Relationship between Genetic Variability and Land Use and Land Cover in Populations of Campomanesia adamantium (Myrtaceae)

 , , ,
, , ,
Abstract
1. Introduction
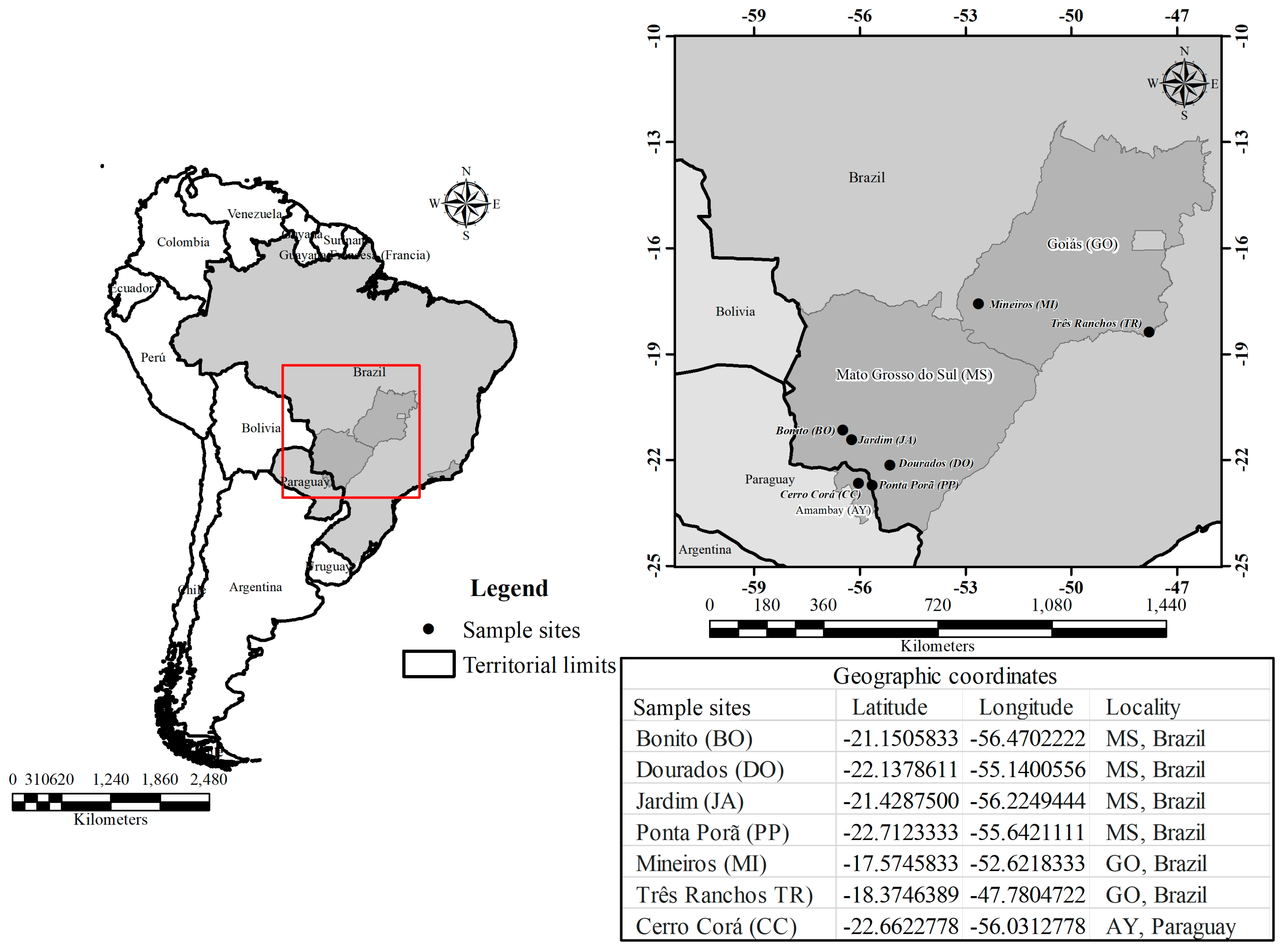
2. Materials and Methods
2.1. Plant Material
2.2. Microsatellite Markers
2.3. Population Genetics Analysis
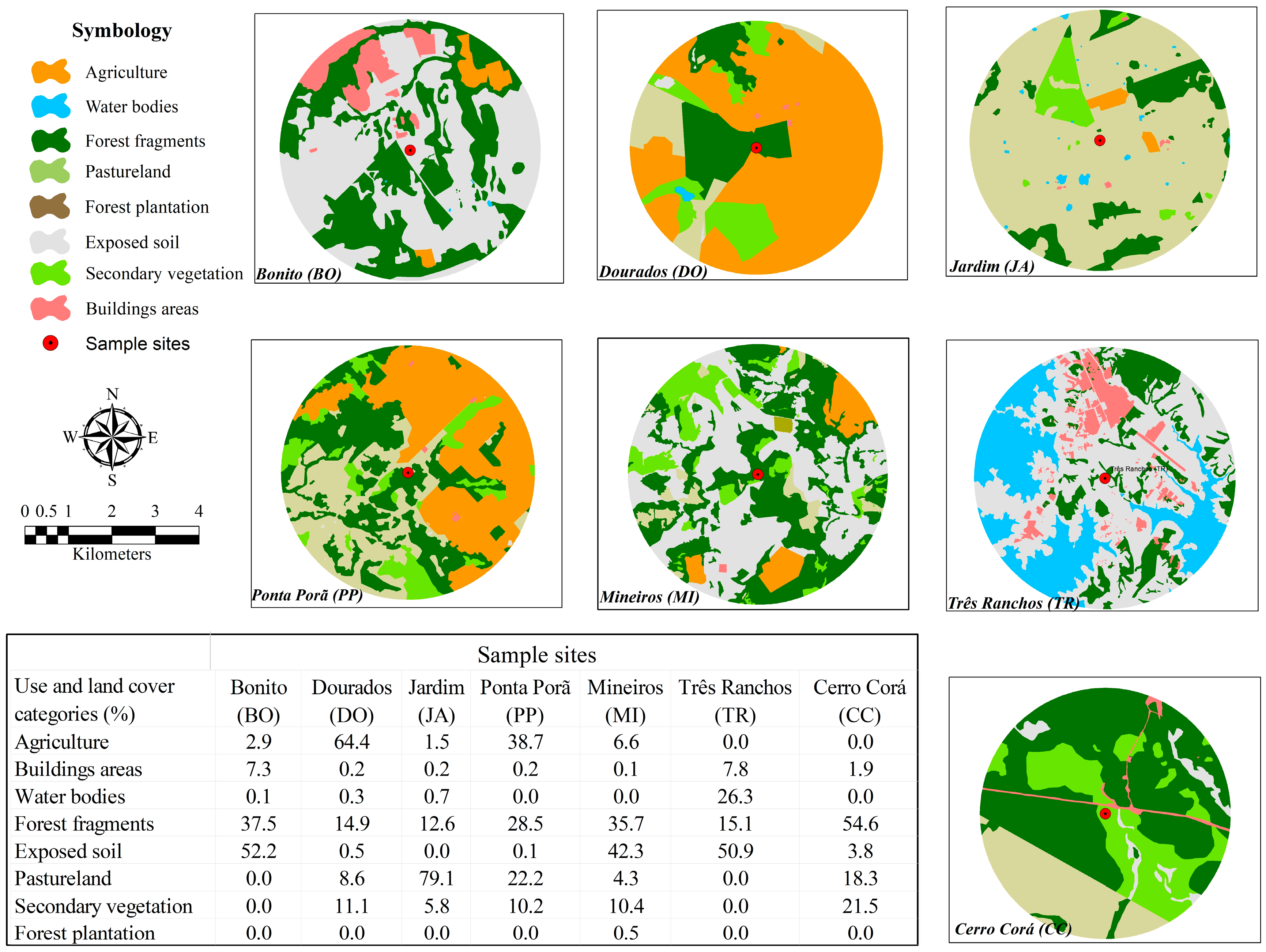
2.4. Use and Land Cover Analysis
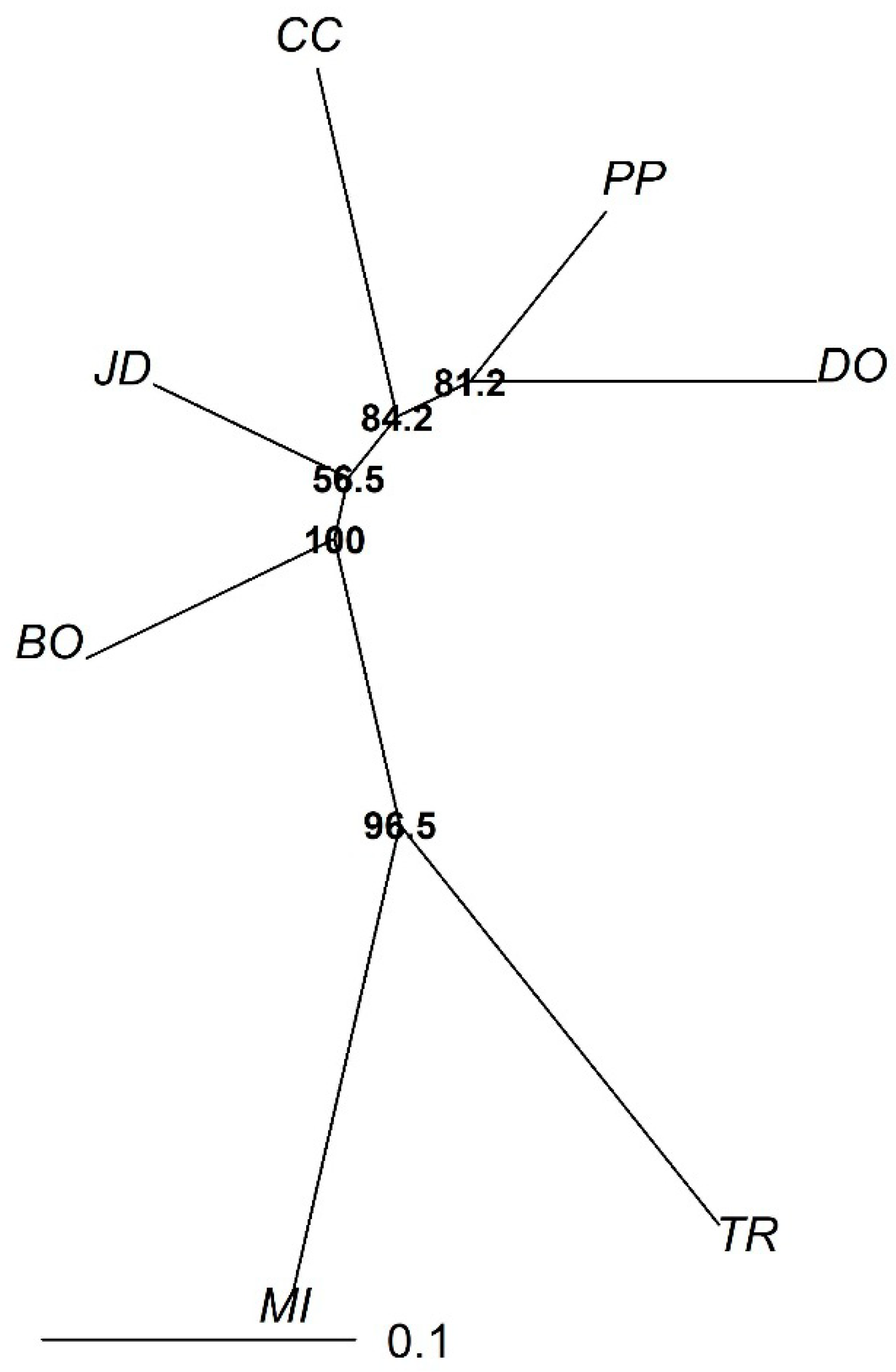
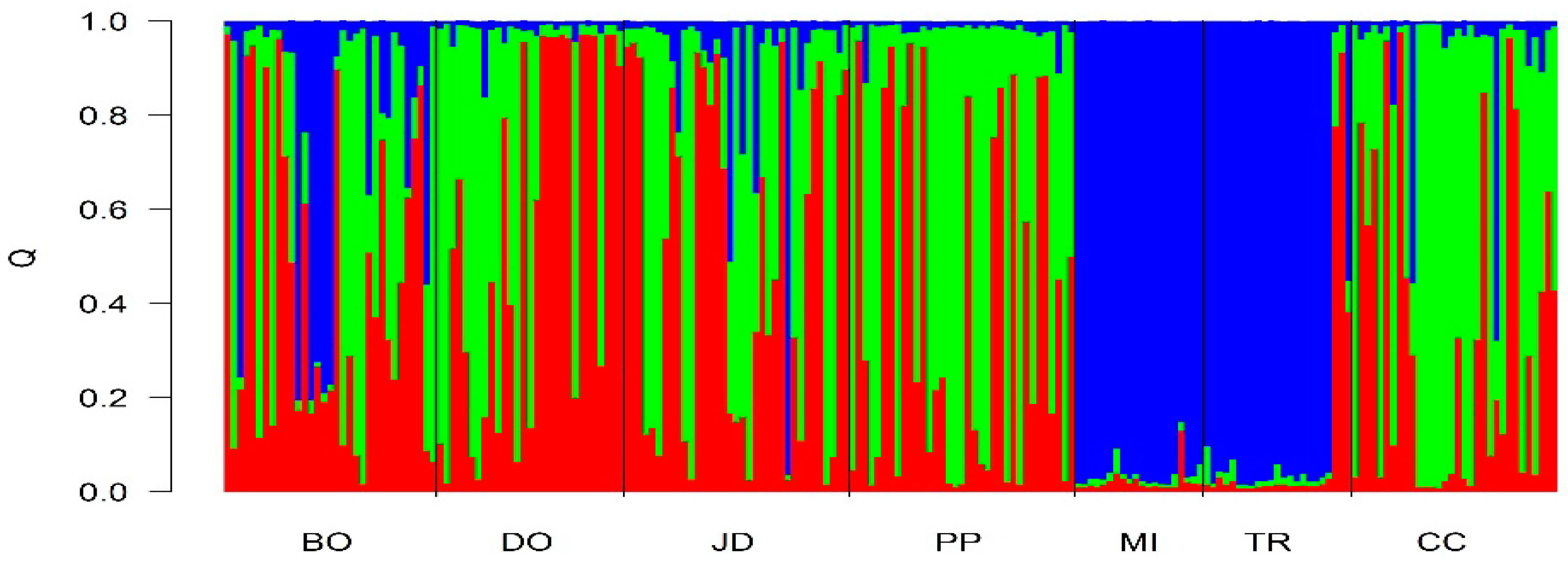
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, D.B.; Silva, J.A.; Junqueira, N.T.V.; Andrade, L.R.M. Frutas do Cerrado; Embrapa Informação Tecnológica: Brasília, Brazil, 2001; p. 179. [Google Scholar]
- Porto, A.C.; Gulias, A.P.M. Gabiroba. In Frutas Nativas da Região Centro-Oeste do Brasil, 1st ed.; Vieira, R.F., Agostini-Costa, T.S., Silva, D.D., Ferreira, F.R., Sano, S.M., Eds.; Embrapa Recursos Genéticos e Biotecnologia: Brasília, Brazil, 2007; pp. 164–172. [Google Scholar]
- Cardoso, C.A.; Salmazzo, G.R.; Honda, N.K.; Prates, C.B.; Vieira, M.C.; Coelho, R.G. Antimicrobial activity of the extracts and fractions of hexanic fruits of Campomanesia species (Myrtaceae). J. Med. Food 2006, 13, 1273–1276. [Google Scholar] [CrossRef] [PubMed]
