Swimming Abilities of Temperate Pelagic Fish Larvae Prove that They May Control Their Dispersion in Coastal Areas

 ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Material and Methods
2.1. Model Species
2.2. Overview of the Experimental Design
2.2.1. Critical Swimming Speed (Ucrit)
2.2.2. Swimming Endurance
2.2.3. Locomotory Behavior
2.2.4. Physiological Condition
2.3. Data Analyses
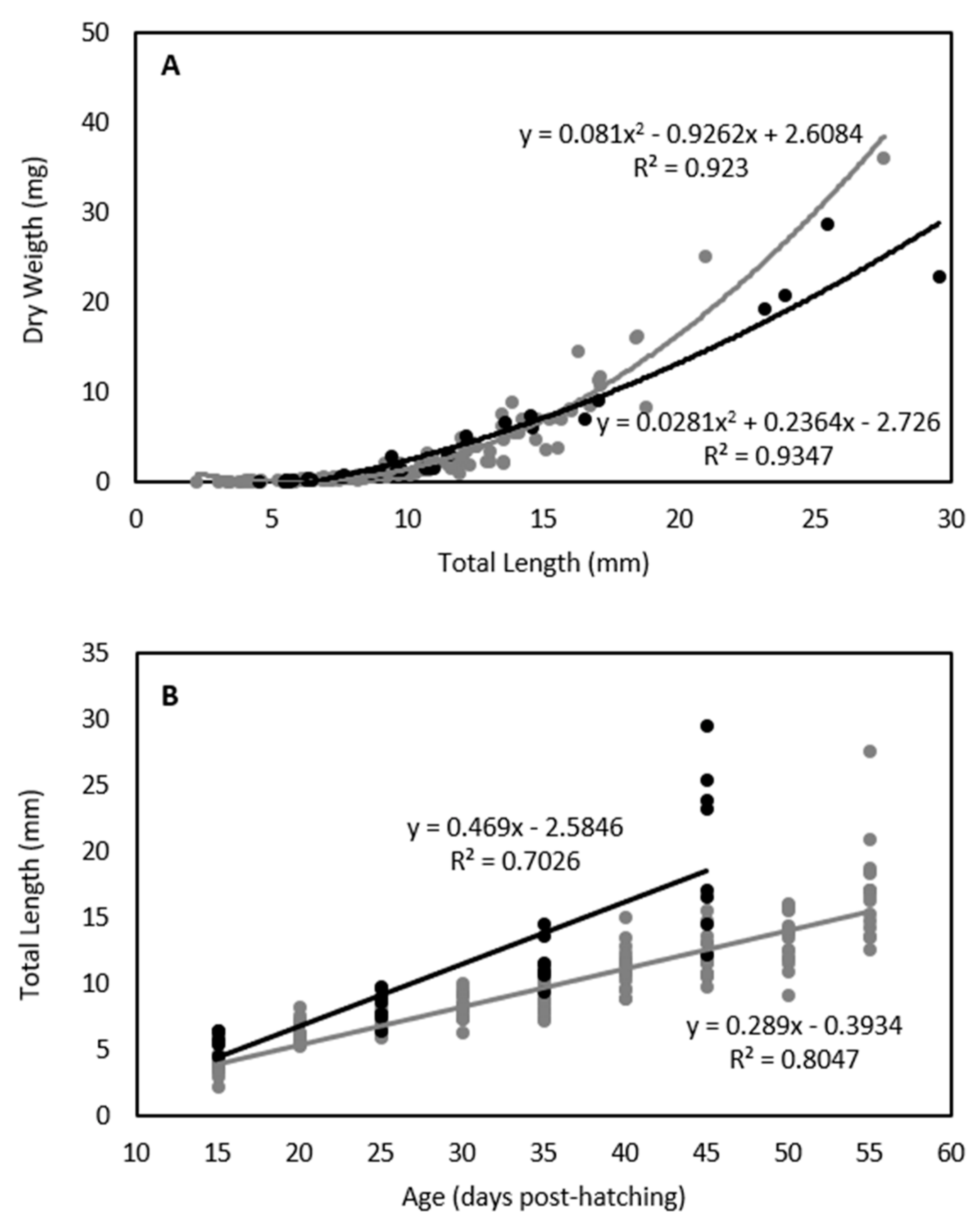
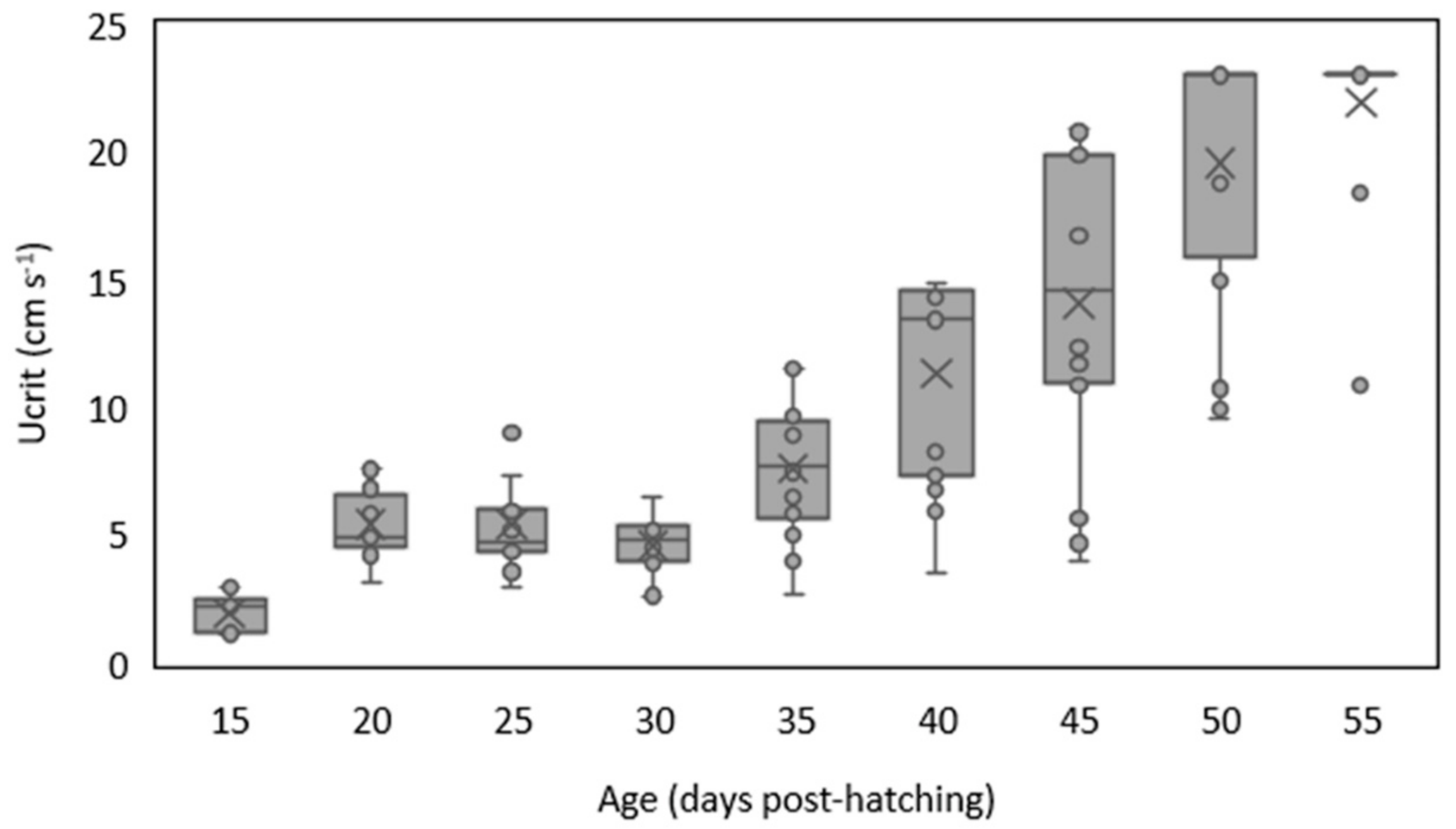
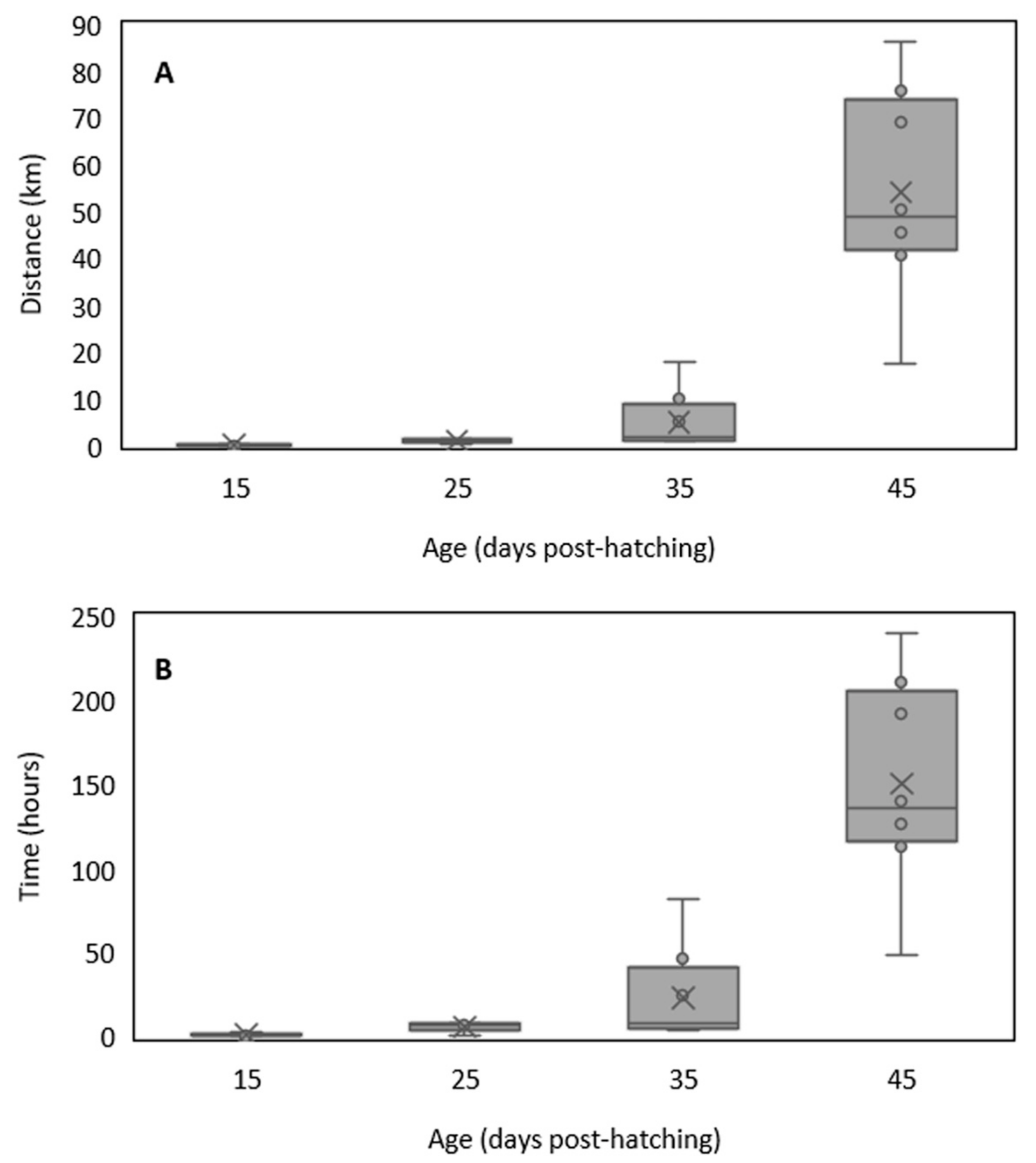
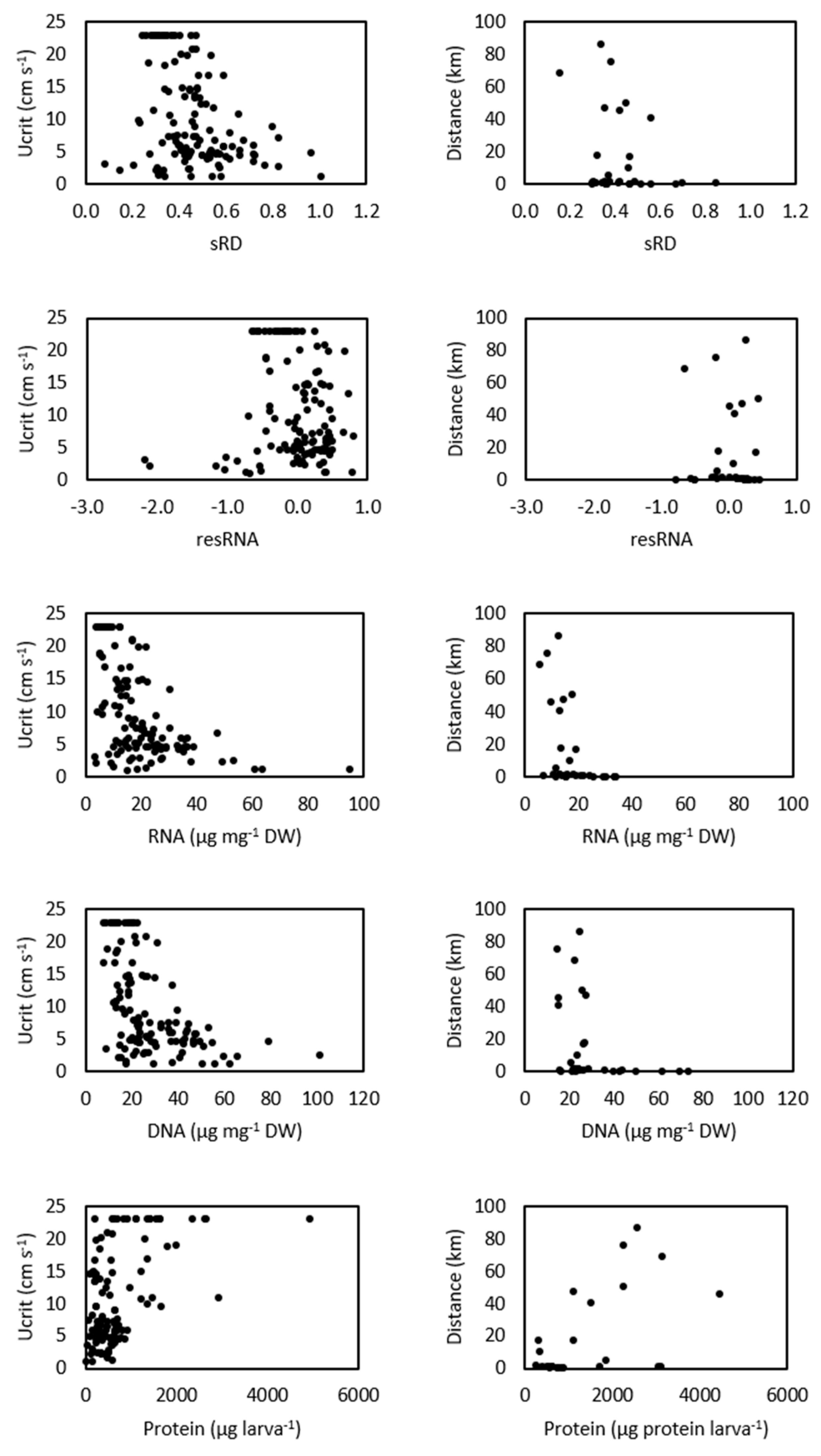
3. Results
4. Discussion
4.1. Swimming Performance of Temperate Fish Larvae
4.2. Swimming Performance and Larval Condition
4.3. Climate Change and the Swimming Performance of Temperate Fish Larvae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Bradbury, I.R.; Snelgrove, P.V.R. Contrasting larval transport in demersal fish and benthic invertebrates: The roles of behaviour and advective processes in determining spatial pattern. Can. J. Fish. Aquat. Sci. 2001, 58, 811–823. [Google Scholar] [CrossRef]
- Teodósio, M.A.; Paris, C.B.; Wolanski, E.; Morais, P. Biophysical processes leading to the ingress of temperate fish larvae into estuarine nursery areas: A review. Estuar. Coast. Shelf Sci. 2016, 183, 187–202. [Google Scholar] [CrossRef]
- Montgomery, J.C.; Tolimieri, N.; Haine, O.S. Active habitat selection by pre-settlement reef fishes. Fish Fish. 2001, 2, 261–277. [Google Scholar] [CrossRef]
- Leis, J.M. Are larvae of demersal fishes plankton or nekton? Adv. Mar. Biol. 2006, 51, 59–141. [Google Scholar] [CrossRef]
- Houde, E.D. Emerging from Hjort’s shadow. J. Northwest Atl. Fish Sci. 2008, 41, 53–70. [Google Scholar] [CrossRef]
- Faillettaz, R.; Blandin, A.; Paris, C.B.; Koubbi, P.; Irisson, J.O. Suncompass orientation in Mediterranean fish larvae. PLoS ONE 2015, 10, e0135213. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, H.; Atema, J.; Kingsford, M.J.; Gerlach, G. Sun compass orientation helps coral reef fish larvae return to their natal reef. PLoS ONE 2013, 8, e66039. [Google Scholar] [CrossRef]
- Atema, J.; Kingsford, M.J.; Gerlach, G. Larval reef fish could use odour for detection, retention and orientation to reefs. Mar. Ecol. Prog. Ser. 2002, 241, 151–160. [Google Scholar] [CrossRef]
- Watt-Pringle, P.; Strydom, N.A. Habitat use by larval fishes in a temperate South African surf zone. Estuar. Coast. Shelf Sci. 2003, 58, 765–774. [Google Scholar] [CrossRef]
