Cross-Shelf Variation in Coral Community Response to Disturbance on the Great Barrier Reef


Abstract
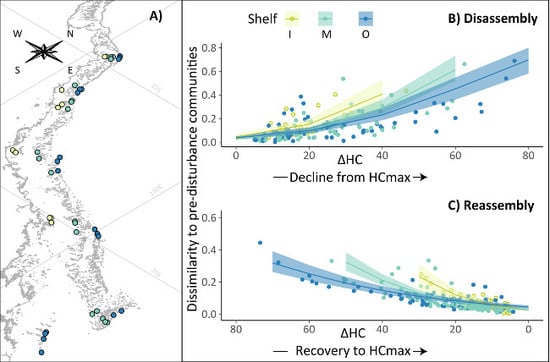
:
1. Introduction
2. Materials and Methods
2.1. Survey Reefs
2.2. Survey Methods
2.3. Disturbance Data
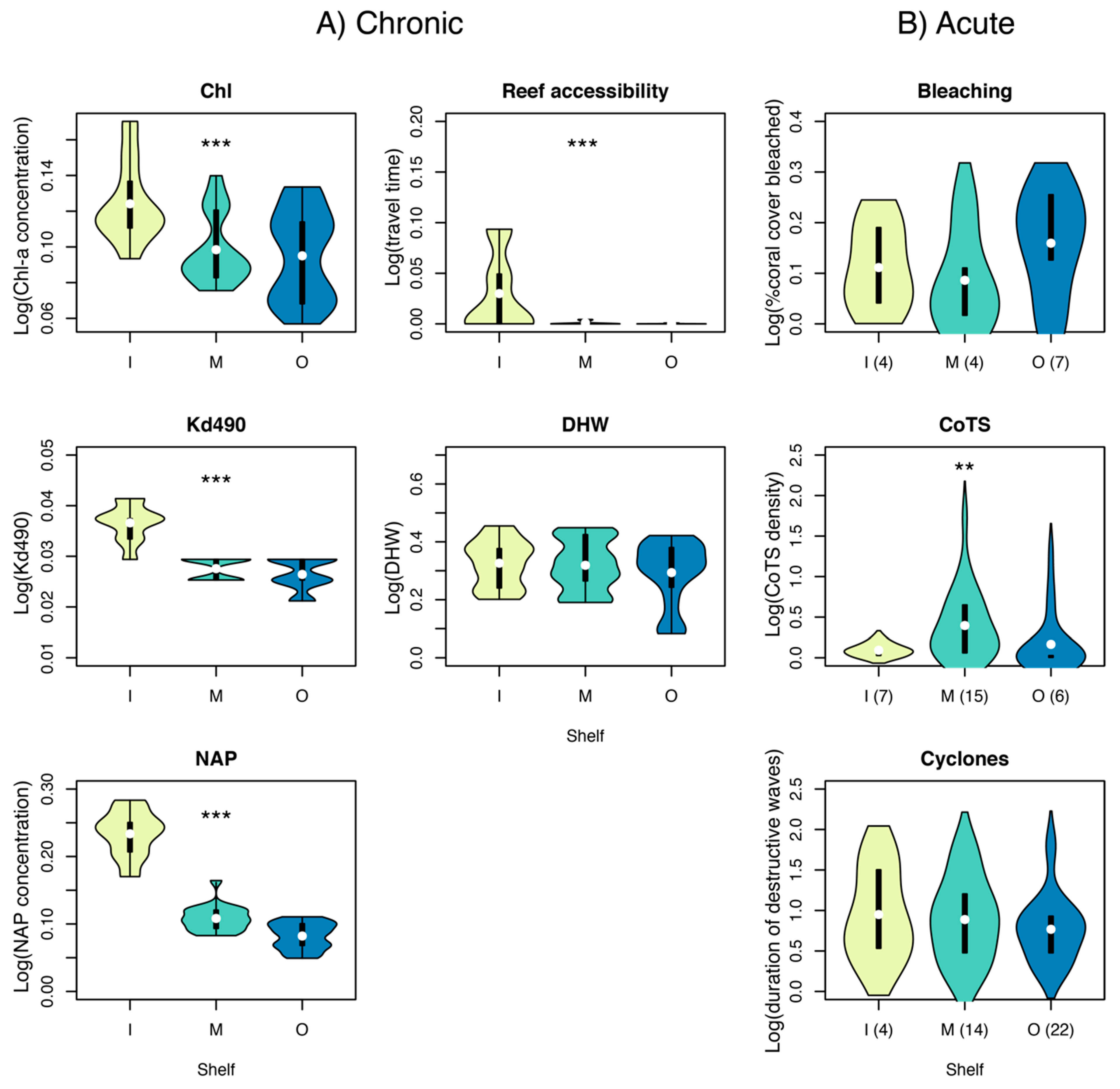
2.3.1. Chronic Stressors
2.3.2. Acute Stressors
2.4. Modelling
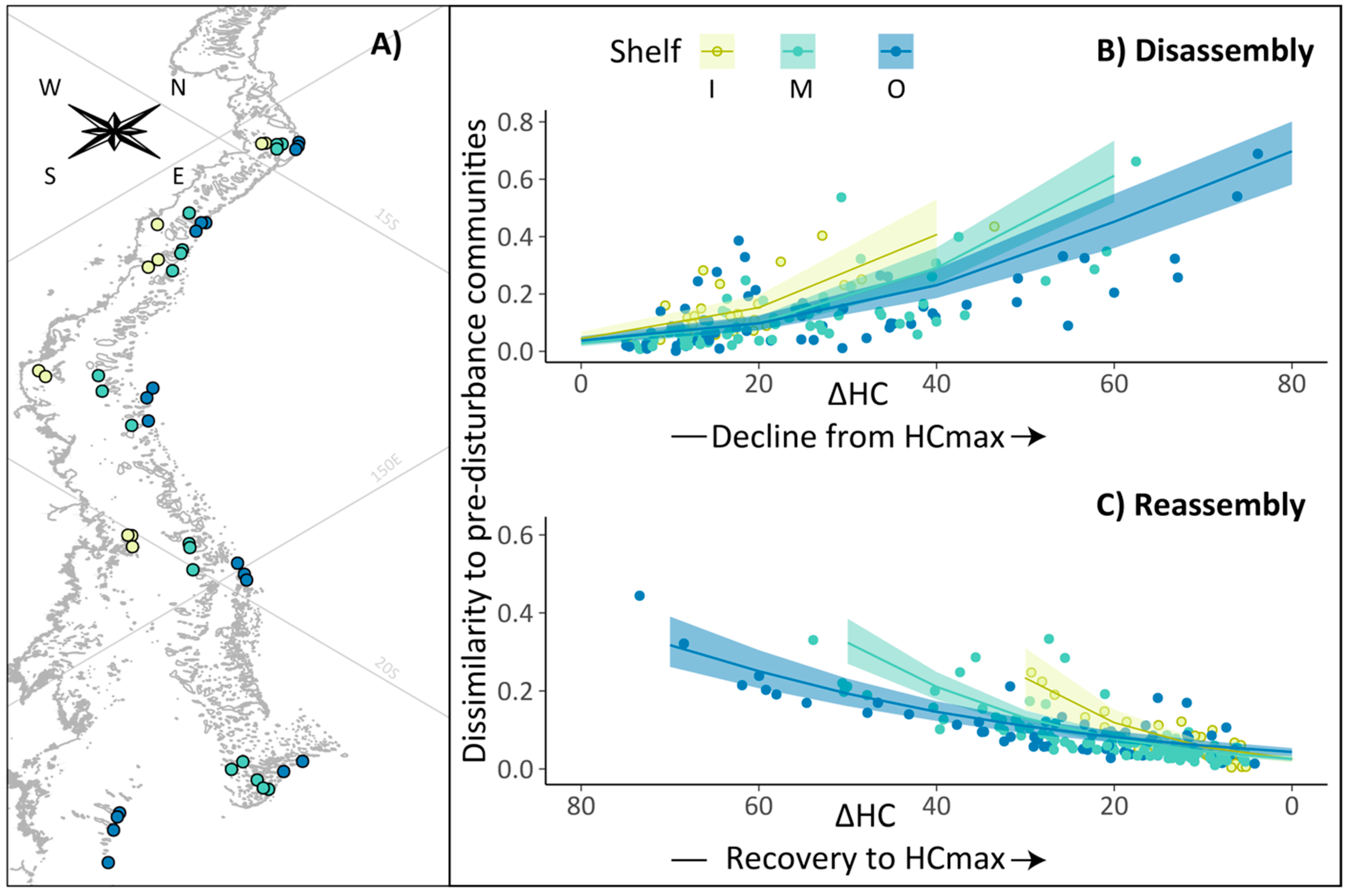
2.4.1. Calculation of Disassembly and Reassembly
2.4.2. Pre-Disturbance Communities
2.4.3. Hierarchical Linear Models
3. Results
3.1. Community Disassembly and Reassembly
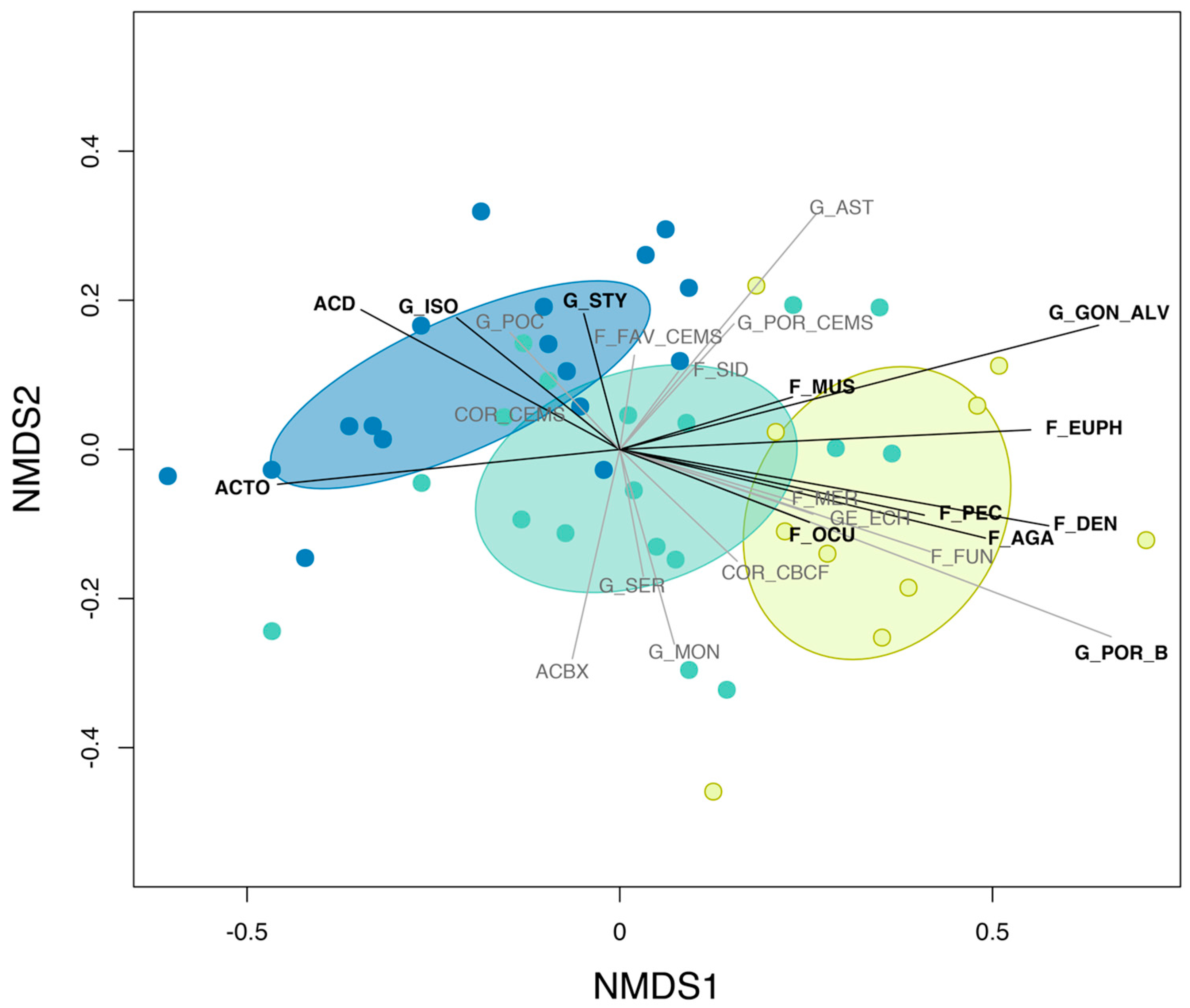
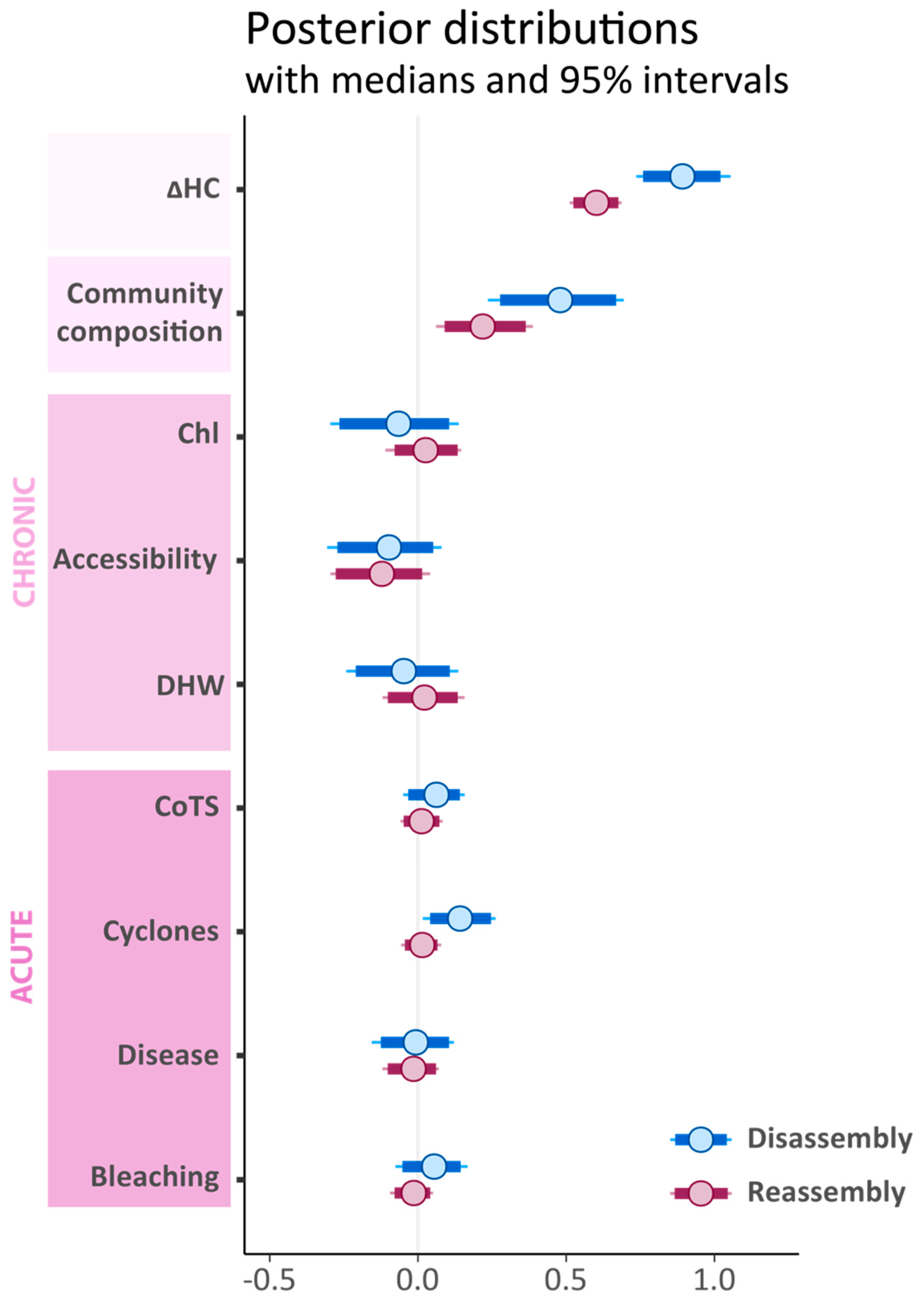
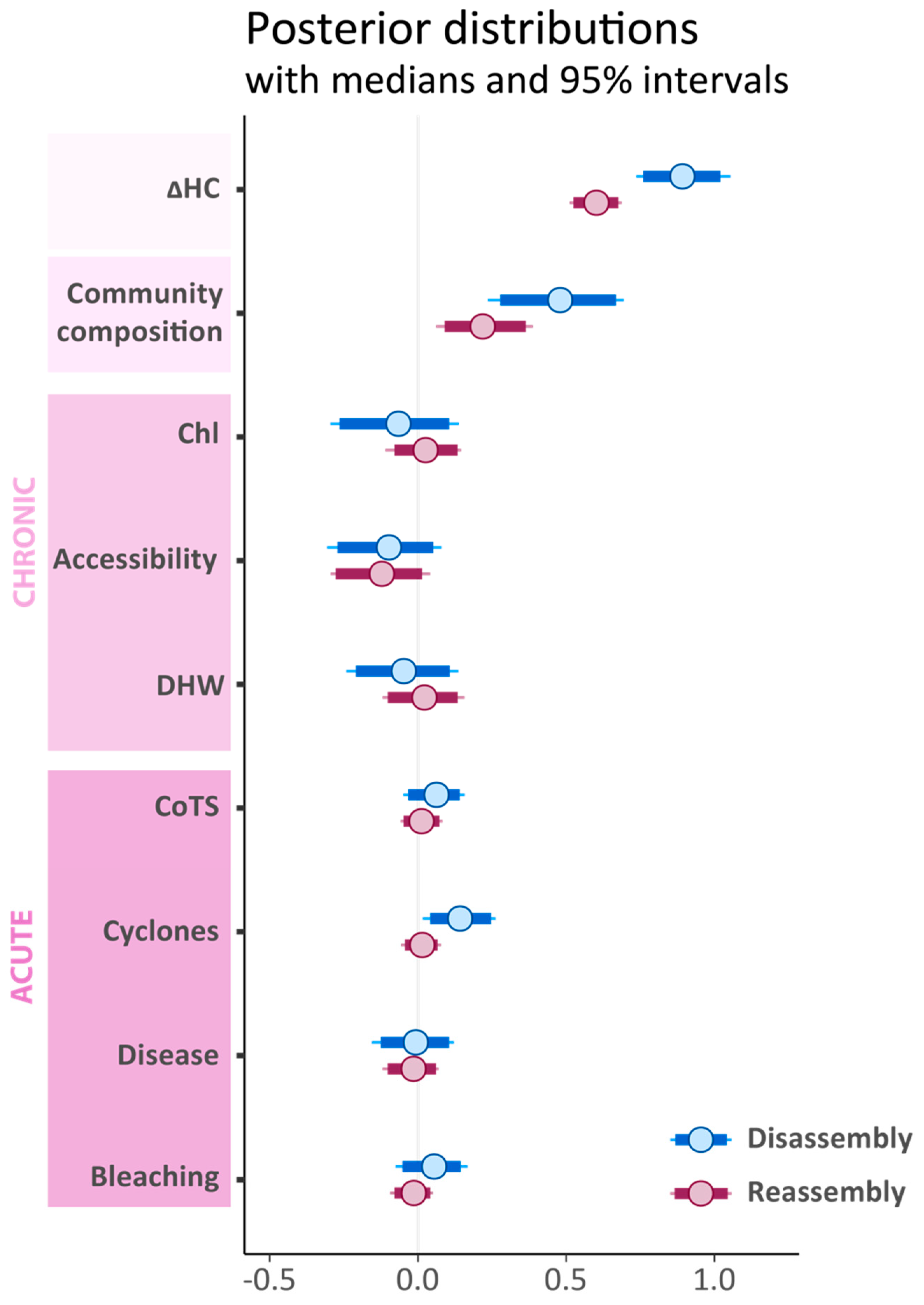
3.2. Effects of Pre-Disturbance Community Composition, Chronic and Acute Stressors
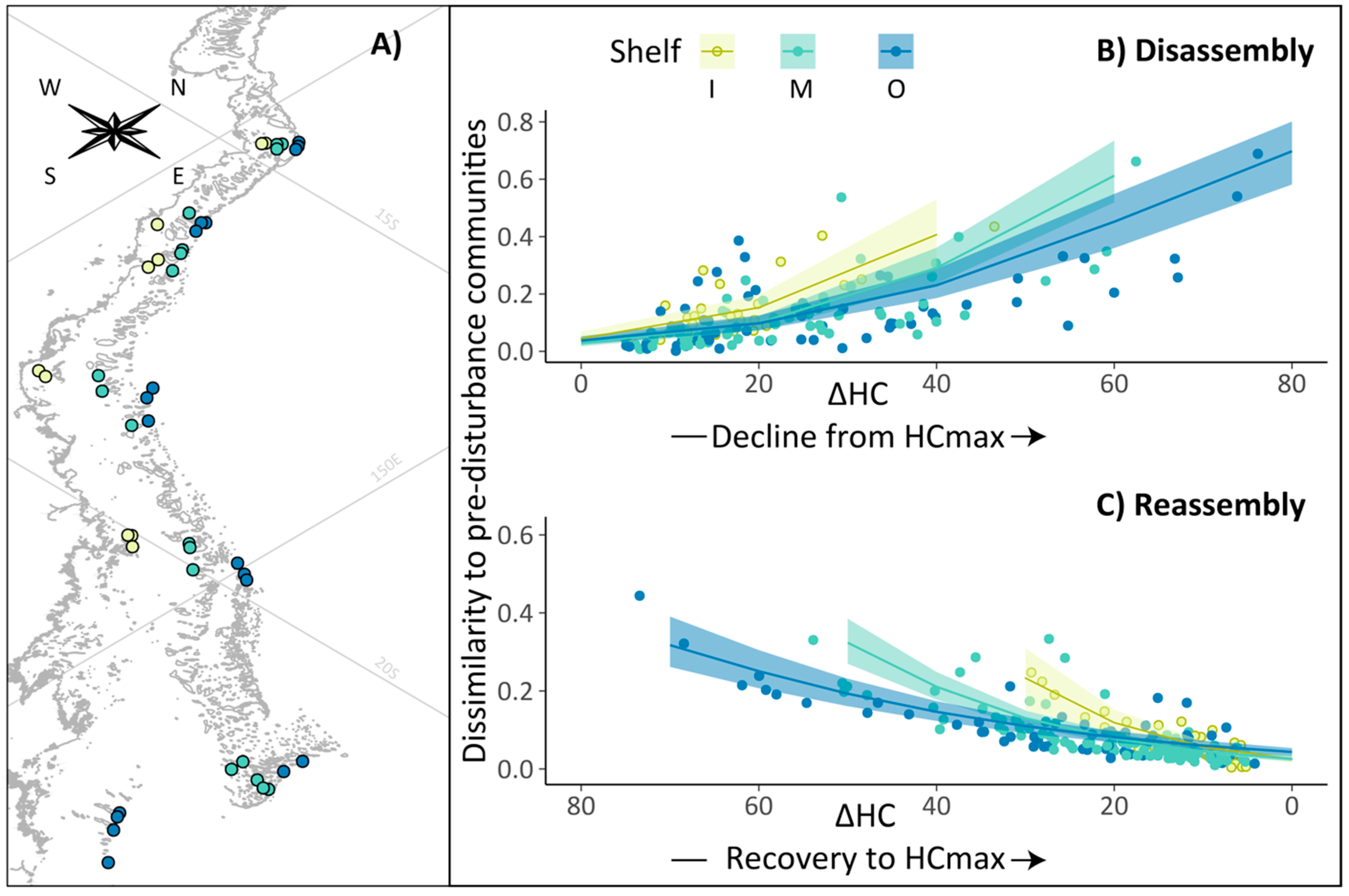
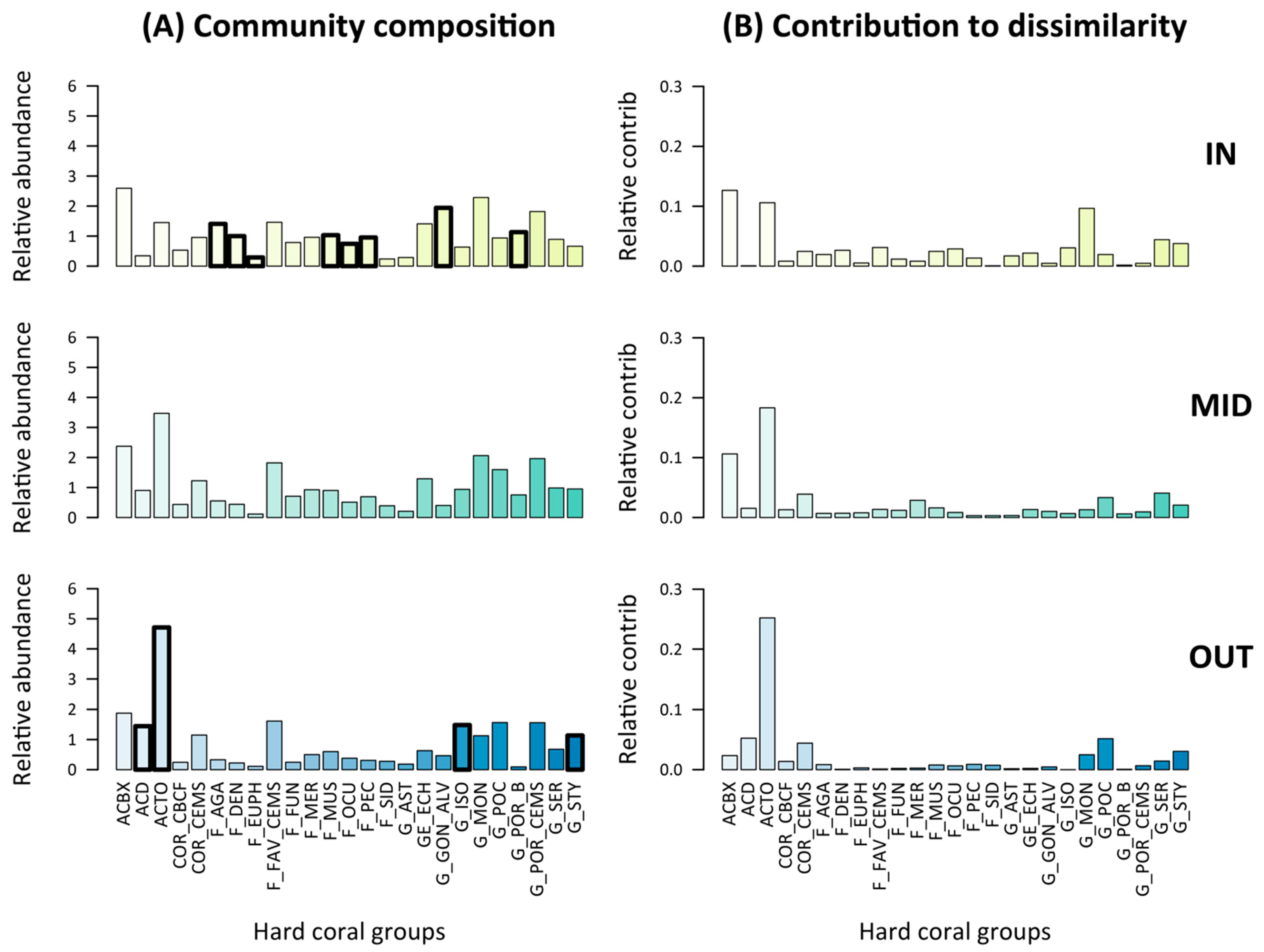
