The Metal-Dependent Regulators FurA and FurB from Mycobacterium Tuberculosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Discussion
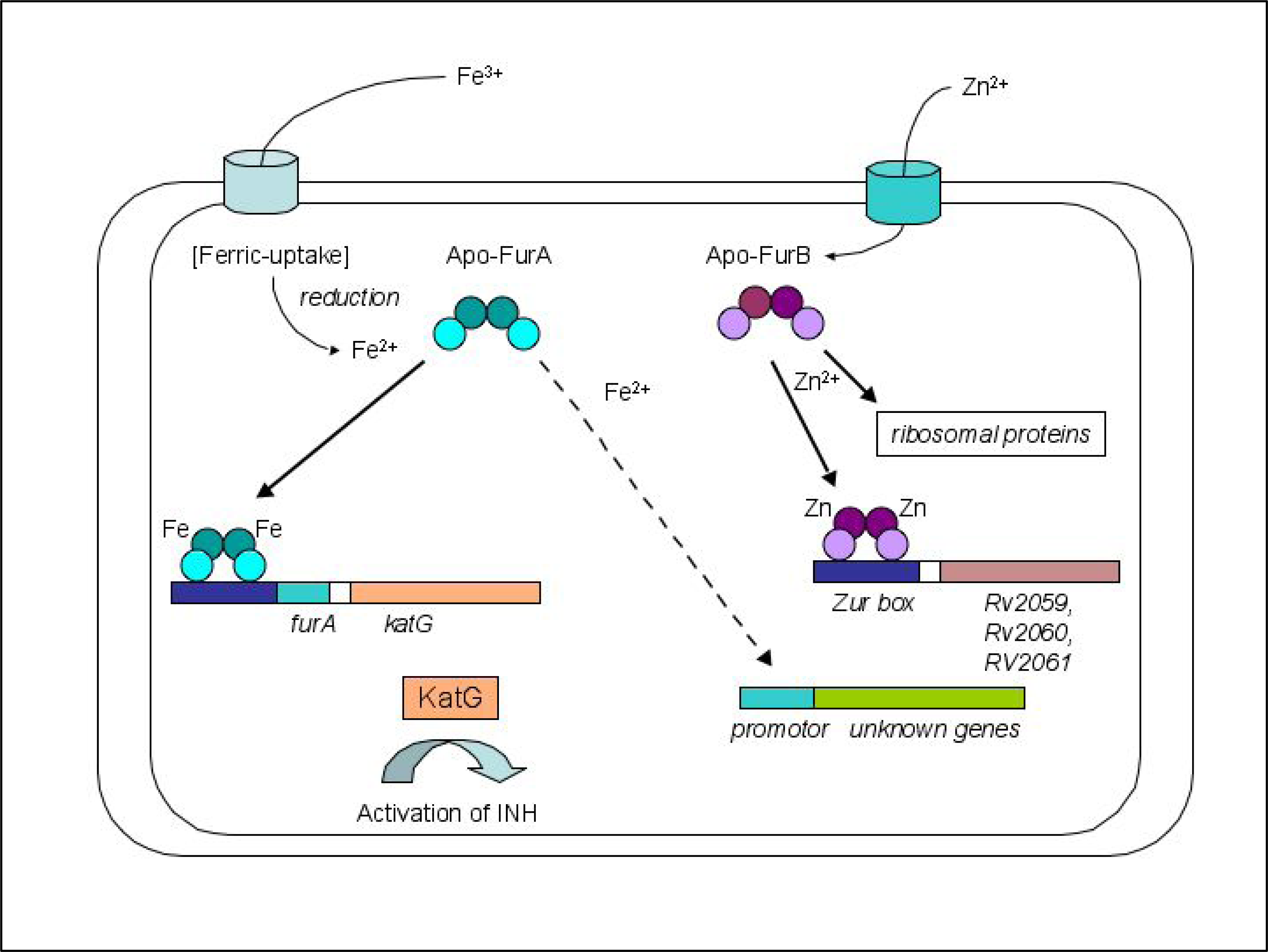
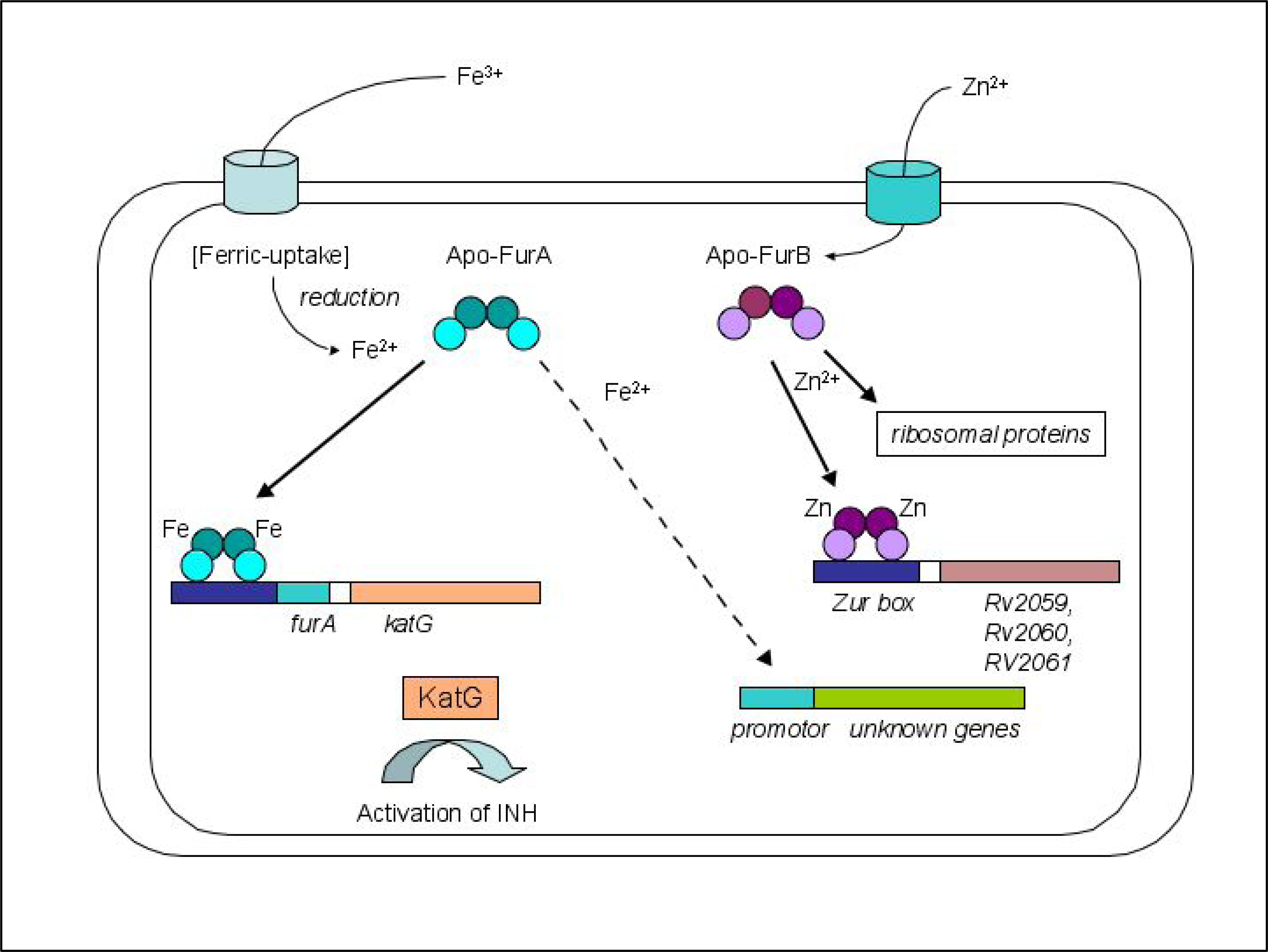
2.1. Biological