- Agostini-Costa, T.S.; Silva, D.B.; Vieira, R.F.; Sano, S.M.; Ferreira, F.R. Espécies de maior relevância para a região Centro-Oeste. In Frutas Nativas da Região Centro-Oeste do Brasil, 1st ed.; Vieira, R.F., Agostini-Costa, T.S., Silva, D.D., Ferreira, F.R., Sano, S.M., Eds.; Embrapa Recursos Genéticos e Biotecnologia: Brasília, Brazil, 2007; pp. 12–24. [Google Scholar]
- Melchior, S.J.; Custódio, C.C.; Marques, T.A.; Machado, N.B. Colheita e armazenamento de sementes de gabiroba (Campomanesia adamantium Camb.-Myrtaceae) e implicações na germinação. Rev. Bras. Sementes 2006, 28, 141–150. [Google Scholar] [CrossRef]
- Resende, H.C.; Teixeira, T.A. Diversidade genética em Campomanesia (MYRTACEA) estimada por análise multivariada de características fenotípicas. Ceres 2009, 56, 85–92. [Google Scholar]
- Oliveira, M.C.; Santana, D.G.; Santos, C.M. Biometrics of fruits and seeds and seedling emergence of two species fruit of the Campomanesia genus. Rev. Bras. Frutic. 2011, 33, 446–455. [Google Scholar] [CrossRef]
- Dresch, D.M.; Scalon, S.P.Q.; Masetto, T.E.; Vieira, M.C. Germination and vigor of Campomanesia adamantium seeds according to fruit and seed size. Pesqui. Agropecu. Trop. 2013, 43, 262–271. [Google Scholar] [CrossRef]
