Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
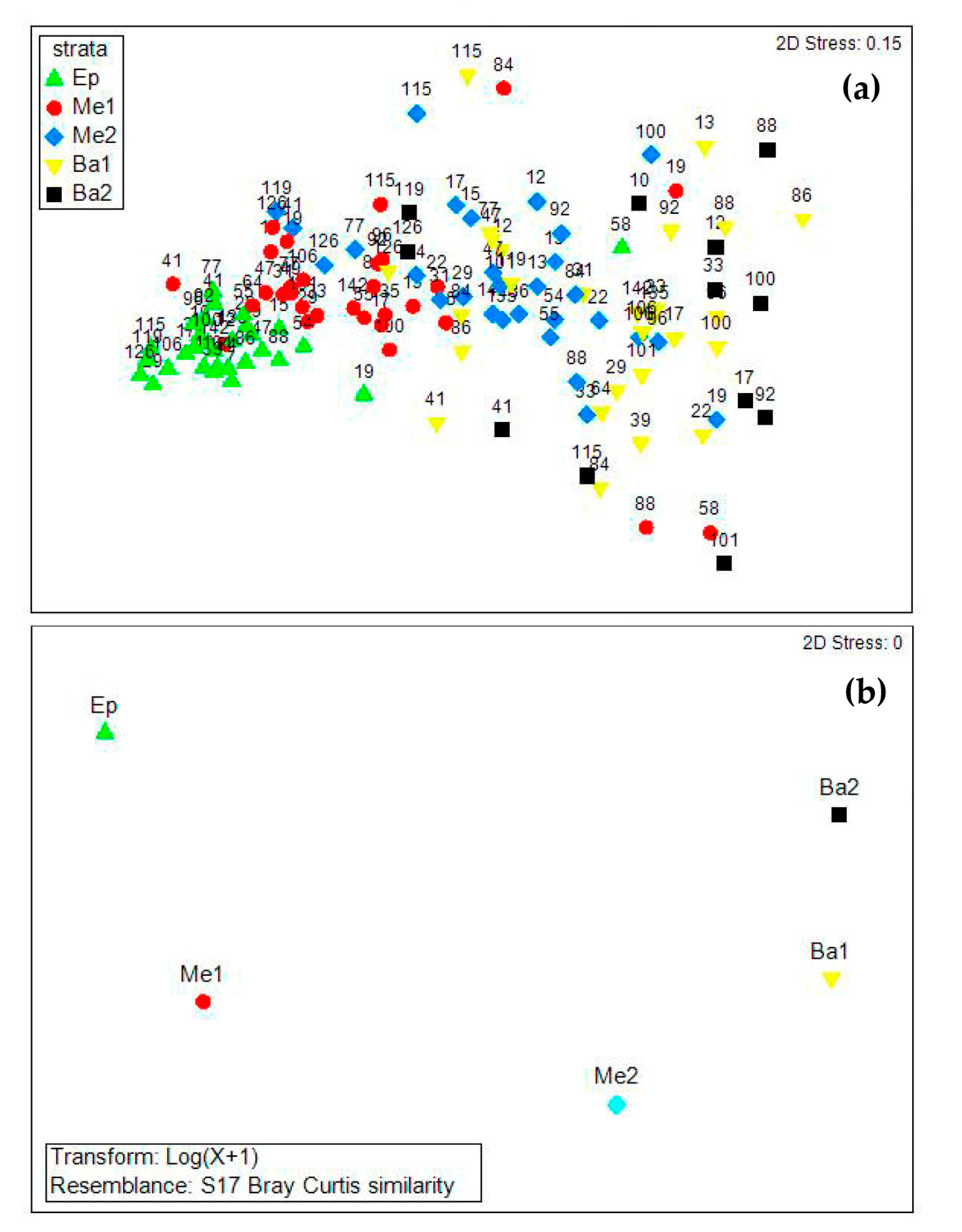
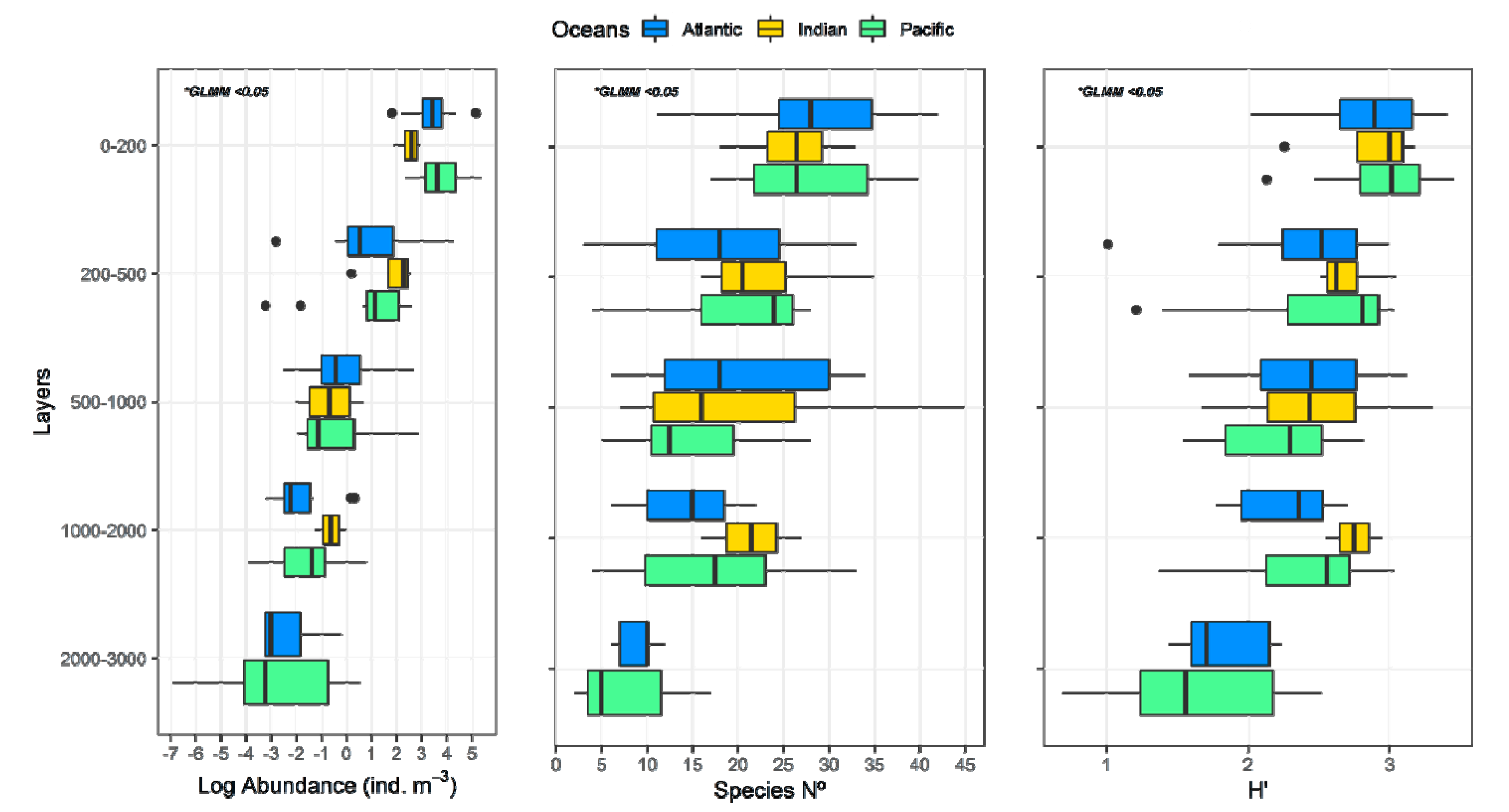
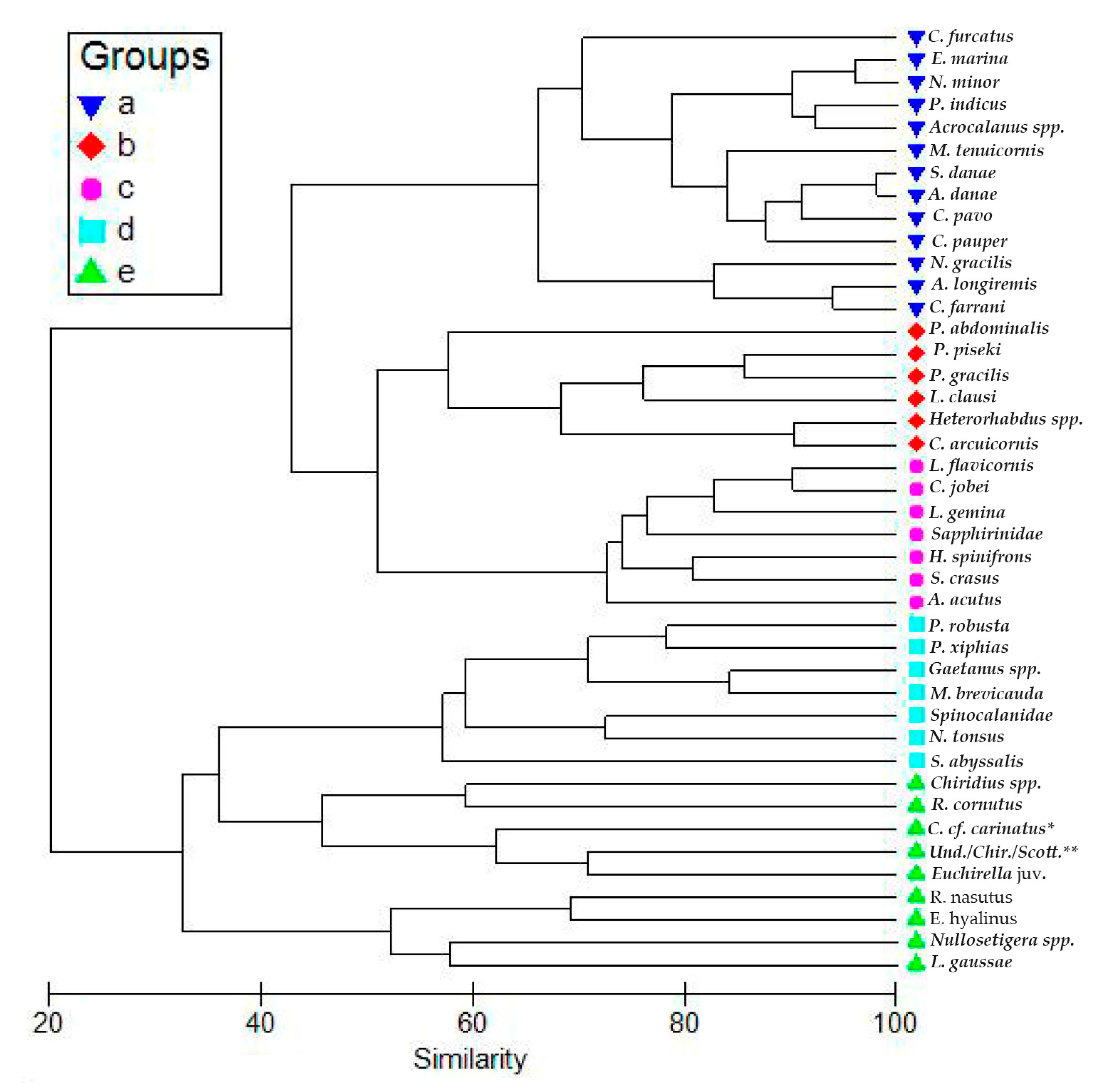
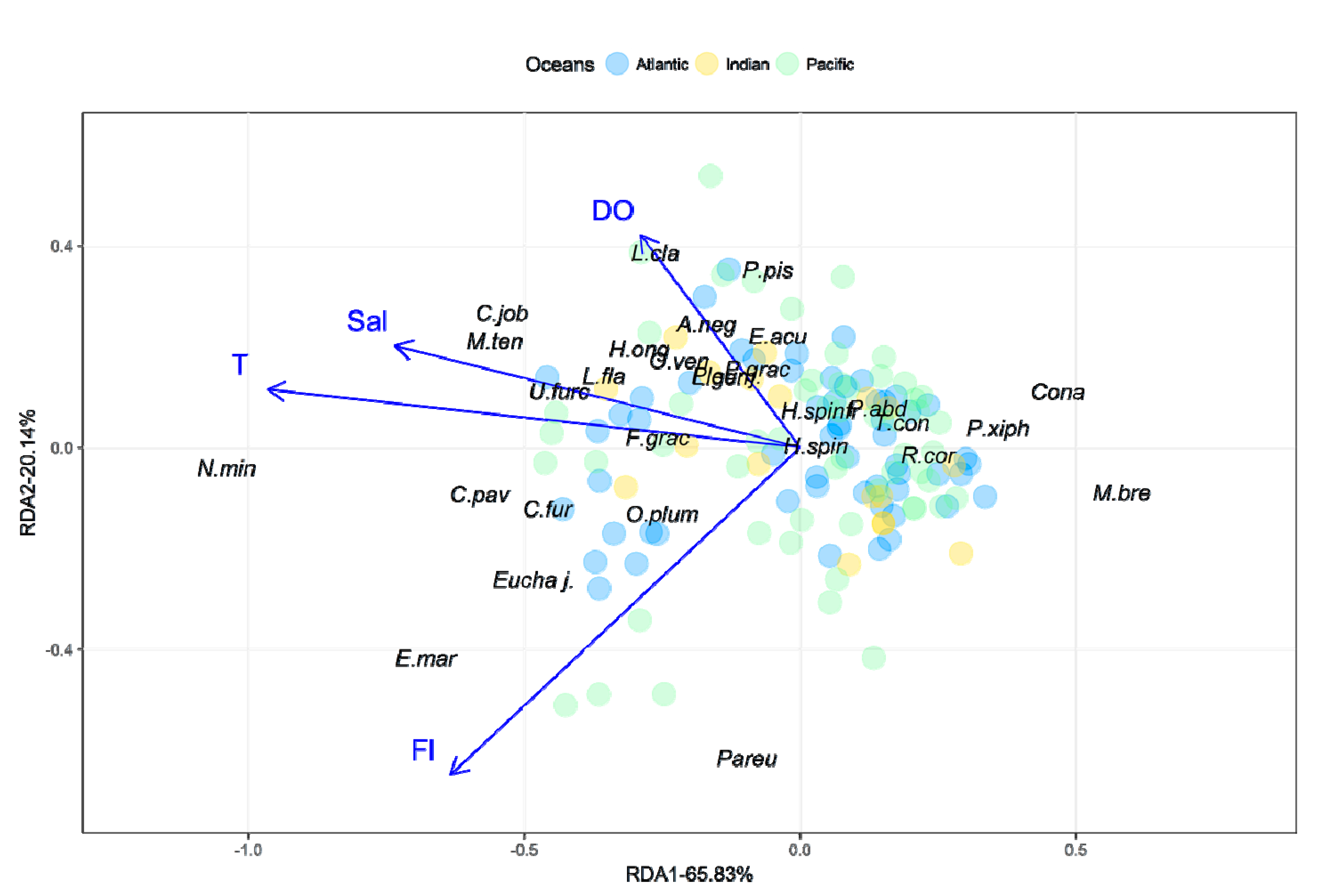
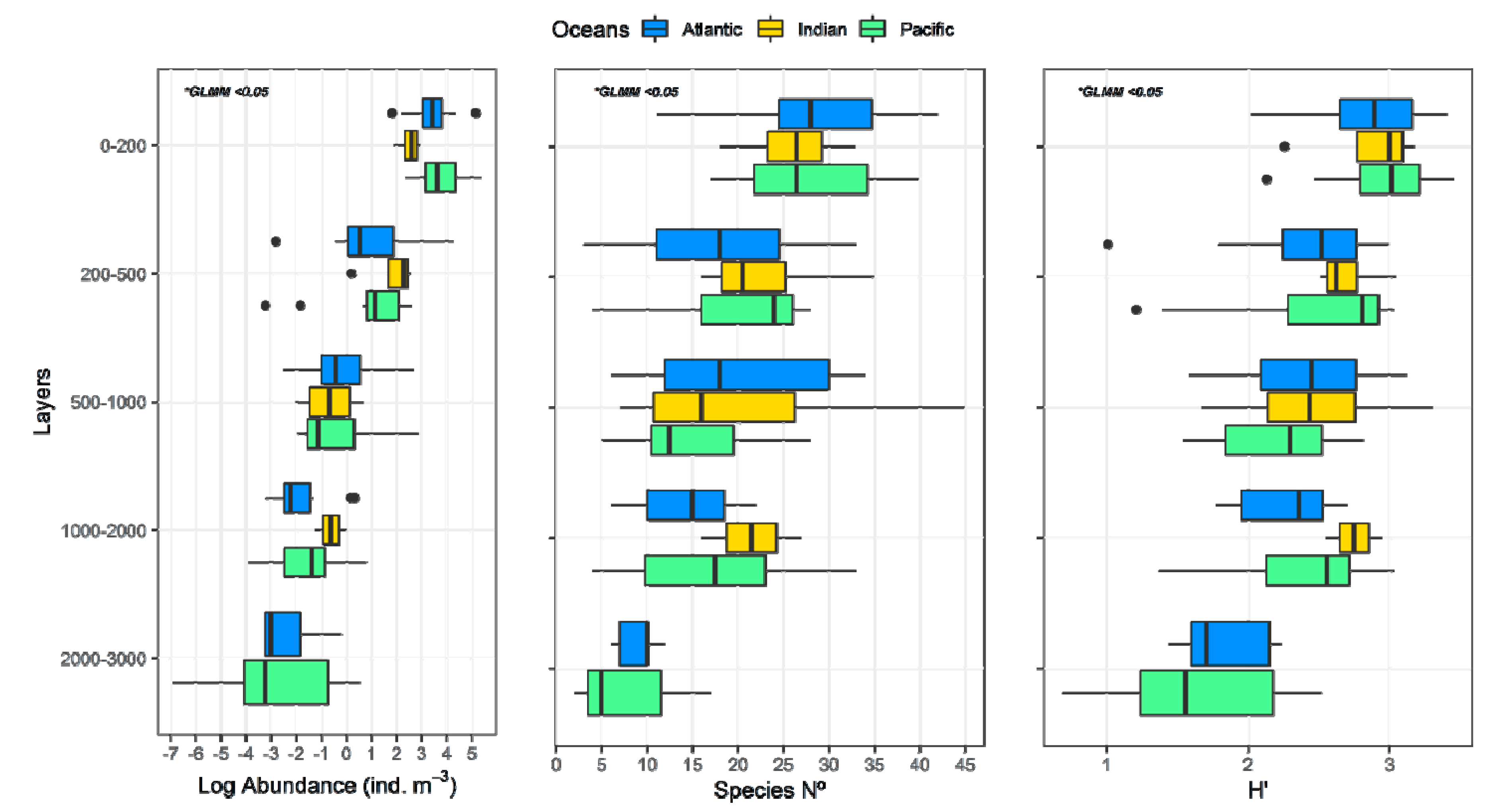
3. Results
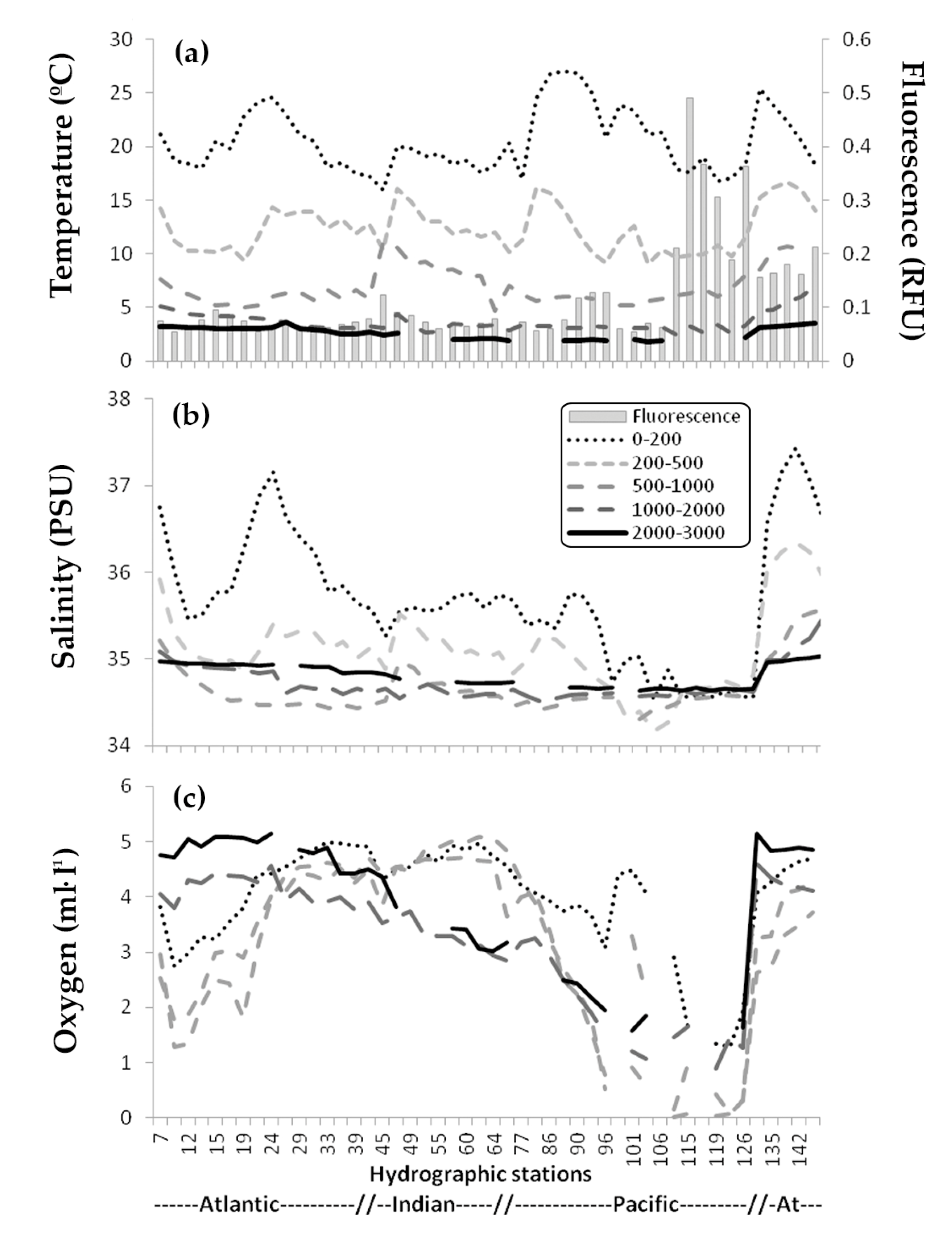
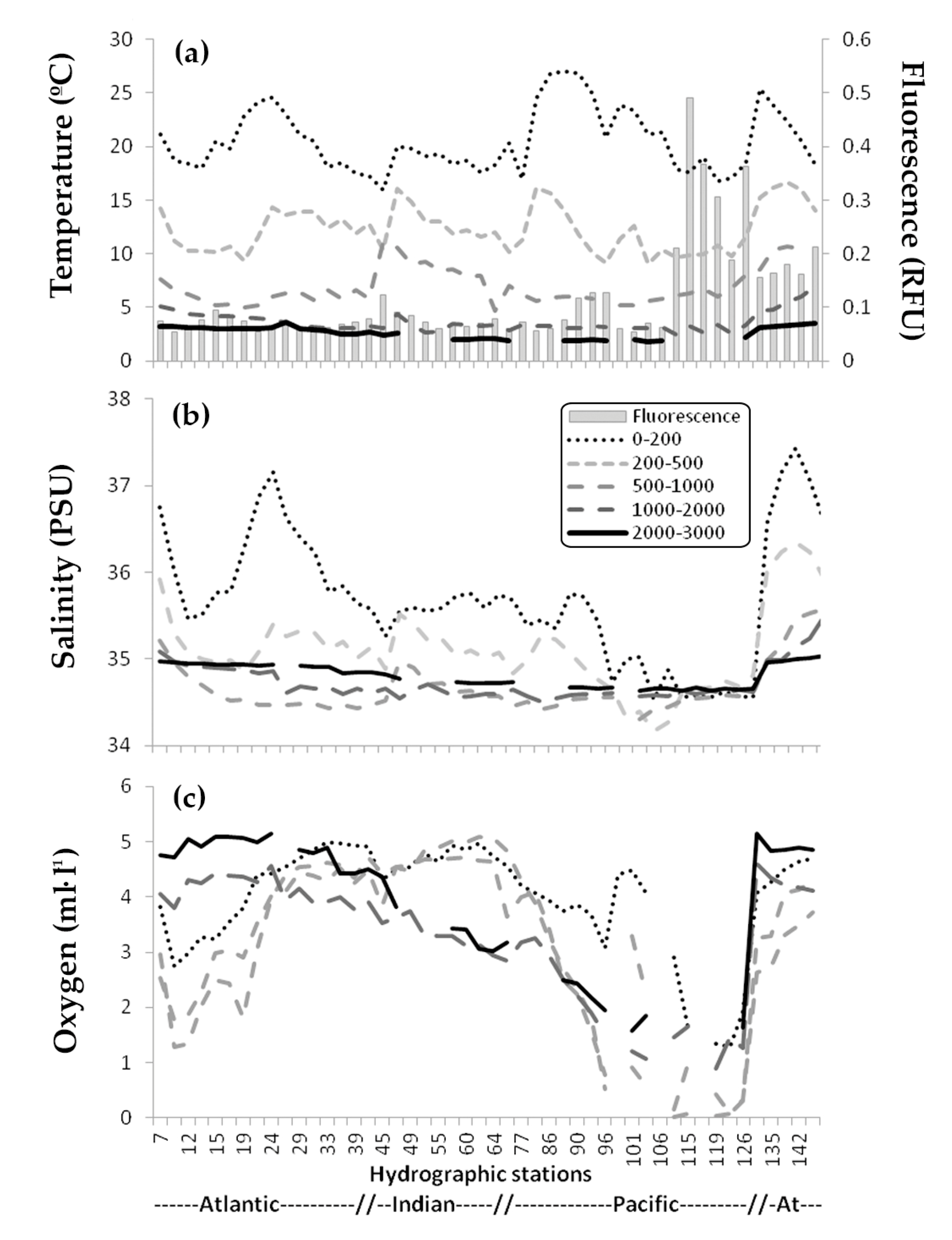
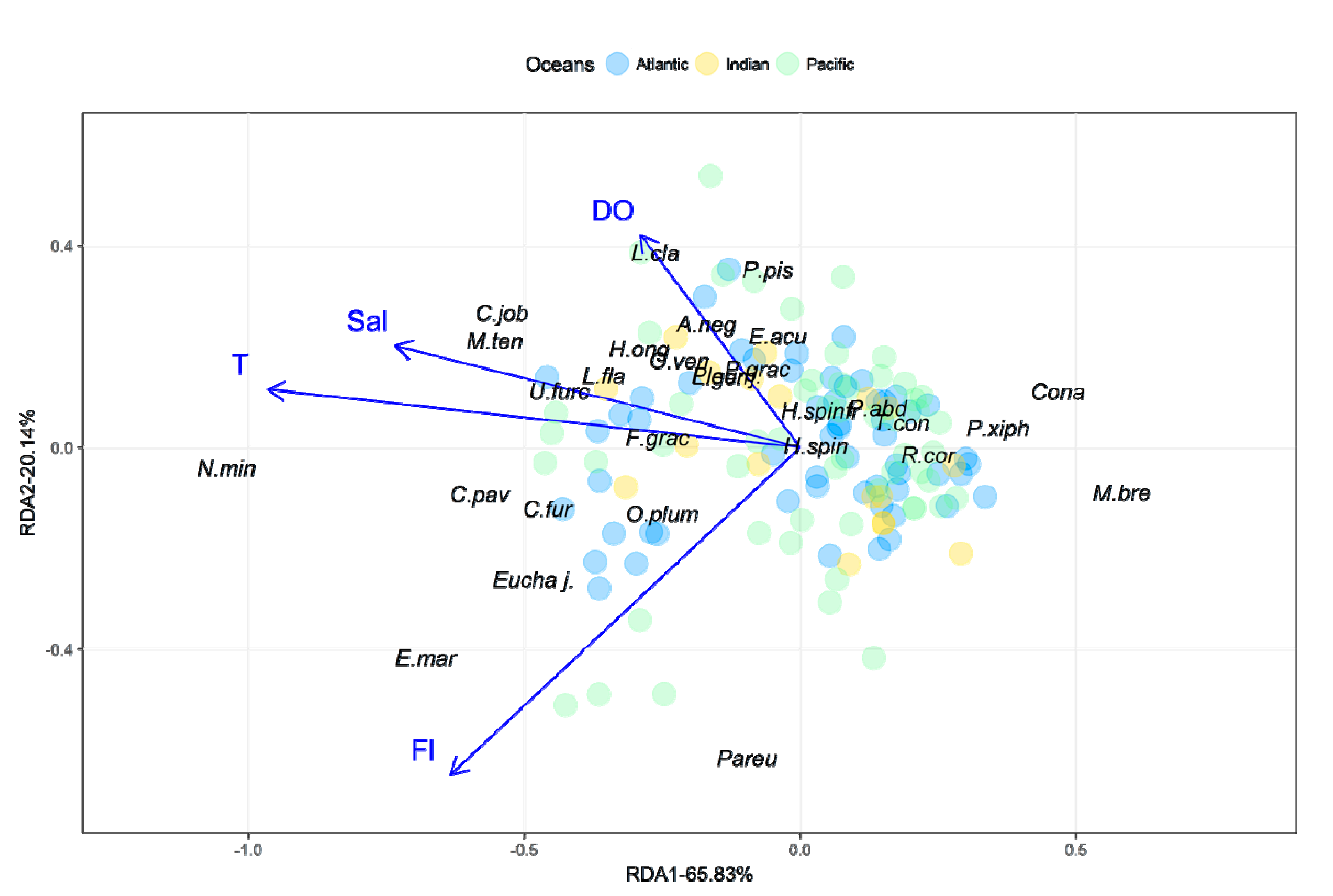
3.1. Environmental Data
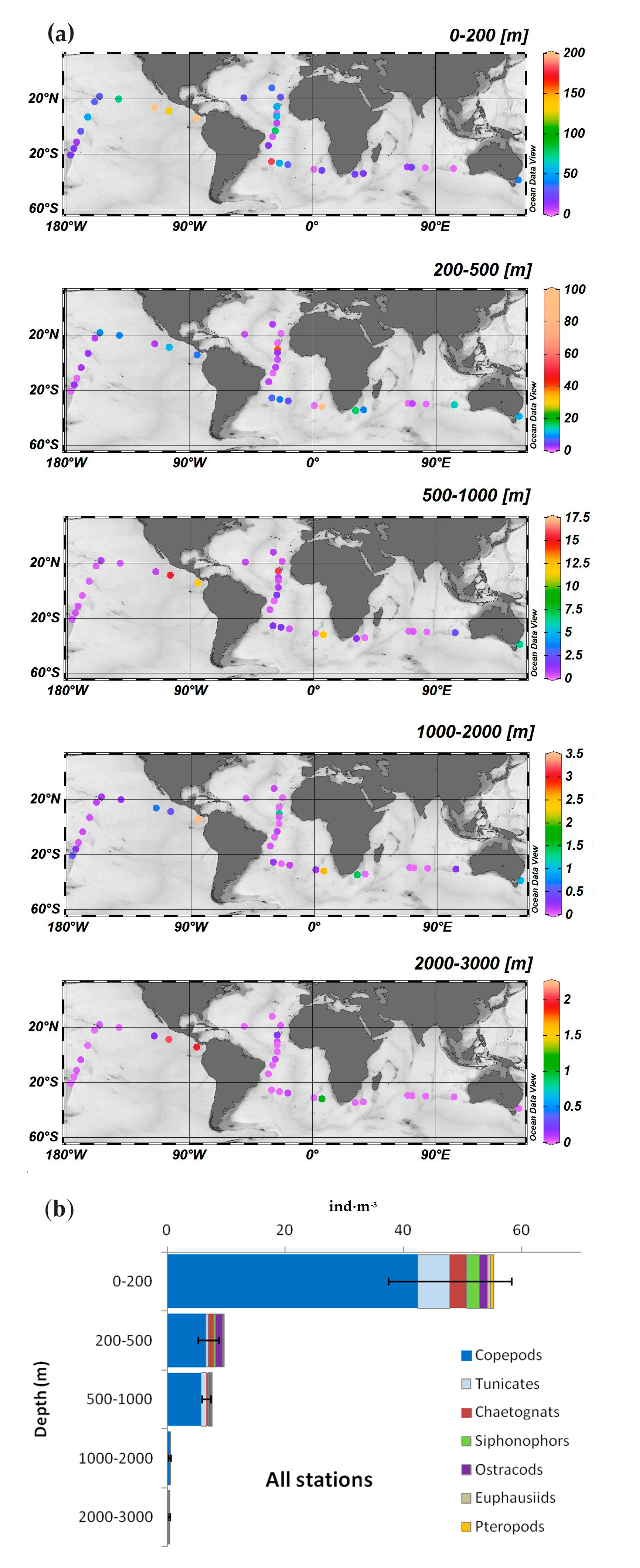
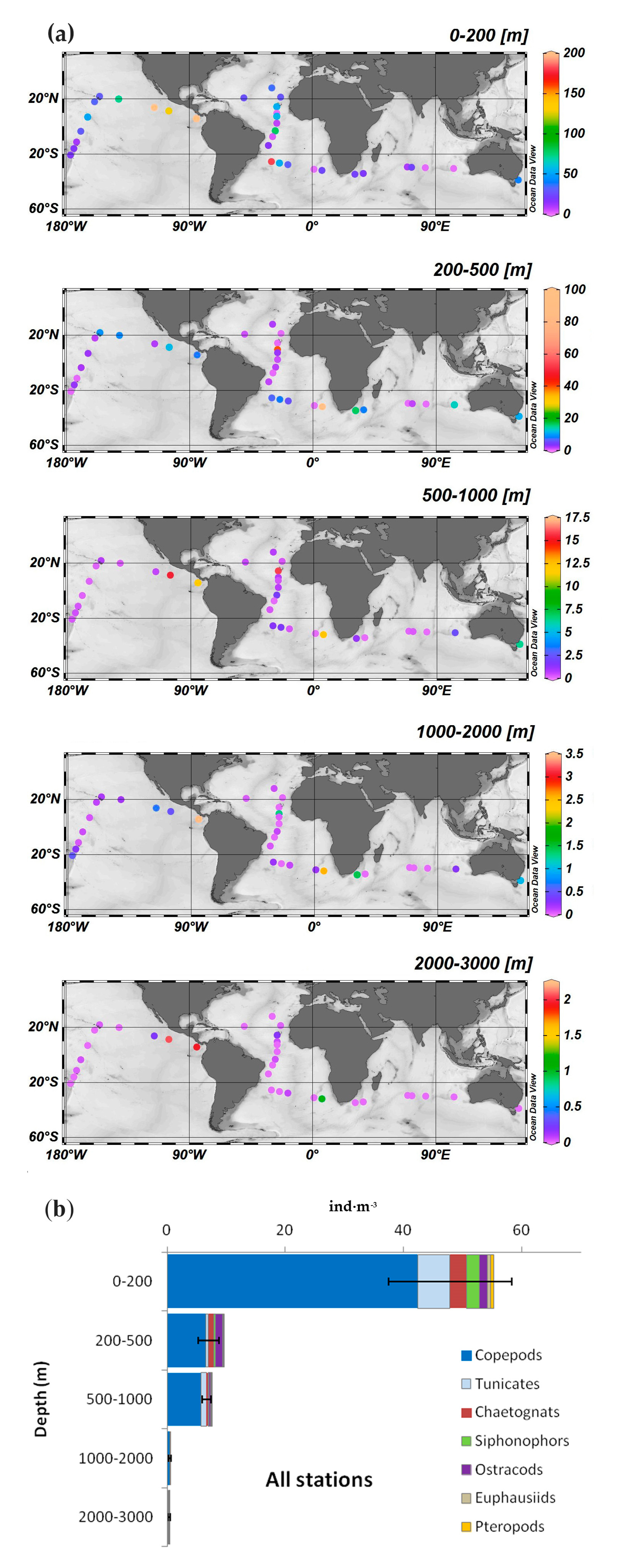
3.2. Zooplankton Abundance and Main Groups
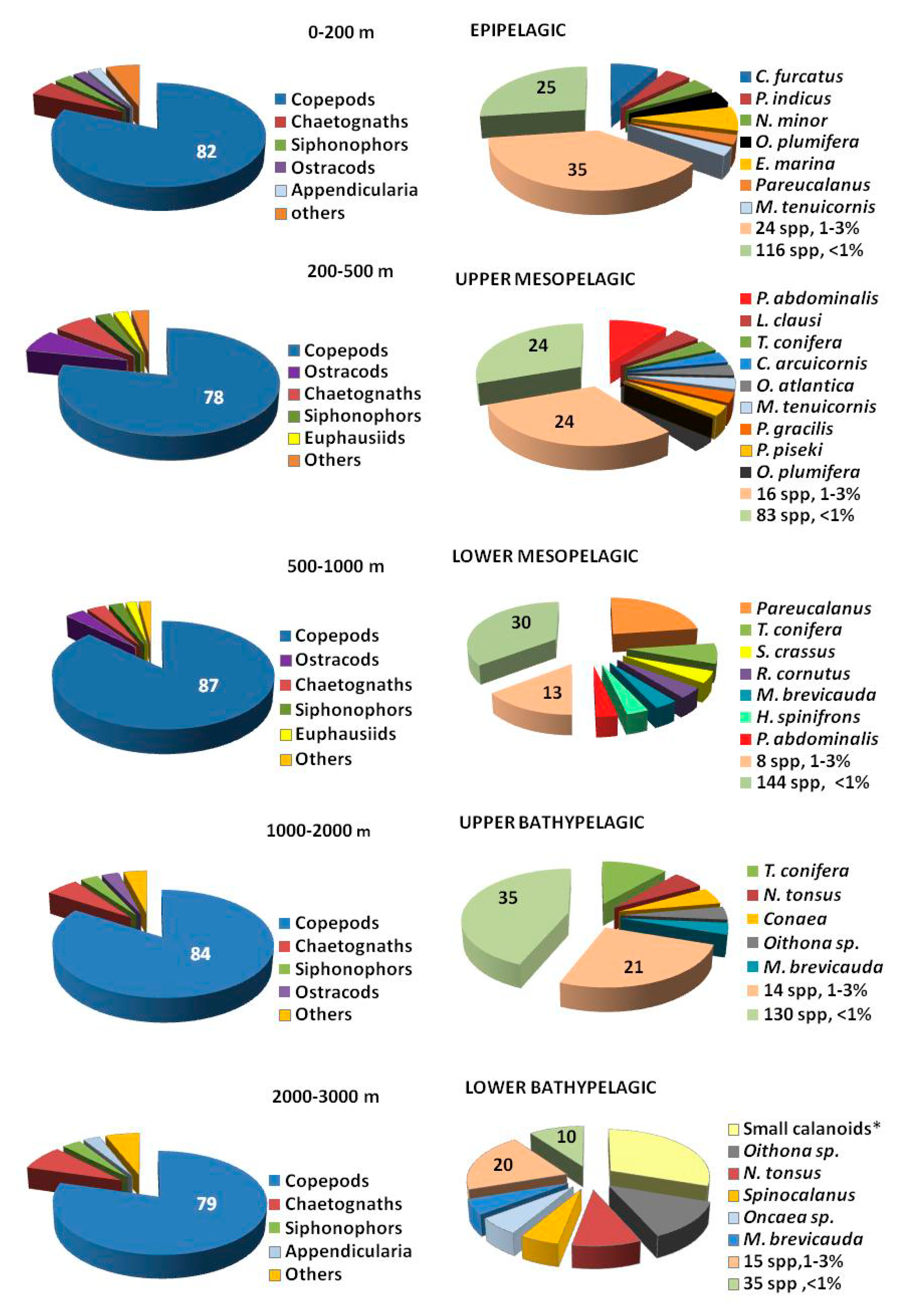
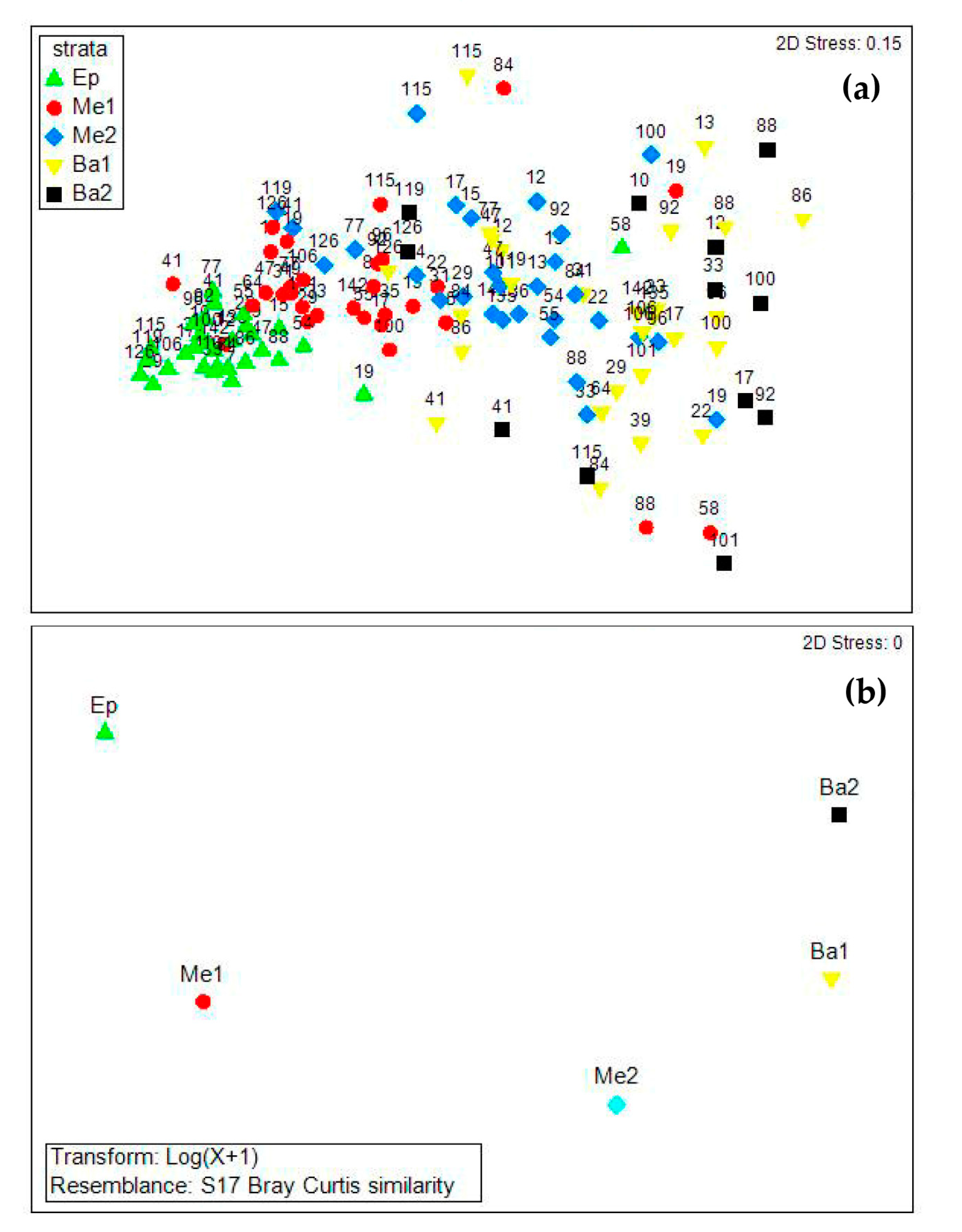
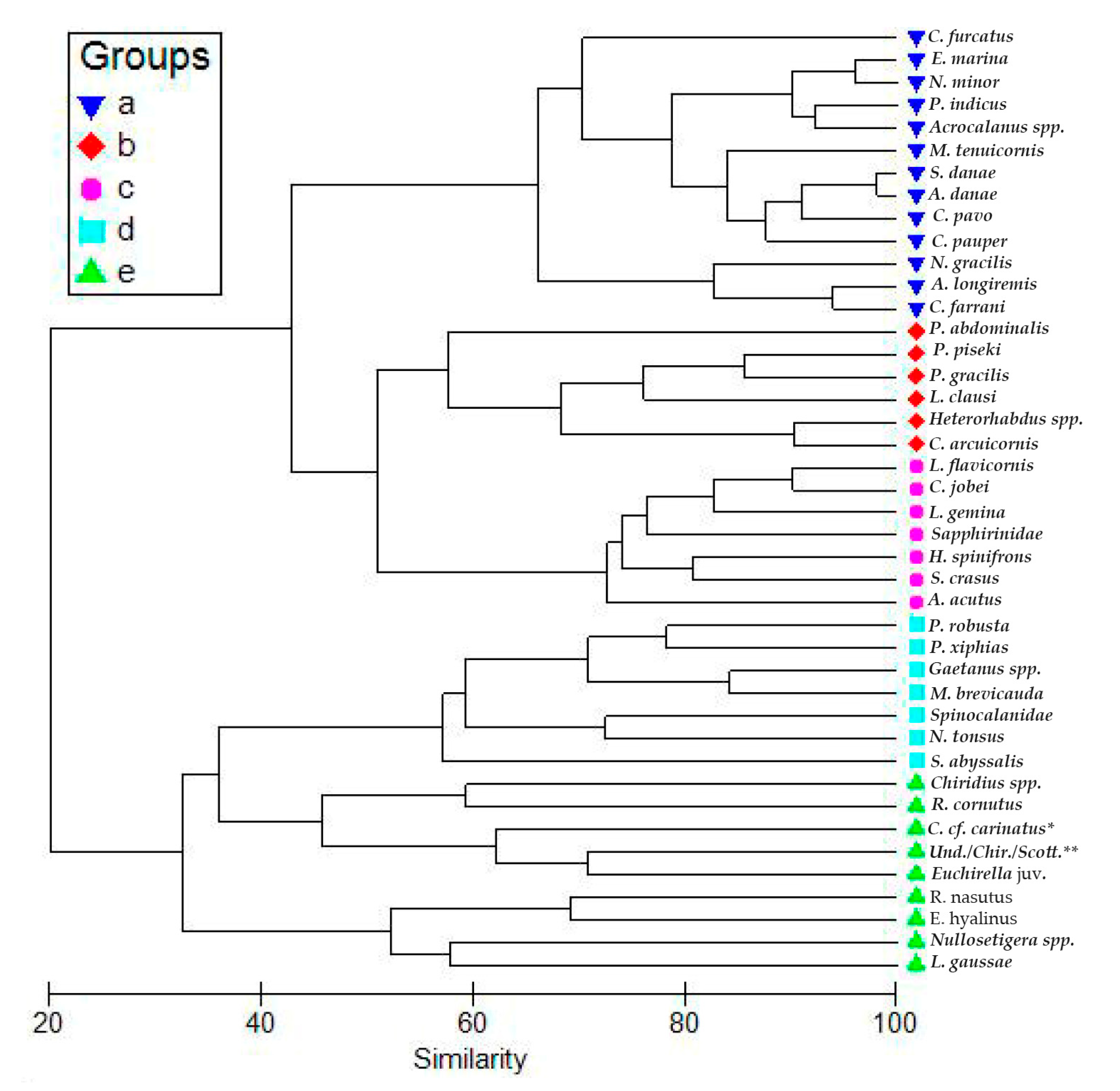
3.3. Copepod Composition, Dominant Species, and Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bucklin, A.; Nishida, S.; Schnack-Schiel, S.; Wiebe, P.H.; Lindsay, D.; Machida, R.J.; Copley, N.J. A Census of Zooplankton of the Global Ocean. In Wiley Online Books Life in the World’s Oceans; McIntyre, A.D., Ed.; Wiley-Blackwell Publisher: West Sussex, UK, 2010; pp. 247–263. [Google Scholar]
- Hering, P. The Biology of the Deep Ocean; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef]
- Longhurst, A.R. Relationship between diversity and the vertical structure of the upper ocean. Deep Sea Res. Part A Oceanogr. Res. Pap. 1985, 32, 1535–1570. [Google Scholar] [CrossRef]
- Vinogradov, M.E.; Gitelzon, I.I.; Sorokin, Y.I. The vertical structure of a pelagic community in the tropical ocean. Mar. Biol. 1970, 6, 187–194. [Google Scholar] [CrossRef]
- Beaugrand, G.; Reid, P.C.; Ibanez, F.; Lindley, J.A.; Edwards, M. Reorganization of North Atlantic marine copepod biodiversity and climate. Science 2002, 296, 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, A.R. Ecological Geography of the Sea; Burlington, Ed.; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Fiedler, P.C. The annual cycle and biological effects of the Costa Rica Dome. Deep Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 321–338. [Google Scholar] [CrossRef]