role of FurA and FurB in M. tuberculosis
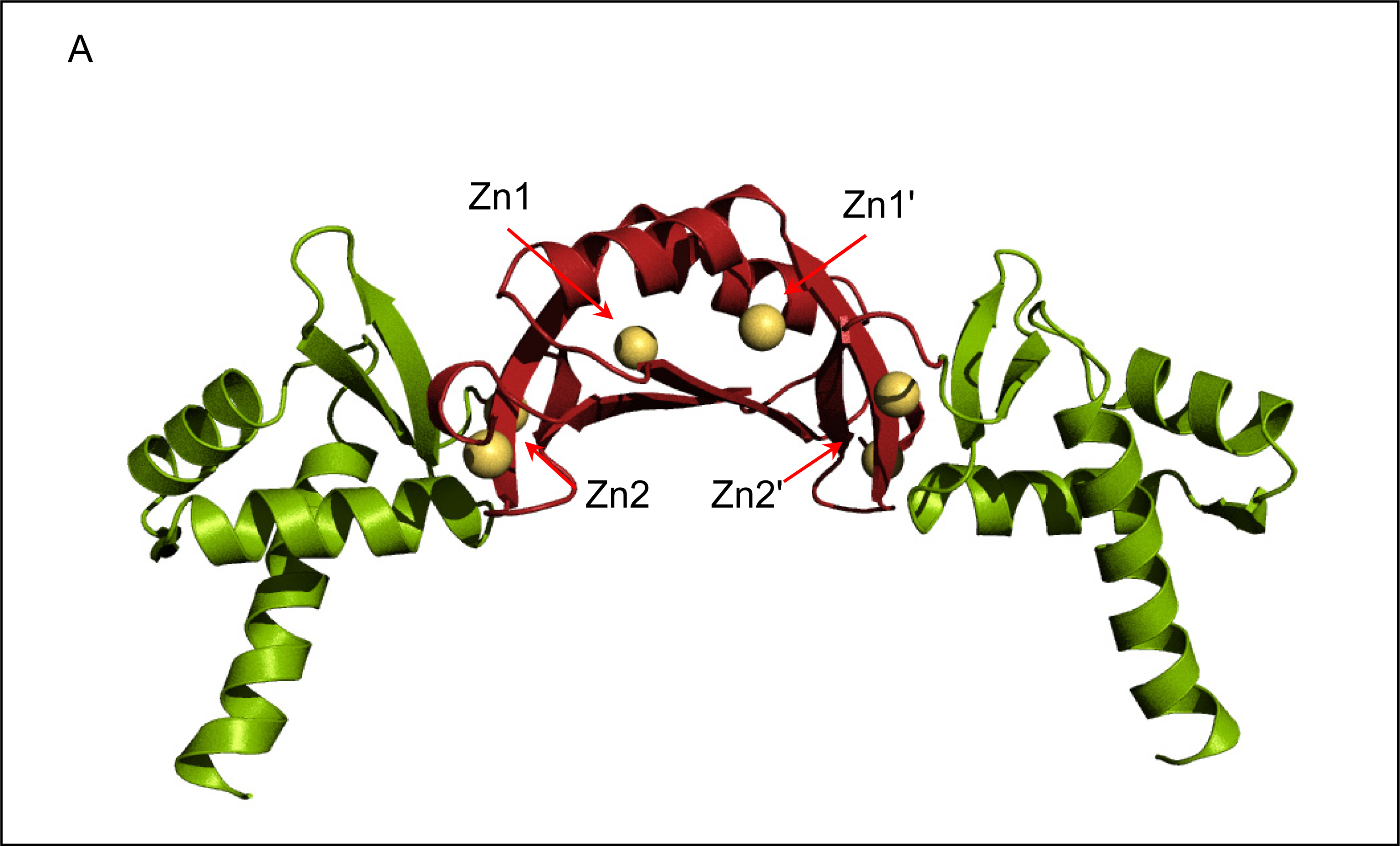