- Wesp, C.L.; Santana, M.A.; Gasparetto, B.F.; Barros, I.B.I. Caracterização física de frutos de guabirobeiras (Campomanesia spp.) coletados no estado do Rio Grande do Sul. Cad. Agroecol. 2013, 8, 1–4. [Google Scholar]
- Sehgal, D.; Raina, S.N. DNA markers and germplasm resource diagnostics: New perspectives in crop improvement and conservation strategies. In Utilization of Biotechnology in Plant Sciences; Arya, I.D., Arya, S., Eds.; Forest Research Institute: Dehradun, India, 2008; pp. 39–54. [Google Scholar]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Brondani, R.P.V.; Brondani, C.; Tarchini, R.; Grattapaglia, D. Development, characterization and mapping of microsatellite markers in Eucalyptus grandis and E. urophylla. Theor. Appl. Genet. 1998, 97, 816–827. [Google Scholar] [CrossRef]
- Parida, S.K.; Kalia, S.K.; Kaul, S.; Dalal, V.; Hemaprabha, G.; Selvi, A.; Pandit, A.; Singh, A.; Gaikwad, K.; Sharma, T.R.; et al. Informative genomic microsatellite markers for efficient genotyping applications in sugarcane. Theor. Appl. Genet. 2009, 118, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.C.; Shih, H.C.; Chang, L.W.; Li, W.R.; Lin, H.Y.; Ju, L.P. Isolation of 16 polymorphic microsatellite markers from an endangered and endemic species. Podocarpus nakaii (Podocarpaceae). Am. J. Bot. 2011, 98, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.J.; Pádua, J.G.; Zucchi, M.I.; Vencovsky, R.; Vieira, M.L.C. Origin, evolution and genome distribution of microsatellites. Genet. Mol. Biol. 2006, 29, 294–307. [Google Scholar] [CrossRef]
- Fagundes, B.S.; Silva, L.F.; Giacomin, R.M.; Secco, D.; Díaz-Cruz, J.A.; Da-Silva, P.R. Transferability of microsatellite markers among Myrtaceae species and their use to obtain population genetics data to help the conservation of the Brazilian Atlantic Forest. Trop. Conserv. Sci. 2016, 9, 408–422. [Google Scholar] [CrossRef]
- Assis, E.S.; Reis, E.F.; Pinto, J.F.; Contim, L.A.; Dias, L.A. Genetic diversity of gabiroba based on random amplified polymorphic DNA markers and morphological characteristics. Genet. Mol. Res. 2013, 12, 3500–3509. [Google Scholar] [CrossRef] [PubMed]
