Effects of Habitat Structure, Plant Cover, and Successional Stage on the Bat Assemblage of a Tropical Dry Forest at Different Spatial Scales




Abstract
:1. Introduction
2. Materials and Methods
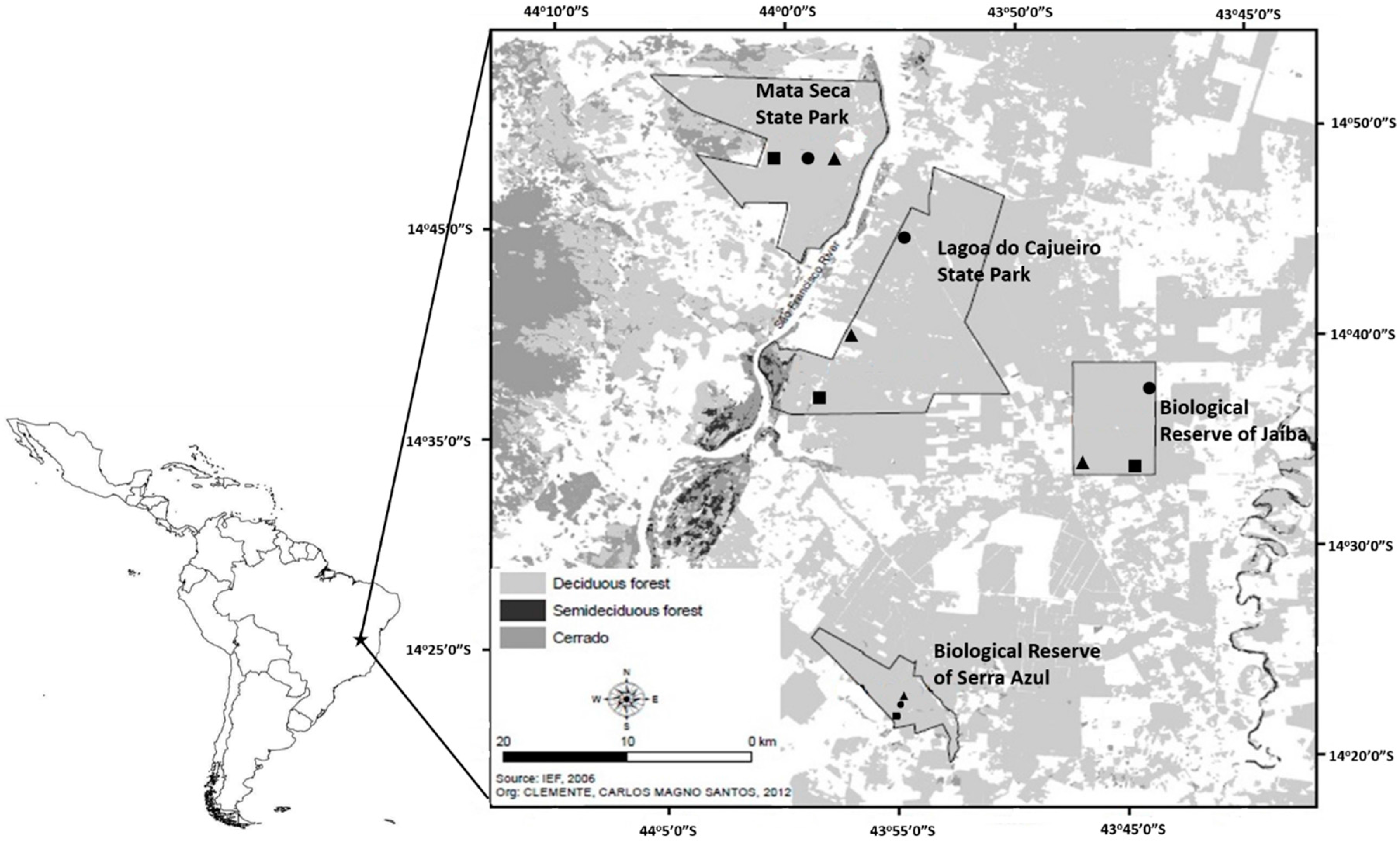
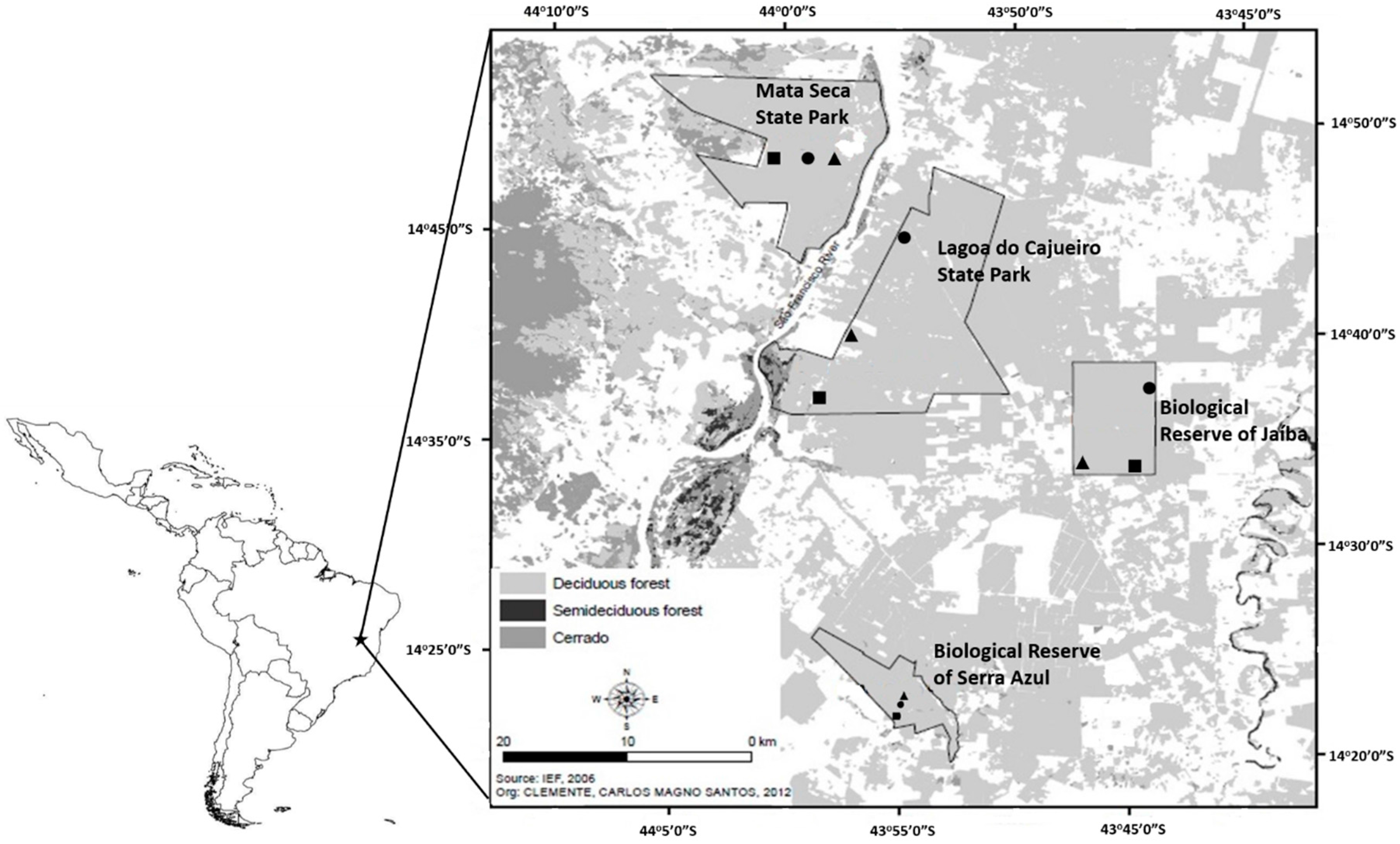
2.1. Sampling Sites
2.2. Bat Captures
2.3. Statistical Analysis
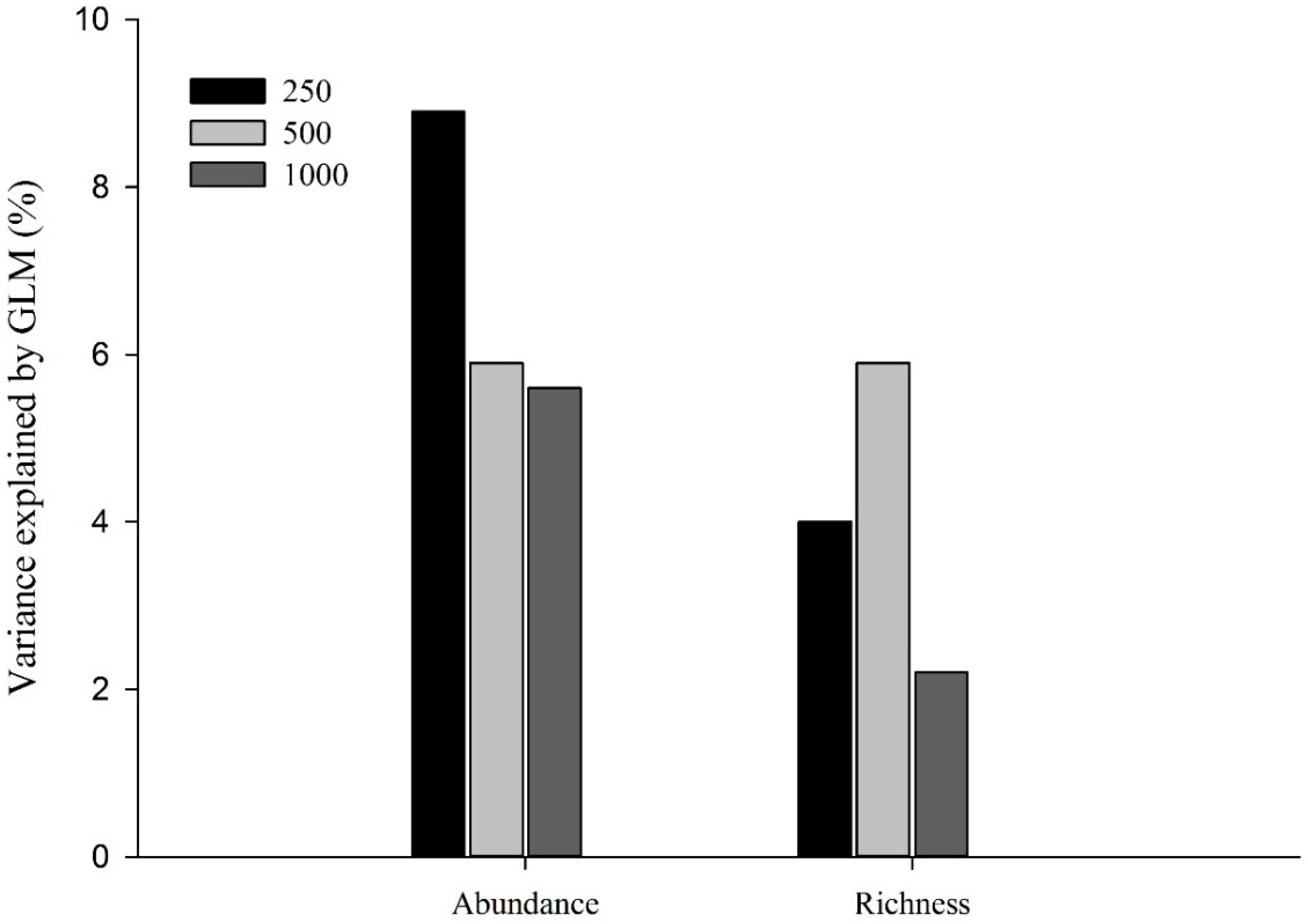
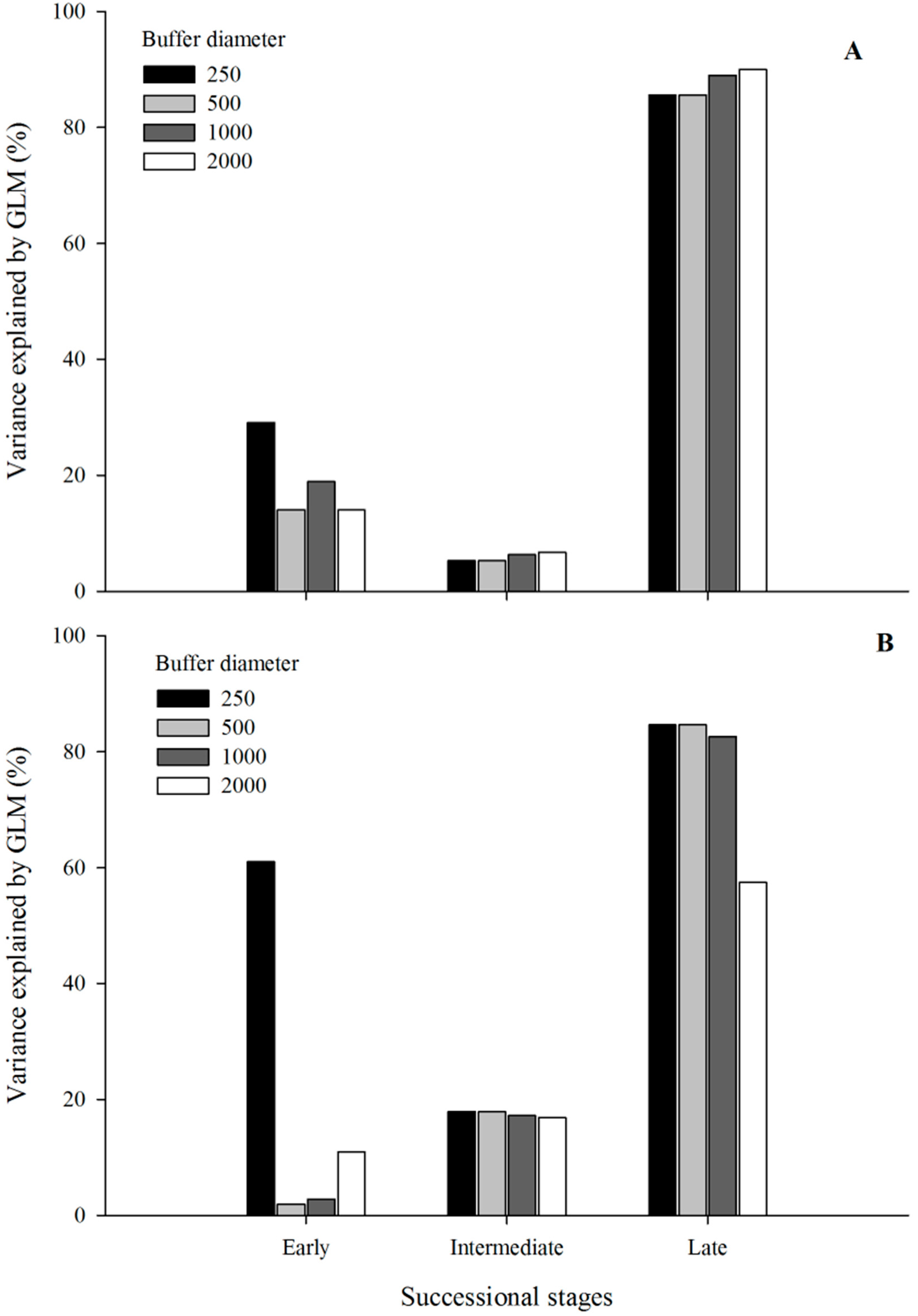
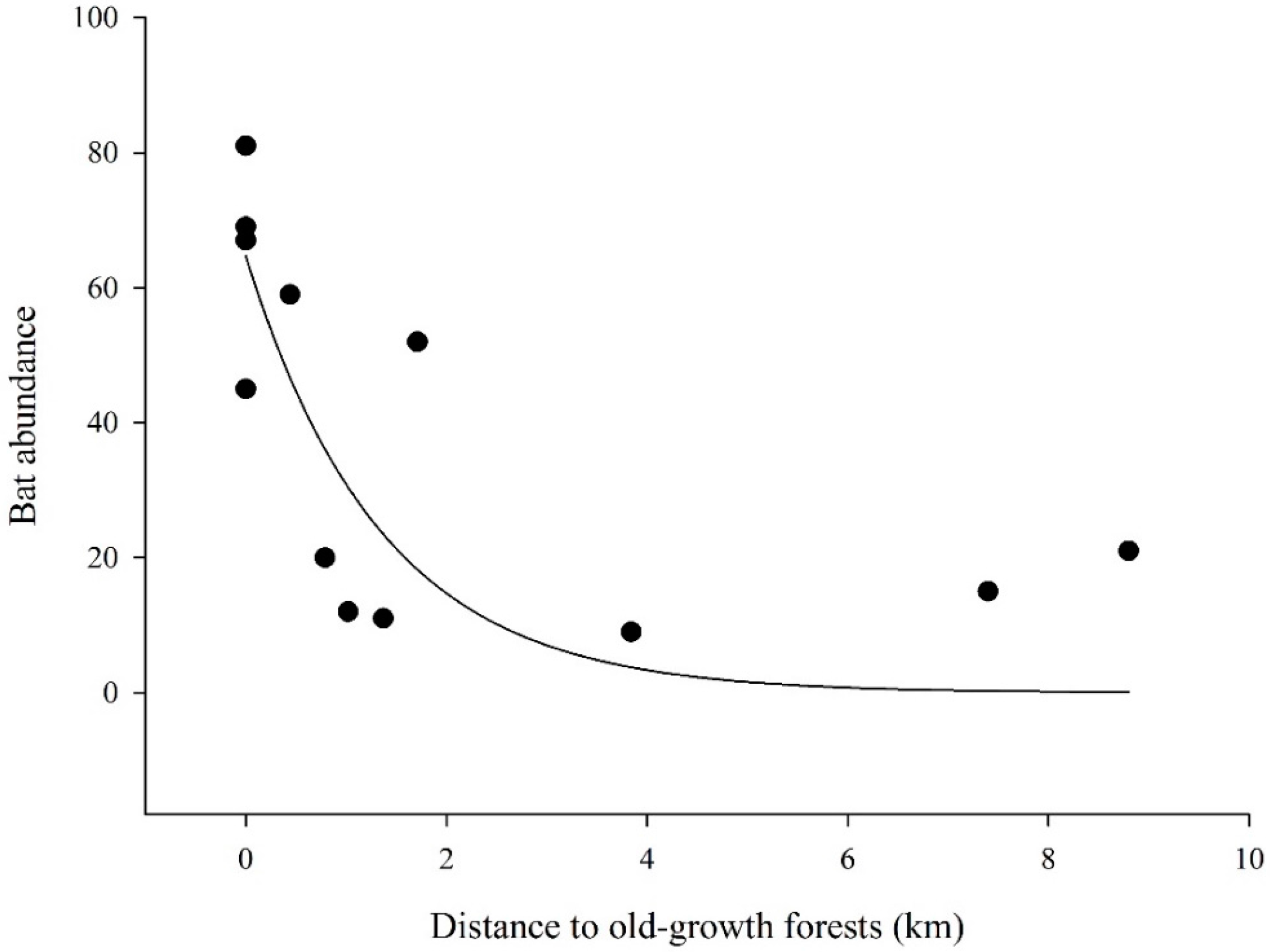
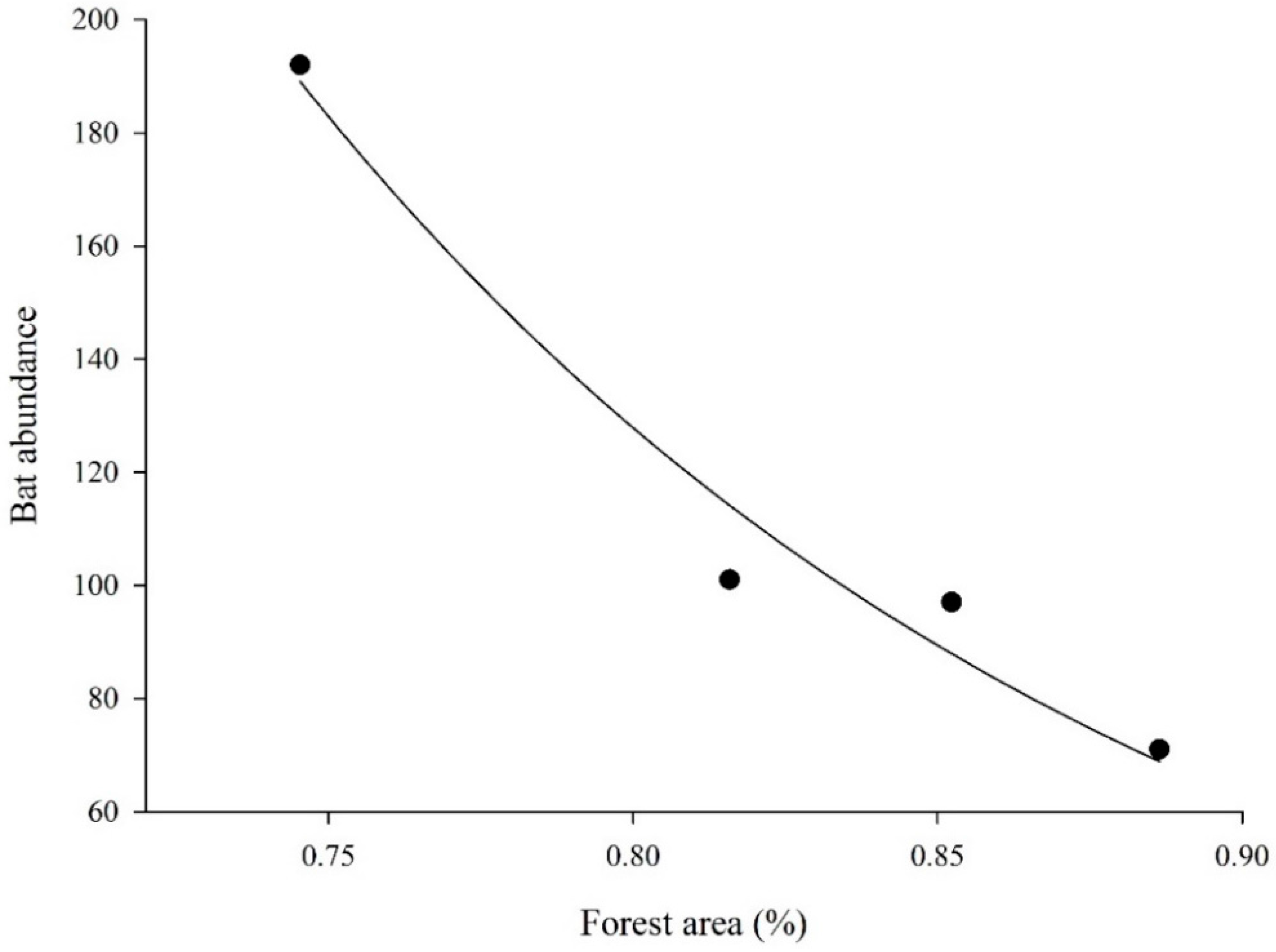
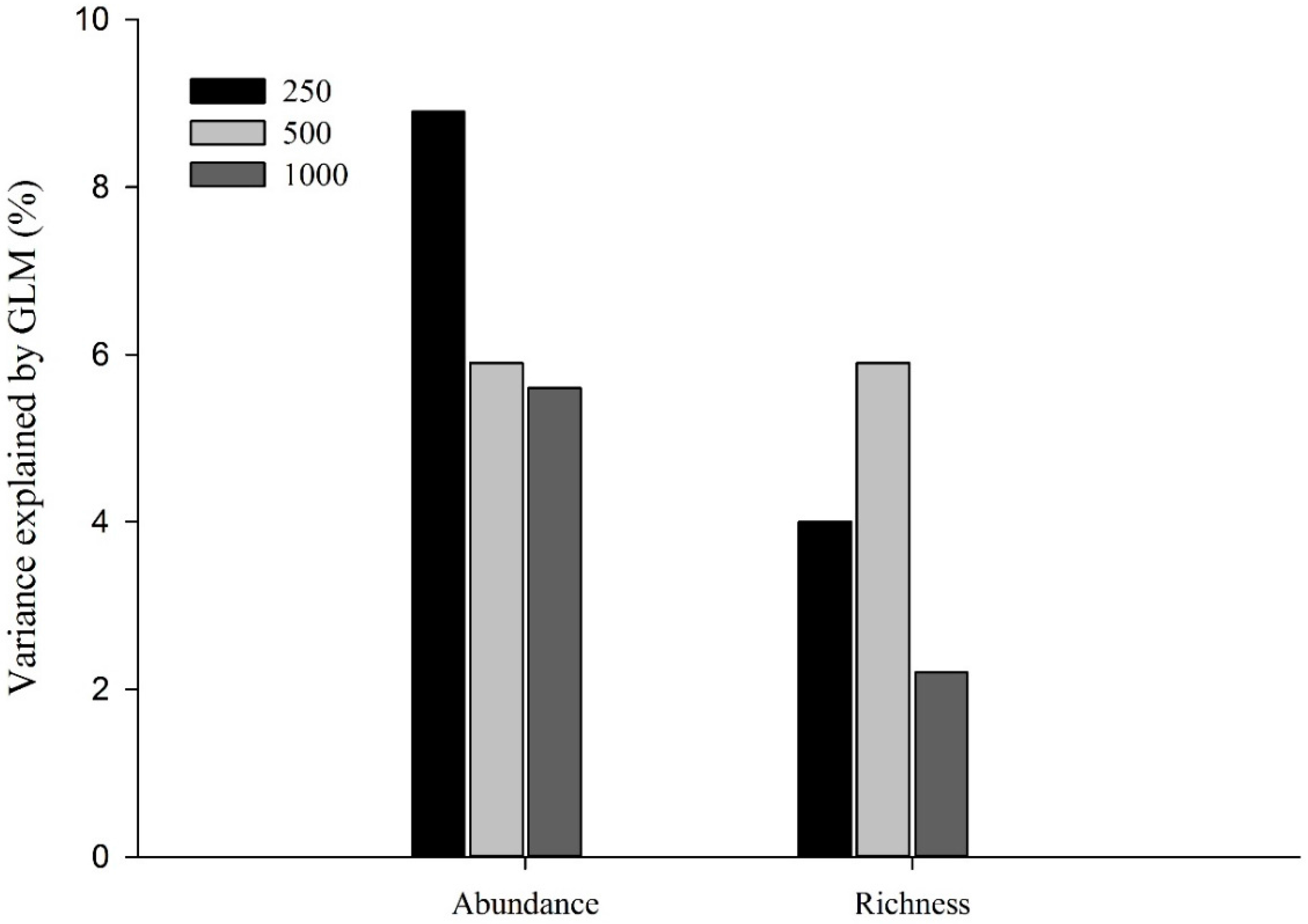
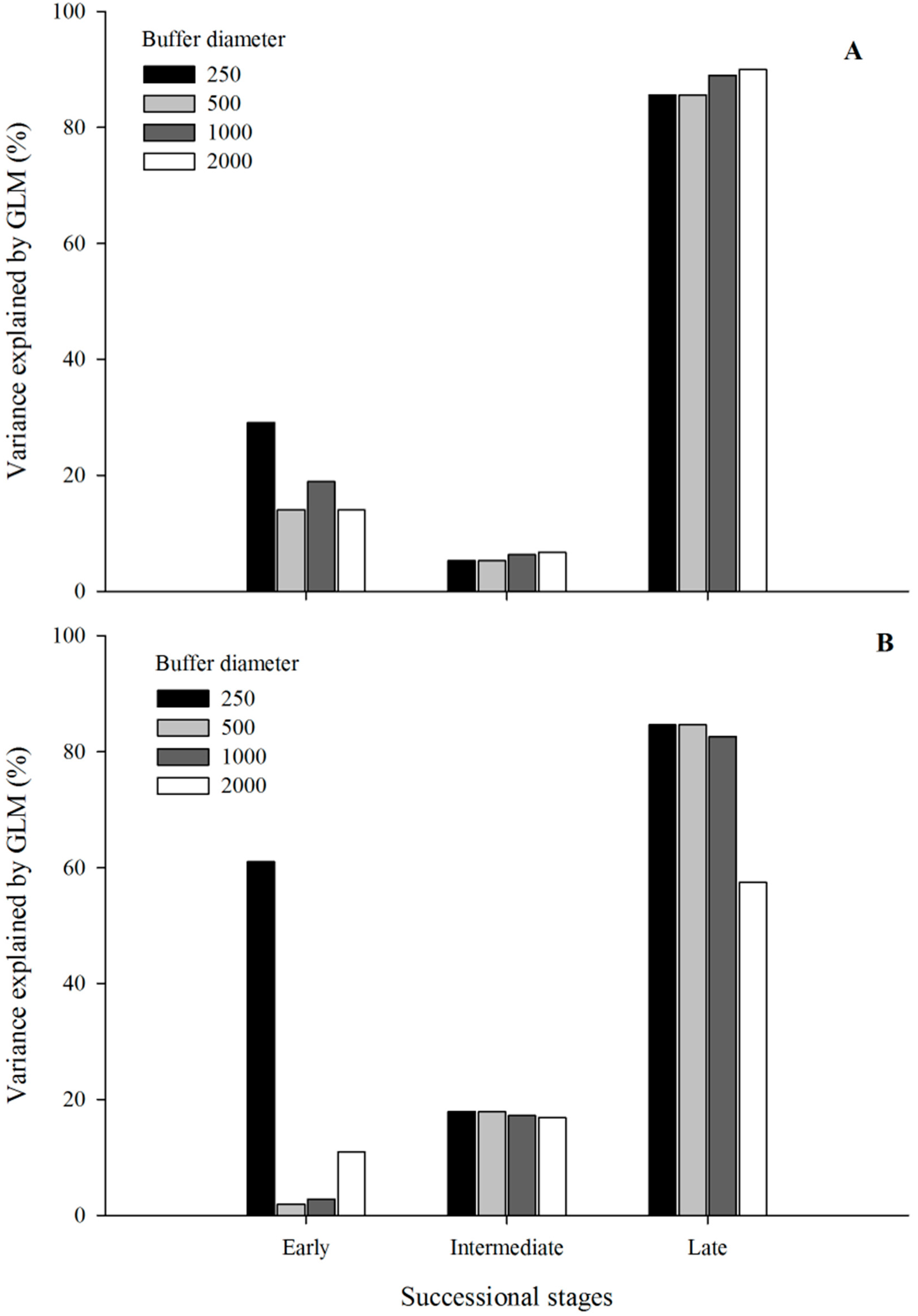
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Henry, M.; Jouard, S. Effect of bat exclusion on patterns of seed rain in tropical rain forest in French Guiana. Biotropica 2007, 39, 510–518. [Google Scholar] [CrossRef]
- Kalka, M.B.; Smith, A.R.; Kalko, E.K.V. Bats limit arthropods and herbivory in a tropical forest. Science 2008, 320, 71. [Google Scholar] [CrossRef] [PubMed]
- Quesada, M.; Stoner, K.E.; Rosas-Guerrero, V.; Palacios-Guevara, C.; Lobo, J.A. Effects of habitat disruption on the activity of nectarivorous bats (Chiroptera: Phyllostomidae) in a dry tropical forest: Implications for the reproductive success of the neotropical tree Ceiba grandiflora. Oecologia 2003, 135, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.L.; Lugo, A. Tropical secondary forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Kalacska, M.; Sanchez-azofeifa, G.A.; Calvo-alvarado, J.C.; Quesada, M.; Rivard, B.; Janzen, D.H. Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest. For. Ecol. Manag. 2004, 200, 227–247. [Google Scholar] [CrossRef]
- Madeira, B.G.; Espírito-Santo, M.M.; Neto, S.D.; Nunes, Y.R.F.; Azofeifa, G.A.S.; Fernandes, G.W.; Quesada, M. Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant Ecol. 2009, 201, 291–304. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Scariot, A. Principles of natural regeneration of tropical dry forests for restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef]
- Fenton, M.; Acharya, L.; Audet, D.; Hickey, M.; Merriman, C.; Obrist, M.; Syme, D.; Adkins, B. Phyllostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the Neotropics. Biotropica 1992, 24, 440–446. [Google Scholar] [CrossRef]
- Medellin, R.A.; Equihua, M.; Amin, M.A. Bat diversity and abundance as indicators of disturbance in neotropical rainforests. Conserv. Biol. 2000, 14, 1666–1675. [Google Scholar] [CrossRef]
- Falcão, L.A.D.; do Espírito-Santo, M.M.; Leite, L.O.; Garro, R.N.S.L.; Avila-Cabadilla, L.D.; Stoner, K.E. Spatiotemporal variation in phyllostomid bat assemblages over a successional gradient in a tropical dry forest in southeastern Brazil. J. Trop. Ecol. 2014, 30, 123–132. [Google Scholar] [CrossRef]
