Planktotrophic Brachiopod Larvae from the Pacific and Caribbean of Panama

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Sequencing
2.3. Sequence Analysis
3. Results
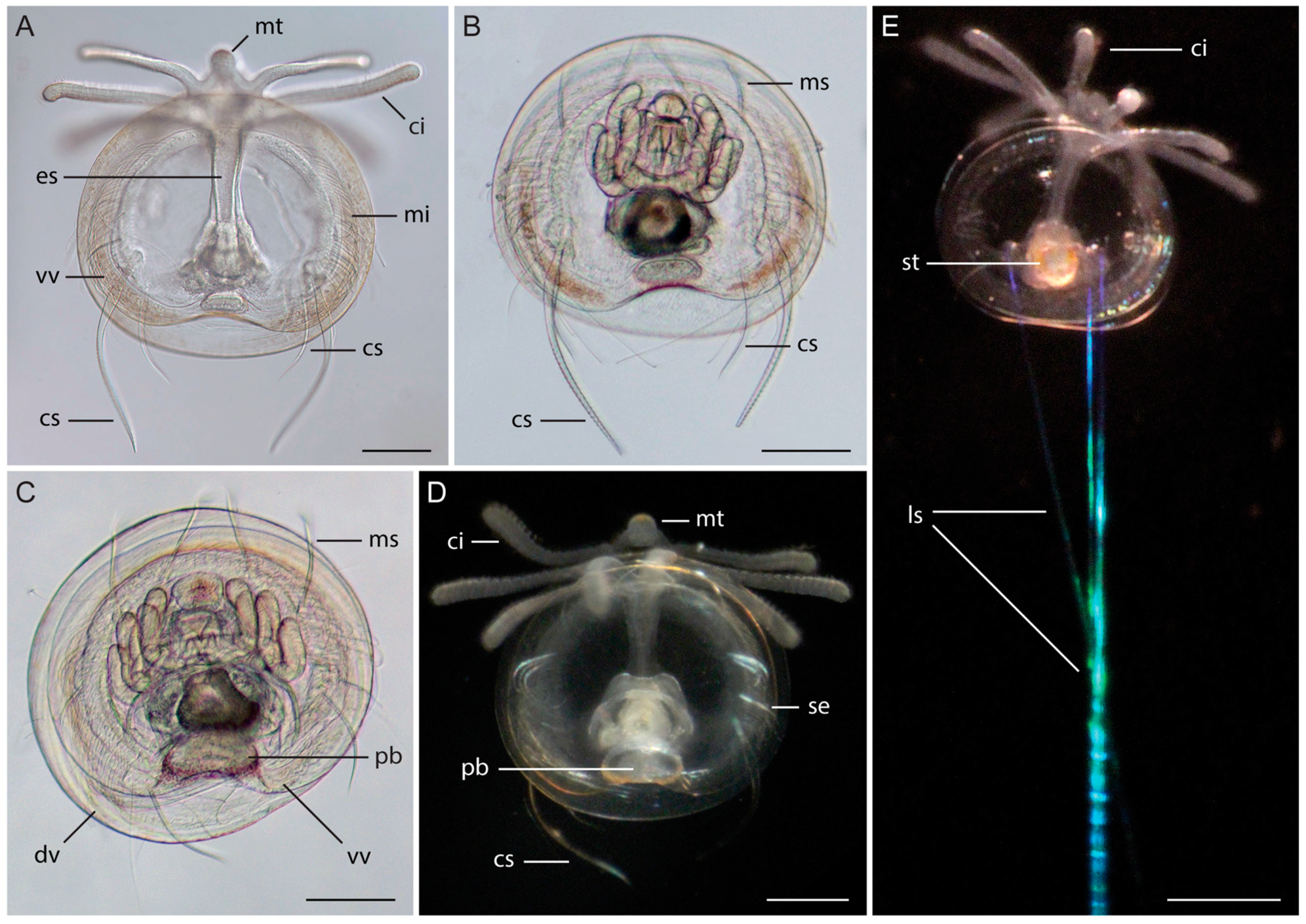
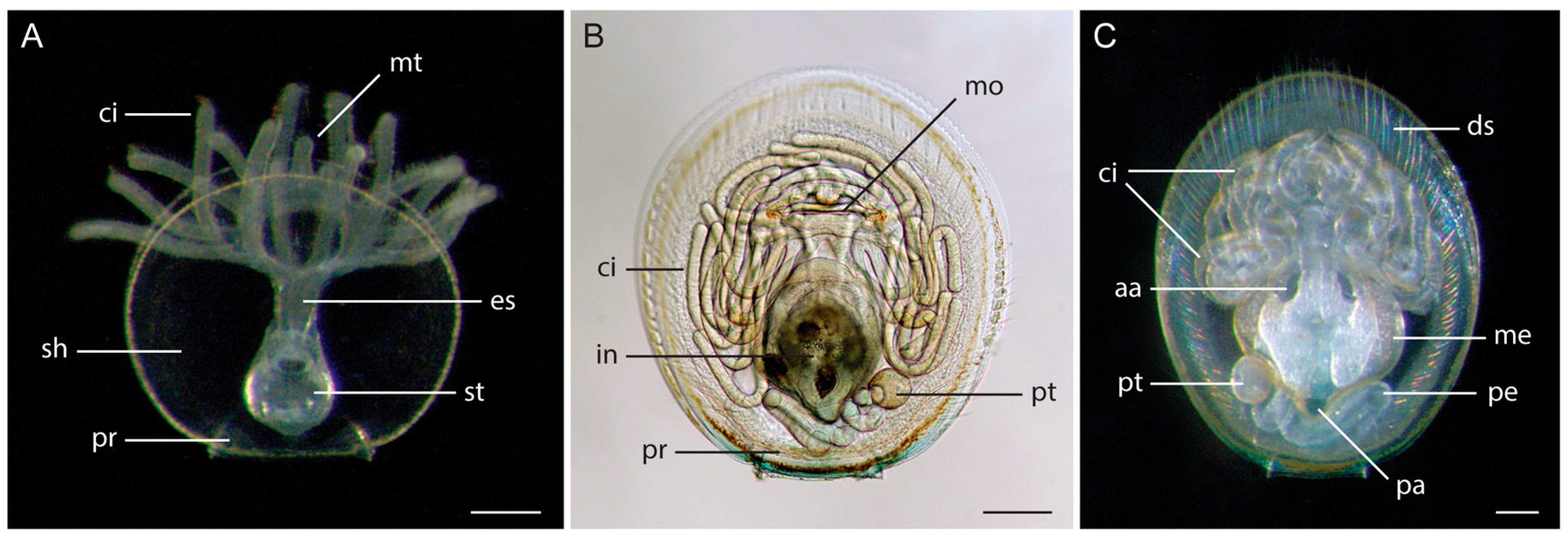
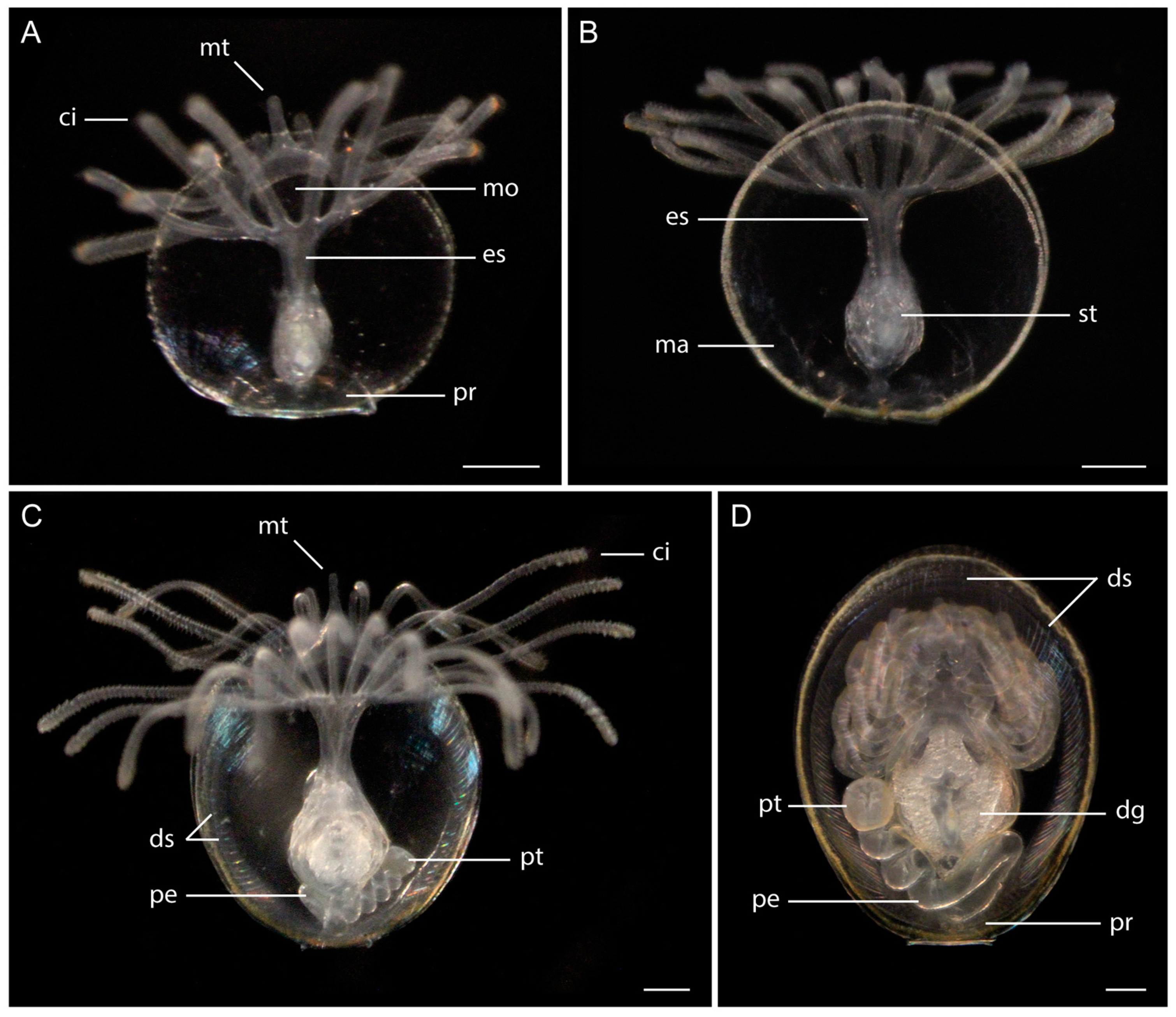
3.1. Morphology of Discinid Larvae
3.2. Morphology of Glottidia Larvae
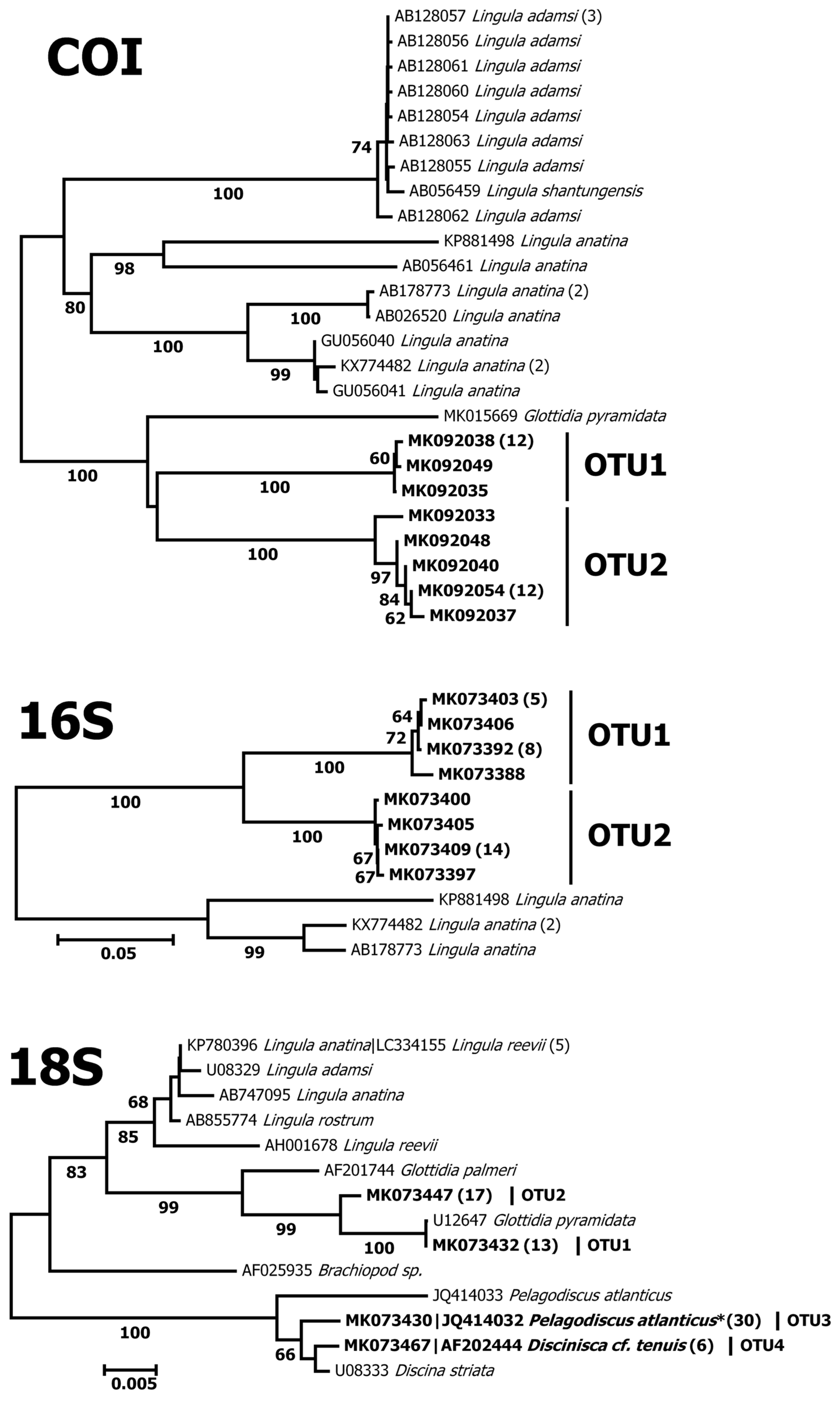
3.3. DNA Barcoding
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pennington, J.T.; Stricker, S.A. Phylum Brachiopoda. In Atlas of Marine Invertebrate Larval Forms; Young, C.M., Sewell, M.A., Rice, M.E., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 441–461. [Google Scholar]
- Yatsu, N. On the development of Lingula anatina. J. Coll. Sci. Imp. Univ. Tokyo 1902, 17, 1–112. [Google Scholar]
- Long, J.A.; Stricker, S.A. Brachiopoda. In Reproduction of Marine Invertebrates, Volume VI, Echinoderms and Lophophorates; Giese, A.C., Pearse, J.S., Pearse, V., Eds.; The Boxwood Press: Pacific Grove, CA, USA, 1991; pp. 47–84. [Google Scholar]
- Lüter, C. Brachiopod larval setae—A key to the phylum’s ancestral life cycle? In Brachiopods; CRC Press: Boca Raton, FL, USA, 2003; pp. 60–69. [Google Scholar]
- Strathmann, R.R. Progressive vacating of adaptive types during the Phanerozoic. Evolution 1978, 32, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Valentine, J.W.; Jablonski, D. Larval adaptations and patterns of brachiopod diversity in space and time. Evolution 1983, 37, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Freeman, G.; Lundelius, J.W. Changes in the timing of mantle formation and larval life history traits in linguliform and craniiform brachiopods. Lethaia 1999, 32, 197–216. [Google Scholar] [CrossRef]
