Spatial and Temporal Variation in Fecundity of Acropora spp. in the Northern Great Barrier Reef

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
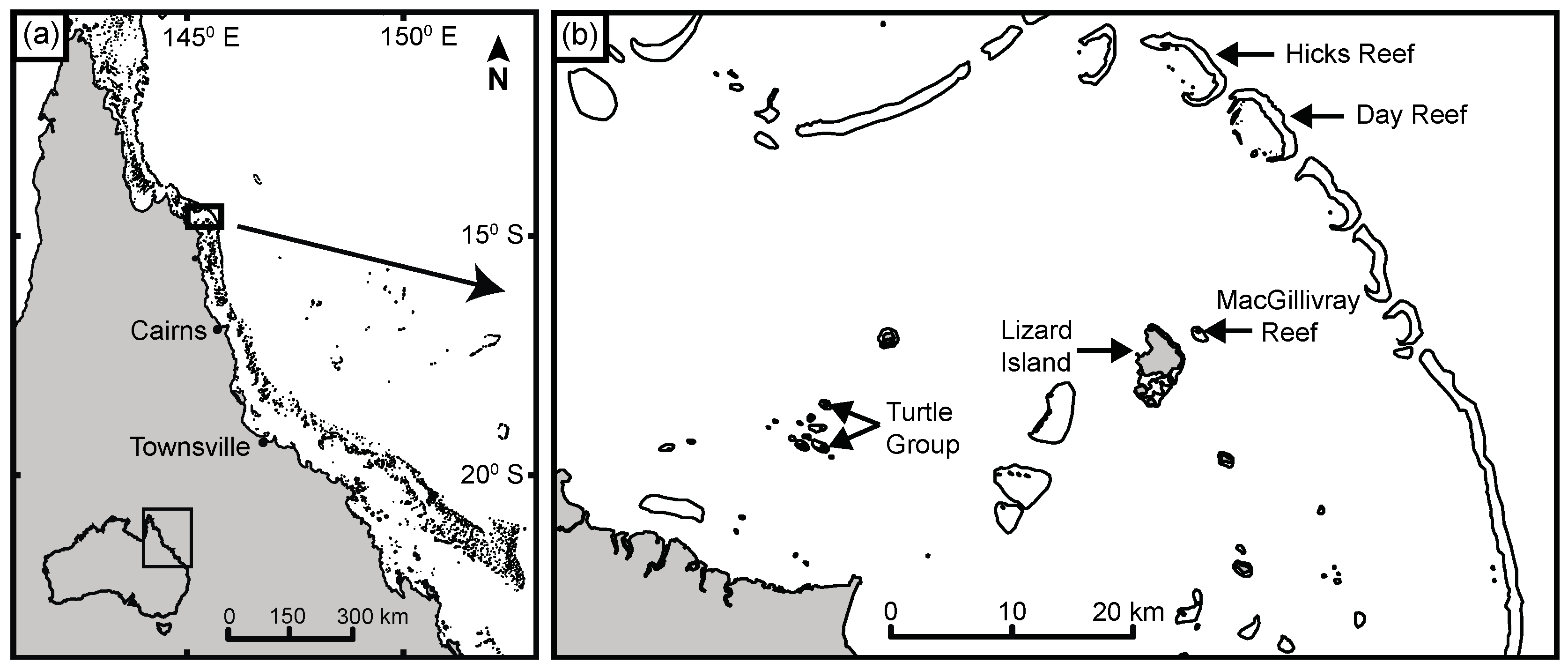
2.1. Field Sampling
2.2. Data Analyses
3. Results
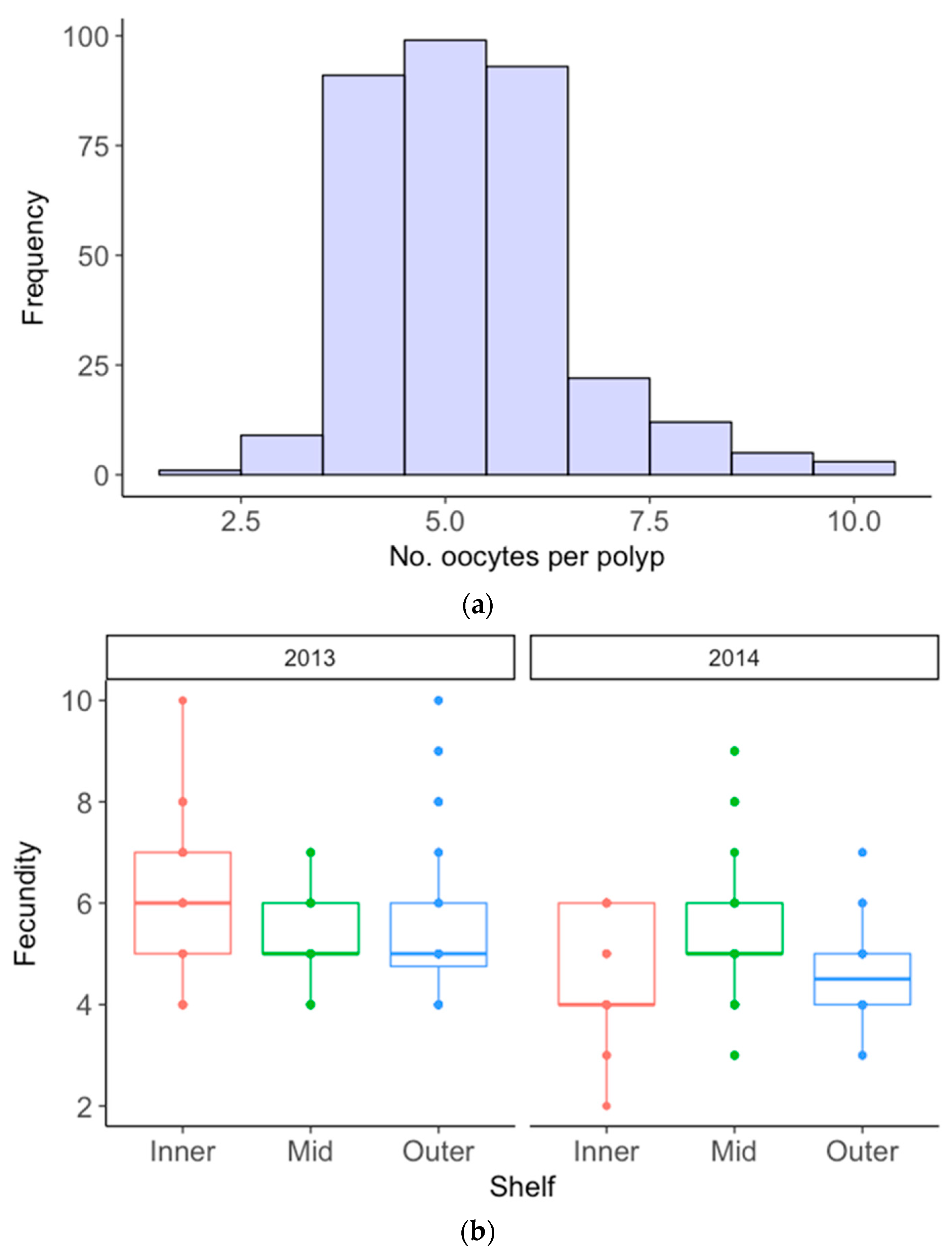
3.1. Acropora Hyacinthus
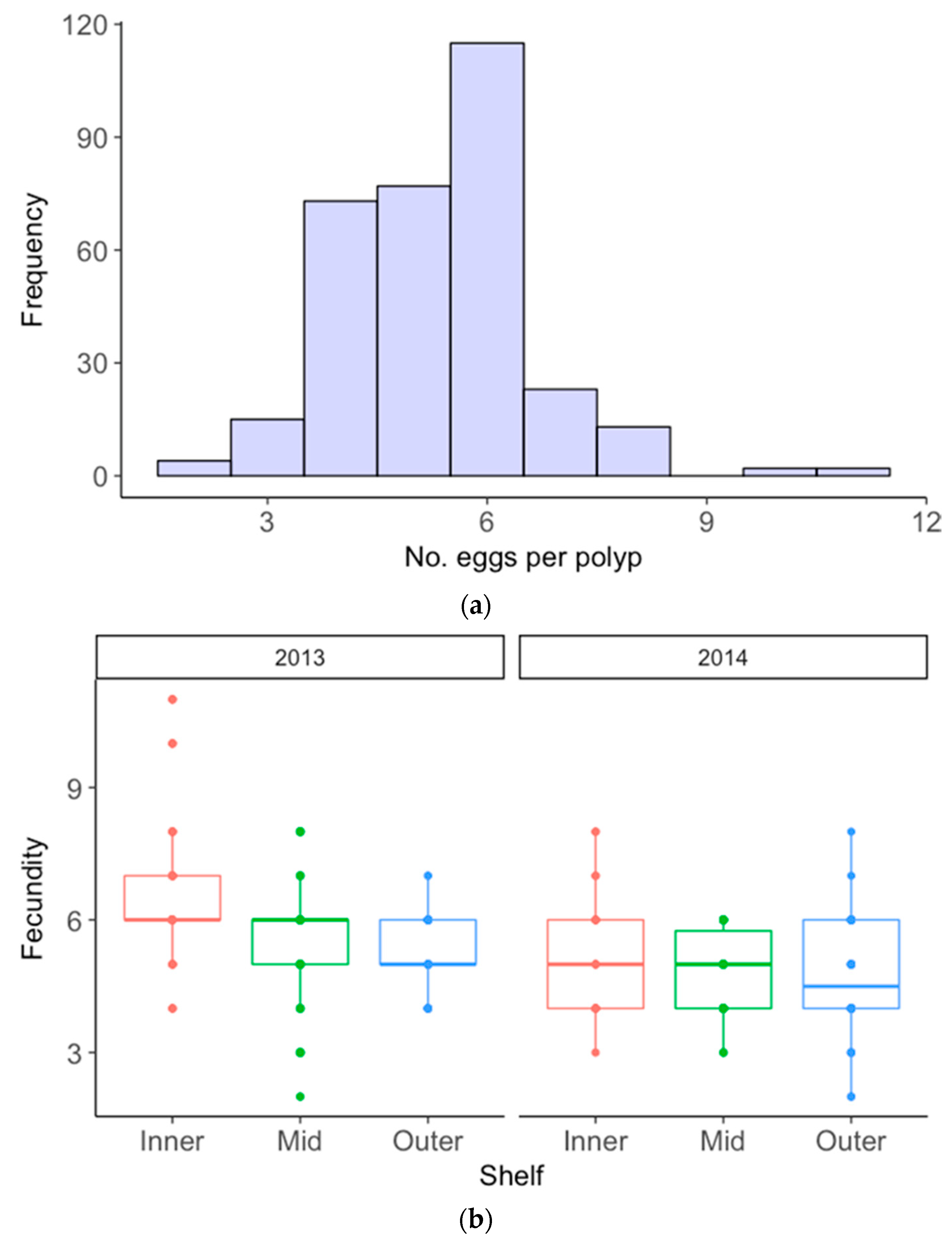
3.2. Acropora Nasuta
3.3. Acropora Spathulata
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, T.P. Recruitment limitation, mortality, and population regulation in open systems: A case study. Ecology 1990, 71, 12–20. [Google Scholar] [CrossRef]
- Baird, A.H.; Marshall, P.A. Mortality, growth and reproduction in scleractinian corals following bleaching on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2002, 237, 133–141. [Google Scholar] [CrossRef]
- Palmer, C.V.; Baird, A.H. Tumor-like growths induce an immune response and reduce fitness in a common reef-building coral. Dis. Aquat. Org. 2018, 130, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Babcock, R.C. Comparative demography of three species of scleractinian corals using age-and size-dependent classifications. Ecol. Monogr. 1991, 61, 225–244. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.A.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Supply-side ecology works both ways: The link between benthic adults, fecundity, and larval recruits. Ecology 2000, 81, 2241–2249. [Google Scholar] [CrossRef]
- Hall, V.R.; Hughes, T.P. Reproductive strategies of modular organisms: Comparative studies of reef-building corals. Ecology 1996, 77, 950–963. [Google Scholar] [CrossRef]
- Alvarez-Noriega, M.; Baird, A.H.; Dornelas, M.; Madin, J.S.; Cumbo, V.R.; Connolly, S.R. Fecundity and the demographic strategies of coral morphologies. Ecology 2016, 97, 3485–3493. [Google Scholar] [CrossRef]
- Tan, C.H.; Pratchett, M.S.; Bay, L.K.; Graham, E.M.; Baird, A.H. Spatial and temporal variation in fecundity among populations of Acropora millepora on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2016, 561, 147–153. [Google Scholar] [CrossRef]
