Subterranean Invasion by Gapped Ringed Crayfish: Effectiveness of a Removal Effort and Barrier Installation


Abstract
1. Introduction
2. Materials and Methods
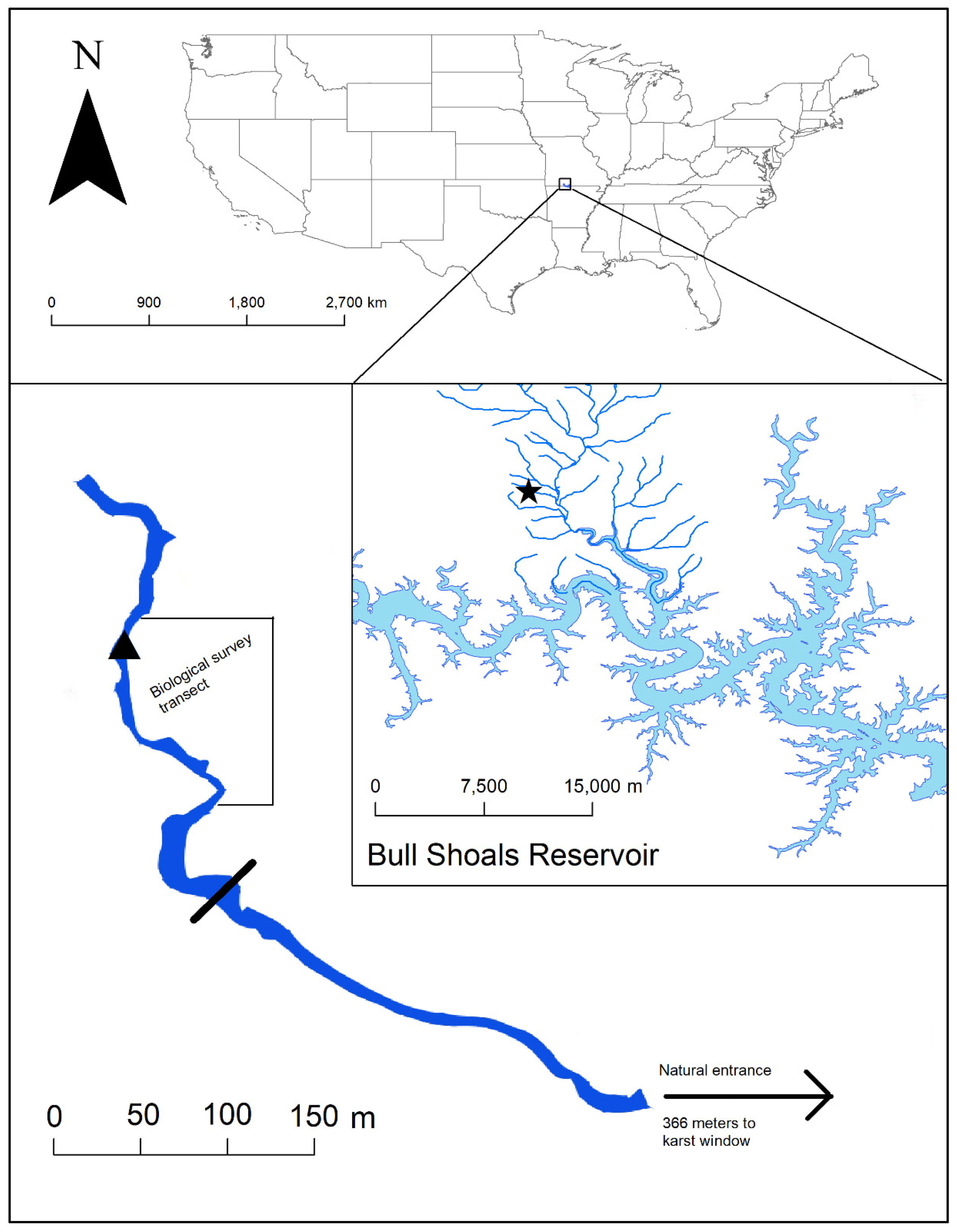
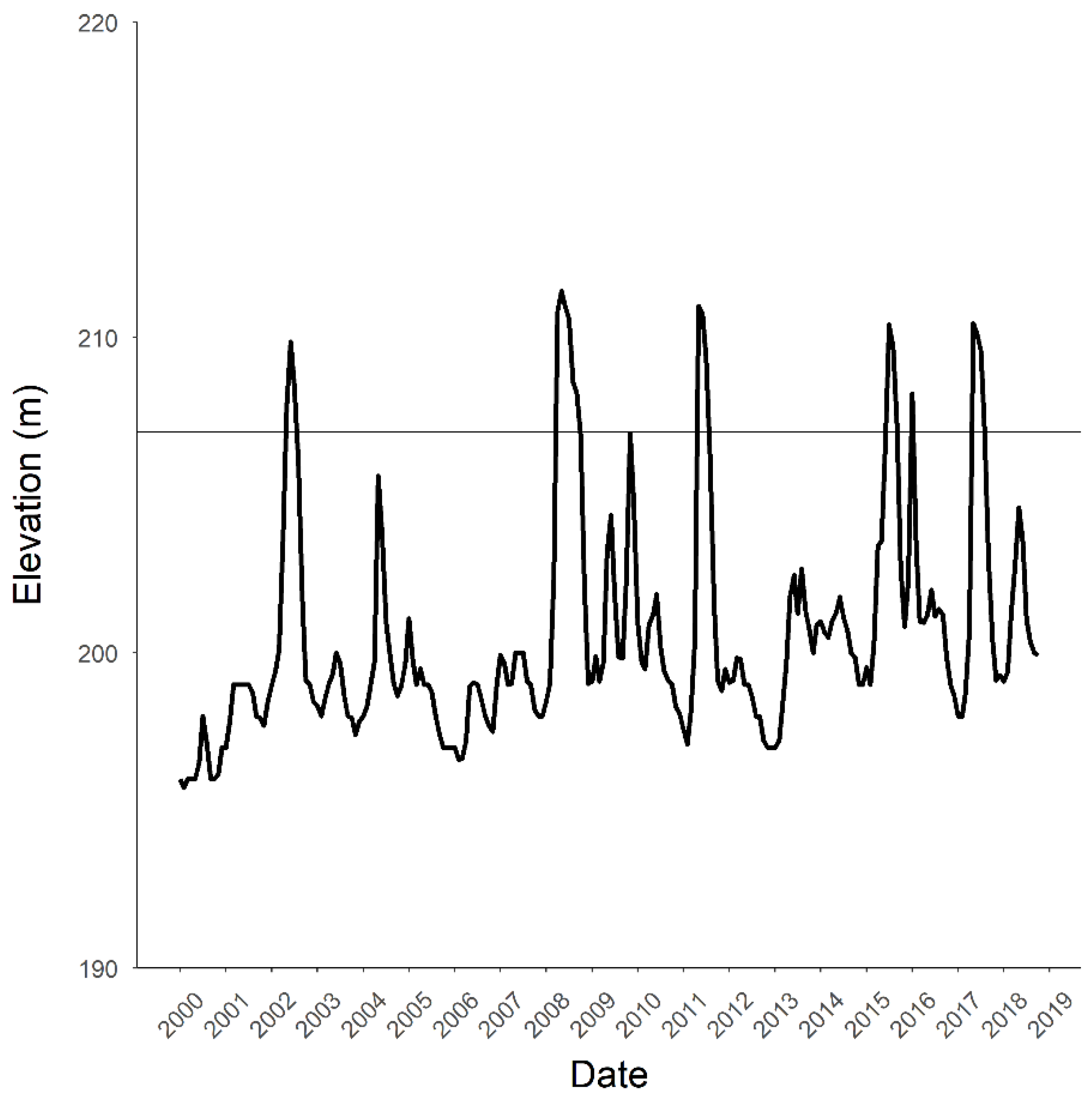
2.1. Study Area
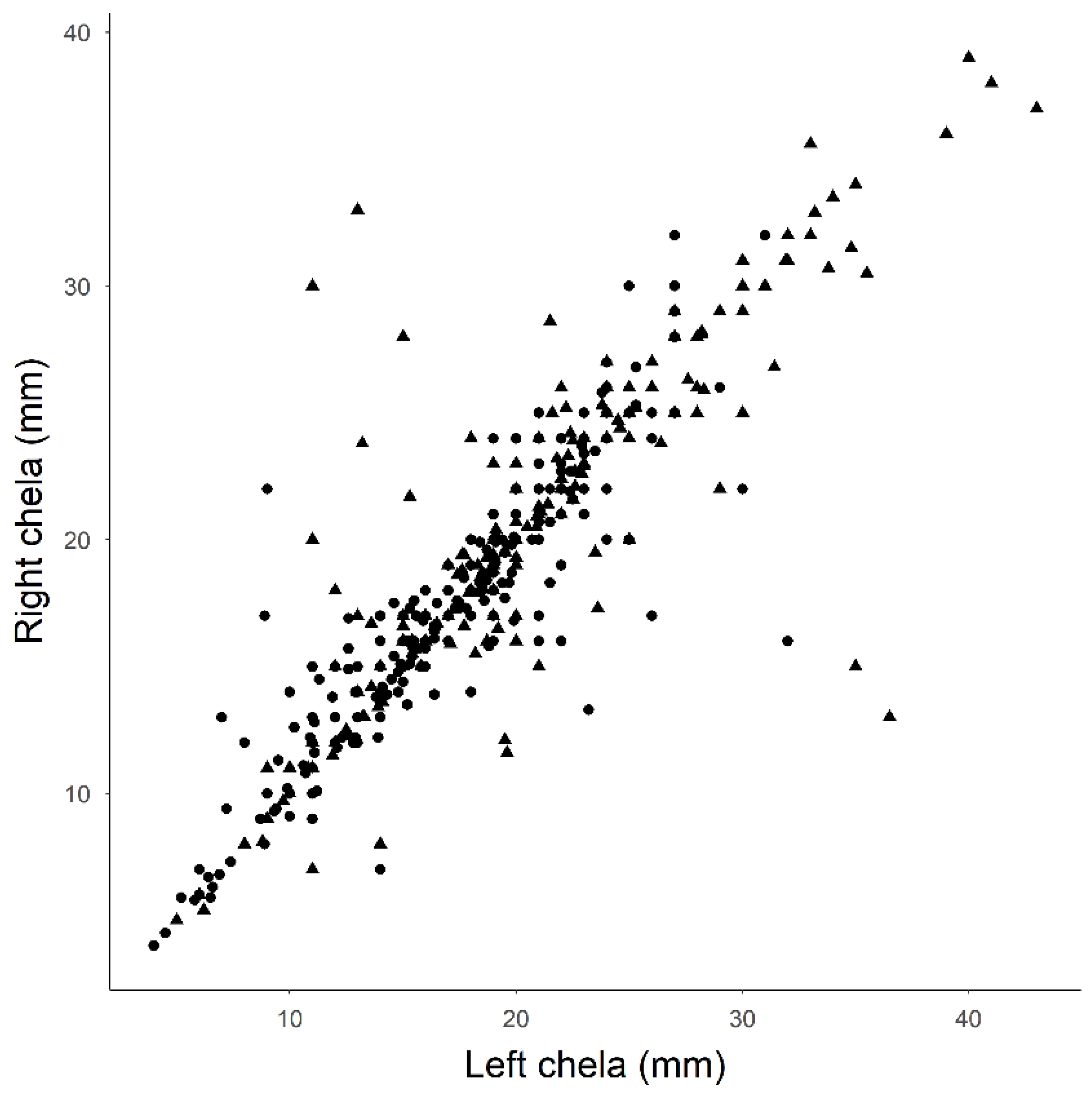
2.2. Crayfish Surveys and Morphometric Traits
2.3. Crayfish Removal
2.4. Statistical Analysis
2.5. Age Comparison of Surface and Subterranean Populations
2.6. Barrier Installations
3. Results
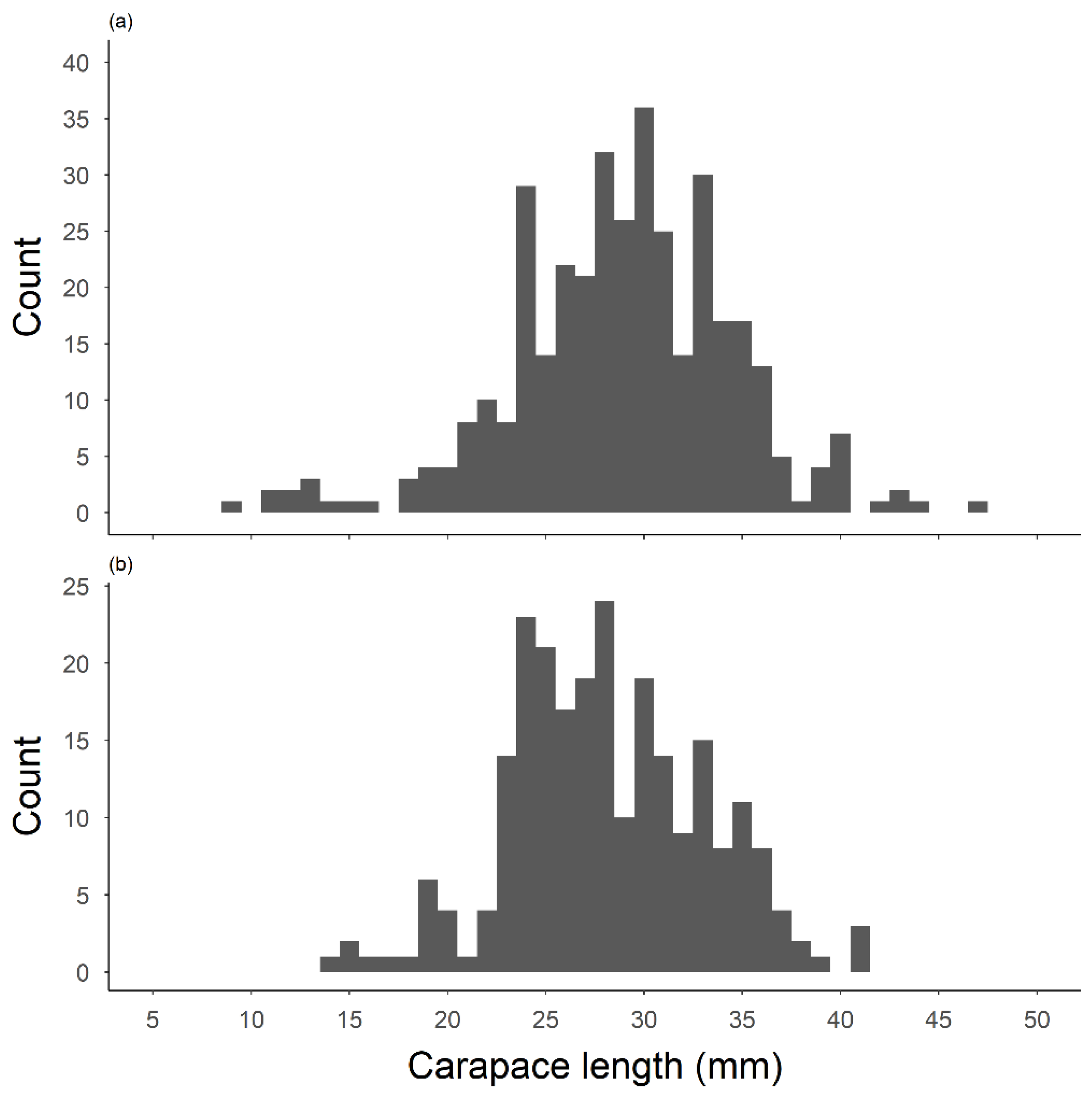
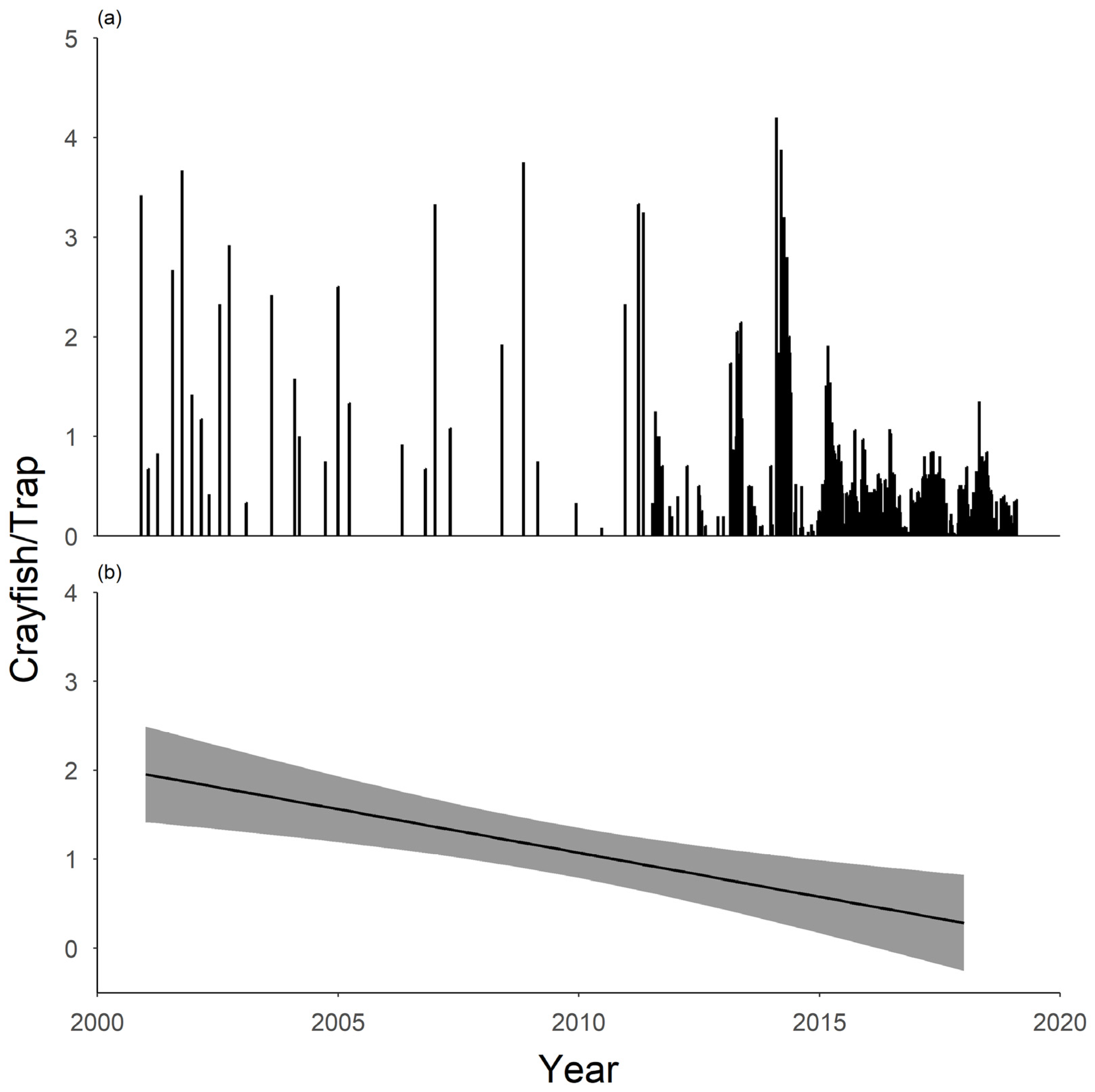
3.1. Crayfish Surveys and Morphometric Traits
3.2. Crayfish Removal
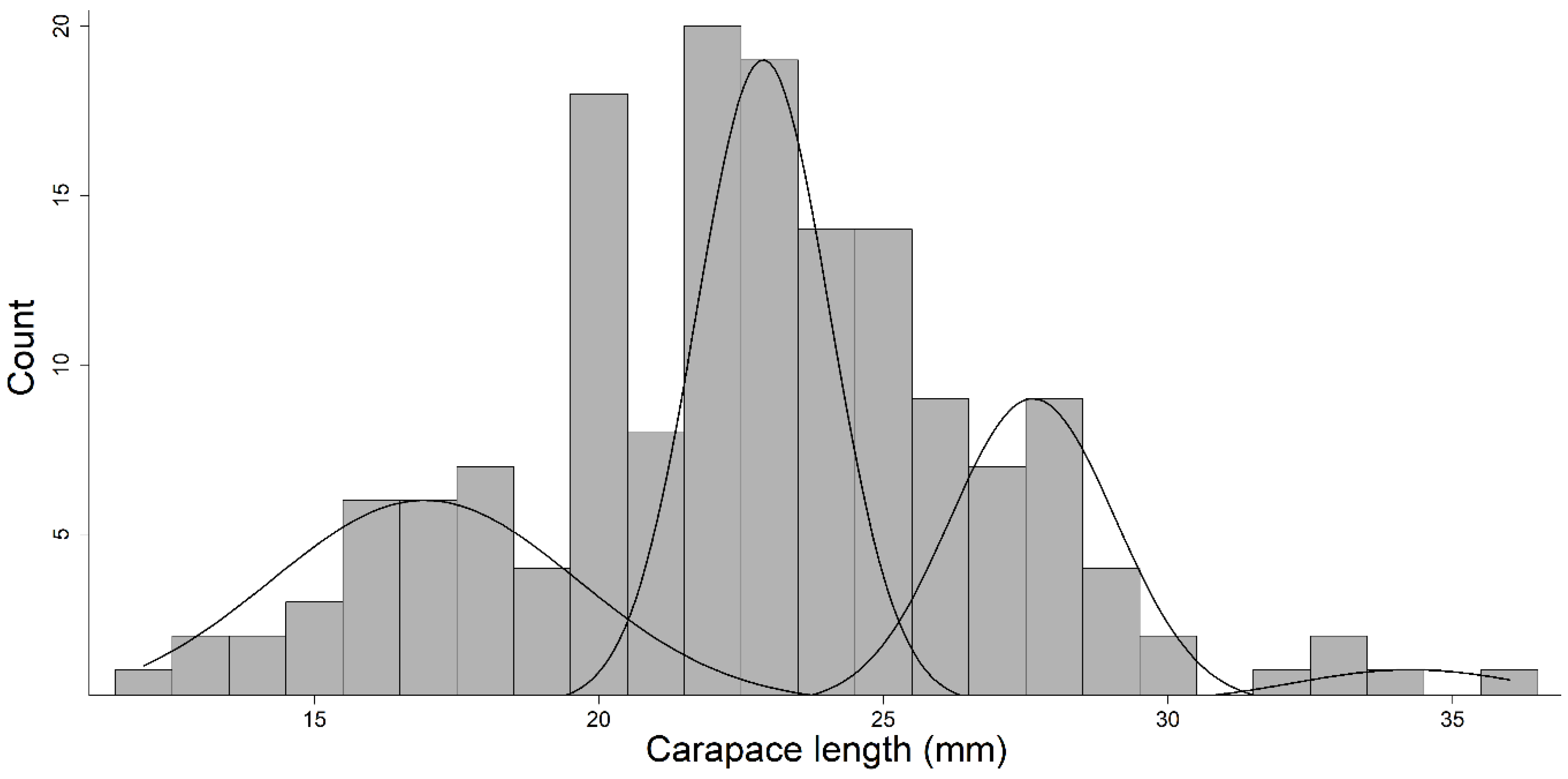
3.3. Age Comparison of Surface and Subterranean Populations
3.4. Barrier Installations
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S., III; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Schuster, G.A.; Cooper, J.E.; DiStefano, R.J.; Eversole, A.G.; Hamr, P.; Hobbs, H.H., III; Robison, H.W.; Skelton, C.E.; Thoma, R.F. A