- Paine, R.T. Ecology of the brachiopod Glottidia pyramidata. Ecol. Monogr. 1963, 33, 187–213. [Google Scholar] [CrossRef]
- Ashworth, J.H. On the larvae of Lingula and Pelagodiscus (Discinisca). Trans. R. Soc. Edinb. 1915, 51, 45–69. [Google Scholar] [CrossRef]
- Chuang, S.H. The larvae of a discinid (Inarticulata, Brachiopoda). Biol. Bull. 1968, 135, 263–272. [Google Scholar] [CrossRef]
- Chuang, S.H. Larval development in Discinisca (inarticulate brachiopod). Am. Zool. 1977, 17, 39–53. [Google Scholar] [CrossRef]
- Emig, C.C. Brachiopods. In Marine Biodiversity of Costa Rica, Central America; Whertmann, I.S., Cortés, J., Eds.; Springer: Berlin, Germany, 2008; pp. 417–420. ISBN 978-1-4020-8278-8. [Google Scholar]
- Cooper, G.A. Brachiopods from the Caribbean Sea and adjacent waters. Stud. Trop. Oceanogr. Miami 1977, 14, 1–211. [Google Scholar]
- Emig, C.C. Taxonomie du genre Glottidia (Brachiopodes Inarticulés). Bull. Mus. Natl. Hist. Nat. Paris 1983, 4, 469–489. [Google Scholar]
- Labarbera, M. Mechanisms of spatial competition of Discinisca strigata (Inarticulata: Brachiopoda) in the intertidal of Panama. Biol. Bull. 1985, 168, 91–105. [Google Scholar] [CrossRef]
- Emig, C.C.; Bitner, M.A.; Alvarez, F. Brachiopoda Database. Available online: http://paleopolis.rediris.es/brachiopoda_database/index.html (accessed on 30 December 2018).
- Santagata, S.; Tunnell, J.W., Jr. Brachiopoda of the Gulf of Mexico. In Gulf of Mexico Origin, Waters, and Biota: Biodiversity, 1st ed.; Texas A&M University Press: College Station, TX, USA, 2009; Volume 1, pp. 1137–1141. ISBN 1603440941. [Google Scholar]
