Cross-Shelf Differences in the Response of Herbivorous Fish Assemblages to Severe Environmental Disturbances


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
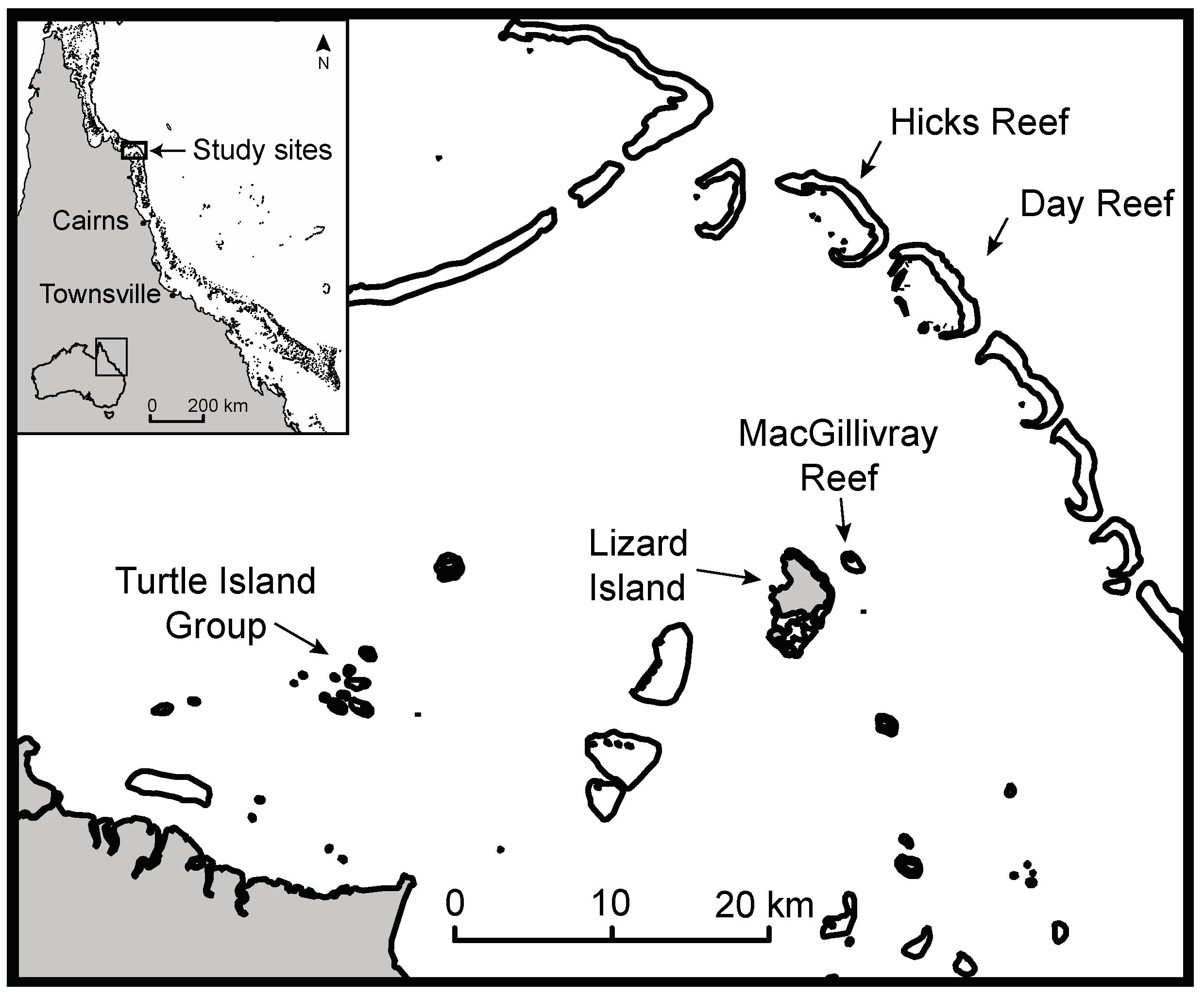
2.1. Study Area
2.2. Assessment of Benthic and Fish Assemblages
2.3. Statistical Analyses
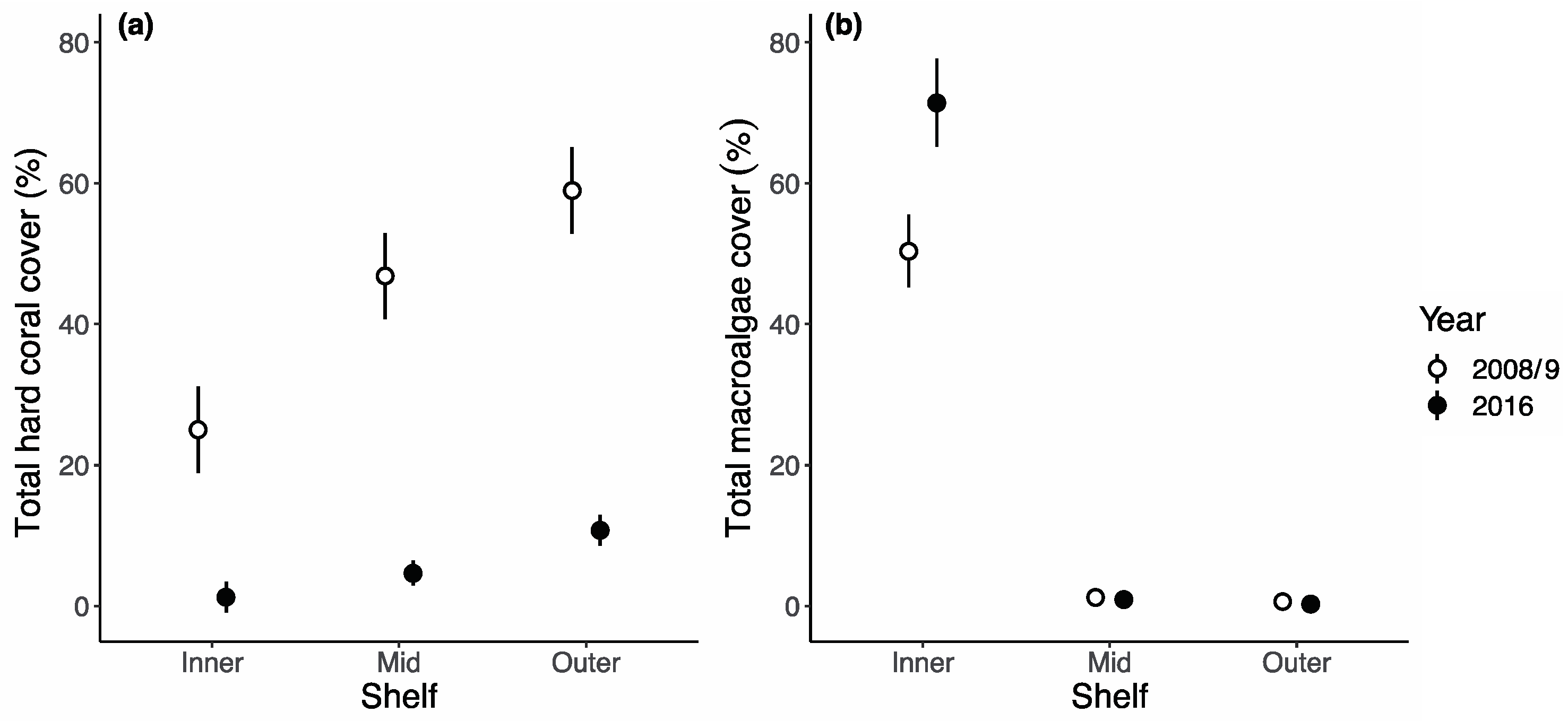
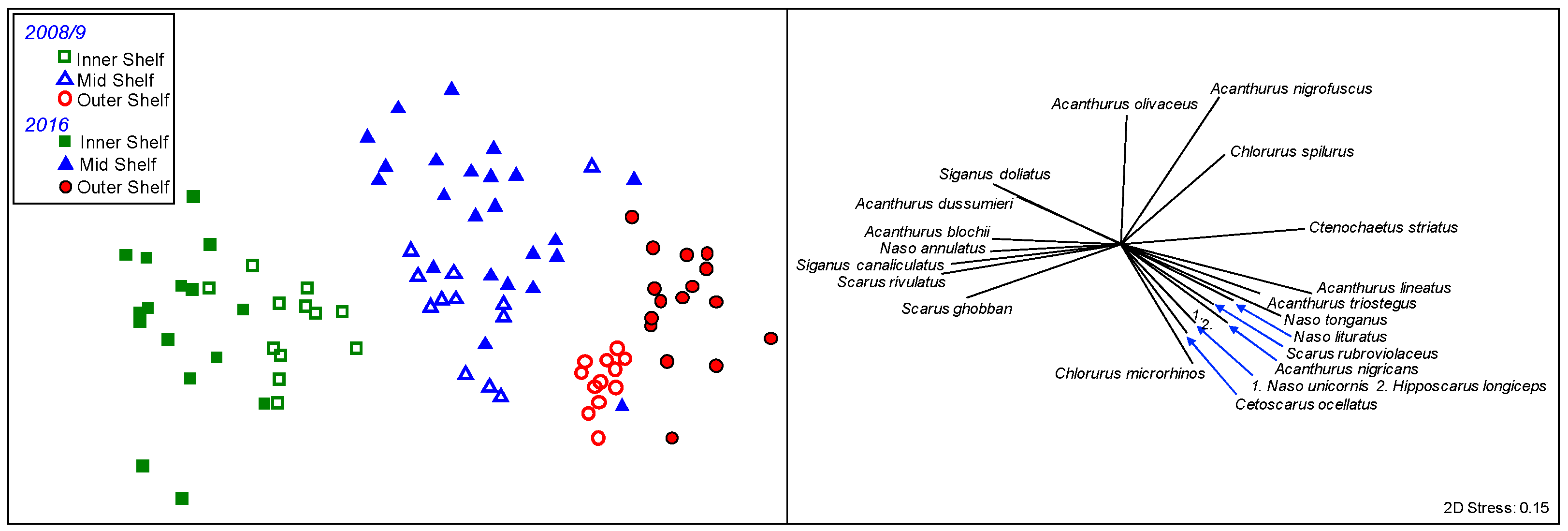
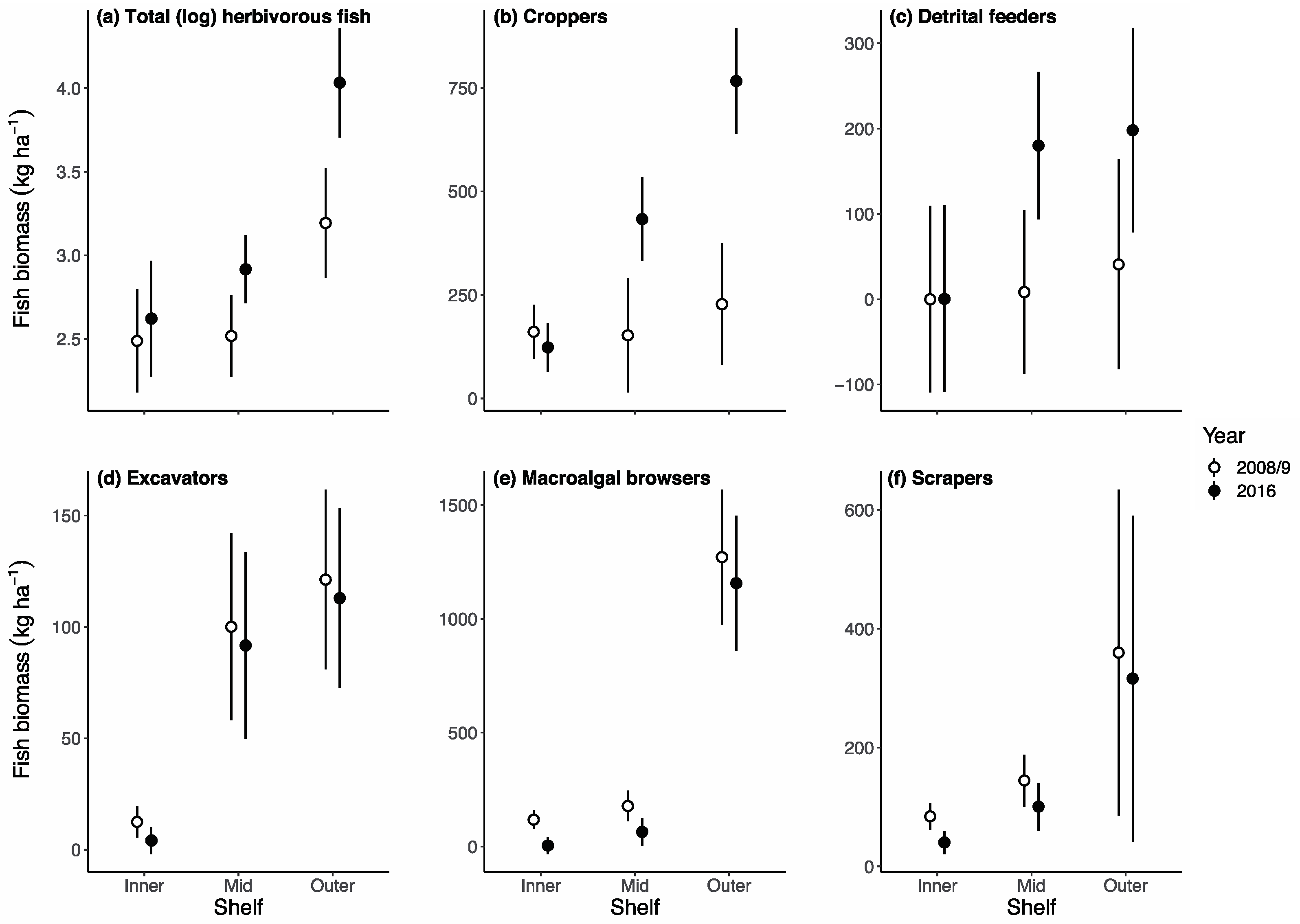
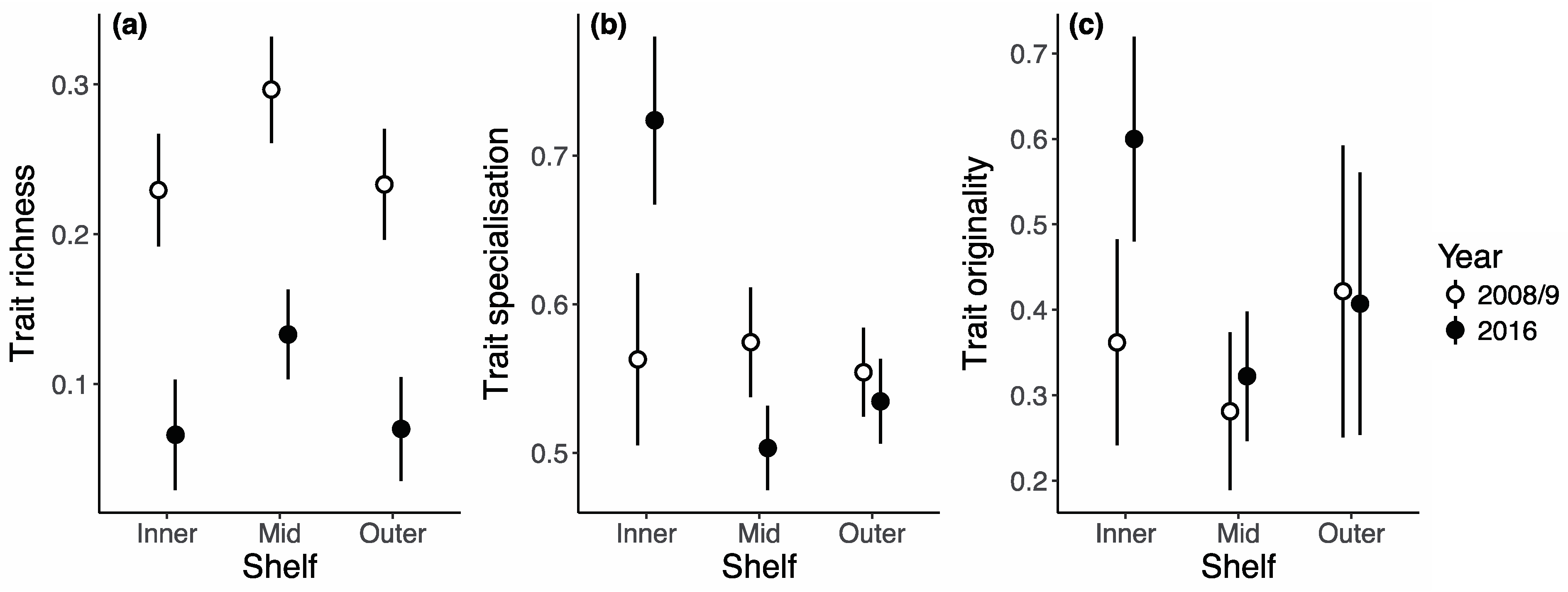
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blake, J.G.; Loiselle, B.A. Diversity of birds along an elevational gradient in the Cordillera Central, Costa Rica. Auk 2000, 117, 663–686. [Google Scholar] [CrossRef]
