1795) | - | - | - | - | 11 | 1.50 | 1 | 1.39 | - | - |
| Ss | SF | Asperarca nodulosa (O. F. Müller, 1776) | - | - | - | - | 5 | 0.68 | - | - | - | - |
| Ss | SF | Barbatia barbata (Linnaes, 1758) | - | - | - | - | 9 | 1.23 | - | - | - | - |
| Ss | SF | Chama gryphoides (Linnaeus, 1758) | 10 | 0.84 | - | - | 7 | 0.95 | 1 | 1.39 | - | - |
| Ss | SF | Crassostrea gigas (Thunberg, 1793) | 2 | 0.17 | - | - | - | - | - | - | - | - |
| Ss | SF | Crenella arenaria (Monterosato, 1875 ex H. Martin, ms.) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| Sd | SF | Flexopecten hyalinus (Poli, 1795) | - | - | - | - | - | - | 2 | 2.78 | - | - |
| Sd | SF | Gouldia minima (Montagu, 1803) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| Ss | SF | Gregariella petagnae (Scacchi, 1832) | 2 | 0.17 | 5 | 0.45 | 3 | 0.41 | 1 | 1.39 | - | - |
| Ss | SF | Gregariella semigranata (Reeve, 1858) | 11 | 0.93 | 3 | 0.27 | 36 | 4.91 | 1 | 1.39 | - | - |
| Ss | SF | Hiatella arctica (Linnaeus, 1767) | 99 | 8.33 | 4 | 0.36 | 73 | 9.96 | 2 | 2.78 | 1 | 3.85 |
| Ss | SF | Irus irus (Linnaeus, 1758) | 8 | 0.67 | 1 | 0.09 | 3 | 0.41 | - | - | - | - |
| Ss | SF | Lima lima (Linnaeus, 1758) | - | - | - | - | 1 | 0.14 | 1 | 1.39 | - | - |
| Ss | SF | Limaria hians (Gmelin, 1791) | 5 | 0.42 | - | - | 1 | 0.14 | - | - | - | - |
| Ss | SF | Lithophaga lithophaga (Linnaeus, 1758) | 2 | 0.17 | 1 | 0.09 | 43 | 5.87 | - | - | - | - |
| Sd | SF | Mimachlamys varia (Linnaeus, 1758) | 1 | 0.08 | - | - | - | - | - | - | - | - |
| Ss | SF | Modiolula phaseolina (Philippi, 1844) | 8 | 0.67 | 5 | 0.45 | 1 | 0.14 | - | - | - | - |
| Ss | SF | Modiolus barbatus (Linnaeus, 1758) | 13 | 1.09 | - | - | 5 | 0.68 | 1 | 1.39 | - | - |
| Ss | SF | Musculus costulatus (Risso, 1826) | 5 | 0.42 | 7 | 0.63 | 27 | 3.68 | 4 | 5.56 | 4 | 15.38 |
| Ss | SF | Musculus subpictus (Cantraine, 1835) | 2 | 0.17 | - | - | - | - | - | - | - | - |
| Ss | SF | Mytilaster minimus (Poli, 1795) | - | - | 8 | 0.72 | - | - | - | - | - | - |
| Ss | SF | Mytilus galloprovincialis (Lamarck, 1819) | 133 | 11.19 | 983 | 88.24 | - | - | - | - | - | - |
| Sd | SF | Neolepton sulcatulum (Jeffreys, 1859) | - | - | - | - | 7 | 0.95 | - | - | - | - |
| Ss | SF | Ostrea edulis (Linnaeus, 1758) | 3 | 0.25 | 2 | 0.18 | - | - | - | - | - | - |
| Sd | SF | Palliolum incomparabile (Risso, 1826) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| Sd | SF | Papillicardium papillosum (Poli, 1791) | - | - | - | - | 2 | 0.27 | - | - | - | - |
| Sd | SF | Parvicardium exiguum (Gmelin, 1791) | 1 | 0.08 | - | - | - | - | - | - | - | - |
| Sd | SF | Parvicardium scriptum (Bucquoy, Dautzenberg & Dollfus, 1892) | 15 | 1.26 | - | - | - | - | - | - | - | - |
| Ss | SF | Petricola lithophaga (Retzius, 1788) | 7 | 0.59 | 1 | 0.09 | 1 | 0.14 | - | - | - | - |
| Ss | SF | Pseudochama gryphina (Lamarck, 1819) | - | - | - | - | 4 | 0.55 | - | - | - | - |
| Ss | SF | Rocellaria dubia (Pennant, 1777) | 3 | 0.25 | - | - | 10 | 1.36 | - | - | - | - |
| Ss | SF | Spondylus gaederopus (Linnaeus, 1758) | - | - | - | - | 1 | 0.14 | - | - | - | - |
| Ss | SF | Striarca lactea (Linnaeus, 1758) | 753 | 63.33 | 4 | 0.36 | 326 | 44.47 | 5 | 6.94 | - | - |
| Sd | SF | Talochlamys multistriata (Poli, 1795) | - | - | - | - | - | - | 1 | 1.39 | - | - |
| Sd | SF | Venerupis corrugata (Gmelin, 1791) | 1 | 0.08 | 1 | 0.09 | - | - | - | - | - | - |