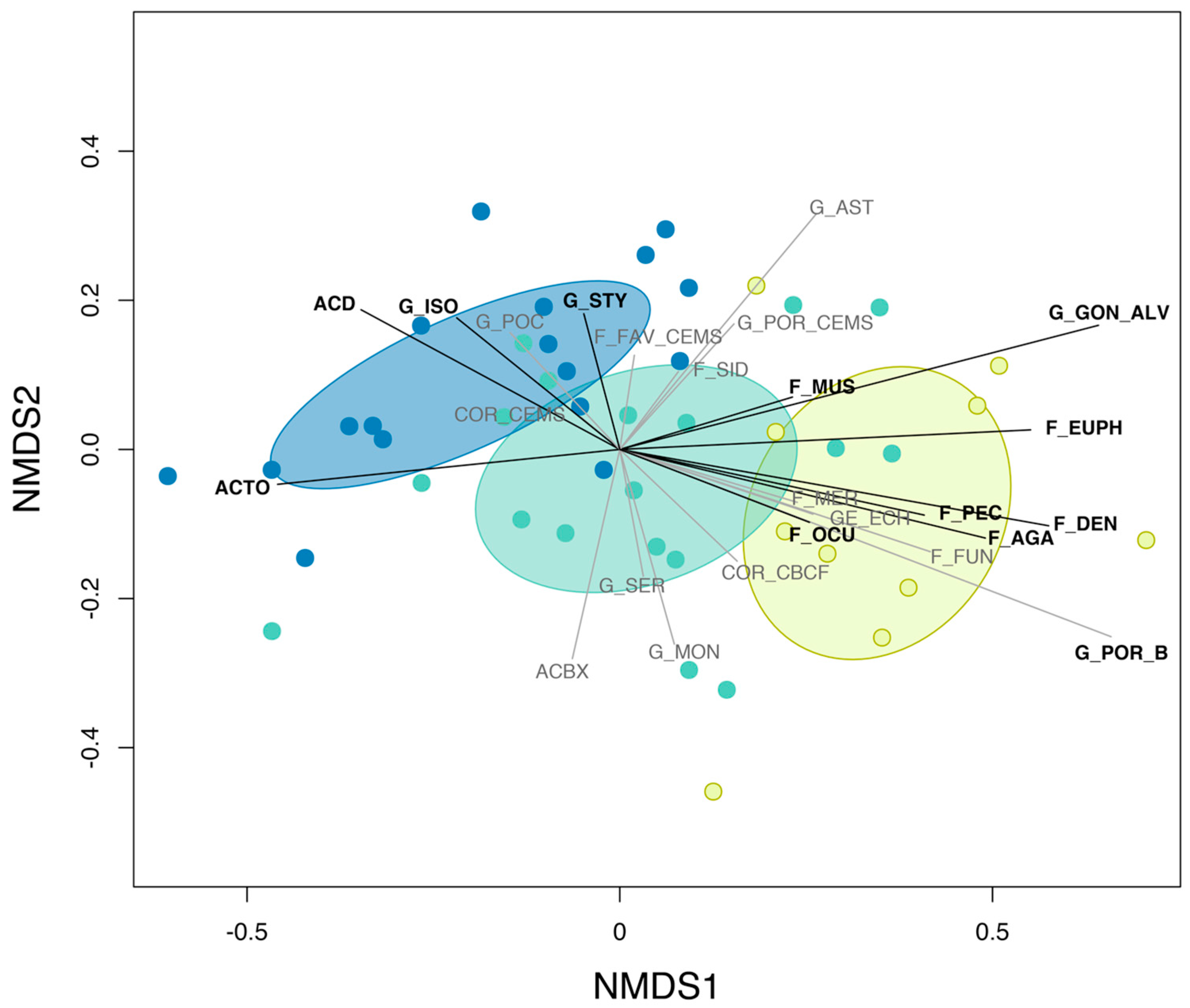
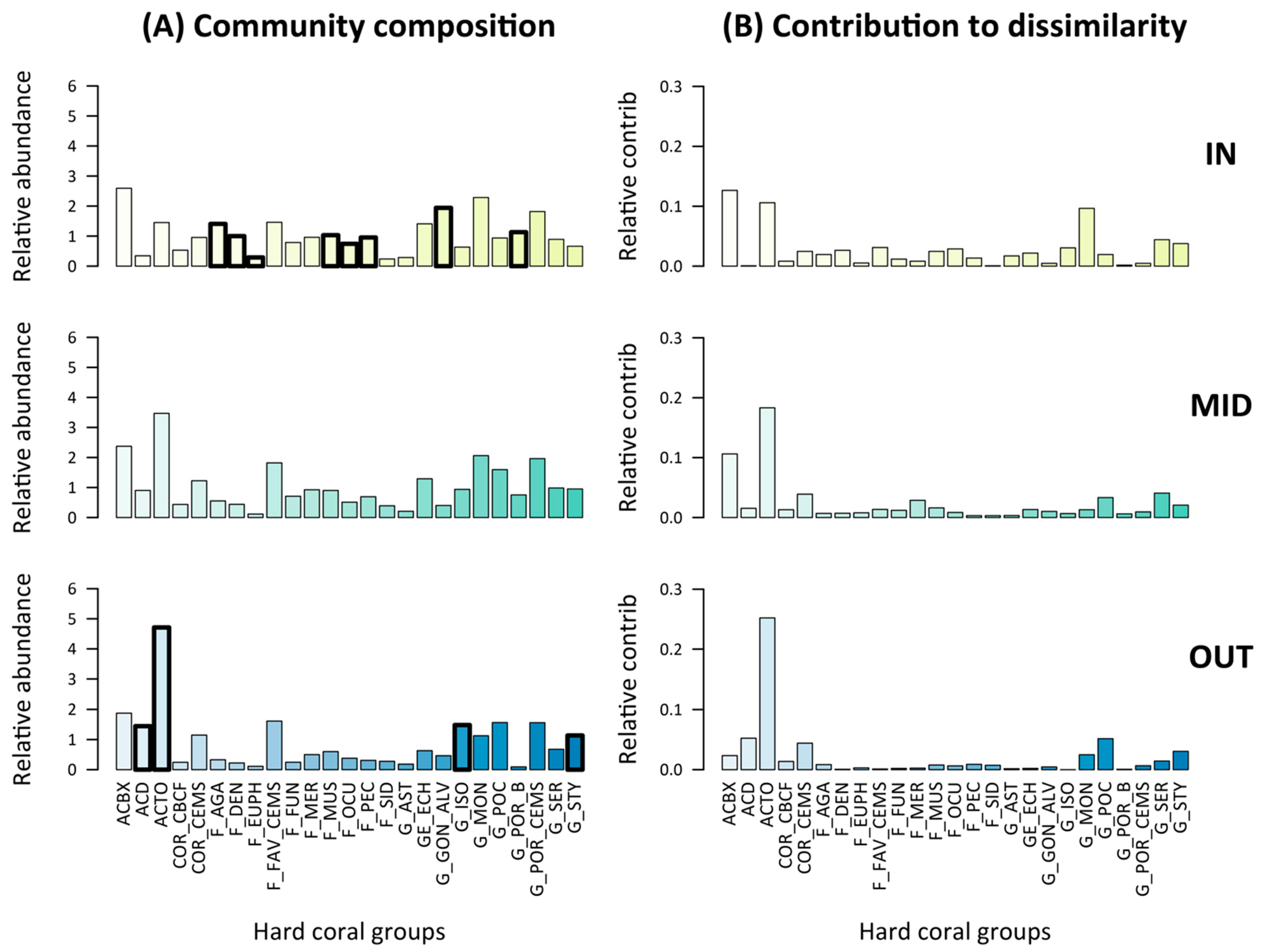
3.3. Pre-Disturbance Community Composition and Taxon-Specific Contribution to Disassembly and Reassembly
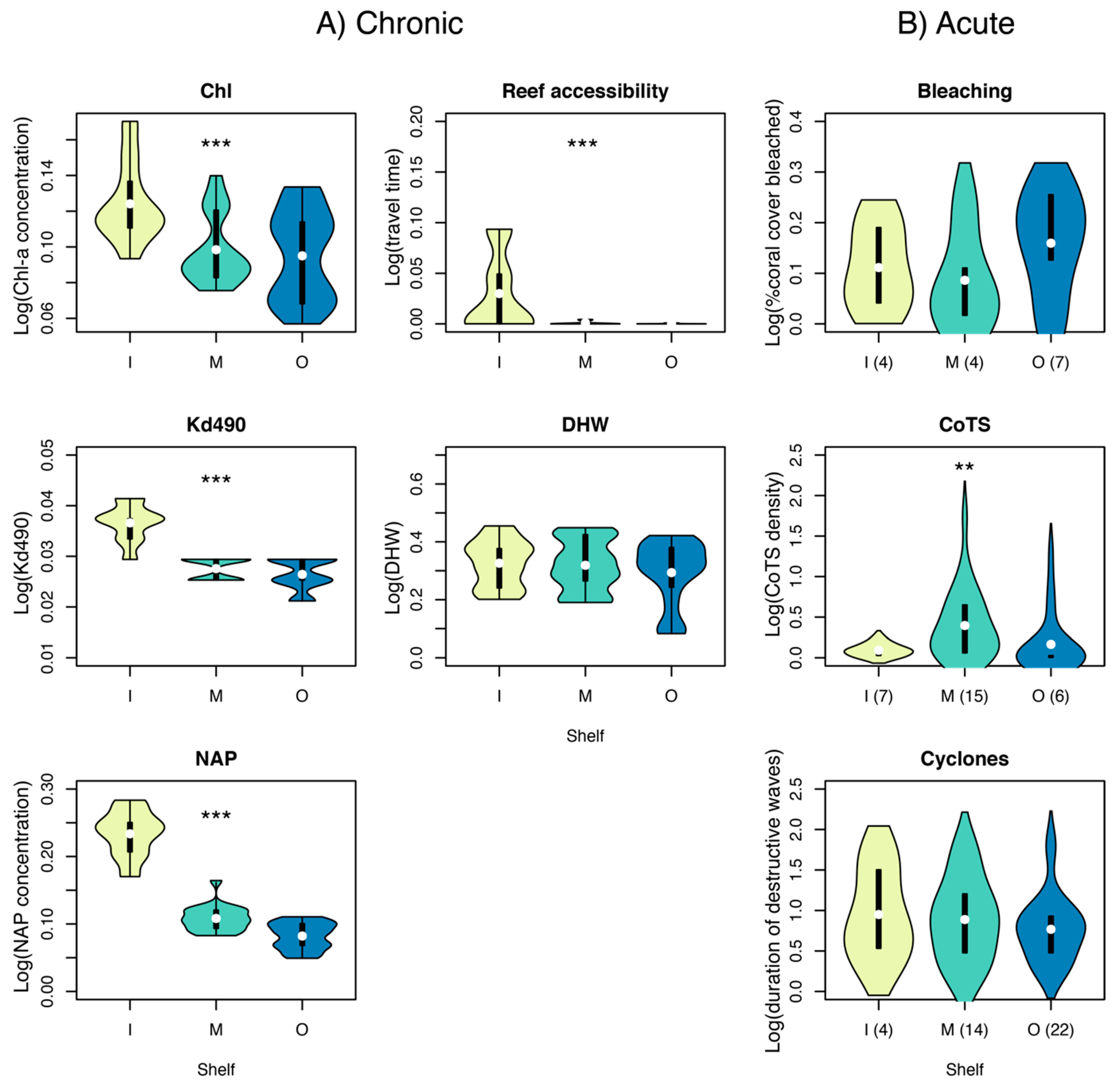
3.4. Cross-Shelf Variation in Chronic and Acute Disturbance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Alvarez-Noriega, M.; Alvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373. [Google Scholar] [CrossRef] [PubMed]
- Cheal, A.J.; Macneil, M.A.; Emslie, M.J.; Sweatman, H. The threat to coral reefs from more intense cyclones under climate change. Glob. Chang. Biol. 2017, 23, 1511–1524. [Google Scholar] [CrossRef] [PubMed]
- Osborne, K.; Thompson, A.A.; Cheal, A.J.; Emslie, M.J.; Johns, K.A.; Jonker, M.J.; Logan, M.; Miller, I.R.; Sweatman, H.P.A. Delayed coral recovery in a warming ocean. Glob. Chang. Biol. 2017, 23, 