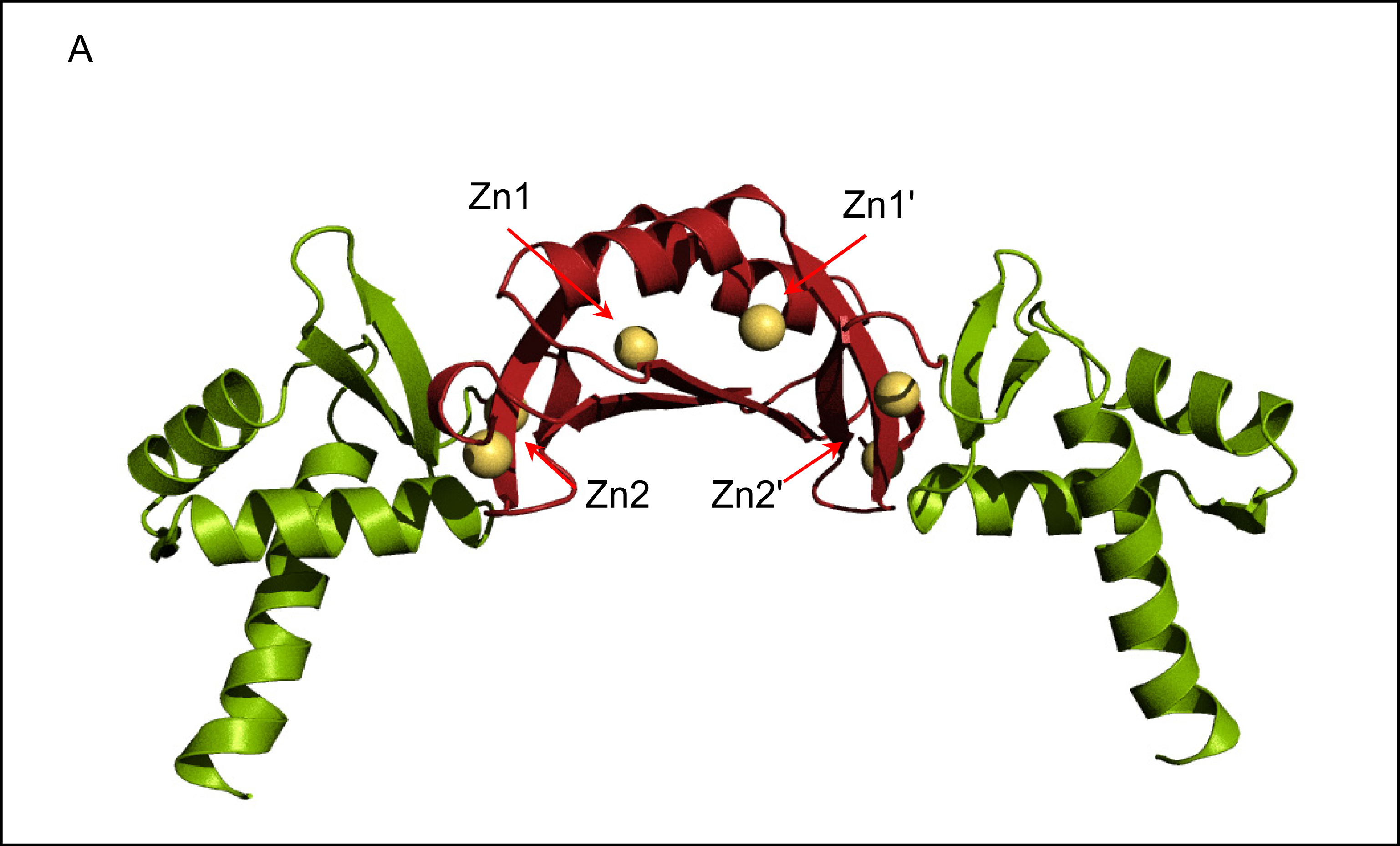
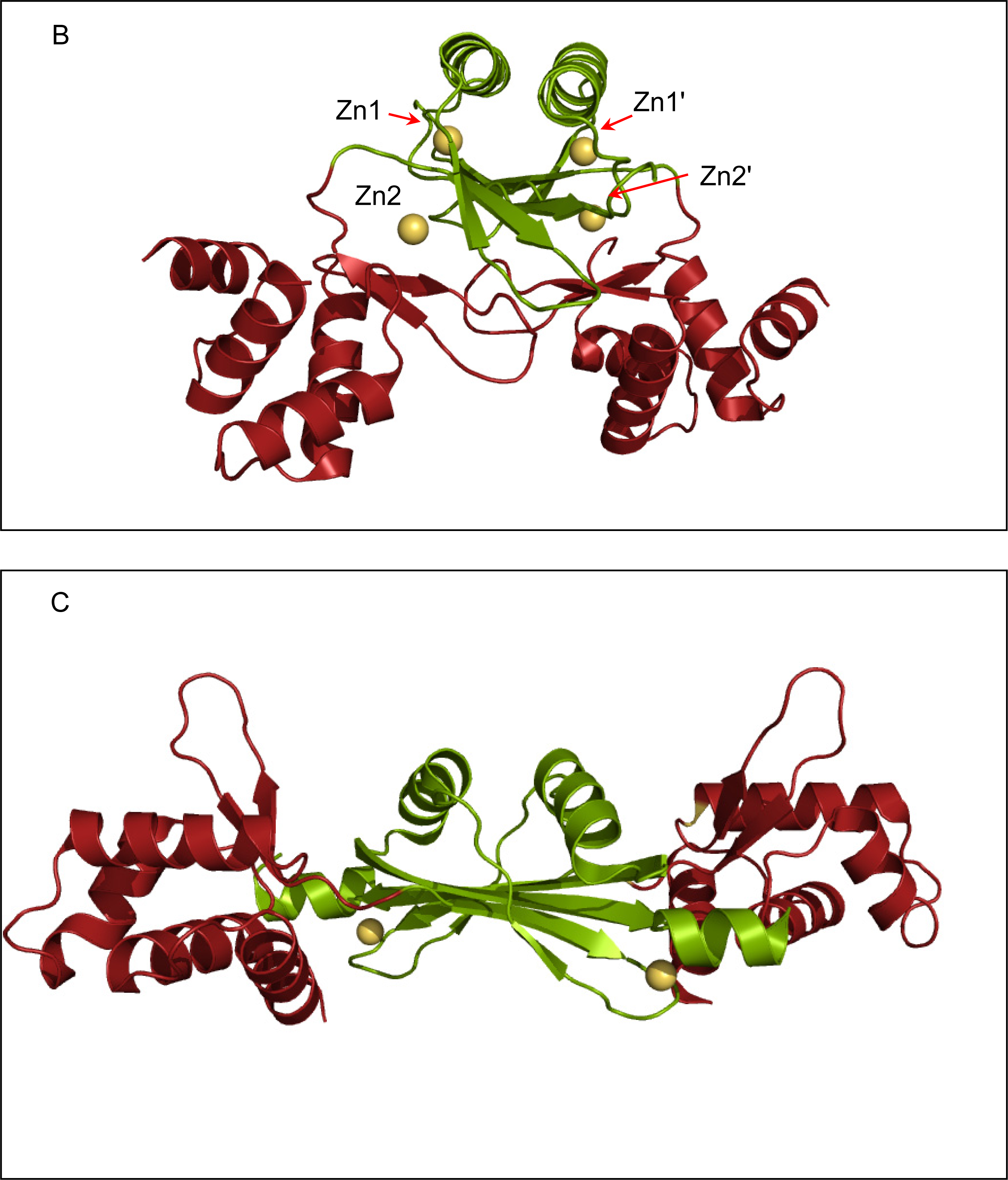