- Fu, C.; Hua, X.; Li, J.; Chang, Z.; Pu, Z.; Chen, J. Elevation patterns of frog species richness and endemic richness in Hengduan Mountains, China: Geometric constrains, area and climate effects. Ecography 2006, 29, 919–927. [Google Scholar] [CrossRef]
- Di Musciano, M.; Carranza, M.L.; Frate, L.; Di Cecco, V.; Di Martino, L.; Frattaroli, A.R.; Stanisci, A. Distribution of plant species and dispersal traits along environmental gradients in central Mediterranean summits. Diversity 2018, 10, 58. [Google Scholar] [CrossRef]
- Thiel, R.; Sepulveda, A.; Kafemann, R.; Nellen, W. Environmental factors as forces structuring the fish community of the Elbe Estuary. J. Fish Biol. 1995, 46, 47–69. [Google Scholar] [CrossRef]
- Wilkinson, C.R.; Cheshire, A.C. Cross-shelf variations in coral reef structure and function—influences of land and ocean. In Proceedings of the 6th international Coral Reef Symposium, Townsville, Australia, 8–12 August 1988. [Google Scholar]
- Fabricius, K.; De’ath, G.; McCook, L.; Turak, E.; Williams, D.M.B. Changes in algal, coral and fish assemblages along water quality gradients on the inshore Great Barrier Reef. Mar. Pollut. Bull. 2005, 51, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Done, T.J. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Williams, D.M. Patterns and processes in the distribution of coral reef fishes. In Ecology of Coral Reef Fishes; Sale, P., Ed.; Academic Press: San Diego, CA, USA, 1991; pp. 437–474. [Google Scholar]
- Neves, L.M.; Teixeira-Neves, T.P.; Pereira-Filho, G.H.; Araújo, F.G. The farther the better: Effects of multiple environmental variables on reef fish assemblages along a distance gradient from river influences. PLoS ONE 2016, 11, e0166679. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K. Water quality as a regional driver of coral biodiversity and macroalgae on the Great Barrier Reef. Ecol. Appl. 2010, 20, 840–850. [Google Scholar] [CrossRef]
- Balmford, A.; Bruner, A.; Cooper, P.; Costanza, R.; Farber, S.; Green, R.E.; Jenkins, M.; Jefferiss, P.; Jessamy, V.; Madden, J.; et al. Economic reasons for conserving wild nature. Science 2002, 297, 950–953. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; MacE, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Pianka, E.R. Evolutionary Ecology; Harper & Row: New York, NY, USA, 1974. [Google Scholar]
- Williams, D.M.; Hatcher, A.I. Structure of Fish Communities on Outer Slopes of Inshore, Mid-Shelf and Outer Shelf Reefs of the Great Barrier Reef. Mar. Ecol. Prog. Ser. 1983, 10, 239–250. [Google Scholar] [CrossRef]
- Williams, D.M.; Russ, G.G.; Doherty, P.P. Reef fishes: Large-scale distributions, trophic interactions and life cycles. Oceanus 1986, 29, 76–82. [Google Scholar]
- Mapstone, B.D.; Ayling, A.M.; Choat, J.H. Habitat, Cross Shelf and Regional Patterns in the Distributions and Abundances of Some Coral Reef Organisms on the Northern Great Barrier Reef—With Comment on the Implications for Future Monitoring; Great Barrier Reef Marine Park Authority: Townsville, Australia, 1998; p. 71. Available online: https://core.ac.uk/download/pdf/50850531.pdf (accessed on 11 November 2018).