- Miranda, E.A.; Boaventura-Novaes, C.R.; Braga, R.S.; Reis, E.F.; Pinto, J.F.; Telles, M. P Validation of EST-derived microsatellite markers for two Cerrado-endemic Campomanesia (Myrtaceae) species. Genet. Mol. Res. 2016, 15, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vellend, M. Parallel effects of land-use history on species diversity and genetic diversity of forest herbs. Ecology 2004, 85, 3043–3055. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Grattapaglia, D.; Mamani, E.M.; Silva-Junior, O.B.; Faria, D.A. A novel genome-wide microsatellite resource for species of Eucalyptus with linkage-to-physical correspondence on the reference genome sequence. Mol. Ecol. Resour. 2015, 15, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J. FSTAT: A Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3.2); University of Lausanne, Department of Ecology and Evolution: Lausanne, Switzerland, 2002; Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 25 May 2018).
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Roessli, D.; Excoffier, L. Arlequin: A Software for Population Genetics Data Analysis. User Manual ver 2.000; Genetics and Biometry Laboratory Department of Anthropology, University of Geneva: Geneva, Switzerland, 2000; Available online: http://www.cecalc.ula.ve/BIOINFO/servicios/herr1/ARLEQUIN/Arlequin.pdf (accessed on 15 June 2018).