- Tanner, J.E. Interspecific competition reduces fitness in scleractinian corals. J. Exp. Mar. Biol. Ecol. 1997, 214, 19–34. [Google Scholar] [CrossRef]
- Wallace, C.C. Reproduction, recruitment and fragmentation in nine sympatric species of the coral genus Acropora. Mar. Biol. 1985, 88, 217–233. [Google Scholar] [CrossRef]
- Michalek-Wagner, K.; Willis, B.L. Impacts of bleaching on the soft coral Lobophytum compactum. II. Biochemical changes in adults and their eggs. Coral Reefs 2001, 19, 240–246. [Google Scholar] [CrossRef]
- Baird, A.H.; Álvarez-Noriega, M.; Cumbo, V.R.; Connolly, S.R.; Dornelas, M.; Madin, J.S. Effects of tropical storms on the demography of reef corals. Mar. Ecol. Prog. Ser. 2018, 606, 29–38. [Google Scholar] [CrossRef]
- Hughes, T.P.; Day, J.C.; Brodie, J. Securing the future of the Great Barrier Reef. Nat. Clim. Chang. 2015, 5, 508–511. [Google Scholar] [CrossRef]
- Kroon, F.J.; Thorburn, P.; Schaffelke, B.; Whitten, S. Towards protecting the Great Barrier Reef from land-based pollution. Glob. Chang. Biol. 2016, 22, 1985–2002. [Google Scholar] [CrossRef]
- Roff, G.; Clark, T.R.; Reymond, C.E.; Zhao, J.X.; Feng, Y.; McCook, L.J.; Done, T.J.; Pandolfi, J.M. Palaeoecological evidence of a historical collapse of corals at Pelorus Island, inshore Great Barrier Reef, following European settlement. Proc. R. Soc. B 2013, 280, 20122100. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Bridge, T.C.; Brodie, J.; Cameron, D.S.; Day, J.C.; Emslie, M.J.; Grech, A.; Hamann, M.; Heron, S.F.; Hoey, A.S.; et al. Australia’s Great Barrier Reef. In World Seas: An Environmental Evaluation; Academic Press: Cambridge, MA, USA, 2013; pp. 333–362. [Google Scholar]
- Anthony, K.R.; Fabricius, K.E. Shifting roles of heterotrophy and autotrophy in coral energetics under varying turbidity. J. Exp. Mar. Biol. Ecol. 2000, 252, 221–253. [Google Scholar] [CrossRef]
- Anthony, K.R. Enhanced energy status of corals on coastal, high-turbidity reefs. Mar. Ecol. Prog. Ser. 2006, 319, 111–116. [Google Scholar] [CrossRef]