- Sameoto, D.D. Environmental factors influencing diurnal distribution of zooplankton and ichthyoplankton. J. Plankton Res. 1984, 6, 767–792. [Google Scholar] [CrossRef] [Green Version]
- Vinogradov, M.E. Some Problems of Vertical Distribution of Meso- and Macroplankton in the Ocean. In The Biogeography of the Oceans; Blaxter, J.H.S., Southward, A.J., Gebruk, A.V., Southward, E.C., Tyler, P., Eds.; Academic Press: Cambridge, MA, USA, 1997; Volume 32, pp. 1–92. ISBN 0065-2881. [Google Scholar]
- Cavalcanti, E.A.H.; de Larrazábal, M.E.L. Macrozooplâncton da Zona Econômica Exclusiva do Nordeste do Brasil (segunda expedição oceanográfica-REVIZEE/NE II) com ênfase em Copepoda (Crustacea). Rev. Bras. Zool. 2004, 21, 467–475. [Google Scholar] [CrossRef]
- Verheye, H.M.; Hagen, W.; Auel, H.; Ekau, W.; Loick, N.; Rheenen, I.; Wencke, P.; Jones, S. Life strategies, energetics and growth characteristics of Calanoides carinatus (Copepoda) in the Angola-Benguela frontal region. Afr. J. Mar. Sci. 2005, 27, 641–651. [Google Scholar] [CrossRef]
- Longhurst, A.; Williams, R. Materials for plankton modelling: Vertical distribution of Atlantic zooplankton in summer. J. Plankton Res. 1979, 1, 1–28. [Google Scholar] [CrossRef]
- Gaard, E.; Gislason, A.; Falkenhaug, T.; Søiland, H.; Musaeva, E.; Vereshchaka, A.; Vinogradov, G. Horizontal and vertical copepod distribution and abundance on the Mid-Atlantic Ridge in June 2004. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 59–71. [Google Scholar] [CrossRef]
- Schnack-Schiel, S.B.; Mizdalski, E.; Cornils, A. Copepod abundance and species composition in the Eastern subtropical/tropical Atlantic. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 2064–2075. [Google Scholar] [CrossRef]
- Vereshchaka, A.; Abyzova, G.; Lunina, A.; Musaeva, E. The Deep-Sea zooplankton of the North, Central, and South Atlantic: Biomass, abundance, diversity. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 137, 89–101. [Google Scholar] [CrossRef]