| Sd | SF | Venus casina (Linnaeus, 1758) | - | - | - | - | 8 | 1.09 | - | - | - | - |
| Site | Habitat | SR | N | J | H′ |
|---|---|---|---|---|---|
| Lido di Ostia | SABEL | 22.25 ± 3.77 | 297.25 ± 192.21 | 0.52 ± 0.14 | 2.30 ± 0.55 |
| HARDB | 10.75 ± 3.59 | 278.50 ± 164.29 | 0.30 ± 0.21 | 1.04 ± 0.72 | |
| Punta del Lazzaretto | CORAL | 34.50 ± 2.88 | 366 ± 97.43 | 0.68 ± 0.08 | 3.44 ± 0.32 |
| PHYLL | 12.5 ± 1.291 | 18 ± 2 | 0.92 ± 0.06 | 3.35 ± 0.36 | |
| PHOTO | 4.25 ± 0.96 | 6.5 ± 1.29 | 0.96 ± 0.06 | 1.97 ± 0.36 | |
| PERMANOVA test | F | 72.942 | 7.7411 | 19.719 | 17.251 |
| p(perm) | 0.0002 | 0.0028 | 0.0002 | 0.0002 | |
| Unique perms | 1927 | 4975 | 4982 | 4988 |
| PERMANOVA Pair-Wise Test | t | p | Pair-Wise Test | t | p |
|---|---|---|---|---|---|
| SABEL vs. PHYLL | 2.9552 | 0.031 | CORAL vs. PHYLL | 3.0418 | 0.0308 |
| SABEL vs. PHOTO | 3.2232 | 0.0292 | CORAL vs. PHOTO | 3.3665 | 0.0318 |
| SABEL vs. HARDB | 3.2896 | 0.0272 | CORAL vs. HARDB | 5.1407 | 0.0284 |
| SABEL vs. CORAL | 3.7467 | 0.0272 | PHYLL vs. PHOTO | 1.9856 | 0.0308 |
| HARDB vs. PHYLL | 3.0418 | 0.0268 | HARDB vs. PHOTO | 3.3091 | 0.0308 |
| Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% |
|---|---|---|---|---|---|
| Striarca lactea | 2.26 | 2.90 | 0.75 | 13.12 | 13.12 |
| Bittium latreillii | 1.07 | 2.86 | 0.52 | 12.94 | 26.08 |
| Bittium reticulatum | 1.12 | 2.41 | 0.56 | 10.91 | 36.98 |
| Mytilus galloprovincialis | 1.72 | 2.35 | 0.37 | 10.65 | 47.63 |
| Average Similarity | Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% |
|---|---|---|---|---|---|---|
| CORAL—68.95% | Striarca lactea | 5.04 | 7.96 | 31.94 | 11.55 | 11.55 |
| Hiatella arctica | 3.43 | 4.89 | 7.54 | 7.10 | 18.64 | |
| Lithophaga lithophaga | 3.07 | 4.77 | 11.73 | 6.92 | 25.56 | |
| Musculus costulatus | 2.67 | 4.33 | 19.60 | 6.27 | 31.83 | |
| Gregariella semigranata | 2.83 | 4.13 | 4 | 5.99 | 37.82 | |
| Crisilla semistriata | 2.39 | 3.56 | 8.12 | 5.17 | 43.99 | |
| Bittium latreillii | 2.47 | 3.49 | 3.53 | 5.06 | 48.05 | |
| SABEL—67.10% | Striarca lactea | 4.93 | 13.49 | 11.11 | 20.11 | 20.11 |
| Mytilus galloprovincialis | 3.38 | 9.36 | 15.85 | 13.95 | 34.06 | |
| Hiatella arctica | 3.11 | 8.47 | 4.93 | 12.63 | 46.69 | |
| HARDB—64.65% | Mytilus galloprovincialis | 5.21 | 28.13 | 4.59 | 43.51 | 43.51 |
| PHOTO—42.21% | Bittium reticulatum | 0.9 | 20.22 | 4.19 | 47.91 | 47.91 |
| PHYLL—37.17% | Bittium latreillii | 1.57 | 11.4 | 2.23 | 30.67 | 30.67 |
| Pusia savignyi | 0.79 | 6.72 | 22.28 | 18.09 | 48.76 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F.; Russo, G.F.; Sandulli, R.; Donnarumma, L. Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity 2019, 11, 74. https://doi.org/10.3390/d11050074
Casoli E, Bonifazi A, Ardizzone G, Gravina MF, Russo GF, Sandulli R, Donnarumma L. Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity. 2019; 11(5):74. https://doi.org/10.3390/d11050074
Chicago/Turabian StyleCasoli, Edoardo, Andrea Bonifazi, Giandomenico Ardizzone, Maria Flavia Gravina, Giovanni Fulvio Russo, Roberto Sandulli, and Luigia Donnarumma. 2019. "Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea" Diversity 11, no. 5: 74. https://doi.org/10.3390/d11050074
APA StyleCasoli, E., Bonifazi, A., Ardizzone, G., Gravina, M. F., Russo, G. F., Sandulli, R., & Donnarumma, L. (2019). Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity, 11(5), 74. https://doi.org/10.3390/d11050074