- Faillettaz, R.; Paris, C.B.; Irisson, J.O. Larval fish swimming behavior alters dispersal patterns from marine protected areas in the North-Western Mediterranean Sea. Front. Mar. Sci. 2018, 5, 97. [Google Scholar] [CrossRef]
- Lett, C.; Barriera, N.; Ourmièresb, Y.; Petita, C.; Labonnea, M.; Bourjeaa, J.; Darnaudea, A.M. Modeling larval dispersal for the gilthead seabream in the northwestern Mediterranean Sea. Mar. Environ. Res. 2019, 104781. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.J.; Higgs, D.M.; Belanger, A.J.; Leis, J.M. Auditory and olfactory abilities of pre-settlement larvae and post-settlement juveniles of a coral reef damselfish (Pisces: Pomacentridae). Mar. Biol. 2005, 147, 1425–1434. [Google Scholar] [CrossRef]
- Wolanski, E.; Kingsford, M.J. Oceanographic and behavioural assumptions in models of the fate of coral and coral reef fish larvae. J. R. Soc. Interface 2014, 11, 20140209. [Google Scholar] [CrossRef] [PubMed]
- Stobutzki, I.C.; Bellwood, D.R. Sustained swimming abilities of the late pelagic stages of coral reef fishes. Mar. Ecol. Prog. Ser. 1997, 149, 35–41. [Google Scholar] [CrossRef]
- Atema, J.; Gerlach, G.; Paris, C.B. Sensory biology and navigation behavior of reef fish larvae. In Ecology of Fishes on Coral Reefs; Mora, C., Ed.; Cambridge University Press: Cambridge, UK, 2015; pp. 3–15. [Google Scholar]
- Paris, C.; Cowen, R.K. Direct evidence of a biophysical retention mechanism for coral reef fish larvae. Limnol. Oceanogr. 2004, 49, 1964–1979. [Google Scholar] [CrossRef]
- Jørgensen, C.; Opdal, A.F.; Fiksen, Ø. Can behavioural ecology unite hypotheses for fish recruitment? ICES J. Mar. Sci. 2014, 71, 909–917. [Google Scholar] [CrossRef]
- Weins, J.A.; Stenseth, N.C.; van Horne, B.; Ins, R.A. Ecological mechanisms and landscape ecology. Oikos 1995, 66, 369–380. [Google Scholar] [CrossRef]
- Faria, A.M.; Muha, T.; Morote, E.; Chícharo, M.A. Influence of starvation on the critical swimming behavior of the Senegalese sole (Solea senegalensis) and its relationship with RNA:DNA ratios during ontogeny. Sci. Mar. 2011, 75, 87–94. [Google Scholar] [CrossRef]
- Kashef, N.S.; Sogard, S.M.; Fisher, R.; Largier, J.L. Ontogeny of critical swimming speeds for larval and pelagic juvenile rockfishes (Sebastes spp., family Scorpaenidae). Mar. Ecol. Prog. Ser. 2014, 500, 231–243. [Google Scholar] [CrossRef]
- Fisher, R.; Bellwood, D.R.; Job, S.D. Development of swimming abilities in reef fish larvae. Mar. Ecol. Prog. Ser. 2000, 202, 163–173. [Google Scholar] [CrossRef]