- Bode, M.; Hagen, W.; Cornils, A.; Kaiser, P.; Auel, H. Copepod distribution and biodiversity patterns from the surface to the deep sea along a latitudinal transect in the eastern Atlantic Ocean (24°N to 21°S). Prog. Oceanogr. 2018, 161, 66–77. [Google Scholar] [CrossRef]
- Grice, G.D.; Hulseman, K. Bathypelagic Calanoid Copepods of the Western Indian Ocean. Proc. United States Natl. Mus. 1967, 122, 1–67. [Google Scholar] [CrossRef] [Green Version]
- Conway, D.V.P. Guide to the Coastal and Surface Zooplankton of the South-Western Indian Ocean; Marine Biological Association of the United Kingdom: Plymouth, UK, 2003. [Google Scholar]
- Vereshchaka, A.L. Vertical distribution of euphausiids, pelagic decapods, mysids in the Near-Bottom layer of the western Indian Ocean. Okeanologiya 1990, 30, 126–131. [Google Scholar]
- Vereshchaka, A.L. Macroplankton in the Near-Bottom layer of continental slopes and seamounts. Deep Sea Res. Part I Oceanogr. Res. Pap. 1995, 42, 1639–1668. [Google Scholar] [CrossRef]
- Roemmich, D.; McGowan, J. Climatic Warming and the Decline of Zooplankton in the California Current. Science 1995, 267, 1324–1326. [Google Scholar] [CrossRef]
- McGowan, J.A.; Walker, P.W. Structure in the Copepod Community of the North Pacific Central Gyre. Ecol. Monogr. 1979, 49, 195–226. [Google Scholar] [CrossRef]
- Fernández-Álamo, M.A.; Färber-Lorda, J. Zooplankton and the oceanography of the eastern tropical Pacific: A review. Prog. Oceanogr. 2006, 69, 318–359. [Google Scholar] [CrossRef]
- Piontkovski, S.A.; Landry, M.R.; Finenko, Z.Z.; Kovalev, A.V.; Williams, R.; Gallienne, C.P.; Mishonov, A.V.; Skryabin, V.A.; Tokarev, Y.N.; Nikolsky, V.N. Plankton communities of the South Atlantic anticyclonic gyre. Oceanol. Acta 2003, 26, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Teuber, L.; Schukat, A.; Hagen, W.; Auel, H. Distribution and Ecophysiology of Calanoid Copepods in Relation to the Oxygen Minimum Zone in the Eastern Tropical Atlantic. PLoS ONE 2013, 8, e77590. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, A. Seasonal cycles of pelagic production and consumption. Prog. Oceanogr. 1995, 36, 77–167. [Google Scholar] [CrossRef]
- Angel, M.V.; de Baker, A.C. Vertical Distribution of the Standing Crop of Plankton and Micronekton at Three Stations in the Northeast Atlantic. Biol. Oceanogr. 1982, 2, 1–30. [Google Scholar]
- Van der Spoel, S.; Heyman, R.P. A Comparative Atlas of Zooplankton; Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- Weikert, H.; Koppelmann, R. Mid-Water zooplankton profiles from the temperate ocean and partially landlocked seas. A Re-Evaluation of interoceanic differences. Oceanol. Acta 1996, 19, 657–664. [Google Scholar]
- Benedetti, F.; Gasparini, S.; Ayata, S.D. Identifying copepod functional groups from species functional traits. J. Plankton Res. 2016, 38, 159–166. [Google Scholar] [CrossRef]
- Bernal, A.; Olivar, M.P.; Maynou, F.; Fernández de Puelles, M.L. Diet and feeding strategies of mesopelagic fishes in the western Mediterranean. Prog. Oceanogr. 2015, 135, 1–17. [Google Scholar] [CrossRef]
- Moku, M.; Kawaguchi, K.; Watanabe, H. Feeding habits of three dominant myctophid fishes, Diaphus theta, Stenobrachius leucopsarus and S. nannochir, in the subarctic and transitional waters of the western North Pacific. Mar. Ecol. Prog. Ser. 2000, 207, 129–140. [Google Scholar] [CrossRef]
- Gartner, J.V.; Crabtree, R.E.; Sulak, K.J. Feeding at depth. In Deep Sea Fishes; Randall, D.J., Farrell, A.P., Eds.; Academic Press: London, UK, 1997; pp. 115–194. [Google Scholar]
- Hernández-León, S.; Ikeda, T. A global assessment of mesozooplankton respiration in the ocean. J. Plankton Res. 2005, 27, 153–158. [Google Scholar] [CrossRef]