- Klingbeil, B.T.; Willig, M.R. Guild-specific responses of bats to landscape composition and configuration in fragmented Amazonian rainforest. J. Appl. Ecol. 2009, 46, 203–213. [Google Scholar] [CrossRef]
- Henry, M.; Cosson, J.F.; Pons, J.M. Modelling multi-scale spatial variation in species richness from abundance data in a complex neotropical bat assemblage. Ecol. Model. 2010, 221, 2018–2027. [Google Scholar] [CrossRef]
- Avila-Cabadilla, L.D.; Sanchez-Azofeifa, G.A.; Stoner, K.E.; Alvarez-Añorve, M.Y.; Quesada, M.; Portillo-Quintero, C.A. Local and landscape factors determining occurrence of phyllostomid bats in tropical secondary forests. PLoS ONE 2012, 7, e35228. [Google Scholar] [CrossRef] [PubMed]
- Kalda, O.; Kalda, R.; Liira, J. Multi-scale ecology of insectivorous bats in agricultural landscapes. Agric. Ecosyst. Environ. 2015, 199, 105–113. [Google Scholar] [CrossRef]
- Cisneros, L.M.; Fagan, M.E.; Willig, M.R. Effects of human-modified landscapes on taxonomic, functional and phylogenetic dimensions of bat biodiversity. Divers. Distrib. 2015, 21, 523–533. [Google Scholar] [CrossRef]
- Gorresen, P.M.M.; Willig, M.R.M.R.; Strauss, R.E. Multivariate analysis of scale-dependent associations between bats and landscape structure. Ecol. Appl. 2005, 15, 2126–2136. [Google Scholar] [CrossRef]