- Collin, R.; Venera-Pontón, D.E.; Driskell, A.C.; Chan, K.-Y.K.; MacDonald, K.S.; Boyle, M.J. Understanding Neotropical diversity of phoronids with DNA barcoding of planktonic larvae. Invertebr. Biol. 2018. accepted. [Google Scholar]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Meyer, C.P. Molecular systematics of cowries (Gastropoda: Cypraeidae) and diversification patterns in the tropics. Biol. J. Linn. Soc. Lond. 2003, 79, 401–459. [Google Scholar] [CrossRef]
- Palumbi, S.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR, Version 2; Department of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Gascuel, O. BIONJ: An improved version of the NJ algorithm based on a simple model of sequence data. Mol. Biol. Evol. 1997, 14, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The barcode of life data system (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Lassmann, T.; Sonnhammer, E.L.L. Kalign—An accurate and fast multiple sequence alignment algorithm. BMC Bioinform. 2005, 6, 298. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Saito, M.; Kojima, S.; Endo, K. Mitochondrial COI sequences of brachiopods: Genetic code shared with protostomes and limits of utility for phylogenetic reconstruction. Mol. Phylogenet. Evol. 2000, 15, 331–344. [Google Scholar] [CrossRef]
- Wu, L.J.; Sheng, G.L.; Lai, X.L.; Hou, X.; Yuan, J. Geographical pattern analysis of the brachiopod Lingula anatina based on mitochondrial COI gene sequences. Geol. Sci. Technol. Inform. 2010, 29, 17–22. [Google Scholar]
- Endo, K.; Ozawa, T.; Kojima, S. Nuclear and mitochondrial gene sequences reveal unexpected genetic heterogeneity among northern Pacific populations of the brachiopod Lingula anatina. Mar. Biol. 2001, 139, 105–112. [Google Scholar] [CrossRef]