- Stobutzki, I.C.; Bellwood, D.R. An analysis of the sustained swimming abilities of pre- and post-settlement coral reef fishes. J. Exp. Mar. Bio. Ecol. 1994, 175, 275–286. [Google Scholar] [CrossRef]
- Fisher, R.; Leis, J.M.; Clark, D.; Wilson, S.K. Critical swimming speeds of late-stage coral reef fish larvae: Variation within species, among species and between locations. Mar. Biol. 2005, 147, 1201–1212. [Google Scholar] [CrossRef]
- Leis, J.M.; Wright, K.J.; Johnson, R.N. Behaviour that influences dispersal and connectivity in the small, young larvae of a reef fish. Mar. Biol. 2007, 153, 103–117. [Google Scholar] [CrossRef]
- Leis, J.M.; Hay, A.C.; Howarth, G.J. Ontogeny of in situ behaviours relevant to dispersal and population connectivity in larvae of coral-reef fishes. Mar. Ecol. Prog. Ser. 2009, 379, 163–179. [Google Scholar] [CrossRef]
- Koumoundouros, G.; Ashton, C.; Xenikoudakis, G.; Giopanou, I.; Georgakopoulou, E.; Stickland, N. Ontogenetic differentiation of swimming performance in gilthead seabream (Sparus aurata, Linnaeus 1758) during metamorphosis. J. Exp. Mar. Bio. Ecol. 2009, 370, 75–81. [Google Scholar] [CrossRef]
- Faria, A.M.; Borges, R.; Gonçalves, E.J. Critical swimming speeds of wild-caught sand-smelt Atherina presbyter larvae. J. Fish. Biol. 2014, 85, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.A.; Huebert, K.B.; Llopiz, J.K. Intrinsic and extrinsic factors driving match-mismatch dynamics during the early life history of marine fishes. Adv. Ecol. Res. 2012, 47, 177–302. [Google Scholar] [CrossRef]
- Silva, L.; Moyano, M.; Illing, B.; Faria, A.M.; Garrido, S.; Peck, M.A. Ontogeny of swimming capacity in plaice (Pleuronectes platessa) larvae. Mar. Biol. 2015, 162, 753–761. [Google Scholar] [CrossRef]
- Faillettaz, R.; Durand, E.; Paris, C.B.; Koubbi, P.; Irisson, J.-O. Swimming speeds of Mediterranean settlement-stage fish larvae nuance Hjort’s aberrant drift hypothesis. Limnol. Oceanogr. 2018, 63, 509–523. [Google Scholar] [CrossRef]
- Rossi, A.; Levaray, M.; Paillon, C.; Durieux, E.D.H.; Pasqualini, V.; Agostini, S. Relationship between swimming capacities and morphological traits of fish larvae at settlement stage: A study of several coastal Mediterranean species. J. Fish Biol. 2019. [Google Scholar] [CrossRef]
- Leis, J.M. Ontogeny of behaviour in larvae of marine demersal fishes. Ichthyol. Res. 2010, 57, 325–342. [Google Scholar] [CrossRef]