- Cheal, A.; Emslie, M.; Miller, I.; Sweatman, H. The distribution of herbivorous fishes on the Great Barrier Reef. Mar. Biol. 2012, 159, 1143–1154. [Google Scholar] [CrossRef]
- Johnson, G.; Taylor, B.; Robbins, W.; Franklin, E.; Toonen, R.; Bowen, B.; Choat, J. Diversity and Structure of Parrotfish Assemblages across the Northern Great Barrier Reef. Diversity 2019, 11, 14. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Wainwright, P. Locomotion in labrid fishes: Implications for habitat use and cross-shelf biogeography on the Great Barrier Reef. Coral Reefs 2001, 20, 139–150. [Google Scholar] [CrossRef]
- Wismer, S.; Hoey, A.S.; Bellwood, D.R. Cross-shelf benthic community structure on the Great Barrier Reef: Relationships between macroalgal cover and herbivore biomass. Mar. Ecol. Prog. Ser. 2009, 376, 45–54. [Google Scholar] [CrossRef]
- Emslie, M.J.; Pratchett, M.S.; Cheal, A.J.; Osborne, K. Great Barrier Reef butterflyfish community structure: The role of shelf position and benthic community type. Coral Reefs 2010, 29, 705–715. [Google Scholar] [CrossRef]
- Cheal, A.J.; Emslie, M.; MacNeil, M.A.; Miller, I.; Sweatman, H. Spatial variation in the functional characteristics of herbivorous fish communities and the resilience of coral reefs. Ecol. Appl. 2013, 23, 174–188. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in browsing intensity on the Great Barrier Reef. Coral Reefs 2010, 29, 499–508. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.V.C. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Chang. Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K.; Messmer, V.; Graham, N.A.J. Changes in biodiversity and functioning of reef fish assemblages following coral bleaching and coral loss. Diversity 2011, 3, 424–452. [Google Scholar] [CrossRef]
- Adam, T.C.; Schmitt, R.J.; Holbrook, S.J.; Brooks, A.J.; Edmunds, P.J.; Carpenter, R.C.; Bernardi, G. Herbivory, connectivity, and ecosystem resilience: Response of a coral reef to a large-scale perturbation. PLoS ONE 2011, 6, e23717. [Google Scholar] [CrossRef] [PubMed]
- Russ, G.R.; Questel, S.L.A.; Rizzari, J.R.; Alcala, A.C. The parrotfish–coral relationship: Refuting the ubiquity of a prevailing paradigm. Mar. Biol. 2015, 162, 2029–2045. [Google Scholar] [CrossRef]
- Russ, G.R.; Payne, C.S.; Bergseth, B.J.; Rizzari, J.R.; Abesamis, R.A.; Alcala, A.C. Decadal-scale response of detritivorous surgeonfishes (family Acanthuridae) to no-take marine reserve protection and changes in benthic habitat. J. Fish Biol. 2018, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hoey, A.S.; Ackerman, J.L.; Depczynski, M. Coral bleaching, reef fish community phase shifts and the resilience of coral reefs. Glob. Chang. Biol. 2006, 12, 1587–1594. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Baird, A.H.; Depczynski, M.; González-Cabello, A.; Hoey, A.S.; Lefèvre, C.D.; Tanner, J.K. Coral recovery may not herald the return of fishes on damaged coral reefs. Oecologia 2012, 170, 567–573. [Google Scholar] [CrossRef]
- Hoey, A.S.; Howells, E.; Johansen, J.L.; Hobbs, J.P.A.; Messmer, V.; McCowan, D.M.; Wilson, S.K.; Pratchett, M.S. Recent advances in understanding the effects of climate change on coral reefs. Diversity 2016, 8, 12. [Google Scholar] [CrossRef]
- Mumby, P.J.; Dahlgren, C.P.; Harborne, A.R.; Kappel, C.V.; Micheli, F.; Brumbaugh, D.R.; Holmes, K.E.; Mendes, J.M.; Broad, K.; Sanchirico, J.N.; et al. Fishing, trophic cascades, and the process of grazing on coral reefs. Science 2006, 311, 98–101. [Google Scholar] [CrossRef]
- Russ, G. Distribution and abundance of herbivorous grazing fishes in the central Great Barrier Reef. I. Levels of variability across the entire continental shelf. Mar. Ecol. Prog. Ser. 1984, 20, 23–34. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Hoey, A.S.; Brandl, S.J.; Bellwood, D.R. Diet and cross-shelf distribution of rabbitfishes (f. Siganidae) on the northern Great Barrier Reef: Implications for ecosystem function. Coral Reefs 2013, 32, 973–984. [Google Scholar] [CrossRef]
- Emslie, M.J.; Logan, M.; Ceccarelli, D.M.; Cheal, A.J.; Hoey, A.S.; Miller, I.; Sweatman, H.P.A. Regional-scale variation in the distribution and abundance of farming damselfishes on Australia’s Great Barrier Reef. Mar. Biol. 2012, 159, 1293–1304. [Google Scholar] [CrossRef]