3869–3881. [Google Scholar] [CrossRef] [PubMed]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Hughes, T.P.; Graham, N.A.J.; Jackson, J.B.C.; Mumby, P.J.; Steneck, R.S. Rising to the challenge of sustaining coral reef resilience. Trends Ecol. Evol. 2010, 25, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Darling, E.S.; McClanahan, T.R.; Cote, I.M. Life histories predict coral community disassembly under multiple stressors. Glob. Chang. Biol. 2013, 19, 1930–1940. [Google Scholar] [CrossRef]
- Graham, N.A.; Chabanet, P.; Evans, R.D.; Jennings, S.; Letourneur, Y.; MacNeil, M.; McClanahan, T.R.; Ohman, M.C.; Polunin, N.V.; Wilson, S.K. Extinction vulnerability of coral reef fishes. Ecol. Lett. 2011, 14, 341–348. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef]
- Johns, K.; Osborne, K.; Logan, M. Contrasting rates of coral recovery and reassembly in coral communities on the Great Barrier Reef. Coral Reefs 2014, 33, 553–563. [Google Scholar] [CrossRef]
- Gilmour, J.P.; Smith, L.D.; Heyward, A.J.; Baird, A.H.; Pratchett, M.S. Recovery of an Isolated Coral Reef System Following Severe Disturbance. Science 2013, 340, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Adjeroud, M.; Michonneau, F.; Edmunds, P.J.; Chancerelle, Y.; de Loma, T.L.; Penin, L.; Thibaut, L.; Vidal-Dupiol, J.; Salvat, B.; Galzin, R. Recurrent disturbances, recovery trajectories, and resilience of coral assemblages on a South Central Pacific reef. Coral Reefs 2009, 28, 775–780. [Google Scholar] [CrossRef]
- Darling, E.S.; Alvarez-Filip, L.; Oliver, T.A.; McClanahan, T.R.; Cote, I.M. Evaluating life-history strategies of reef corals from species traits. Ecol. Lett. 2012, 15, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Baird, A.H.; Marshall, P.A. Mortality, growth and reproduction in scleractinian corals following bleaching on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2002, 237, 133–141. [Google Scholar] [CrossRef]
- Kayal, M.; Lenihan, H.S.; Brooks, A.J.; Holbrook, S.J.; Schmitt, R.J.; Kendall, B.E. Predicting coral community recovery using multi-species population dynamics models. Ecol. Lett. 2018, 21, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Mellin, C.; Bradshaw, C.J.A.; Meekan, M.G.; Caley, M.J. Environmental and spatial predictors of species richness and abundance in coral reef fishes. Glob. Ecol. Biogeogr. 2010, 19, 212–222. [Google Scholar] [CrossRef]
- Pitcher, C.R.; Lawton, P.; Ellis, N.; Smith, S.J.; Incze, L.S.; Wei, C.L.; Greenlaw, M.E.; Wolff, N.H.; Sameoto, J.A.; Snelgrove, P.V. Exploring the role of environmental variables in shaping patterns of seabed biodiversity composition in regional-scale ecosystems. J. Appl. Ecol. 2012, 49, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Petus, C.; da Silva, E.; Tracey, D.; Wolff, N.H.; Waterhouse, J.; Brodie, J. Water quality and river plume monitoring in the Great Barrier Reef: An overview of methods based on ocean colour satellite data. Remote Sens.-Basel 2015, 7, 12909–12941. [Google Scholar] [CrossRef]
- Petus, C.; da Silva, E.T.; Devlin, M.; Wenger, A.S.; Alvarez-Romero, J.G. Using MODIS data for mapping of water types within river plumes in the Great Barrier Reef, Australia: Towards the production of river plume risk maps for reef and seagrass ecosystems. J Env. Manag. 2014, 137, 163–177. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, M.A.; Mellin, C.; Matthews, S.; Wolff, N.H.; McClanahan, T.R.; Devlin, M.; Drovandi, C.; Mengersen, K.; Graham, N.A.J. Water quality mediated resilience on the Great Barrier Reef. Nat. Ecol. Evol. 2018, in press. [Google Scholar]
- Emslie, M.J.; Pratchett, M.S.; Cheal, A.J.; Osborne, K. Great Barrier Reef butterflyfish community structure: The role of shelf position and benthic community type. Coral Reefs 2010, 29, 705–715. [Google Scholar] [CrossRef]
- Mellin, C.; Bradshaw, C.J.A.; Fordham, D.A.; Caley, M.J. Strong but opposing beta-diversity-stability relationships in coral reef fish communities. Proc. Biol. Sci./R. Soc. 2014, 281, 20131993. [Google Scholar] [CrossRef] [PubMed]
- Sweatman, H.; Cheal, A.; Coleman, G.; Jonker, M.; Johns, K.; Emslie, M.; Miller, I.; Osborne, K. Long-Term Monitoring of the Great Barrier Reef. Status Report No 8; Australian Institute of Marine Science: Townsville, Australia, 2008.