- Endo, K.; Noguchi, Y.; Ueshima, R.; Jacobs, H.T. Novel repetitive structures, deviant protein-encoding sequences and unidentified ORFs in the mitochondrial genome of the brachiopod Lingula anatina. J. Mol. Evol. 2005, 6, 36–53. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.J.; Satoh, N.; Endo, K. Mitochondrial gene order variation in the brachiopod Lingula anatina and its implications for mitochondrial evolution in lophotrochozoans. Mar. Genomics 2015, 24, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Mackey, L.Y.; Winnepenninckx, B.; De Watcher, R.; Backeljau, T.; Emschermann, P.; Garey, J.R. 18S rRNA suggests that entoprocta are protostomes, unrelated to ectoprocta. J. Mol. Evol. 1996, 42, 552–559. [Google Scholar] [CrossRef]
- Cohen, B.L.; Gawthrop, A.; Cavalier–Smith, T. Molecular phylogeny of brachiopods and phoronids based on nuclear–encoded small subunit ribosomal RNA gene sequences. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 2039–2061. [Google Scholar] [CrossRef]
- Field, K.G.; Olsen, G.J.; Lane, D.J.; Giovannoni, S.J.; Ghiselin, M.T.; Raff, E.C.; Pace, N.R.; Raff, R.A. Molecular phylogeny of the animal kingdom. Science 1988, 239, 748–753. [Google Scholar] [CrossRef]
- Sato, S.; Endo, K.; Yamashita, H. Morphological and genetic comparisons between Lingula adamsi Dall, 1873 from South Korea and Japan. Jpn. J. Benthol. 2004, 59, 13–18. [Google Scholar] [CrossRef]
- Cohen, B.L. Monophyly of brachiopods and phoronids: Reconciliation of molecular evidence with Linnaean classification (the subphylum Phoroniformea nov.). Proc. R. Soc. Lond. B 2000, 267, 225–231. [Google Scholar] [CrossRef]
- Halanych, K.M.; Bacheller, J.D.; Aguinaldo, A.M.; Liva, S.M.; Hillis, D.M.; Lake, J.A. Evidence from 18S ribosomal DNA that the lophophorates are protostome animals. Science 1995, 267, 1641–1643. [Google Scholar] [CrossRef]