- Green, A.L.; Bellwood, D.R. Monitoring Functional Groups of Herbivorous Reef Fishes as Indicators of Coral Reef Resilience: A Practical Guide for Coral Reef Managers in the Asia Pacific Region (No.7); IUCN working group on Climate Change and Coral Reefs; IUCN: Gland, Switzerland, 2009. [Google Scholar]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term ‘function’ in ecology: A coral reef perspective. Funct. Ecol. 2019. [Google Scholar] [CrossRef]
- Streit, R.P.; Hoey, A.S.; Bellwood, D.R. Feeding characteristics reveal functional distinctions among browsing herbivorous fishes on coral reefs. Coral Reefs 2015, 34, 1037–1047. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bonaldo, R.R. Biology of Parrotfishes; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781482224016. [Google Scholar]
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.; Choat, J.H. Integrating ecological roles and trophic diversification on coral reefs: Multiple lines of evidence identify parrotfishes as microphages. Biol. J. Linn. Soc. 2017, 120, 729–751. [Google Scholar] [CrossRef]
- Hatcher, B.G. Coral reef primary productivity: A beggar’s banquet. Trends Ecol. Evol. 1988, 3, 106–111. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.J.; Villéger, S.; Mason, N.W.H.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Gordon, T.A.C.; Harding, H.R.; Wong, K.E.; Merchant, N.D.; Meekan, M.G.; McCormick, M.I.; Radford, A.N.; Simpson, S.D. Habitat degradation negatively affects auditory settlement behavior of coral reef fishes. Proc. Natl. Acad. Sci. USA 2018, 115, 5193–5198. [Google Scholar] [CrossRef]
- Hoey, A.S.; Pratchett, M.S.; Cvitanovic, C. High macroalgal cover and low coral recruitment undermines the potential resilience of the world’s southernmost coral reef assemblages. PLoS ONE 2011, 6, e25824. [Google Scholar] [CrossRef] [PubMed]
- Dickens, L.C.; Goatley, C.H.R.; Tanner, J.K.; Bellwood, D.R. Quantifying relative diver effects in underwater visual censuses. PLoS ONE 2011, 6, e18965. [Google Scholar] [CrossRef]
- Kulbicki, M.; Guillemot, N.; Amand, M. A general approach to length-weight relationships for New Caledonian lagoon fishes. Cybium 2005, 29, 235–252. [Google Scholar]
- Choat, J.H.; Clements, K.D.; Robbins, W.D. The trophic status of herbivorous fishes on coral reefs 1: Dietary analyses. Mar. Biol. 2002, 140, 613–623. [Google Scholar]
- Froese, R.; Pauly, D. Fishbase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 11 November 2018).
- Kiørboe, T.; Visser, A.; Andersen, K.H. A trait-based approach to ocean ecology. ICES J. Mar. Sci. 2018, 75, 1849–1863. [Google Scholar] [CrossRef]
- Clarke, K.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E, Ltd. Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology: Second English edition. Developments in Environmental Modelling, 20th ed.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Maire, E.; Grenouillet, G.; Brosse, S.; Villéger, S. How many dimensions are needed to accurately assess functional diversity? A pragmatic approach for assessing the quality of functional spaces. Glob. Ecol. Biogeogr. 2015, 24, 728–740. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Plymouth Marine Laboratory: Plymouth, UK, 2008; ISBN 0-12-044620-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; ISBN 3900051070. [Google Scholar]
- Andrews, J.; Gentien, P. Upwelling as a source of nutrients for the Great Barrier Reef Ecosystems: A solution to Darwin’s question? Mar. Ecol. Prog. Ser. 1982, 8, 257–269. [Google Scholar] [CrossRef]
- Richardson, L.E.; Graham, N.A.J.; Pratchett, M.S.; Eurich, J.G.; Hoey, A.S. Mass coral bleaching causes biotic homogenization of reef fish assemblages. Glob. Chang. Biol. 2018, 24, 3117–3129. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K. Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Curr. Opin. Environ. Sustain. 2014, 7, 37–43. [Google Scholar] [CrossRef]
- Hamilton, R.J.; Almany, G.R.; Brown, C.J.; Pita, J.; Peterson, N.A.; Howard Choat, J. Logging degrades nursery habitat for an iconic coral reef fish. Biol. Conserv. 2017, 210, 273–280. [Google Scholar] [CrossRef]