- Dodge, R.E.; Vaisnys, J.R. Coral populations and growth patterns: Responses to sedimentation and turbidity associated with dredging. J. Mar. Sci. 1977, 35, 715–730. [Google Scholar]
- Kojis, B.L.; Quinn, N.J. Seasonal and depth variation in fecundity of Acropora palifera at two reefs in Papua New Guinea. Coral Reefs 1984, 3, 165–172. [Google Scholar] [CrossRef]
- Wismer, S.; Hoey, A.S.; Bellwood, D.R. Cross-shelf benthic community structure on the Great Barrier Reef: Relationships between macroalgal cover and herbivore biomass. Mar. Ecol. Prog. Ser. 2009, 376, 45–54. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in browsing intensity on the Great Barrier Reef. Coral Reefs 2010, 29, 499–508. [Google Scholar] [CrossRef]
- Dikou, A.; Van Woesik, R. Survival under chronic stress from sediment load: Spatial patterns of hard coral communities in the southern islands of Singapore. Mar. Pollut. Bull. 2006, 52, 1340–1354. [Google Scholar] [CrossRef] [PubMed]
- Bauman, A.G.; Hoey, A.S.; Dunshea, G.; Feary, D.A.; Low, J.; Todd, P.A. Macroalgal browsing on a heavily degraded, urbanized equatorial reef system. Sci. Rep. 2017, 7, 8352. [Google Scholar] [CrossRef]
- Foster, N.L.; Box, S.J.; Mumby, P.J. Competitive effects of macroalgae on the fecundity of the reef-building coral Montastraea annularis. Mar. Ecol. Prog. Ser. 2008, 367, 143–152. [Google Scholar] [CrossRef]
- Tanner, J.E. Competition between scleractinian corals and macroalgae: An experimental investigation of coral growth, survival and reproduction. J. Exp. Mar. Biol. Ecol. 1995, 190, 151–168. [Google Scholar] [CrossRef]
- Rasher, D.B.; Hay, M.E. Chemically rich seaweeds poison corals when not controlled by herbivores. Proc. Natl. Acad. Sci. USA 2010, 107, 9683–9688. [Google Scholar] [CrossRef] [PubMed]
- Clements, C.S.; Rasher, D.B.; Hoey, A.S.; Bonito, V.E.; Hay, M.E. Spatial and temporal limits of coral-macroalgal competition: The negative impacts of macroalgal density, proximity, and history of contact. Mar. Ecol. Prog. Ser. 2018, 586, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Hoogenboom, M.O.; Connolly, S.R. Defining fundamental niche dimensions of corals: Synergistic effects of colony size, light, and flow. Ecology 2009, 90, 767–780. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pol. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; De Beer, D.; Lott, C.; Polerecky, L.; Kohls, K.; Abed, R.M.; Ferdelman, T.G.; Fabricius, K.E. Mechanisms of damage to corals exposed to sedimentation. Proc. Natl. Acad. Sci. USA 2012, 109, E1558–E1567. [Google Scholar] [CrossRef]
- Sammarco, P.W. Geographically specific recruitment and postsettlement mortality as influences on coral communities: The cross-continental shelf transplant experiment. Limnol. Oceanogr. 1991, 36, 496–514. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Burn, D.; Pratchett, M.; Heron, S.; Thompson, C.; Pratchett, D.; Hoey, A. Limited Cross-Shelf Variation in the Growth of Three Branching Corals on Australia’s Great Barrier Reef. Diversity 2018, 10, 122. [Google Scholar] [CrossRef]
- Morgan, K.M.; Perry, C.T.; Smithers, S.G.; Johnson, J.A.; Daniell, J.J. Evidence of extensive reef development and high coral cover in nearshore environments: Implications for understanding coral adaptation in turbid settings. Sci. Rep. 2016, 6, 29616. [Google Scholar] [CrossRef] [PubMed]
- Browne, N.K.; Smithers, S.G.; Perry, C.T. Spatial and temporal variations in turbidity on two inshore turbid reefs on the Great Barrier Reef, Australia. Coral Reefs 2013, 32, 195–210. [Google Scholar] [CrossRef]
- Johns, K.A.; Emslie, M.J.; Hoey, A.S.; Osborne, K.; Jonker, M.J.; Cheal, A.J. Macroalgal feedbacks and substrate properties maintain a coral reef regime shift. Ecosphere 2018, 9, e02349. [Google Scholar] [CrossRef]
- Halford, A.; Cheal, A.J.; Ryan, D.; Williams, D.M. Resilience to large-scale disturbance in coral and fish assemblages on the Great Barrier Reef. Ecology 2004, 85, 1892–1905. [Google Scholar] [CrossRef]
- Torda, G.; Sambrook, K.; Cross, P.; Sato, Y.; Bourne, D.G.; Lukoschek, V.; Hill, T.; Jorda, G.T.; Moya, A.; Willis, B.L. Decadal erosion of coral assemblages by multiple disturbances in the Palm Islands, central Great Barrier Reef. Sci. Rep. 2018, 8, 11885. [Google Scholar] [CrossRef]