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Chessel, D.; Duffour, A.; Dray, S. Ade4: Analysis of Ecological Data: Exploratory and Euclidean Methods in Environmental Sciences. 2011. Available online: http://cran.r-project.org/web/packages/ade4/ (accessed on 22 January 2018).
- Raymond, M.; Rousset, F. GENEPOP (version-1.2)—Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 59–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Piry, S.; Luikart, G.; Cornuet, J.M. Bottleneck: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 4, 502–503. [Google Scholar] [CrossRef]
- Luikart, G.; Cornuet, J.M. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv. Biol. 1998, 12, 228–237. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop Trial Version®: Release 10.4. Redlands; Environmental Systems Research Institute: Redlands, CA, USA, 2015. [Google Scholar]
- Phong, D.T.; Lieu, T.T.; Hien, V.T.T.; Hiep, N.T. Genetic diversity of the endemic flat needle pine Pinus krempfii (Pinaceae) from Vietnam revealed by SSR markers. Genet. Mol. Res. 2015, 14, 7727–7739. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, J. A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (Imidacloprid) on honey bees. Ecotoxicology 2011, 20, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.; Steffan-Dewenter, I.; et al. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.W.; Tautz, J.; Grünewald, B.; Fuchs, S. RFID tracking of sublethal effects of two neonicotinoid insecticides on the foraging behavior of Apis mellifera. PLoS ONE 2012, 7, e30023. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, A.; Peterson, A.C.; Garza, J.C.; Valdes, A.M.; Slatkin, M.; Freimer, N. Mutational processes of simple sequence repeat loci in human population. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.W. Genetics of Populations, 4th ed.; Jones and Bartlett: Boston, MA, USA, 2011; p. 675. ISBN 978-0763757373. [Google Scholar]
- Yeh, F.C. Population genetics. In Forest Conservation Genetics: Principles and Practice; Young, A., Boshier, D., Boyle, T., Eds.; CSIRO Publishing: Collingwood, Australia, 2000; pp. 21–37. [Google Scholar]
- Futuyma, D.J. Biologia Evolutiva, 2nd ed.; Sociedade Brasileira de Genética/CNPq: Riberão Preto, Brazil, 1992; p. 646. [Google Scholar]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Charles, H.; Godfray, J.; Garnett, T. Food security and sustainable intensification. Philos. Trans. R. Soc. B 2014, 369, 20120273. [Google Scholar] [CrossRef]
- Lanz, B.; Simon, D.; Tim, S. The expansion of modern agriculture and global biodiversity decline: An integrated assessment. Ecol. Econ. 2018, 144, 260–277. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | N | Ho | He | PIC |
|---|---|---|---|---|
| Embra 1076 | 3 | 0.99 | 0.51 | 0.39 |
| Embra 1335 | 4 | 0.42 | 0.36 | 0.32 |
| Embra 1363 | 16 | 0.80 | 0.89 | 0.88 |
| Embra 1364 | 21 | 0.78 | 0.93 | 0.92 |
| Embra 1374 | 12 | 0.58 | 0.80 | 0.77 |
| Embra 1470 | 6 | 0.53 | 0.52 | 0.47 |
| Embra 1811 | 9 | 0.19 | 0.36 | 0.34 |
| Mean (SD) | 10.14 ± 6.12 | 0.61 ± 0.24 | 0.62 ± 0.22 | 0.58 ± 0.24 |
| Populations | N | ANA | Ho | He | PA | FIS | AR | HWE |
|---|---|---|---|---|---|---|---|---|
| Bonito/MS | 33 | 6.14 ± 3.18 | 0.73 ± 0.03 | 0.63 ± 0.08 | 0.04 | −0.15 | 5.64 | 2 |