- Faria, A.M.; Chícharo, M.A.; Gonçalves, E.J. Effects of starvation on swimming performance and body condition of pre-settlement Sparus aurata larvae. Aquat. Biol. 2011, 12, 281–289. [Google Scholar] [CrossRef]
- Erzini, K.; Gonçalves, J.M.S.; Bentes, L.; Lino, P.J.; Ribeiro, J. Catch composition, catch rates and size selectivity of three long-line methods in the Algarve (southern Portugal). Bol. Inst. Esp. Oceanogr. 1999, 15, 313–323. [Google Scholar]
- Veiga, P.; Ribeiro, J.; Gonçalves, J.M.S.; Erzini, K. Quantifying recreational shore angling catch and harvest in southern Portugal (north-east Atlantic Ocean): Implications for conservation and integrated fisheries management. J. Fish. Biol. 2010, 76, 2216–2237. [Google Scholar] [CrossRef]
- Pajuelo, J.G.; Lorenzo, J.M. Growth and age estimation of Diplodus sargus cadenati (Sparidae) off the Canary Islands. Fish. Res. 2002, 59, 93–100. [Google Scholar] [CrossRef]
- Ribeiro, J.; Bentes, L.; Coelho, R.; Goncalves, J.M.S.; Lino, P.G.; Monteiro, P.; Erzini, K. Seasonal, tidal and diurnal changes in fish assemblages in the Ria Formosa lagoon (Portugal). Estuar. Coast. Shelf Sci. 2006, 67, 461–474. [Google Scholar] [CrossRef]
- Faria, A.; Morais, P.; Chícharo, M.A. Ichthyoplankton dynamics in the Guadiana estuary and adjacent coastal area, South-East Portugal. Estuar. Coast. Shelf Sci. 2006, 70, 85–97. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Leboulleux, V. Microhabitat requirements for settlement of juvenile sparid fishes on Mediterranean rocky shores. Hydrobiologia 1995, 301, 309–320. [Google Scholar] [CrossRef]
- Di Franco, A.; Guidetti, P. Patterns of variability in early-life traits of fishes depend on spatial scale of analysis. Biol. Lett. 2011, 7, 454–456. [Google Scholar] [CrossRef][Green Version]
- Monteiro, C.C.; Lasserre, G.; Lam Hoi, T. Organisation spatiale des communautés ichtyologiques de la Lagune Ria Formosa (Portugal). Oceanol. Acta 1990, 13, 79–96. [Google Scholar]
- Planes, S.; Jouvenel, J.-Y.; Biagi, F.; Francour, P.; Harmelin-Vivien, M.; Macpherson, E.; Tunesi, L.; Galzin, R. Spatio-temporal variability in growth of juvenile sparid fishes from the Mediterranean littoral. J. Mar. Biol. Assoc. UK 1999, 79, 137–143. [Google Scholar] [CrossRef]
- Ortiz-Delgado, J.B.; Darias, M.J.; Cañavate, J.P.; Yúfera, M.; Sarasquete, C. Organogenesis of the digestive tract in the white seabream, Diplodus sargus. Histological and histochemical approaches. Histol. Histopathol. 2003, 18, 1155–1168. [Google Scholar] [CrossRef]