- Jonker, M.; Johns, K.; Osborne, K. Surveys of Benthic Reef Communities Using Digital Photography and Counts of Juvenile Corals. In Long-Term Monitoring of the Great Barrier Reef Standard Operational Procedure Number 10; Australian Institute of Marine Science: Townsville, Australia, 2008. [Google Scholar]
- Maire, E.; Cinner, J.; Velez, L.; Huchery, C.; Mora, C.; Dagata, S.; Vigliola, L.; Wantiez, L.; Kulbicki, M.; Mouillot, D. How accessible are coral reefs to people? A global assessment based on travel time. Ecol. Lett. 2016, 19, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Eakin, C.M.; Morgan, J.A.; Heron, S.F.; Smith, T.B.; Liu, G.; Alvarez-Filip, L.; Baca, B.; Bartels, E.; Bastidas, C.; Bouchon, C.; et al. Caribbean Corals in Crisis: Record Thermal Stress, Bleaching, and Mortality in 2005. PLoS ONE 2010, 5, e13969. [Google Scholar] [CrossRef]
- Howells, E.J.; Beltran, V.H.; Larsen, N.W.; Bay, L.K.; Willis, B.L.; van Oppen, M.J.H. Coral thermal tolerance shaped by local adaptation of photosymbionts. Nat. Clim. Chang. 2011, 2, 116. [Google Scholar] [CrossRef]
- Matthews, S.; Mellin, C.; MacNeil, M.A.; Heron, S.; Puotinen, M.L.; Pratchett, M. Disturbance and environment data for the Great Barrier Reef: A comprehensive characterisation of the abiotic environment and disturbances regimes 1985–2016. Ecology 2019, 100, e02574. [Google Scholar] [CrossRef] [PubMed]
- Osborne, K.; Dolman, A.M.; Burgess, S.C.; Johns, K.A. Disturbance and the dynamics of coral cover on the Great Barrier Reef (1995–2009). PLoS ONE 2011, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Berkelmans, R.; De’ath, G.; Kininmonth, S.; Skirving, W.J. A comparison of the 1998 and 2002 coral bleaching events on the Great Barrier Reef: Spatial correlation, patterns, and predictions. Coral Reefs 2004, 23, 74–83. [Google Scholar] [CrossRef]
- Miller, I.; Müller, R. Validity and reproducibility of benthic cover estimates made during broadscale surveys of coral reefs by manta tow. Coral Reefs 1999, 18, 353–356. [Google Scholar] [CrossRef]
- Puotinen, M.; Maynard, J.A.; Beeden, R.; Radford, B.; Williams, G.J. A robust operational model for predicting where tropical cyclone waves damage coral reefs. Sci. Rep. 2016, 6, 26009. [Google Scholar] [CrossRef]
- Clarke, K.R. Nonparametric Multivariate Analyses of Changes in Community Structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Ferrari, S.L.P.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Figueroa-Zúñiga, J.I.; Arellano-Valle, R.B.; Ferrari, S.L.P. Mixed beta regression: A Bayesian perspective. Comput. Stat. Data Anal. 2013, 61, 137–147. [Google Scholar] [CrossRef]
- Gelman, A. Prior distributions for variance parameters in hierarchical models (comment on article by Browne and Draper). Bayesian Anal. 2006, 1, 515–534. [Google Scholar] [CrossRef]
- Gelman, A.; Goodrich, B.; Gabry, J.; Vehtari, A. R-squared for Bayesian regression models. Am. Stat. 2018, in press. [Google Scholar] [CrossRef]
- Stan Development Team. R Stan: The R Interface to Stan. R Package Version 2.17.3.. 2018. Available online: http://mc-stan.org/ (accessed on 10 February 2019).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 10 February 2019).
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Martinez-Garcia, L.B.; Richardson, S.J.; Peltzer, D.A.; Dickie, I.A. Symmetric assembly and disassembly processes in an ecological network. Ecol. Lett. 2018, 21, 896–904. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J.; Johns, K.A. Retention of habitat complexity minimizes disassembly of reef fish communities following disturbance: A large-scale natural experiment. PLoS ONE 2014, 9, e105384. [Google Scholar] [CrossRef]
- Done, T. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Halford, A.; Cheal, A.J.; Ryan, D.; Williams, D.M. Resilience to large-scale disturbance in coral and fish assemblages on the Great Barrier Reef. Ecology 2004, 85, 1892–1905. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Polunin, N.V.C.; Done, T.J. Ecological states and the resilience of coral reefs. Conserv. Ecol. 2002, 6, 2–18. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Connolly, S.R.; Baird, A.H.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; Liu, G.; et al. Ecological memory modifies the cumulative impact of recurrent climate extremes. Nat. Clim. Chang. 2018, 9, 40. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Fabricius, K.E.; Bongaerts, P.; Wallace, C.C.; Muir, P.R.; Done, T.J.; Webster, J.M. Diversity of Scleractinia and Octocorallia in the mesophotic zone of the Great Barrier Reef, Australia. Coral Reefs 2012, 31, 179–189. [Google Scholar] [CrossRef]
- DeVantier, L.M.; De’ath, G.; Turak, E.; Done, T.J.; Fabricius, K.E. Species richness and community structure of reef-building corals on the nearshore Great Barrier Reef. Coral Reefs 2006, 25, 329–340. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.; Hoey, A.S. Sleeping functional group drives coral-reef recovery. Curr. Biol. 2006, 16, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Mellin, C.; MacNeil, M.A.; Cheal, A.J.; Emslie, M.J.; Caley, M.J. Marine protected areas increase resilience among coral reef communities. Ecol. Lett. 2016, 19, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, K.; De’ath, G.; McCook, L.; Turak, E.; Williams, D.M. Changes in algal, coral and fish assemblages along water quality gradients on the inshore Great Barrier Reef. Mar. Pollut. Bull. 2005, 51, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.; Schroeder, T.; Brando, V.E.; Schaffelke, B. Coral community responses to declining water quality: Whitsunday Islands, Great Barrier Reef, Australia. Coral Reefs 2014, 33, 923–938. [Google Scholar] [CrossRef]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Code | Group Description |
|---|---|
| ACBX | Acropora branching & bottlebrush |
| ACD | Acropora digitate |
| ACTO | Acropora tabulate & corymbose |
| COR_CBCF | non-Acropora coral branching & foliose |
| COR_CEMS | non-Acropora coral encrusting, massive & sub-massive |
| F_AGA | Agariciidae encrusting, foliose & submassive |
| F_DEN | Dendrophyllidae encrusting, foliose & massive |
| F_EUPH | Euphyllidae |
| F_FAV_CEMS | Faviidae encrusting, massive & sub-massive |
| F_FUN | Fungiidae |
| F_MER | Meruliniidae |
| F_MUS | Mussidae |
| F_OCU | Oculinidae |
| F_PEC | Pectiniidae |
| F_SID | Siderastreidae |
| G_AST | Astreopora |
| GE_ECH | Echinopora |
| G_GON_ALV | Goniopora & Alveopora |
| G_ISO | Isopora encrusting & sub-massive |
| G_MON | Montipora encrusting, foliose, massive & sub-massive |
| G_POC | Pocillopora |
| G_POR_B | Porites branching |
| G_POR_CEMS | Porites rus, Porites encrusting, massive & sub-massive |
| G_SER | Seriatopora |
| G_STY | Stylophora |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mellin, C.; Thompson, A.; Jonker, M.J.; Emslie, M.J. Cross-Shelf Variation in Coral Community Response to Disturbance on the Great Barrier Reef. Diversity 2019, 11, 38. https://doi.org/10.3390/d11030038
Mellin C, Thompson A, Jonker MJ, Emslie MJ. Cross-Shelf Variation in Coral Community Response to Disturbance on the Great Barrier Reef. Diversity. 2019; 11(3):38. https://doi.org/10.3390/d11030038
Chicago/Turabian StyleMellin, Camille, Angus Thompson, Michelle J. Jonker, and Michael J. Emslie. 2019. "Cross-Shelf Variation in Coral Community Response to Disturbance on the Great Barrier Reef" Diversity 11, no. 3: 38. https://doi.org/10.3390/d11030038
APA StyleMellin, C., Thompson, A., Jonker, M. J., & Emslie, M. J. (2019). Cross-Shelf Variation in Coral Community Response to Disturbance on the Great Barrier Reef. Diversity, 11(3), 38. https://doi.org/10.3390/d11030038