- Cohen, B.L.; Weydmann, A. Molecular evidence that phoronids are a subtaxon of brachiopods (Brachiopoda: Phoronata) and that genetic divergence of metazoan phyla began long before the early Cambrian. Org. Divers. Evol. 2005, 5, 253–273. [Google Scholar] [CrossRef]
- Cohen, B.L. Rerooting the rRNA tree reveals phoronids to be ’brachiopods without shells’; dangers of wide taxon samples in metazoan phylogenetics (Phoronida; Brachiopoda). Zool. J. Linn. Soc. 2013, 167, 82–92. [Google Scholar] [CrossRef]
- Hammond, L.S. Breeding season, larval development and dispersal of Lingula anatina (Brachiopoda, Inarticulata) from Townsville, Australia. J. Zool. 1982, 198, 183–196. [Google Scholar] [CrossRef]
- Hammond, L.S. The larvae of a discinid (Brachipoda: Inarticulata) from inshore waters near Townsville, Australia, with revised identifications of previous records. J. Natl. Hist. 1980, 14, 647–661. [Google Scholar] [CrossRef]
- Chuang, S.H. The inarticulate brachiopod larvae of the International Indian Ocean Expedition. J. Mar. Biol. Ass. India 1973, 15, 538–544. [Google Scholar]
- Fagetti, E.G. Nota sobre larvas de Brachiopoda Discinidae de la costa Chilena. Montemar 1964, 4, 195–200. [Google Scholar]
- Williams, A.; Cohen, B.L.; Cusack, M.; Long, S.L. Provenance of Atlantic lingulid brachiopods. Palaeontology 2000, 43, 999–1018. [Google Scholar] [CrossRef]
- Barber, P.; Boyce, S.L. Estimating diversity of Indo-Pacific coral reef stomatopods through DNA barcoding of stomatopod larvae. Proc. R. Soc. Lond. B 2006, 273, 2053–2061. [Google Scholar] [CrossRef]