- Faria, A.; Ojanguren, A.; Fuiman, L.; Gonçalves, E. Ontogeny of critical swimming speed of wild-caught and laboratory-reared larvae of red drum (Scieanops ocellatus). Mar. Ecol. Prog. Ser. 2009, 384, 221–230. [Google Scholar] [CrossRef][Green Version]
- Brett, J.R. The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Res. Board. Can. 1964, 21, 1183–1226. [Google Scholar] [CrossRef]
- Pattrick, P.; Strydom, N.A. Swimming abilities of wild-caught, late-stage larvae of Diplodus capensis and Sarpa salpa (Pisces: Sparidae) from temperate South Africa. Estuar. Coast. Shelf Sci. 2009, 85, 547–554. [Google Scholar] [CrossRef]
- Silva, L.; Faria, A.M.; Teodósio, M.A.; Garrido, S. Ontogeny of swimming behaviour in sardine Sardina pilchardus larvae and effect of larval nutritional condition on critical speed. Mar. Ecol. Prog. Ser. 2014, 504, 287–300. [Google Scholar] [CrossRef]
- Barlow, G.W. Ethological units of behavior. In The Central Nervous System and Fish Behavior; Ingle, D.J., Ed.; University of Chicago Press: Chicago, IL, USA, 1968; pp. 217–232. [Google Scholar]
- Chícharo, M.A.; Chícharo, L.; Valdés, L.; López-Jamar, E.; Ré, P. Estimation of starvation and diel variation of the RNA/DNA ratios in field-caught Sardina pilchardus larvae off the north of Spain. Mar. Ecol. Prog. Ser. 1998, 164, 273–283. [Google Scholar] [CrossRef]
- Buckley, L.; Caldarone, E.; Ong, T.L. RNA-DNA ratio and other nucleic acid-based indicators for growth and condition of marine fishes. Hydrobiologia 1999, 401, 265–277. [Google Scholar] [CrossRef]
- Bulow, F.J. RNA–DNA ratios as indicators of recent growth rates of a fish. J. Fish. Res. Board Can. 1970, 27, 2343–2349. [Google Scholar] [CrossRef]
- Wagner, M.M.; Durbin, E.G.; Buckley, L.J. RNA/DNA ratios as indicators of nutritional condition in the copepod Calanus finmarchicus. Mar. Ecol. Prog. Ser. 1998, 162, 173–181. [Google Scholar] [CrossRef]
- Caldarone, E.M.; Wagner, M.; St Onge-Burns, J.; Buckley, L.J. Protocol and guide for estimating nucleic acids in larval fish using a fluorescence microplate reader. In Northeast Fisheries Science Center Reference Document 01–11; Woods Hole: Falmouth, MA, USA, 2001; p. 22. [Google Scholar]
- Caldarone, E.M.; Clemmesen, C.; Berdalet, E.; Miller, T.J.; Folkvord, A.; Holt, G.; Olivar, M.P.; Suthers, I.M. Intercalibration of four spectrofluorometric protocols for measuring RNA/DNA ratios in larval and juvenile fish. Limnol. Oceanogr. Methods 2006, 4, 153–163. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Clark, D.L.; Leis, J.M.; Hay, A.C.; Trnski, T. Swimming ontogeny of larvae of four temperate marine fishes. Mar. Ecol. Prog. Ser. 2005, 292, 287–300. [Google Scholar] [CrossRef]