reassessment of the conservation status of crayfishes in the United States and Canada after 10+ years of increased awareness. Fisheries 2007, 32, 372–389. [Google Scholar] [CrossRef]
- Momot, W.T. Redefining the role of crayfish in aquatic ecosystems. Rev. Fish. Sci. 1995, 3, 33–63. [Google Scholar] [CrossRef]
- Carreira, B.M.; Dias, M.P.; Rebelo, R. How consumption and fragmentation of macrophytes by the invasive crayfish Procambarus clarkii shape the macrophyte communities of temporary ponds. Hydrobiologia 2014, 721, 89–98. [Google Scholar] [CrossRef]
- Mathers, K.L.; Chadd, R.P.; Dunbar, M.J.; Extence, C.A.; Reeds, J.; Rice, S.P.; Wood, P.J. The long-term effects of invasive signal crayfish (Pacifastacus leniusculus) on instream macroinvertebrate communities. Sci. Total Environ. 2016, 556, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Ilhéu, M.; Bernado, J.M.; Fernandes, S. Predation of invasive crayfish on aquatic vertebrates: The effect of Procambarus clarkii on fish assemblages in Mediterranean temporary streams. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Invading Nature—Springer Series in Invasion Ecology; Gherardi, F., Ed.; Springer: Dordrecht, The Netherlands, 2007; Volume 2, pp. 543–558. ISBN 978-1-4020-6028-1. [Google Scholar]
- Nunes, A.L.; Richter-Boix, A.; Laurila, A.; Rebelo, R. Do anuran larvae respond behaviourally to chemical cues from an invasive crayfish predator? A community-wide study. Oecologia 2014, 171, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.M.; Taylor, C.A.; Holdich, D.M.; Skurdal, J. Nonindigenous crayfishes threaten North American freshwater biodiversity: Lessons from Europe. Fisheries 2000, 25, 7–20. [Google Scholar] [CrossRef]