- Mahon, A.R.; Thornhill, D.J.; Norenburg, J.L.; Halanych, K.M. DNA uncovers Antarctic nemertean biodiversity and exposes a decades-old cold case of asymmetric inventory. Polar Biol. 2010, 33, 193–202. [Google Scholar] [CrossRef]
- Maslakova, S.A.; Ellison, C.; Moss, N.D.; Dillenburg, B.; Howland, C.; Robbins, K.; Schwartz, M.L.; Partridge, M.; Zattara, E.; Collin, R.; et al. DNA-barcoding of benthic and planktonic life history stages reveals a large amount of undescribed, cryptic, and unsampled diversity of nemerteans in the Caribbean Sea. In preparation.
- Lee, T.; Ó Foighil, D. Placing the Floridian marine genetic disjunction into a regional evolutionary context using the scorched mussel, Brachidontes exustus, species complex. Evolution 2007, 59, 2139–2158. [Google Scholar] [CrossRef]
- Cowen, R.K.; Paris, C.B.; Srinivasan, A. Scaling of connectivity in marine populations. Science 2006, 311, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Ferguson, E.; Haney, R.; Wares, J.; Silliman, B. Population genetics of a trochid gastropod broadens picture of Caribbean Sea connectivity. PLoS ONE 2010, 5, e12675. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Fragment Length (bp) | References |
|---|---|---|---|
| COI Barcode fragment | jgLCO1490: TITCIACIAAYCAYAARGAYATTGG jgHCO2198: TAIACYTCIGGRTGICCRAARAAYCA or dgLCO1490: GGTCAACAAATCATAAAGAYATYGG dgHCO2198: TAAACTTCAGGGTGACCAAARAAYCA | 654 | [19,20] |
| 16S | 16Sar CGCCTGTTTATCAAAAACAT 16Sbr CCGGTCTGAACTCAGATCACGT | ~525 | [21] |
| 18S | EukF (modified Primer A): AACCTGGTTGATCCTGCCAGT SR7: GTTCAACTACGAGCTTTTTAA | ~600 | [22] |
| OTU/Species 1 | Genbank # COI | Genbank # 16S | Genbank # 18S | Location | Collection Dates |
|---|---|---|---|---|---|
| New Data | |||||