| Dourados/MS | 29 | 5.14 ± 2.73 | 0.61 ± 0.03 | 0.59 ± 0.08 | 0.03 | −0.03 | 4.77 | 4 |
| Jardim/MS | 35 | 6.00 ± 3.21 | 0.73 ± 0.03 | 0.64 ± 0.07 | 0.01 | −0.14 | 5.48 | 1 |
| Ponta Porã/MS | 35 | 5.86 ± 4.06 | 0.61 ± 0.03 | 0.61 ± 0.08 | 0.03 | 0.00 | 5.21 | 2 |
| Mineiros/GO | 20 | 4.86 ± 3.76 | 0.42 ± 0.04 | 0.46 ± 0.13 | 0.04 | 0.08 | 4.81 | 2 |
| Três Ranchos/GO | 20 | 4.57 ± 4.16 | 0.44 ± 0.04 | 0.44 ± 0.15 | NF | 0.00 | 4.50 | 2 |
| Cerro Corá/AY | 35 | 5.86 ± 4.38 | 0.62 ± 0.03 | 0.61 ± 0.08 | 0.03 | −0.01 | 5.47 | 4 |
| BO | DO | JD | PP | MI | TR | CC | |
|---|---|---|---|---|---|---|---|
| BO | 192.883 | 39.941 | 173.368 | 455.686 | 463.372 | 175.778 | |
| DO | 0.073 | 81.961 | 40.363 | 648.250 | 645.465 | 154.300 | |
| JA | 0.009 | 0.035 | 108.585 | 622.807 | 639.049 | 136.921 | |
| PP | 0.050 | 0.018 | 0.021 | 624.752 | 614.854 | 138.004 | |
| MI | 0.051 | 0.109 | 0.081 | 0.104 | 151.377 | 495.130 | |
| TR | 0.074 | 0.111 | 0.084 | 0.103 | 0.051 | 502.809 | |
| CC | 0.046 | 0.038 | 0.021 | 0.011 | 0.012 | 0.010 |
| Populations | Inferred Clusters | |||
|---|---|---|---|---|
| 1 | 2 | 3 | N | |
| Bonito/MS | 0.35 | 0.27 | 0.38 | 33 |
| Dourados/MS | 0.40 | 0.02 | 0.57 | 29 |
| Jardim/MS | 0.41 | 0.10 | 0.48 | 35 |
| Ponta Porã/MS | 0.57 | 0.02 | 0.40 | 35 |
| Mineiros/GO | 0.01 | 0.96 | 0.02 | 20 |
| Três Ranchos/GO | 0.02 | 0.97 | 0.01 | 20 |
| Cerro Corá/AY | 0.56 | 0.08 | 0.35 | 35 |
| Mutation Models | Populations | ||||||
|---|---|---|---|---|---|---|---|
| BO | DO | JD | PP | MI | TR | CC | |
| T.P.M. | 0.34375 | 0.1875 | 0.1875 | 0.01172 * | 0.8125 | 0.40625 | 0.05469 |
| S.M.M. | 0.28906 | 0.8125 | 0.53125 | 0.85156 | 0.96094 | 0.89063 | 0.53125 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crispim, B.D.A.; Bajay, M.M.; Vasconcelos, A.A.d.; Deo, T.G.; Braga, R.D.S.; Telles, M.P.d.C.; Vieira, M.D.C.; Carnevali, T.D.O.; Solórzano, J.C.J.; Grisolia, A.B. Relationship between Genetic Variability and Land Use and Land Cover in Populations of Campomanesia adamantium (Myrtaceae). Diversity 2018, 10, 106. https://doi.org/10.3390/d10040106
Crispim BDA, Bajay MM, Vasconcelos AAd, Deo TG, Braga RDS, Telles MPdC, Vieira MDC, Carnevali TDO, Solórzano JCJ, Grisolia AB. Relationship between Genetic Variability and Land Use and Land Cover in Populations of Campomanesia adamantium (Myrtaceae). Diversity. 2018; 10(4):106. https://doi.org/10.3390/d10040106
Chicago/Turabian StyleCrispim, Bruno Do Amaral, Miklos Maximiliano Bajay, Adrielle Ayumi de Vasconcelos, Thamiris Gatti Deo, Ramilla Dos Santos Braga, Mariana Pires de Campos Telles, Maria Do Carmo Vieira, Thiago De Oliveira Carnevali, Julio César Jut Solórzano, and Alexeia Barufatti Grisolia. 2018. "Relationship between Genetic Variability and Land Use and Land Cover in Populations of Campomanesia adamantium (Myrtaceae)" Diversity 10, no. 4: 106. https://doi.org/10.3390/d10040106
APA StyleCrispim, B. D. A., Bajay, M. M., Vasconcelos, A. A. d., Deo, T. G., Braga, R. D. S., Telles, M. P. d. C., Vieira, M. D. C., Carnevali, T. D. O., Solórzano, J. C. J., & Grisolia, A. B. (2018). Relationship between Genetic Variability and Land Use and Land Cover in Populations of Campomanesia adamantium (Myrtaceae). Diversity, 10(4), 106. https://doi.org/10.3390/d10040106