- Gorresen, P.M.; Willig, M.R. Landscape responses of bats to habitat fragmentation in Atlantic forest of Paraguay. J. Mammal. 2004, 85, 688–697. [Google Scholar] [CrossRef]
- Sanchez-Azofeifa, G.A.; Quesada, M.; Rodriguez, J.P.; Nassar, J.M.; Stoner, K.E.; Castillo, A.; Garvin, T.; Zent, E.L.; Calvo-Alvarado, J.C.; Kalacska, M.E.R.; et al. Research priorities for neotropical dry forests. Biotropica 2005, 37, 477–485. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. Discuss. 2007, 4, 439–473. [Google Scholar] [CrossRef]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.O.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Espírito-Santo, M.M.; Sevilha, A.C.; Anaya, F.C.; Barbosa, R.; Fernandes, G.W.; Sanchez-Azofeifa, G.A.; Scariot, A.; De Noronha, S.E.; Sampaio, C.A. Sustainability of tropical dry forests: Two case studies in southeastern and central Brazil. For. Ecol. Manag. 2009, 258, 922–930. [Google Scholar] [CrossRef]
- Crawley, M. The R Book; Wiley: Hoboken, NJ, USA, 2007; ISBN 978-0-47-051024-7. [Google Scholar]
- R Development Team. R: A Language and Environment for Statistical Computing; R Development Team: Vienna, Austria, 2015. [Google Scholar]
- Avila-Cabadilla, L.D.; Stoner, K.E.; Henry, M.; Añorve, M.Y.A. Composition, structure and diversity of phyllostomid bat assemblages in different successional stages of a tropical dry forest. For. Ecol. Manag. 2009, 258, 986–996. [Google Scholar] [CrossRef]