| Larval OTU1—Glottidia | MK092035, 36, 38–39, 41, 47,49–51, 55–58, 61 | MK073388, 89, 91–93, 95, MK073401, 03–04, 06, 10–13, 16 | MK073422, 26,32–33, 38, 49, 53, 55, 58, 68, 69, 70, 73 | Bahía Almirante | Jul 2013, August & November 2015, February-March & June 2016 |
| Larval OTU2—Glottidia | MK092033, 34, 37,40 42–46, 48, 52–54, 59–60, 62 | MK073386, 87, 90, 94, 96–99, MK073400, 02, 05, 07–09, 14–15, 17 | MK073419, 20, 28, 34, 39, 40, 42–43, 47–48, 52, 62–63, 65, 74–75, 79 | Bay of Panama | April-June & November 2014 |
| Larval OTU3—Discinid | ------- | --------- | MK073423, 24–25, 27, 29,30–31, 35–37, 41, 44–46, 50–51, 54, 56–57, 59, 61, 64, 66, 72, 76–78, 80 | Bahía Almirante and Bay of Panama | July-August 2013, August 2015, November 2014 & 2015, March & June 2016 |
| Larval OTU4—Discinid | -------- | ---------- | MK073421, 60, 67, 71 | Bay of Panama | August 2013, March 2014 |
| Published Data | Reference | ||||
| Lingulids | |||||
| Lingula anatina | AB026520 | Japan | [27] | ||
| Lingula anatina | GU056040-41 | China | [28] | ||
| Lingula anatina | AB056460 | Japan | [29] | ||
| Lingula anatina | AB056461, AB056462 | Hong Kong | [29] | ||
| Lingula anatina | AB178773 | AB178733 | Japan | [30] | |
| Lingula anatina | KX774482, NC_036679 | KX774482, NC_036679 | South Korea | Karagozlu & Kim, Unpublished 2 | |
| Lingula anatina | KP881498 | KP881498 | KP780396 | Japan | [31] |
| Lingula anatina | X81631 | Hong Kong | [32] | ||
| Lingula anatina | AB747095 | Japan | [31] | ||
| Lingula anatina | U08331 | New Caledonia | [33] | ||
| Lingula rostrum | AB855774 | Japan | [31] | ||
| Lingula reevii | AB747096 | Japan | [31] | ||
| Lingula reevii | AH001678 | unknown | [34] | ||
| Lingula reevii | LC334155 | Japan | Kurita et al. Unpublished | ||
| Lingula shantungensis | AB056459 | Japan | [29] | ||