- Mazza, G.; Reboleira, A.S.P.S.; Goncalves, F.; Aquiloni, L.; Inghilesi, A.F.; Spigoli, D.; Stoch, F.; Taiti, S.; Gherardi, F.; Tricarico, E. A new threat to groundwater ecosystems: First occurrences of the invasive crayfish Procambarus clarkii (Girard, 1852) in European caves. J. Caves Karst Stud. 2014, 76, 62–65. [Google Scholar] [CrossRef]
- Gibert, J.; Culver, D.C.; Dole-Olivier, M.; Malard, F.; Christman, M.C.; Deharveng, L. Assessing and conserving groundwater biodiversity: Synthesis and perspectives. Freshw. Biol. 2009, 54, 930–941. [Google Scholar] [CrossRef]
- Venarsky, M.P.; Huryn, A.D.; Benstead, J.P. Re-examining extreme longevity of the cave crayfish Orconectes australis using new mark–recapture data: A lesson on the limitations of iterative size-at-age models. Freshw. Biol. 2012, 57, 1471–1481. [Google Scholar] [CrossRef]
- Notenboom, J.; Plénet, S.; Turquin, M.J. Groundwater contamination and its impact on groundwater animals and ecosystems. In Groundwater Ecology; Gibert, J., Danielopol, D.L., Stanford, J.A., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 477–504. ISBN 978-0-0805-0762-0. [Google Scholar]
- Morris, B.L.; Lawrence, A.R.L.; Chilton, P.J.C.; Adams, B.; Calow, R.C.; Klinck, B.A. Groundwater and Its Susceptibility to Degradation: A Global Assessment of the Problem and Options for Management; Early Warning and Assessment Report Series, RS. 03-3; United Nations Environment Programme: Nairobi, Kenya, 2003; ISBN 92-807-2297-2. [Google Scholar]
- Daniels, R.A.; Murphy, D.C.; Klemens, M.W. Orconectes neglectus is established in the northeast. Northeast. Nat. 2001, 8, 93–100. [Google Scholar] [CrossRef]
- Pearl, C.A.; Adams, M.J.; McCreary, B. Habitat and co-occurrence of native and invasive crayfish in the Pacific Northwest, USA. Aquat. Invasions 2013, 8, 171–184. [Google Scholar] [CrossRef]
- Imhoff, E.M.; Moore, M.J.; DiStefano, R.J. Introduced alien ringed crayfish (Orconectes neglectus neglectus [Faxon, 1885]) threaten imperiled coldwater crayfish (Orconectes eupunctus Williams, 1952) in the Eleven Point River drainage, Missouri, USA. Aquat. Invasions 2012, 7, 129–134. [Google Scholar] [CrossRef]
- Graening, G.O.; Fenolio, D.B.; Slay, M.E. Cave Life of Oklahoma and Arkansas: Exploration and Conservation of Subterranean Biodiversity; University of Oklahoma Press: Norman, OK, USA, 2011; ISBN 978-0-8061-4424-5. [Google Scholar]
- Elliot, W.R. Zoogeography and biodiversity of Missouri caves and karst. J. Caves Karst Stud. 2007, 69, 135–162. [Google Scholar]
- Elliot, W.R. A Guide to Missouri’s Cave Life: 70 Cave Species Brought to Light; Missouri Department of Conservation: Jefferson City, MO, USA, 2003. Available online: http://www.mospeleo.org/sites/default/files/Attachements/A%20Guide%20to%20Missouri%20Cave%20Life.pdf (accessed on 26 September 2018).