- Bernard, E.; Fenton, M.B. Bat mobility and roosts in a fragmented landscape in Central Amazonia, Brazil. Biotropica 2003, 35, 262–277. [Google Scholar] [CrossRef]
- Lundberg, J.; Moberg, F. Mobile link organisms and ecosystem functioning: Implications for ecosystem resilience and management. Ecosystems 2003, 6, 0087–0098. [Google Scholar] [CrossRef]
- Duncan, R.S.; Chapman, C.A. Seed dispersal and potential forest succession in abandoned agriculture in Tropical Africa. Ecol. Appl. 1999, 9, 998–1008. [Google Scholar] [CrossRef]
- Castro-Luna, A.A.; Sosa, V.J.; Castillo-Campos, G. Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Anim. Conserv. 2007, 10, 219–228. [Google Scholar] [CrossRef]
- Ethier, K.; Fahrig, L. Positive effects of forest fragmentation, independent of forest amount, on bat abundance in eastern Ontario, Canada. Landsc. Ecol. 2011, 26, 865–876. [Google Scholar] [CrossRef]
- Caras, T.; Korine, C. Effect of vegetation density on the use of trails by bats in a secondary tropical rain forest. J. Trop. Ecol. 2009, 25, 97–101. [Google Scholar] [CrossRef]
- Falcão, L.A.D.; Alfaro, L.D.; Fernandes, G.W.; Leite, L.O.; Neves, F.S.; Reyes, P.C. Comunidad de murciélagos filostómidos asociada a Caryocar brasiliense. Brenesia 2010, 73–74, 150–153. [Google Scholar]