- Link, J.S.; Brodziak, J.K.T.; Edwards, S.F.; Overholtz, W.J.; Mountain, D.; Jossi, J.W.; Smith, T.D.; Fogarty, M.J. Marine ecosystem assessment in a fisheries management context. Can. J. Fish. Aquat. Sci. 2002, 59, 1429–1440. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Duarte, C.M. Seafaring in the 21St Century: The Malaspina 2010 Circumnavigation Expedition. Limnol. Oceanogr. Bull. 2015, 24, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Bradford-Grieve, J.M.; Markasheva, E.L.; Rocha, C.; Abiahy, B. South Atlantic Zooplankton. In Zooplankton, South Atlantic; Boltovskoy, D., Ed.; Blackhuys Publishers: Leiden, The Netherlands, 1999; p. 1705. [Google Scholar]
- Bradford-Grieve, J.M.; Markasheva, E.I.; Rocha, C.E.F.; Abiahy, B. Copepoda. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1999; Volume 2, pp. 869–1098. [Google Scholar]
- Razouls, C.; de Bovée, F.; Kouwemberg, J.; Desreumaux, N. Diversity and Geographic Distribution of Marine Planktonic Copepods 2019; Sorbonne University; Available online: http://copepodes.obs-banyuls.fr/en (accessed on 22 October 2019).
- Grice, G.D.; Hulseman, K. Abundance, vertical distribution and taxonomy of calanoid copepods at selected stations in the northeast Atlantic. Proc. Zool. Soc. Lond. 1965, 146, 213–262. [Google Scholar] [CrossRef]
- Vives, F.; Shmeleva, A. Crustacea: Copepodos marinos II. Non Calanoida. In Fauna Iberica Vol. 29; Ramos, M.A., Ed.; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2010; p. 486. [Google Scholar]
- Vives, F.; Shmeleva, A. Crustacea: Copepodos marinos I., Calanoida. In Fauna Iberica Vol 29; Ramos, M.A., Ed.; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2006; p. 1156. [Google Scholar]
- Markasheva, E.L. Calanoid Copepods of the Family Aetideidae of the World Ocean; Russian Academy of Science: Moscow, Russia; Zoological Institute: St. Peterburg, Russia, 1996. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Primer-6 Computer Program; Natural Environment Research Council: Plymouth, UK, 2005.