- Leis, J.M.; McCormick, M.I. The biology, behaviour and ecology of the pelagic, larval stage of coral-reef fishes. In Coral Reef Fishes: New Insights into their Ecology; Sale, P.F., Ed.; Academic Press: San Diego, CA, USA, 2002; pp. 171–199. [Google Scholar]
- Sánchez, R.F.; Mason, E.; Relvas, P.; da Silva, A.J.; Peliz, A. On the inner-shelf circulation in the northern Gulf of Cádiz, southern Portuguese shelf. Deep Sea Res. II 2006, 53, 1198–1218. [Google Scholar] [CrossRef]
- Zuo, J.; Du, L.; Peliz, A.; Santos, A.M.P.; Yu, Y. The characteristics of near-surface velocity during the upwelling season on the northern Portugal shelf. J. Ocean Univ. China 2007, 6, 213–225. [Google Scholar] [CrossRef]
- Moyano, M.; Illing, B.; Peschutter, P.; Huebert, K.B.; Peck, M.A. Thermal impacts on the growth, development and ontogeny of critical swimming speed in Atlantic herring larvae. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 197, 23–34. [Google Scholar] [CrossRef]
- Guan, L.; Snelgrove, P.V.R.; Gamperl, A.K. Ontogentic changes in the critical swimming speed of Gadus morhua (Atlantic cod) and Myoxocephalus scorpius (shorthorn sculpin) larvae and the role of temperature. J. Exp. Mar. Biol. Ecol. 2008, 360, 31–38. [Google Scholar] [CrossRef]
- Faria, A.; Gonçalves, E. Ontogeny of swimming behaviour of two temperate clingfishes, Lepadogaster lepadogaster and Lepadogaster purpurea (Gobiesocidae). Mar. Ecol. Prog. Ser. 2010, 414, 237–248. [Google Scholar] [CrossRef][Green Version]
- Houde, E.D. Fish early life dynamics and recruitment variability. Am. Fish. Soc. Symp. 1987, 2, 17–29. [Google Scholar]
- Olivar, M.P.; Diaz, M.V.; Chícharo, M.A. Tissue effect on RNA: DNA ratios of marine fish larvae. Sci. Mar. 2009, 73, 171–182. [Google Scholar] [CrossRef]
- Houlihan, D.F.; Mathers, E.M.; Foster, A. Biochemical correlates of growth rate in fish. In Fish Ecophysiology; Rankin, J.C., Jensen, F.B., Eds.; Chapman and Hall: London, UK, 1993; pp. 45–71. [Google Scholar]
- Houde, E.D. Comparative growth, mortality, and energetics of marine fish larvae: Temperature and implied latitudinal effects. Fish. Bull. 1989, 87, 471–495. [Google Scholar]
- Green, B.S.; Fisher, R. Temperature influences swimming speed, growth and larval duration in coral reef fish larvae. J. Exp. Mar. Bio. Ecol. 2004, 299, 115–132. [Google Scholar] [CrossRef]