- Madin, J.S.; Connolly, S.R. Ecological consequences of major hydrodynamic disturbances on coral reefs. Nature 2006, 444, 477–480. [Google Scholar] [CrossRef]
- Rinkevich, B.; Loya, Y. Intraspecific competition in a reef coral: Effects on growth and reproduction. Oecologia 1985, 66, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Madin, J.S.; Baird, A.H.; Bridge, T.C.; Connolly, S.R.; Zawada, K.J.; Dornelas, M. Cumulative effects of cyclones and bleaching on coral cover and species richness at Lizard Island. Mar. Ecol. Prog. Ser. 2018, 604, 263–268. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock-recruitment dynamics of corals. Nature 2019. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.A.; Dolman, A.M. Coral bleaching: One disturbance too many for near-shore reefs of the Great Barrier Reef. Coral Reefs 2010, 29, 637–648. [Google Scholar] [CrossRef]
- Thompson, A.; Schroeder, T.; Brando, V.E.; Schaffelke, B. Coral community responses to declining water quality: Whitsunday Islands, Great Barrier Reef, Australia. Coral Reefs 2014, 33, 923–938. [Google Scholar] [CrossRef]
- Doszpot, N.E.; McWilliam, M.J.; Pratchett, M.S.; Hoey, A.S.; Figueira, W.F. Plasticity in three-dimensional geometry of branching corals along a cross-shelf cradient. Diversity 2019, 11, 44. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Distance Offshore | A. hyacinthus | A. nasuta | A. spathulata | Total | |||
|---|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | |||
| Turtle South | 12 km | 5 | 5 | 5 | 5 | 5 | 0 | 25 |
| Turtle North | 15 km | 4 | 2 | 5 | 5 | 5 | 5 | 26 |
| Lizard | 30 km | 5 | 5 | 5 | 5 | 5 | 4 | 29 |
| MacGillivray | 34 km | 5 | 5 | 5 | 4 | 5 | 5 | 29 |
| Day | 50 km | 5 | 5 | 5 | 5 | 5 | 5 | 30 |
| Hicks | 51 km | 5 | 5 | 5 | 5 | 5 | 5 | 30 |
| TOTAL | 29 | 27 | 30 | 29 | 30 | 24 | 169 | |
| Coefficient | Estimate | SE | p |
|---|---|---|---|
| Intercept (inner-shelf 2013) | 2351.37 | 494.96 | <0.01 |
| Shelf (mid-shelf) | −2419.19 | 660.58 | <0.01 |