- Hein, C.L.; Zanden, M.J.V.; Magnuson, J.J. Intensive trapping and increased fish predation cause massive population decline of an invasive crayfish. Freshw. Biol. 2007, 52, 1134–1146. [Google Scholar] [CrossRef]
- Woods, A.J.; Omernik, J.M.; Butler, D.R.; Ford, J.G.; Henley, J.E.; Hoagland, B.W.; Arndt, D.S.; Morgan, B.C. Ecoregions of Oklahoma. U.S. Geological Survey, map scale 1:1,250,000; U.S. Geological Survey: Reston, VA, USA, 2005. Available online: https://www.epa.gov/eco-research/ecoregion-download-files-state-region-6 (accessed on 26 September 2018).
- Dillman, C.B. Molecular Systematics, Biogeography, and Phylogeography of North American Freshwater Crayfishes (Decapoda: Cambaridae): With Emphasis on the Genera Cambarus and Orconectes. Ph.D. Thesis, Saint Louis University, Saint Louis, MO, USA, 2008. [Google Scholar]
- Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; Listing the Tumbling Creek Cavesnail as Endangered; Fish and Wildlife Service: Columbia, SC, USA, 2001. Available online: www.gpo.gov/fdsys/search/citation.result.FR.action?federalRegister.volume=2001&federalRegister.page=66803&publication=FR (accessed on 26 September 2018).
- Missouri Department of Conservation. Missouri Species and Communities of Conservation Concern Checklist; Missouri Department of Conservation: Jefferson City, MO, USA, 2018. Available online: https://nature.mdc.mo.gov/sites/default/files/downloads/2018_SOCC.pdf (accessed on 26 September 2018).
- Phillips, J.G.; Fenolio, D.B.; Emel, S.L.; Bonett, R.M. Hydrologic and geologic history of the Ozark Plateau drive phylogenomic patterns in a cave-obligate salamander. J. Biogeogr. 2017, 440, 2463–2474. [Google Scholar] [CrossRef]
- Jurcak, A.M.; Lahman, S.E.; Wofford, S.J.; Moore, P.A. Behavior of Crayfish. In Biology and Ecology of Crayfish; Longshaw, M., Stebbing, B., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 117–131. ISBN 978-1-4987-6732-3. [Google Scholar]
- France, R.; Holmes, J.; Lynch, A. Use of size–frequency data to estimate the age composition of crayfish populations. Can. J. Fish. Aquat. Sci. 1991, 48, 2324–2332. [Google Scholar] [CrossRef]
- Kilada, R.; Sainte-Marie, B.; Rochette, R.; Davis, N.; Vanier, C.; Campana, S. Direct determination of age in shrimps, crabs, and lobsters. Can. J. Fish. Aquat. Sci. 2012, 69, 1728–1733. [Google Scholar] [CrossRef]
- Mouser, J.M.; Glover, J.; Brewer, S.K. Comparing age estimates of freshwater crayfish using gastric mill bands and length-frequency histograms. Unpublished: Manuscript in preparation.
- Bhattacharya, C.G. A simple method of resolution of a distribution into Gaussian components. Biometrics 1967, 23, 115–135. [Google Scholar] [CrossRef] [PubMed]
- Mildenberger, T.K.; Taylor, M.H.; Wolff, M. TropFishR: An R package for fisheries analysis with length-frequency data. Methods Ecol. Evol. 2017, 8, 1520–1527. [Google Scholar] [CrossRef]
- Gayanilo, F.C.; Sparre, P.; Pauly, D. FAO-ICLARM Stock Assessment Tools II: User’s Guide; Food and Agriculture Organization: Rome, Italy, 2005; ISBN 92-5-105300-6. [Google Scholar]
- Chu, C.; Mandrak, N.E.; Minns, C.K. Potential impacts of climate change on the distributions of several common and rare freshwater fishes in Canada. Divers. Distrib. 2005, 11, 299–310. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.N.; Bryga, H. Life-history evolution in guppies (Poecilia reticulata): 1. Phenotypic and genetic changes in an introduction experiment. Evolution 1987, 41, 1370–1385. [Google Scholar] [CrossRef] [PubMed]
- Huppop, K. Adaptation to low food. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Eds.; Academic Press: Waltham, MA, USA, 2012; pp. 99–102. ISBN 978-0-123-83832-2. [Google Scholar]
- Pflieger, W.L. The Crayfishes of Missouri; Missouri Department of Conservation: Jefferson City, MO, USA, 1996; ISBN 1-887247-05-X.