- Hart, A.M.; Russ, G.R. Response of herbivorous fishes to crown-of-thorns starfish Acanthaster planci outbreaks. III. Age, growth, mortality and maturity indices of Acanthurus nigrofuscus. Mar. Ecol. Prog. Ser. 1996, 136, 25–35. [Google Scholar] [CrossRef]
- Gilmour, J.P.; Smith, L.D.; Heyward, A.J.; Baird, A.H.; Pratchett, M.S. Recovery of an isolated coral reef system following severe disturbance. Science 2013, 340, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Lamy, T.; Legendre, P.; Chancerelle, Y.; Siu, G.; Claudet, J. Understanding the spatio-temporal response of coral reef fish communities to natural disturbances: Insights from beta-diversity decomposition. PLoS ONE 2015, 10, e0138696. [Google Scholar] [CrossRef] [PubMed]
- Rizzari, J.R.; Bergseth, B.J.; Frisch, A.J. Impact of conservation areas on trophic interactions between apex predators and herbivores on coral reefs. Conserv. Biol. 2015, 29, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime Shifts, Resilience, and Biodiversity in Ecosystem Management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Engelen, A.H.; Breeman, A.M.; Olsen, J.L.; Stam, W.T.; Åberg, P. Life history flexibility allows Sargassum polyceratium to persist in different environments subjected to stochastic disturbance events. Coral Reefs 2005, 24, 670–680. [Google Scholar] [CrossRef]
- Loffler, Z.; Graba-Landry, A.; Kidgell, J.T.; McClure, E.C.; Pratchett, M.S.; Hoey, A.S. Holdfasts of Sargassum swartzii are resistant to herbivory and resilient to damage. Coral Reefs 2018, 37, 1075–1084. [Google Scholar] [CrossRef]
- Clements, C.S.; Rasher, D.B.; Hoey, A.S.; Bonito, V.E.; Hay, M.E. Spatial and temporal limits of coral-macroalgal competition: The negative impacts of macroalgal density, proximity, and history of contact. Mar. Ecol. Prog. Ser. 2018, 586, 11–20. [Google Scholar] [CrossRef]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase Shifts, Herbivory, and the Resilience of Coral Reefs to Climate Change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef]
- Newman, S.J.; Williams, D.M.B. Variation in reef associated assemblages of the Lutjanidae and Lethrinidae at different distances offshore in the central Great Barrier Reef. Environ. Biol. Fishes 1996, 46, 123–138. [Google Scholar] [CrossRef]
- Newman, S.J.; Williams, D.M.B.; Russ, G.R. Patterns of zonation of assemblages of the Lutjanidae, Lethrinidae and Serranidae (Epinephelinae) within and among mid-shelf and outer-shelf reefs in the central Great Barrier Reef. Mar. Freshw. Res. 1997, 48, 119–128. [Google Scholar] [CrossRef]
- Gust, N.; Choat, J.H.; Ackerman, J.L. Demographic plasticity in tropical reef fishes. Mar. Biol. 2002, 140, 1039–1051. [Google Scholar]
- Frisch, A.J.; Baker, R.; Hobbs, J.P.A.; Nankervis, L. A quantitative comparison of recreational spearfishing and linefishing on the Great Barrier Reef: Implications for management of multi-sector coral reef fisheries. Coral Reefs 2008, 27, 85–95. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McClure, E.C.; Richardson, L.E.; Graba-Landry, A.; Loffler, Z.; Russ, G.R.; Hoey, A.S. Cross-Shelf Differences in the Response of Herbivorous Fish Assemblages to Severe Environmental Disturbances. Diversity 2019, 11, 23. https://doi.org/10.3390/d11020023
McClure EC, Richardson LE, Graba-Landry A, Loffler Z, Russ GR, Hoey AS. Cross-Shelf Differences in the Response of Herbivorous Fish Assemblages to Severe Environmental Disturbances. Diversity. 2019; 11(2):23. https://doi.org/10.3390/d11020023
Chicago/Turabian StyleMcClure, Eva C., Laura E. Richardson, Alexia Graba-Landry, Zoe Loffler, Garry R. Russ, and Andrew S. Hoey. 2019. "Cross-Shelf Differences in the Response of Herbivorous Fish Assemblages to Severe Environmental Disturbances" Diversity 11, no. 2: 23. https://doi.org/10.3390/d11020023
APA StyleMcClure, E. C., Richardson, L. E., Graba-Landry, A., Loffler, Z., Russ, G. R., & Hoey, A. S. (2019). Cross-Shelf Differences in the Response of Herbivorous Fish Assemblages to Severe Environmental Disturbances. Diversity, 11(2), 23. https://doi.org/10.3390/d11020023