- Castro, I.; Michalski, F. Effects of logging on bats in tropical forests. Nat. Conserv. 2014, 2, 99–105. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Guild | Successional Stages | Total | ||
|---|---|---|---|---|---|
| Early | Intermediate | Late | |||
| Subfamily Carollinae | |||||
| Carollia perspicillata | Frugivore | 54 | 27 | 68 | 149 |
| Desmodontinae | |||||
| Desmodus rotundus | Sanguinivore | 4 | 6 | 24 | 34 |
| Diphylla ecaudata | Sanguinivore | -- | -- | 3 | 3 |
| Glossophaginae | |||||
| Glossophaga soricina | Nectarivore | 3 | 5 | 5 | 13 |
| Lonchophylla dekeyseri | Nectarivore | -- | 1 | 2 | 3 |
| Phyllostominae | |||||
| Chrotopterus auritus | Carnivore | -- | -- | 2 | 2 |
| Micronycteris sanborni | Insectivore | 1 | -- | 3 | 4 |
| Mimon benneti | Insectivore | -- | -- | 2 | 2 |
| Mimon crenulatum | Insectivore | -- | 2 | 11 | 13 |
| Phylloderma stenops | Omnivore | -- | 2 | 2 | 4 |
| Phyllostomus discolor | Omnivore | -- | 6 | 5 | 11 |
| Phyllostomus hastatus | Omnivore | -- | 3 | 4 | 7 |
| Tonatia bidens | Insectivore | -- | -- | 1 | 1 |
| Stenodermatinae | |||||
| Artibeus lituratus | Frugivore | 1 | 1 | 4 | 6 |
| Artibeus planirostris | Frugivore | 24 | 57 | 123 | 204 |
| Plathyrrinus lineatus | Frugivore | 1 | 1 | 2 | 4 |
| Uroderma magnirostrum | Frugivore | -- | -- | 1 | 1 |