- Fuiman, L.; Batty, R.S. What a drag it is getting cold: Partitioning the physical and physiological effects of temperature on fish swimming. J. Exp. Biol. 1997, 200, 1745–1755. [Google Scholar] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Climate change 2007: The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Munday, P.L.; Jones, G.P.; Pratchett, M.S.; Williams, A.J. Climate change and the future for coral reef fishes. Fish Fish. 2008, 9, 261–285. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
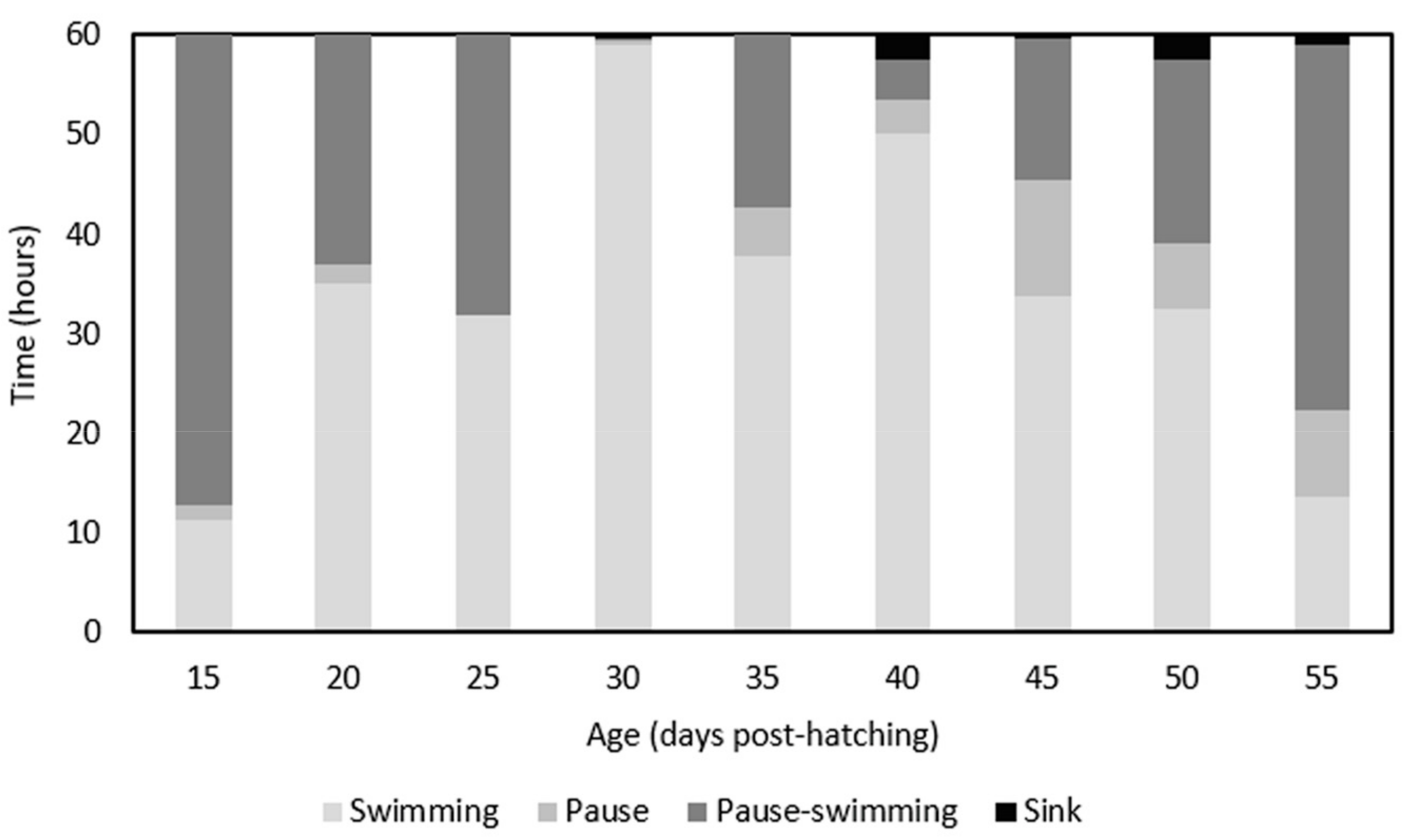{kind=link}
| Experiment | 15 DPH | 20 DPH | 25 DPH | 30 DPH | 35 DPH | 40 DPH | 45 DPH | 50 DPH | 55 DPH |
|---|---|---|---|---|---|---|---|---|---|
| UCrit | 15 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 |
| Endurance flow speed (cm s−1) | 8 1.5 | - | 8 5.0 | - | 8 6.0 | - | 8 10.0 | - | - |
| MAPs | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Experiment | Correlation with | ρ | p-value |
|---|---|---|---|
| Ucrit | DPH | 0.85 | <0.001 |
| TL | 0.83 | <0.001 | |
| RNA | −0.61 | <0.001 | |
| DNA | −0.58 | <0.001 | |
| sRD | −0.37 | <0.001 | |
| resRNA | −0.16 | 0.061 | |
| Proteins | −0.55 | <0.001 | |
| Endurance | DPH | 0.93 | <0.001 |
| TL | 0.88 | <0.001 | |
| RNA | −0.56 | 0.001 | |
| DNA | −0.38 | 0.033 | |
| sRD | −0.16 | 0.388 | |
| resRNA | −0.08 | 0.668 | |
| Proteins | −0.89 | <0.001 |
| Order | Family | Species | Size (mm) | Ucrit (cm s−1) | Ucrit (bl s−1) | Endurance (km) | Reference |
|---|---|---|---|---|---|---|---|
| Atheriniformes | Atheriniidae | Atherina presbyter | 6.6–21.0 | 3.6–18.7 | 5.2–17.6 | – | [27] |
| Clupeiformes | Clupeidae | Sardina pilchardus | 7.9–23.4 | 1.6–9.5 | 1.8–6.9 | – | [47] |
| Clupea harengus | 8.0–22.0 | 0.0–9.2 | 0.0–4.8 | – | [60] | ||
| Gadiformes | Gadidae | Gadus morhua | – | 1.2–9.7 | 2.2–6.9 | – | [61] |
| Gobiesociformes | Gobiesocidae | Lepadogaster lepadogaster | 5.1–10.9 | 1.0–9.4 | 1.5–9.6 | – | [62] |
| Mugiliformes | Mugilidae | Mugilidae sp. | 9.8–13.4 | 21.1–37.4 | 20.5–30.4 | – | [30] |
| Perciformes | Apogonidae | Apogon imberbis | – | 26.3 ± 1.2 | – | – | [31] |
| Carangidae | Carangidae sp. | – | 19.4 ± 2.9 | – | – | [31] | |
| Percichthyidae | Macquaria novemaculeata | 4.0–10.4 | 0–1.5 | – | 0–12.5 | [56] | |
| Pomacentridae | Chromis chromis | 8.3–12.4 | 19.0–35.6 | 17.0–30.4 | – | [30] | |
| Sciaenidae | Argyrosomus japonicus | 3.0–14.0 | 4.0–16.6 | – | 0–22.4 | [56] | |