- Stebbing, P.; Longshaw, M.; Scott, A. Review of methods for the management of nonindigenous crayfish, with particular reference to Great Britain. Ethol. Ecol. Evol. 2014, 26, 204–231. [Google Scholar] [CrossRef]
- Reynolds, J.D. Growth and reproduction. In Biology of Freshwater Crayfish; Holdich, D.M., Ed.; Blackwell Science: Oxford, UK, 2002; pp. 152–191. ISBN 978-0-632-05431-2. [Google Scholar]
- Ashley, D.C. Missouri cave biology. In Hitchhiker’s Guide to Missouri Caving. 2015 NSS Convention Guidebook; Slais, D., Ed.; World Press: Fenton, MO, USA, 2015; pp. 67–70. [Google Scholar]
- Fenolio, D.; Niemiller, M.L.; Soares, D.; Slay, M.; Harris, A.; Harris, N. Subterranean reproduction of the Ringed Crayfish, Orconectes neglectus Faxon 1885 (Astacoidea: Cambaridae) within an Ozark Highlands cave in Oklahoma, USA. Speleobiol. Notes 2013, 5, 43–46. [Google Scholar] [CrossRef]
- Fenolio, D.B.; Graening, G.O.; Collier, B.A.; Stout, J.F. Coprophagy in a cave-adapted salamander; the importance of bat guano examined through nutritional and stable isotope analyses. Proc. R. Soc. B 2006, 273, 439–443. [Google Scholar] [CrossRef]
- Humphreys, W.F. Experimental re-establishment of pulse-driven populations in a terrestrial troglobite community. J. Anim. Ecol. 1991, 60, 609–623. [Google Scholar] [CrossRef]
- Poulson, T.L. Food sources. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Eds.; Academic Press: Waltham, MA, USA, 2012; pp. 255–264. ISBN 978-0-123-83832-2. [Google Scholar]
- Kats, L.B.; Bucciarelli, G.; Vandergon, T.L.; Honeycutt, R.L.; Mattiasen, E.; Sanders, A.; Riley, S.P.D.; Kerby, J.L.; Fisher, R.N. Effects of natural flooding and manual trapping on the facilitation of invasive crayfish-native amphibian coexistence in a semi-arid perennial stream. J. Arid Environ. 2013, 98, 109–112. [Google Scholar] [CrossRef]
- Kerby, J.L.; Riley, S.P.D.; Kats, LB.; Wilson, P. Barriers and flow as limiting factors in the spread of an invasive crayfish (Procambarus clarkii) in southern California streams. Biol. Conserv. 2005, 126, 402–409. [Google Scholar] [CrossRef]
- Ellis, L.E.; Jones, N.E. Longitudinal trends in regulated rivers: A review and synthesis within the context of the serial discontinuity concept. Environ. Rev. 2013, 21, 136–148. [Google Scholar] [CrossRef]
- Wilson, P.N. Movement patterns of the cave salamander (Eurycea lucifuga) in Sauerkraut Cave, Kentucky. Honors Thesis, University of Louisville, Louisville, KY, USA, 2016. Available online: http://doi.org/10.18297/honors/121 (accessed on 2 December 2018).
- Moore, M.J.; DiStefano, R.J.; Larson, E.R. An assessment of life-history studies for USA and Canadian crayfishes: Identifying biases and knowledge gaps to improve conservation and management. Freshw. Sci. 2013, 32, 1276–1287. [Google Scholar] [CrossRef]
- Larson, E.R.; Olden, J.D. Latent extinction and invasion risk of crayfishes in the Southeastern United States. Conserv. Biol. 2010, 24, 1099–1110. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | CL range (mm) | Mean (mm) | SI |
|---|---|---|---|
| 0 | 12–23 | 16.9 ± 2.7 | 3.9 |
| 1 | 21–26 | 22.9 ± 1.2 | 17.2 |
| 2 | 24–31 | 27.6 ± 1.5 | 10.8 |
| 3 | 31–36 | 34.3 ± 2.1 | NA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouser, J.; Ashley, D.C.; Aley, T.; Brewer, S.K. Subterranean Invasion by Gapped Ringed Crayfish: Effectiveness of a Removal Effort and Barrier Installation. Diversity 2019, 11, 3. https://doi.org/10.3390/d11010003
Mouser J, Ashley DC, Aley T, Brewer SK. Subterranean Invasion by Gapped Ringed Crayfish: Effectiveness of a Removal Effort and Barrier Installation. Diversity. 2019; 11(1):3. https://doi.org/10.3390/d11010003
Chicago/Turabian StyleMouser, Joshua, David C. Ashley, Tom Aley, and Shannon K. Brewer. 2019. "Subterranean Invasion by Gapped Ringed Crayfish: Effectiveness of a Removal Effort and Barrier Installation" Diversity 11, no. 1: 3. https://doi.org/10.3390/d11010003
APA StyleMouser, J., Ashley, D. C., Aley, T., & Brewer, S. K. (2019). Subterranean Invasion by Gapped Ringed Crayfish: Effectiveness of a Removal Effort and Barrier Installation. Diversity, 11(1), 3. https://doi.org/10.3390/d11010003