| Shelf (outer-shelf) | −366.27 | 659.49 | 0.57 |
| Year (2014) | −1.65 | 0.25 | <0.01 |
| Shelf (mid-shelf): Year (2014) | 1.21 | 0.32 | <0.01 |
| Shelf (outer-shelf): Year (2104) | 0.18 | 0.32 | 0.57 |
| Coefficient | Estimate | SE | p |
|---|---|---|---|
| Intercept (inner-shelf 2013) | 5646.88 | 932.03 | <0.01 |
| Shelf (mid-shelf) | −6788.74 | 1337.45 | <0.01 |
| Shelf (outer-shelf | −5270.63 | 1318.10 | <0.01 |
| Year (2014) | −2.810 | 0.46 | <0.01 |
| Shelf (mid-shelf): Year (2014) | 3.37 | 0.66 | <0.01 |
| Shelf (outer-shelf): Year (2104) | 2.62 | 0.65 | <0.01 |
| Coefficient | Estimate | SE | p |
|---|---|---|---|
| Intercept (inner-shelf 2013) | 2858.27 | 191.99 | <0.01 |
| Shelf (mid-shelf) | −1770.71 | 255.02 | <0.01 |
| Shelf (outer-shelf | −1510.93 | 252.99 | 0.02 |
| Year (2014) | −1.42 | 0.26 | <0.01 |
| Shelf (mid-shelf): Year (2014) | 0.88 | 0.33 | <0.01 |
| Shelf (outer-shelf): Year (2104) | 0.75 | 0.33 | 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratchett, M.S.; Hoey, A.S.; Tan, C.-H.; Kuo, C.-Y.; Bauman, A.G.; Kumaraswamy, R.; Baird, A.H. Spatial and Temporal Variation in Fecundity of Acropora spp. in the Northern Great Barrier Reef. Diversity 2019, 11, 60. https://doi.org/10.3390/d11040060
Pratchett MS, Hoey AS, Tan C-H, Kuo C-Y, Bauman AG, Kumaraswamy R, Baird AH. Spatial and Temporal Variation in Fecundity of Acropora spp. in the Northern Great Barrier Reef. Diversity. 2019; 11(4):60. https://doi.org/10.3390/d11040060
Chicago/Turabian StylePratchett, Morgan S., Andrew S. Hoey, Chun-Hong Tan, Chao-Yang Kuo, Andrew G. Bauman, Rajani Kumaraswamy, and Andrew H. Baird. 2019. "Spatial and Temporal Variation in Fecundity of Acropora spp. in the Northern Great Barrier Reef" Diversity 11, no. 4: 60. https://doi.org/10.3390/d11040060
APA StylePratchett, M. S., Hoey, A. S., Tan, C.-H., Kuo, C.-Y., Bauman, A. G., Kumaraswamy, R., & Baird, A. H. (2019). Spatial and Temporal Variation in Fecundity of Acropora spp. in the Northern Great Barrier Reef. Diversity, 11(4), 60. https://doi.org/10.3390/d11040060