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.2-0. 2014. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 22 October 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bradford-Grieve, J.M.; Blanco-Bercial, L.; Prusova, I. Calanoides natalis Brady, 1914 (Copepoda: Calanoida: Calanidae): Identity and distribution in relation to coastal oceanography of the eastern Atlantic and western Indian Oceans. J. Nat. Hist. 2017, 51, 807–836. [Google Scholar] [CrossRef]
- Estrada, M.; Delgado, M.; Blasco, D.; Latasa, M.; Cabello, A.M.; Benítez-Barrios, V.; Fraile-Nuez, E.; Mozetic, P.; Vidal, M. Phytoplankton across Tropical and Subtropical Regions of the Atlantic, Indian and Pacific Oceans. PLoS ONE 2016, 11, e0151699. [Google Scholar] [CrossRef]
- Vinogradov, M.E. (Ed.) Vertical distribution of the oceanic zooplankton. In Oceanology, Biology of the Ocean; Nauka: Moscow, Russia, 1977; pp. 132–151. [Google Scholar]
- Yamaguchi, A.; Matsuno, K.; Homma, T. Spatial changes in the vertical distribution of calanoid copepods down to great depths in the North Pacific. Zool. Stud. 2015, 54, 13–17. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef]
- Deevey, G.B.; Brooks, A.L. Copepods of the Sargasso Sea off Bermuda: Species Composition, and Vertical and Seasonal Distribution between the Surface and 2000 M. Bull. Mar. Sci. 1977, 27, 256–291. [Google Scholar]
- Brugnano, C.; Granata, A.; Guglielmo, L.; Zagami, G. Spring diel vertical distribution of copepod abundances and diversity in the open Central Tyrrhenian Sea (Western Mediterranean). J. Mar. Syst. 2012, 105–108, 207–220. [Google Scholar] [CrossRef]
- Dias, C.O.; Araujo, A.V.; Paranhos, R.; Bonecker, S.L.C. Vertical copepod assemblages (0–2300 m) off southern Brazil. Zool. Stud. 2010, 49, 230–242. [Google Scholar]
- Chang, W.B.; Fang, L.S. Temporal and spatial variations in the species composition, distribution, and abundance of copepods in Kaohsiung Harbor, Taiwan. Zool. Stud. 2004, 43, 454–463. [Google Scholar]
- Vinogradov, M.E.; Vereshchaka, A.L.; Shushkina, E.A. Vertical structure of the zooplankton community in the oligotrophic areas of the north Atlantic and influence on the hydrothermal vent. Okeanologiya 1996, 36, 64–71. [Google Scholar]
- Angel, M.V. The pelagic environment of pelagic ocean. In Ecosystems of the Deep Oceans; Tyler, P.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 39–79. [Google Scholar]
- Scotto di Carlo, B.; Ianora, A.; Mazzocchi, M.G.; Scardi, M. Atlantis II Cruise: Uniformity of deep copepod assemblages in the Mediterranean Sea. J. Plankton Res. 1991, 13, 263–277. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Carlson, C.A.; Bates, N.R.; Goldthwait, S.A.; Madin, L.P.; Michaels, A.F. Zooplankton vertical migration and the active transport of dissolved organic and inorganic carbon in the Sargasso Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2000, 47, 137–158. [Google Scholar] [CrossRef]
- Webber, M.K.; Roff, J.C. Annual structure of the copepod community and its associated pelagic environment off Discovery Bay, Jamaica. Mar. Biol. 1995, 123, 467–479. [Google Scholar] [CrossRef]
- Dam, H.G.; Miller, C.A.; Jonasdottir, S.H. The trophic role of mesozooplankton at 47°N, 20°W during the North Atlantic Bloom Experiment. Deep Sea Res. Part II Top. Stud. Oceanogr. 1993, 40, 197–212. [Google Scholar] [CrossRef]
- Calbet, A.; Landry, M.R. Mesozooplankton influences on the microbial food web: Direct and indirect trophic interactions in the oligotrophic open ocean. Limnol. Oceanogr. 1999, 44, 1370–1380. [Google Scholar] [CrossRef]
- Siokou, I.; Zervoudaki, S.; Christou, E.D. Mesozooplankton community distribution down to 1000 m along a gradient of oligotrophy in the Eastern Mediterranean Sea (Aegean Sea). J. Plankton Res. 2013, 35, 1313–1330. [Google Scholar] [CrossRef] [Green Version]
- Cornils, A.; Schulz, J.; Schmitt, P.; Lanuru, M.; Richter, C.; Schnack-Schiel, S.B. Mesozooplankton distribution in the Spermonde Archipelago (Indonesia, Sulawesi) with special reference to the Calanoida (Copepoda). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 2076–2088. [Google Scholar] [CrossRef]
- Maycas, E.R.; Bourdillon, A.; Macquart-Moulin, C.; Passelaigue, F.; Patriti, G. Diel variations of the bathymetric distribution of zooplankton groups and biomass in Cap-Ferret Canyon, France. Deep Sea Res. Part II Top. Stud. Oceanogr. 1999, 46, 2081–2099. [Google Scholar] [CrossRef]
- Deevey, G.B. The annual cycle in quality and composition of the zooplankton of the Sargasso sea off Bermuda. I. The upper 500 m1. Limnol. Oceanogr. 1971, 16, 219–240. [Google Scholar] [CrossRef]
- Deevey, G.B.; Brooks, A.L. The annual cycle in quality and composition of the zooplankton of the Sargasso sea off Bermuda. II. The surface to 2,000 m1. Limnol. Oceanogr. 1971, 16, 927–943. [Google Scholar] [CrossRef]
- Wheeler, E.H.J. Atlantic Deep-Sea Galanoid Copepoda. Smithson. Contrib. Zool. 1970, 55, 1–31. [Google Scholar] [CrossRef]
- Hays, G.C.; Kennedy, H.; Frost, B.W. Individual variability in diel vertical migration of a marine copepod: Why some individuals remain at depth when others migrate. Limnol. Oceanogr. 2001, 46, 2050–2054. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J. Habitat dimensions of calanoid copepods in the western Gulf of Mexico. J. Mar. Res. 1983, 41, 163–188. [Google Scholar] [CrossRef]
- Jackson, M.L.; Smith, S.L. Vertical distribution of Eucalanoid copepods within the Costa Rica Dome area of the Eastern Tropical Pacific. J. Plankton Res. 2016, 38, 305–316. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
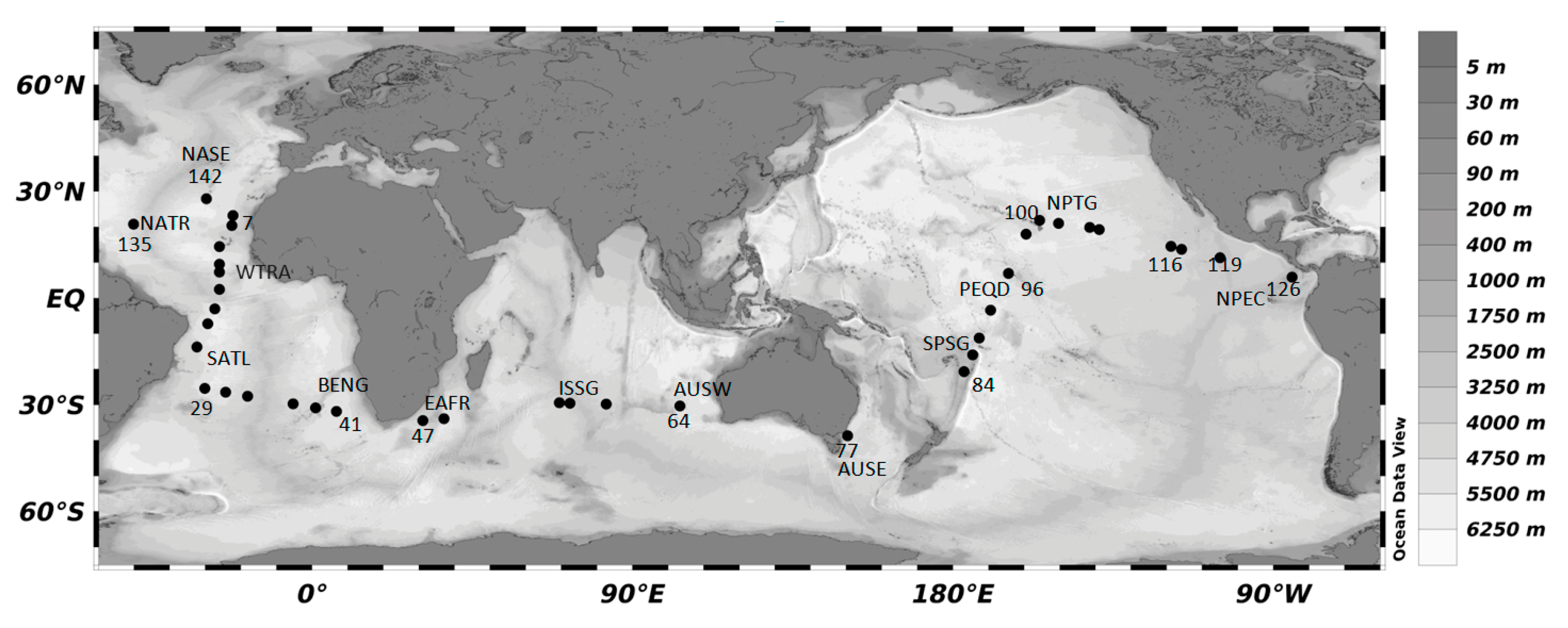
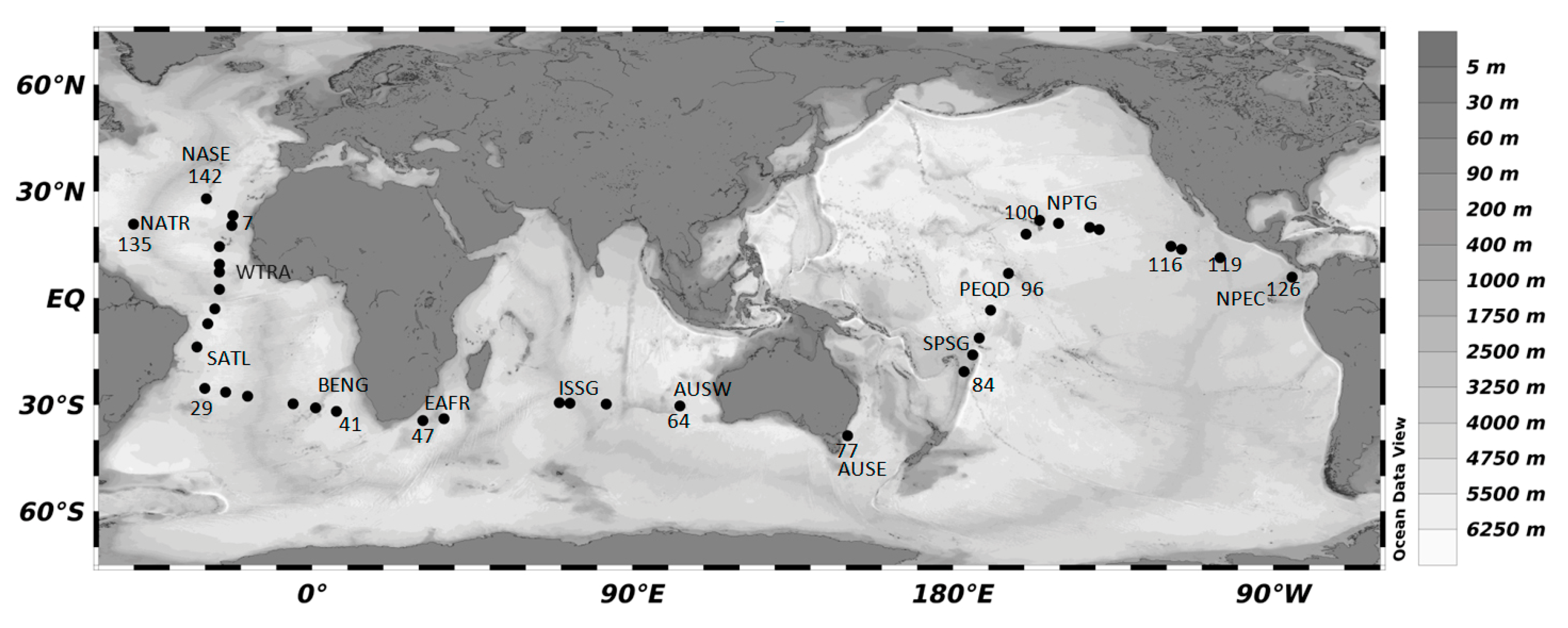
| Leg | Starting (City, Date) | End (City, Date) | Provinces | Abbrev. (Stations) |
|---|---|---|---|---|
| 1 | Cadiz (14/12/2011) | Rio de Janeiro (13/01/2011) | North Atlantic Tropical Gyre | NATR (5–10) |
| Western Tropical Atlantic | WTRA(11–18) | |||
| South Atlantic Gyre | SATL (19–26) | |||
| 2 | Rio de Janeiro (17/01/2011) | Cape Town (06/02/2011) | South Atlantic Gyre | SATL (27–40) |
| Benguela Current Coastal | BENG (41–44) | |||
| 3 | Cape Town (11/02/2011) | Perth (11/02/2011) | East Africa Coastal | EAFR(45–47) |
| Indian South Tropical Gyre West Australian Current | ISSG (48–63) AUSW (64–65) | |||
| 4 | Perth (17/03/2011) | Sydney (30/03/2011) | East Australian Current | AUSE (77) |
| 5 | Aukland (16/03/2011) | Honolulu (8/05/2011) | South Pacific Subtropical Gyre | SPSG (80–89) |
| Pacific Equatorial Divergence | PEQD (90–97) | |||
| North Pacific Tropical Gyre | NPTG (98–100) | |||
| 6 | Honolulu (10/05/2011) | Cartagena de Indias (10/06/2011) | North Pacific Tropical Gyre | NPTG (101–115) |
| North Pacific Equatorial Countercurrent | NPEC (116–126) | |||
| 7 | Cartagena de Indias (19/06/2011) | Cadiz (14/07/2011) | North Atlantic Tropical Gyre | NATR (131–139) |
| North Atlantic Subtropical Gyre | NASE (142–147) |
| Groups | Av. Abund. | % | Cum. % | |
|---|---|---|---|---|
| Epipelagic (0–200 m) Average similarity: 63% | Copepods | 3.4 | 54.2 | 54 |
| Chaetognaths | 1.2 | 14.5 | 69 | |
| Siphonophors | 0.9 | 11.3 | 80 | |
| Ostracods | 0.7 | 7.2 | 87 | |
| Euphausiids | 0.5 | 4.7 | 92 | |
| Upper mesopelagic (200–500 m) Average similarity: 45% | Copepods | 1.5 | 62.5 | 62 |
| Ostracods | 0.5 | 12.5 | 75 | |
| Chaetognaths | 0.4 | 8.8 | 84 | |
| Siphonophors | 0.2 | 7.9 | 92 | |
| Lower mesopelagic (500–1000 m) Average similarity: 42% | Copepods | 0.8 | 79.2 | 79 |
| Chaetognaths | 0.1 | 8.1 | 87 | |
| Siphonophors | 0.1 | 6.2 | 93 | |
| Upper bathypelagic (1000–2000 m) Average similarity: 41% | Copepods | 0.3 | 87 | 87 |
| Chaetognaths | 0.03 | 5.6 | 93 | |
| Lower bathypelagic (2000–3000 m) Average similarity: 50% | Copepods | 0.14 | 91.7 | 92 |
| Acartiidae | Arietellidae | Candaciidae (Continuation) | Euchaetidae (Continuation) |
|---|---|---|---|