| Scianops ocellatus | 3.0–19.1 | 1.1–20.5 | 3.1–20.4 | – | [44] | ||
| Sparidae | Acanthopagrus australis | 4.9–11.1 | 2.0–27.0 | – | – | [56] | |
| Boops boops | 8.2–13.8 | 5.3–25.3 | 4.5–22.6 | – | [30] | ||
| Dentex dentex | – | 17.8 | – | – | [30] | ||
| Diplodus annularis | 7.8–11.6 | 6.1–20.9 | 7.0–23.2 | – | [30] | ||
| Diplodus capensis | 8.9–16.0 | 2.8–35.2 | max 26.1 | 0.2–32.4 | [46] | ||
| Diplodus sargus | – | 10.8 ± 2.8 | – | – | [31] | ||
| 1.3–29.5 | 1.1–23.0 | 2.0–27.2 | 0.0–86.5 | this study | |||
| Oblada melanura | – | 21.3 ± 3.1 | – | – | [31] | ||
| Pagellus acarne | – | 21.3 ± 5.5 | – | – | [31] | ||
| Pagrus auratus | 2.2–10.4 | 2.0–27.3 | – | 4.0–9.9 | [56] | ||
| Sarpa salpa | 12.1–21.3 | 4.8–33.4 | max 15.7 | 0.1–64.8 | [46] | ||
| Sparus aurata | 6.2–14.1 | 3.0–19.3 | 4.5–20.9 | 10.2–23.9 | [33] | ||
| Spicara smaris | 9.0–13.9 | 7.2–26.6 | 5.5–22.4 | – | [30] | ||
| Spondyliosoma cantharus | 11.7–17.3 | 12.0–26.6 | 9.3–17.5 | – | [30] | ||
| Pleuronectiformes | Pleuronectidae | Pleuronectes platessa | 7.0–13.0 | 1.0–3.9 | 1.3–3.5 | – | [29] |
| Soleidae | Solea senegalensis | 3.5–7.5 | 0.4–5.0 | 0.2–11.9 | – | [19] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baptista, V.; Morais, P.; Cruz, J.; Castanho, S.; Ribeiro, L.; Pousão-Ferreira, P.; Leitão, F.; Wolanski, E.; Teodósio, M.A. Swimming Abilities of Temperate Pelagic Fish Larvae Prove that They May Control Their Dispersion in Coastal Areas. Diversity 2019, 11, 185. https://doi.org/10.3390/d11100185
Baptista V, Morais P, Cruz J, Castanho S, Ribeiro L, Pousão-Ferreira P, Leitão F, Wolanski E, Teodósio MA. Swimming Abilities of Temperate Pelagic Fish Larvae Prove that They May Control Their Dispersion in Coastal Areas. Diversity. 2019; 11(10):185. https://doi.org/10.3390/d11100185
Chicago/Turabian StyleBaptista, Vânia, Pedro Morais, Joana Cruz, Sara Castanho, Laura Ribeiro, Pedro Pousão-Ferreira, Francisco Leitão, Eric Wolanski, and Maria Alexandra Teodósio. 2019. "Swimming Abilities of Temperate Pelagic Fish Larvae Prove that They May Control Their Dispersion in Coastal Areas" Diversity 11, no. 10: 185. https://doi.org/10.3390/d11100185
APA StyleBaptista, V., Morais, P., Cruz, J., Castanho, S., Ribeiro, L., Pousão-Ferreira, P., Leitão, F., Wolanski, E., & Teodósio, M. A. (2019). Swimming Abilities of Temperate Pelagic Fish Larvae Prove that They May Control Their Dispersion in Coastal Areas. Diversity, 11(10), 185. https://doi.org/10.3390/d11100185