| Total | 88 | 111 | 262 | 461 | |
| Response Variable | Explanatory Variable | Df | Deviance | Residual Df | Residual Deviance | p * |
|---|---|---|---|---|---|---|
| Abundance | ||||||
| Local scale | ||||||
| Plant richness | 1 | 2.076 | 10 | 209.21 | 0.73 | |
| Canopy height | 1 | 42.734 | 9 | 166.48 | 0.14 | |
| Plant density | 1 | 44.123 | 8 | 122.36 | 0.14 | |
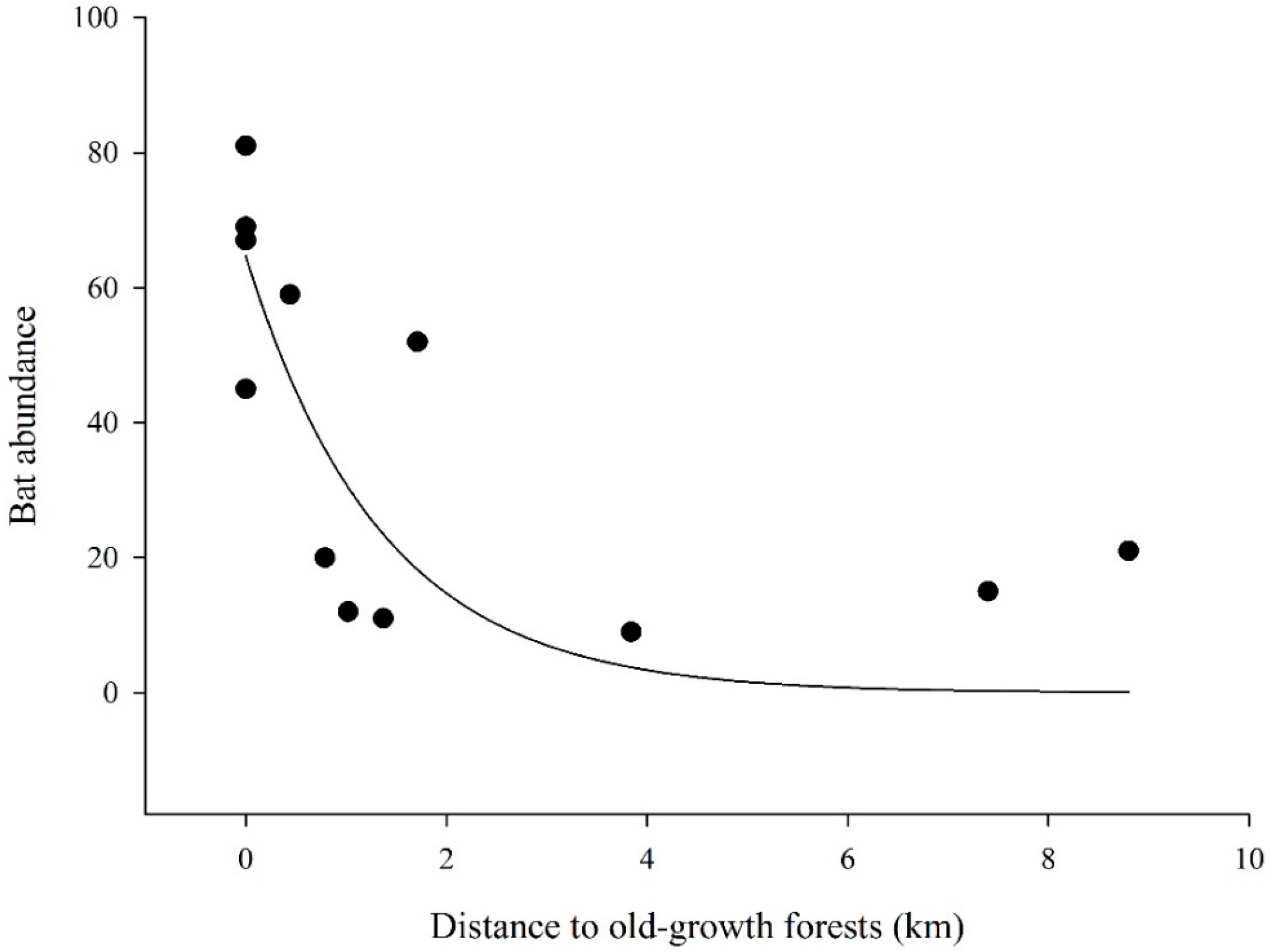
| Distance to old-growth forests | 1 | 78.364 | 10 | 132.92 | 0.02 (-) | |
| Landscape scale | ||||||
| Plant richness | 1 | 0.069 | 2 | 67.028 | 0.96 | |
| Canopy height | 1 | 0.576 | 2 | 66.521 | 0.91 | |
| Plant density | 1 | 20.967 | 2 | 46.131 | 0.42 | |
| Forest cover | 1 | 4.153 | 2 | 62.944 | 0.12 | |
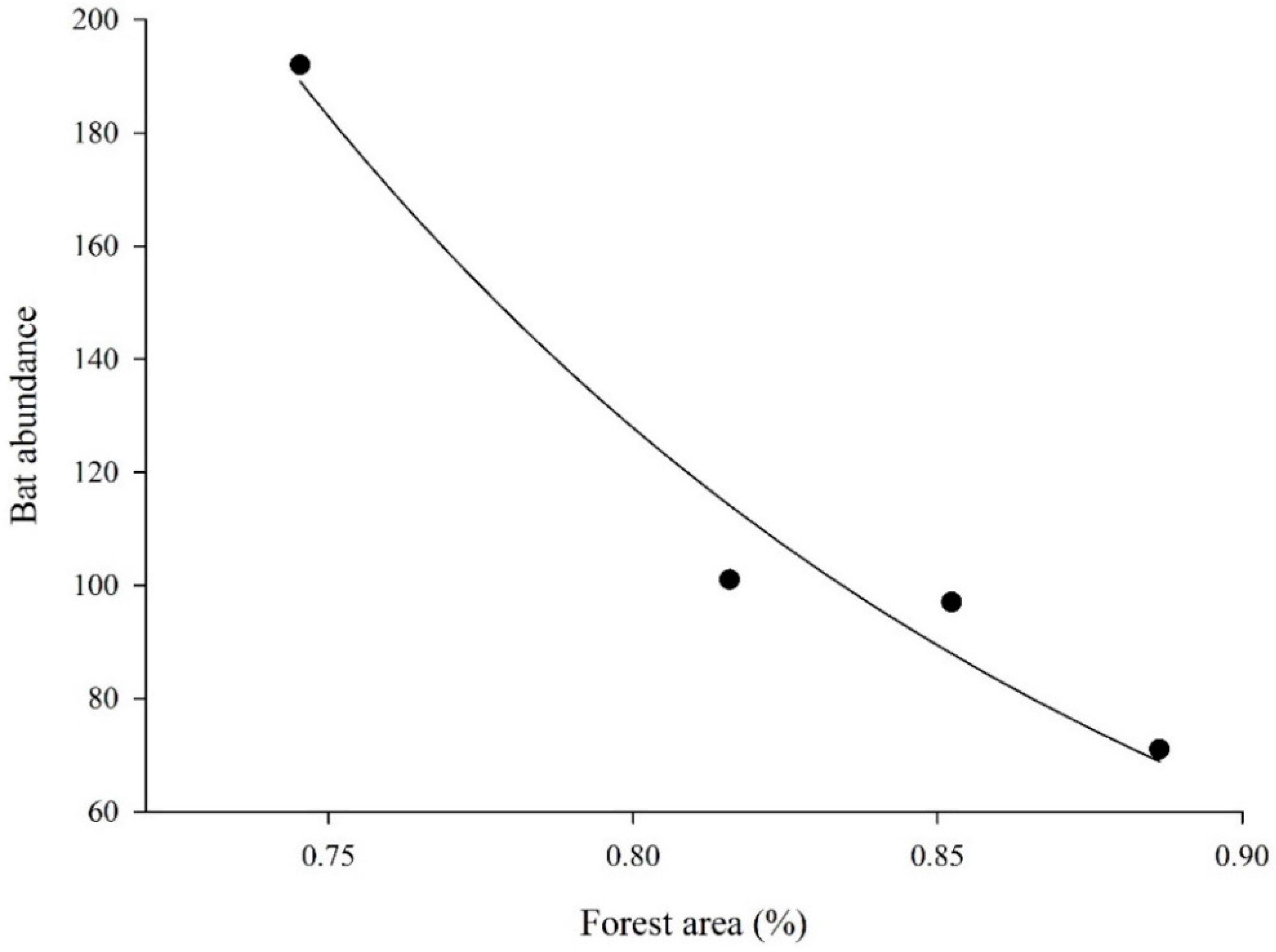
| Forest cover at 1 km | 1 | 64.578 | 2 | 2.52 | 0.01 (-) | |
| Forest cover at 5 km | 1 | 50.305 | 2 | 16.792 | 0.12 | |
| Richness | ||||||
| Local scale | ||||||
| Plant richness | 1 | 0.003 | 10 | 26.553 | 0.99 | |
| Canopy height | 1 | 8.558 | 9 | 17.995 | 0.08 | |
| Plant density | 1 | 0.151 | 8 | 17.843 | 0.80 | |
| Distance to old-growth forests | 1 | 2.44 | 10 | 24.114 | 0.32 | |
| Landscape scale | ||||||
| Plant richness | 1 | 1.9091 | 2 | 1.46 | 0.25 | |
| Canopy height | 1 | 0.565 | 2 | 2.8 | 0.58 | |
| Plant density | 1 | 0.346 | 2 | 3.022 | 0.68 | |