| Acartia danae Giesbrecht, 1889 | Arietellus aculeatus (T. Scott, 1894) | Candacia ethiopica Dana, 1849 | Euchaeta media Giesbrecht, 1888 |
| Acartia enzoi (Crisafi, 1974 ) | Arietellus giesbrechti Sars G.O., 1905 | Candacia katchumi Grice, 1961 | Euchaeta spinosa Giesbrecht, 1892 |
| Acartia longiremis (Lilljeborg, 1853) | Arietellus pavoninus Sars G.O., 1905 | Candacia longimana Claus, 1863 | Paraeuchaeta exigua (Wolfenden, 1911) |
| Acartia negligens Dana, 1849 | Augaptilidae | Candacia pachydactila Dana, 1849 | Paraeuchaeta tonsa (Giesbrecht, 1895) |
| Aetideidae | Augaptilus longicaudatus (Claus, 1863) | Candacia tenuimana Giesbrecht, 1889 | Fosshageniidae |
| Aetideus acutus (Farran, 1929) | Centraugaptilus rattrayi (T. Scott, 1894) | Candacia truncata (Dana, 1849) | Temoropia spp. T. Scott, 1894 |
| Aetideus australis (Vervoort, 1957) | Euaugaptilus hecticus (Giesbrecht, 1889) | Candacia simplex Giesbrecht, 1889 | Temoropia mayumbaensis T. Scott, 1894 |
| Aetideus armatus (Boeck, 1872) | Euaugaptilus magnus (Wolfenden, 1904) | Candacia varicans Giesbrecht, 1893 | Heterorhabdidae |
| Aetideus giesbrechti Cleve, 1904 | Euaugaptilus marginatus (Tanaka, 1964) | Centropagidae | Disseta magna Bradford, 1971 |
| Aetideus pseudoarmatus (Bradford, 1971) | Euaugaptilus palumboi Giesbrecht, 1889 | Centropages sp. Kröyer, 1849 | Disseta palumboi Giesbrecht, 1889 |
| Bradyidius armatus (Giesbrecht, 1897) | Haloptilus acutifrons (Giesbrecht, 1892) | Centropages calaninus (Dana, 1849) | Heterorhabdus clausi (Giesbrecht, 1889) |
| Chiridius gracilis (Farran, 1908) | Haloptilus aculeatus (Brady, 1883) | Centropages elongatus (Giesbrecht, 1896 ) | Heterorhabdus papilliger (Claus, 1863 ) |
| Chiridius poppei Giesbrecht, 1892 | Haloptilus fons Farran, 1908 | Centropages furcatus (Dana, 1849) | Heterorhabdus spinifer Park, 1970 |
| Chirundina streetsii Giesbrecht, 1895 | Haloptilus longicornis Claus, 1863 | Centropages longicornis Mori, 1932 | Heterorhabdus spinifrons (Claus, 1863) |
| Euchirella amoena Giesbrecht, 1888 | Haloptilus ornatus (Giesbrecht, 1893) | Clausocalanidae | Heterostylites longicornis (Giesbrecht, 1889) |
| Euchirella curticauda Giesbrecht, 1888 | Haloptilus oxycephalus (Giesbrecht, 1889) | Clausocalanus arcuicornis (Dana, 1849) | Heterostylites major (F. Dahl, 1894) |
| Euchirella máxima Wolfenden, 1905 | Haloptilus spiniceps (Giesbrecht, 1893) | Clausocalanus farrani (Sewell, 1929) | Paraheterorhabdus compactus (G.O. Sars, 1900) |
| Euchirella messiniensis (Claus, 1863) | Pachyptilus eurygnathus G.O. Sars, 1920 | Clausocalanus furcatus (Brady, 1883) | Lucicutiidae |
| Euchirella pulchra (Lubbock, 1856) | Pachyptilus sp. G.O. Sars, 1920 | Clausocalanus ingens Frost & Fleminger, 1968 | Lucicutia bicornuta Wolfenden, 1905 |
| Euchirella rostrata (Claus, 1866) | Bathypontiidae | Clausocalanus jobei Frost & Fleminger, 1968 | Lucicutia clausi (Giesbrecht, 1889) |
| Euchirella splendens (Vervoort, 1963) | Temorites spp. G.O. Sars, 1900 | Clausocalanus lividus Frost & Fleminger, 1968 | Lucicutia curta Farran, 1905 |
| Euchirella truncata Esterly, 1911 | Temorites brevis G.O. Sars,1900 | Clausocalanus mastigophorus (Claus, 1863) | Lucicutia flavicornis (Claus, 1863) |
| Gaetanus spp. Giesbrecht, 1888 | Calanidae | Clausocalanus minor (Sewell, 1929) | Lucicutia gaussae Grice, 1963 |
| Gaetanus armiger Giesbrecht, 1888 | Canthocalanus pauper (Giesbrecht, 1888) | Clausocalanus parapergens Frost & Fleminger, 1968 | Lucicutia gemina Farran, 1926 |
| Gaetanus brevicaudatus G.O. Sars, 1907 | Calanoides cf. carinatus (Krøyer, 1848)* | Clausocalanus paululus Farran, 1926 | Lucicutia grandis (Giesbrecht, 1895) |
| Gaetanus brevicornis Esterly, 1906 | Calanoides macrocarinatus (Brodsky, 1967)* | Clausocalanus pergens Farran, 1926 | Lucicutia intermedia G.O. Sars, 1905 |
| Gaetanus kruppii Giesbrecht, 1903 | Cosmocalanus darwini (Lubbock, 1860) | Ctenocalanus vanus Giesbrecht, 1888 | Lucicutia longicornis (Giesbrecht, 1889) |
| Gaetanus latifrons G.O. Sars, 1905 | Mesocalanus tenuicornis (Dana, 1849) | Eucalanidae | Lucicutia longiserrata (Giesbrecht, 1889) |
| Gaetanus miles Giesbrecht, 1888 | Nannocalanus minor (Claus, 1863) | Eucalanus hyalinus (Claus, 1866) | Lucicutia magna Wolfenden, 1903 |
| Gaetanus minor Farran, 1905 | Neocalanus gracilis (Dana, 1849) | Pareucalanus sewelli (Fleminger, 1973) | Lucicutia maxima Steuer, 1904 |
| Gaetanus pileatus Farran, 1903 | Neocalanus robustior (Giesbrecht, 1888) | Pareucalanus attenuatus (Dana, 1849) | Lucicutia ovalis (Giesbrecht, 1889) |
| Gaetanus pungens Giesbrecht, 1895 | Neocalanus tonsus (Brady, 1883) | Subeucalanus crassus (Giesbrecht, 1888) | Lucicutia wolfendini (Sewell, 1932) |
| Gaetanus brevispinus G.O. Sars, 1900 | Undinula vulgaris (Dana, 1849) | Subeucalanus monachus (Giesbrecht, 1888) | Megacalanidae |
| Gaetanus tenuispinus G.O. Sars, 1900 | Candaciidae | Subeucalanus subtenuis (Giesbrecht, 1888) | Bradycalanus sarsi (Farran, 1939) |
| Pseudochirella sp. G.O. Sars, 1920 | Candacia bipinnata Giesbrecht, 1889 | Subeucalanus longiceps (Matthews, 1925) | Megacalanus princeps Wolfenden, 1904 |
| Pseudochirella spinosa (Wolfenden, 1905) | Candacia bispinosa Claus, 1863 | Euchaetidae | Metridinidae |
| Pseudochirella major (Sars, 1907) | Candacia catula (Giesbrecht, 1889) | Euchaeta acuta Giesbrecht, 1893 | Metridia sp. Boeck, 1865 |
| Undeuchaeta major Giesbrecht, 1888 | Candacia cheirura Cleve, 1904 | Euchaeta concinna Dana, 1849 | Metridia brevicauda Giesbrecht, 1889 |
| Undeuchaeta plumosa (Lubbock, 1856) | Candacia elongata Boeck, 1872 | Euchaeta marina (Prestandrea, 1833) | |
| Metridinidae | Rhincalanidae | Tortanidae | Mormonillidae |
| Metridia curticauda Giesbrecht, 1889 | Rhincalanus cornutus (Dana, 1849) | Tortanus spp. (Giesbrecht, 1898) | Mormonilla spp. Giesbrecht, 1891 |
| Metridia longa (Lubbock, 1854) | Rhincalanus gigas Brady, 1883 | Phaennidae | Oithonidae |
| Metridia lucens Boeck, 1864 | Rhincalanus nasutus Giesbrecht, 1888 | Onchocalanus spp.(G.O. Sars, 1905) | Oithona atlantica Farran, 1908 |
| Metridia princeps Giesbrecht, 1889 | Scolecitrichidae | Phaenna spinifera (Claus, 1863) | Oithona brevicornis Giesbrecht, 1891 |
| Metridia venusta Giesbrecht, 1889 | Amallothrix farrani Rose, 1942 | Xanthocalanus spp. Giesbrecht, 1893 | Oithona nana Giesbrecht, 1892 |
| Pleuromamma abdominalis (Lubbock, 1856) | Amallothrix spp. G.O. Sars, 1925 | Xanthocalanus agilis Giesbrecht, 1893 | Oithona plumifera Baird, 1843 |
| Pleuromamma borealis (F. Dahl, 1893) | Landrumius gigas (A. Scott, 1909) | Aegisthidae | Oithona robusta Giesbrecht, 1891 |
| Pleuromamma gracilis (Claus, 1863) | Lophothrix frontalis Giesbrecht, 1895 | Aegisthus spp. Giesbrecht, 1891 | Oithona setigera (Dana, 1849) |
| Pleuromamma piseki Farran, 1929 | Lophothrix humilifrons G.O. Sars, 1905 | Aegisthus aculeatus Giesbrecht, 1891 | Oithona tenuis Rosendorn, 1917 |
| Pleuromamma quadrungulata (F. Dahl, 1893) | Lophothrix latipes (T. Scott, 1894) | Aegisthus mucronatus Giesbrecht, 1891 | Oithona spp. Baird, 1843 |
| Pleuromamma robusta (F. Dahl, 1893) | Lophothrix quadrispinosa Wolfenden, 1911 | Corycaeidae | Oncaeidae |
| Pleuromamma xiphias (Giesbrecht, 1889) | Pseudoamallothrix emarginata (Farran, 1905) | Agetus flaccus (Giesbrecht, 1891) | Oncaea spp. Philippi, 1843 |
| Nullosetigeridae | Pseudoamallothrix obtusifrons (G.O. Sars, 1905) | Agetus limbatus (Brady, 1883) | Oncaea media Giesbrecht, 1891 |
| Nullosetigera aequalis (G.O. Sars, 1920) | Scaphocalanus sp. G.O. Sars, 1900 | Agetus typicus Krøyer, 1849 | Oncaea mediterranea (Claus, 1863) |
| Nullosetigera bidentata (Brady, 1883) | Scaphocalanus affinis (G.O. Sars, 1905) | Corycaeus clausi F. Dahl, 1894 | Oncaea mediterranea flava Giesbrecht, 1892 |
| Nullosetigera helgae (Farran, 1908) | Scaphocalanus brevicornis (G.O. Sars, 1900) | Corycaeus crassiusculus Dana, 1849 | Oncaea venusta Philippi, 1843 |