| Lingula adamsi | AB128054-63 | South Korea | [35] | ||
| Lingula adamsi | U08329 | New Caledonia | [33] | ||
| Glottidia palmeri | AF201744 | Baja California | [36] | ||
| Glottidia pyramidata | U12647 | Florida, Gulf Coast (K Halanych, pers.com) | [37] | ||
| Glottidia pyramidata | MK015669 | Florida, Gulf Coast | K. Kocot, unpublished | ||
| Discinids | |||||
| Discinisca cf. tenuis | AF202444 | Namibia | [36] | ||
| Discinisca cf. tenuis | U08327 | Namibia | [33] | ||
| Discinisca cf. tenuis | AY842020 | Panama Bay, Panama | [38] | ||
| Discina striata | U08333 | Gambia | [33] | ||
| Pelagodiscus atlanticus | JQ414032 | Antarctica | [39] | ||
| Pelagodiscus atlanticus | JQ414033 | Uncertain 3 | [39] | ||
| Brachiopod sp. | AF025935 | Guam, Pacific | [33] | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collin, R.; Venera-Pontón, D.E.; Driskell, A.C.; Macdonald, K.S., III; Boyle, M.J. Planktotrophic Brachiopod Larvae from the Pacific and Caribbean of Panama. Diversity 2019, 11, 2. https://doi.org/10.3390/d11010002
Collin R, Venera-Pontón DE, Driskell AC, Macdonald KS III, Boyle MJ. Planktotrophic Brachiopod Larvae from the Pacific and Caribbean of Panama. Diversity. 2019; 11(1):2. https://doi.org/10.3390/d11010002
Chicago/Turabian StyleCollin, Rachel, Dagoberto E. Venera-Pontón, Amy C. Driskell, Kenneth S. Macdonald, III, and Michael J. Boyle. 2019. "Planktotrophic Brachiopod Larvae from the Pacific and Caribbean of Panama" Diversity 11, no. 1: 2. https://doi.org/10.3390/d11010002
APA StyleCollin, R., Venera-Pontón, D. E., Driskell, A. C., Macdonald, K. S., III, & Boyle, M. J. (2019). Planktotrophic Brachiopod Larvae from the Pacific and Caribbean of Panama. Diversity, 11(1), 2. https://doi.org/10.3390/d11010002