| Forest cover | 1 | 0.145 | 2 | 3.224 | 0.78 | |
| Forest cover at 1 km | 1 | 0.379 | 2 | 2.99 | 0.66 | |
| Forest cover at 5 km | 1 | 1.395 | 2 | 1.974 | 0.35 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falcão, L.A.D.; Espírito-Santo, M.M.; Fernandes, G.W.; Paglia, A.P. Effects of Habitat Structure, Plant Cover, and Successional Stage on the Bat Assemblage of a Tropical Dry Forest at Different Spatial Scales. Diversity 2018, 10, 41. https://doi.org/10.3390/d10020041
Falcão LAD, Espírito-Santo MM, Fernandes GW, Paglia AP. Effects of Habitat Structure, Plant Cover, and Successional Stage on the Bat Assemblage of a Tropical Dry Forest at Different Spatial Scales. Diversity. 2018; 10(2):41. https://doi.org/10.3390/d10020041
Chicago/Turabian StyleFalcão, Luiz A. D., Mário M. Espírito-Santo, G. W. Fernandes, and Adriano P. Paglia. 2018. "Effects of Habitat Structure, Plant Cover, and Successional Stage on the Bat Assemblage of a Tropical Dry Forest at Different Spatial Scales" Diversity 10, no. 2: 41. https://doi.org/10.3390/d10020041
APA StyleFalcão, L. A. D., Espírito-Santo, M. M., Fernandes, G. W., & Paglia, A. P. (2018). Effects of Habitat Structure, Plant Cover, and Successional Stage on the Bat Assemblage of a Tropical Dry Forest at Different Spatial Scales. Diversity, 10(2), 41. https://doi.org/10.3390/d10020041