| Nullosetigera impar (Farran, 1908) | Scaphocalanus brevirostris Park, 1970 | Corycaeus speciosus Dana, 1849 | Triconia conifera (Giesbrecht, 1891) |
| Nullosetigera mutica (G.O. Sars, 1907) | Scaphocalanus emine Uysal & Shmeleva, 2002 | Ditrichocorycaeus amazonicus (F. Dahl, 1894 ) | Triconia dentipes (Giesbrecht, 1891) |
| Paracalanidae | Scaphocalanus insignis Brodsky, 1950 | Farranula carinata (Giesbrecht, 1891) | Conaea spp. Giesbrecht, 1891 |
| Acrocalanus andersoni Bowman, 1958 | Scaphocalanus magnus (T. Scott, 1894) | Farranula gracilis (Dana, 1849) | Pachos punctatum (Claus, 1863) |
| Acrocalanus gibber Giesbrecht, 1888 | Scolecithricella abyssalis (Giesbrecht, 1888) | Farranula rostrata (Claus, 1863) | Pachos tuberosum (Giesbrecht, 1891) |
| Acrocalanus gracilis Giesbrecht, 1888 | Scolecithricella profunda (Giesbrecht, 1892) | Onychocorycaeus agilis ( Dana, 1849 ) | Sapphirinidae |
| Acrocalanus longicornis Giesbrecht, 1888 | Scolecithricella dentata (Giesbrecht, 1892) | Onychocorycaeus giesbrechti (F. Dahl, 1894) | Sapphirina spp. J. V. Thompson, 1830 |
| Acrocalanus monachus Giesbrecht, 1888 | Scolecithricella minor (Brady, 1883) | Urocorycaeus furcifer (Claus, 1863) | Copilia quadrata Dana, 1849 |
| Calocalanus contractus Farran, 1926 | Scolecithricella tenuiserrata (Giesbrecht, 1893) | Urocorycaeus longistylis (Dana, 1849) | Copilia hendorffi ( Dahl, 1892) |
| Calocalanus elegans Shmeleva, 1965 | Scolecithricella vittata (Giesbrecht, 1893) | Clytemnestridae | Copilia mirabilis Dana, 1849 |
| Calocalanus indicus Shmeleva, 1974 | Scolecithrix bradyi Giesbrecht, 1888 | Clytemnestra spp. Dana, 1847 | |
| Calocalanus pavo (Dana, 1849) | Scolecithrix danae (Lubbock, 1856) | Ectinosomatidae | |
| Calocalanus pavoninus Farran, 1936 | Scottocalanus securifrons (T. Scott, 1894) | Microsetella norvegica (Boeck, 1864) | |
| Calocalanus plumulosus (Claus, 1863) | Spinocalanidae | Microsetella rosea (Dana, 1848) | |
| Calocalanus styliremis Giesbrecht, 1888 | Spinocalanus spp.(Giesbrecht, 1888) | Lubbockiidae | |
| Calocalanus tenuis Farran, 1926 | Spinocalanus longicornis G.O. Sars,1900 | Lubbockia aculeata Giesbrecht, 1891) | |
| Paracalanus indicus Wolfenden, 1905 | Monacilla sp. G.O. Sars, 1905 | Lubbockia squillimana Claus, 1863 | |
| Paracalanus nanus G.O. Sars, 1907) | Monacilla typica G.O. Sars, 1905 | Miraciidae | |
| Paracalanus parvus (Claus, 1863) | Temoridae | Miracia efferata Dana, 1849 | |
| Mecynocera clausi I.C.Thompson, 1888 | Temora discaudata Giesbrecht, 1889 | Macrosetella gracilis (Dana, 1847) | |
| Pontellidae | Temora stylifera Dana, 1849 | ||
| Calanopia elliptica (Dana, 1849) | |||
| Calanopia minor A. Scott, 1902 | |||
| Labidocera acutifrons (Dana, 1849) | |||
| Pontellina plumata (Dana, 1849) |
| Atlantic | Indian | Pacific | ||||
|---|---|---|---|---|---|---|
| Species | % | Species | % | Species | % | |
| Epipelagic | C. furcatus | 11.3 | A. negligens | 7 | N. minor | 10 |
| O. plumifera | 8.4 | L. flavicornis | 6.1 | Acrocalanus | 6.2 | |
| N. minor | 8.1 | U. furcifer | 5.4 | A. danae | 4.6 | |
| M. tenuicornis | 5.9 | C. pavo | 5.2 | E. marina | 4.5 | |
| E. marina | 5.4 | M. tenuicornis | 5.1 | C. furcatus | 4.5 | |
| A. danae | 4.9 | O. plumifera | 4.9 | F. gracilis | 4.3 | |
| C. pavo | 4.8 | Pleuromamma juv. | 4.9 | O. plumifera | 4.1 | |
| C. arcuicornis | 4.2 | M. tenuicornis | 4.1 | |||
| H. spinifrons | 3.9 | N. gracilis | 3.3 | |||
| P. indicus | 3 | |||||
| AvS: 22% | AvS: 14% | AvS: 24% | ||||
| Upper Mesopelagic | P. gracilis | 16 | Pleuromamma juv. | 25 | T. conifera | 14 |
| P. abdominalis | 15.2 | T. conifera | 10.7 | P. abdominalis | 8.7 | |
| P. piseki | 10.5 | M. tenuicornis | 11 | L. clausi | 6.8 | |
| O. mediter. flava | 6.1 | C. jobei | 10.2 | P. xiphias | 6.8 | |
| U. furcifer | 4.8 | P. gracilis | 7.7 | P. piseki | 6.1 | |
| T. conifera | 2.7 | P. robusta | 6 | |||
| AvS: 14% | AvS: 17% | AvS: 13% | ||||
| Lower Mesopelagic | M. brevicauda | 22 | P. abdominalis | 58 | Conaea | 27 |
| Conaea | 10.4 | T. conifera | 28 | M. brevicauda | 13.5 | |
| P. xiphias | 9.1 | Heterostylites | 13.5 | P. abdominalis | 7.1 | |
| N. helgae | 8.2 | T. conifera | 5.2 | |||
| R. cornutus | 8.2 | P. attenuatus | 4.2 | |||
| AvS: 12% | AvS: 6% | AvS: 8% | ||||
| Upper Bathypelagic | Conaea | 31 | M. brevicauda | 66.6 | M. brevicauda | 17 |
| Copepodites | 10 | L. gaussae | 33.4 | T. conifera | 14.6 | |
| M. brevicauda | 7.7 | Copepodites | 10.7 | |||
| T. conifera | 7.1 | Small calanoids* | 12.9 | |||
| AvS: 10% | AvS: 4% | AvS: 7% | ||||
| Lower Bathypelagic | Copepodites | 63.2 | Small calanoids* | 60.6 | ||
| T. conifera | 12.8 | Copepodites | 12 | |||
| Conaea | 8.5 | M. brevicauda | 10 | |||
| Conaea | 8 | |||||
| AvS: 7% | AvS: 4% | |||||
| Epipelagic AvS: 23% | Upper Mesopelagic AvS: 11% | Lower Mesopelagic AvS: 9% | Upper Bathypelagic AvS: 7% | Lower Bathypelagic AvS: 5% | |||||
|---|---|---|---|---|---|---|---|---|---|
| Species | % | Species | % | Species | % | Species | % | Species | % |
| C. furcatus | 8.9 | P.abdominalis | 12.9 | M. brevicauda | 18.3 | Conaea | 17.4 | Calanoids BC | 31.6 |
| N. minor | 8.6 | P. gracilis | 10 | Conaea | 16.7 | M. brevicauda | 10.4 | Small calanoids* | 31.5 |
| O. plumifera | 5.6 | T. conifera | 8.4 | P. xiphias | 9 | Calanoids BC | 7.1 | T. conifera | 6.1 |
| E.marina | 5.4 | P. piseki | 7.6 | P. abdominalis | 6.3 | Small calanoids* | 6.9 | Copepodites | 4.9 |
| M. tenuicornis | 4.8 | O. mediter. flava | 4.7 | R. cornutus | 5.4 | T. conifera | 6.4 | Oithona spp. | 4.7 |
| C. pavo | 3.9 | Pleuromamma juv. | 4.4 | T. conifera | 5.3 | N. helgae | 3.5 | Oncaea spp. | 4.6 |
| A. danae | 3.5 | Copepodites | 3.9 | N. helgae | 5.3 | H. spinifrons | 3.5 | Conaea | 4.1 |
| A. negligens | 3.3 | M. brevicauda | 3.7 | Copepodites | 3.5 | O. plumifera | 3.5 | M. brevicauda | 3.7 |
| Acrocalanus spp. | 3.3 | L. gemina | 3.4 | O. plumifera | 3.3 | Aegisthus | 3.4 | ||
| F. gracilis | 2.6 | U. furcifer | 3.2 | H. spinifrons | 2.8 | P. abdominalis | 2.4 | ||
| S. danae | 2.4 | L. clausi | 2.7 | Heterostylites | 2 | M. princeps | 2.4 | ||
| Euchaeta juv. | 2.1 | P. xiphias | 2.3 | Pareucalanus | 1.9 | ||||
| C. jobei | 2.0 | P. robusta | 2.3 | C. furcatus | 1.8 | ||||
| N. gracilis | 1.9 | O. plumifera | 1.9 | ||||||
| N. robustior | 1.7 | H. spinifrons | 1.9 | ||||||
| O. atlantica | 1.6 | ||||||||
| H. longicornis | 1.6 | ||||||||
| L. gemina | 1.5 | ||||||||
| O. agilis | 1.5 | ||||||||
| O. setigera | 1.5 | ||||||||
| O. venusta | 1.5 | ||||||||
| Copepodites | 1.3 | ||||||||
| Pareucalanus | 1.2 | ||||||||
| C. farrani | 1.1 | ||||||||
| M. clausi | 1.1 | ||||||||
| Total | 78.4 | 73.3 | 81.6 | 66.9 | 91.2 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández de Puelles, M.L.; Gazá, M.; Cabanellas-Reboredo, M.; Santandreu, M.d.M.; Irigoien, X.; González-Gordillo, J.I.; Duarte, C.M.; Hernández-León, S. Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean. Diversity 2019, 11, 203. https://doi.org/10.3390/d11110203
Fernández de Puelles ML, Gazá M, Cabanellas-Reboredo M, Santandreu MdM, Irigoien X, González-Gordillo JI, Duarte CM, Hernández-León S. Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean. Diversity. 2019; 11(11):203. https://doi.org/10.3390/d11110203
Chicago/Turabian StyleFernández de Puelles, Mᵃ Luz, Magdalena Gazá, Miguel Cabanellas-Reboredo, Mᵃ del Mar Santandreu, Xabier Irigoien, Juan Ignacio González-Gordillo, Carlos M. Duarte, and Santiago Hernández-León. 2019. "Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean" Diversity 11, no. 11: 203. https://doi.org/10.3390/d11110203
APA StyleFernández de Puelles, M. L., Gazá, M., Cabanellas-Reboredo, M., Santandreu, M. d. M., Irigoien, X., González-Gordillo, J. I., Duarte, C. M., & Hernández-León, S. (2019). Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean. Diversity, 11(11), 203. https://doi.org/10.3390/d11110203