2.2. Crystal structure of FurB
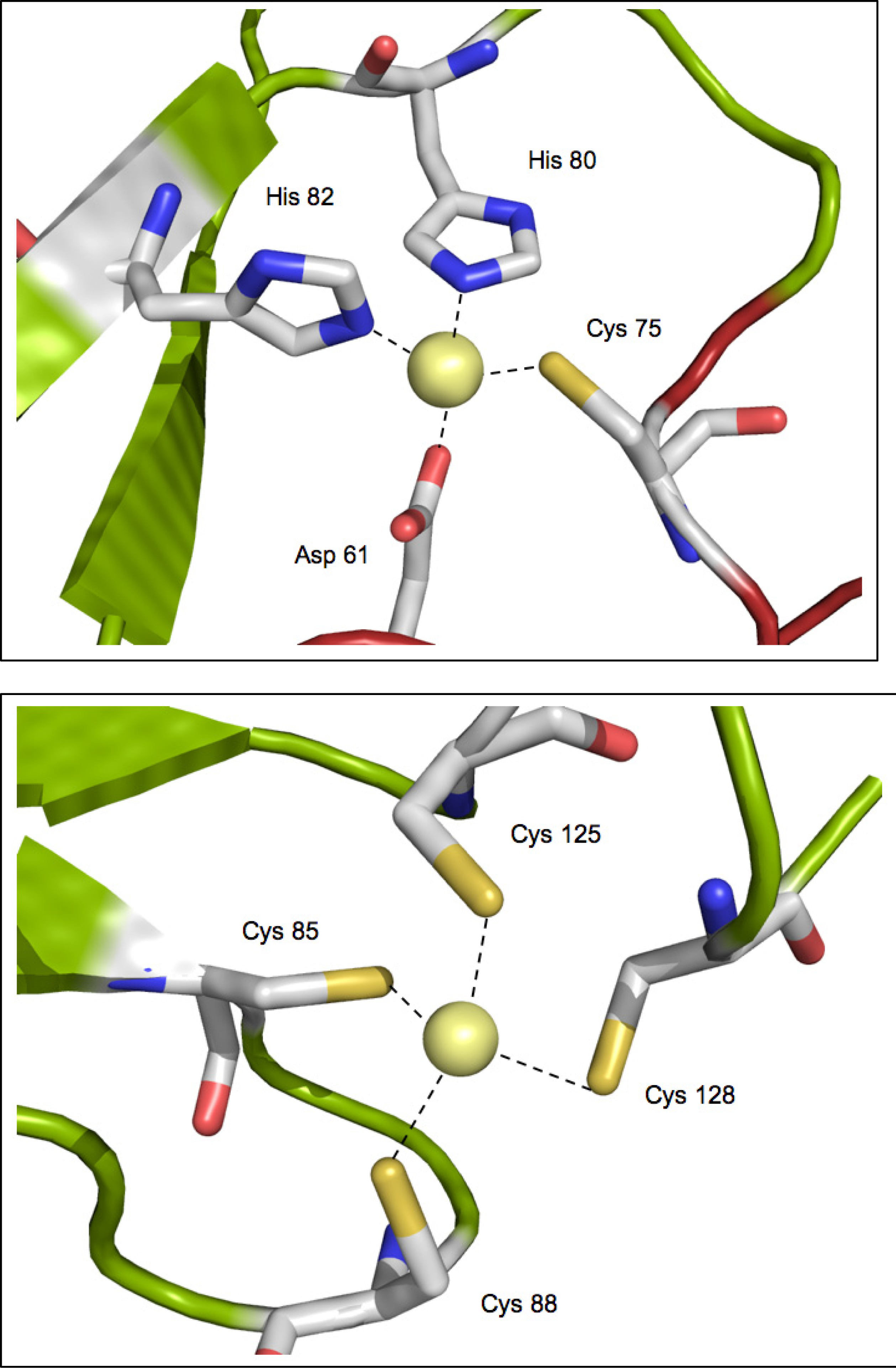
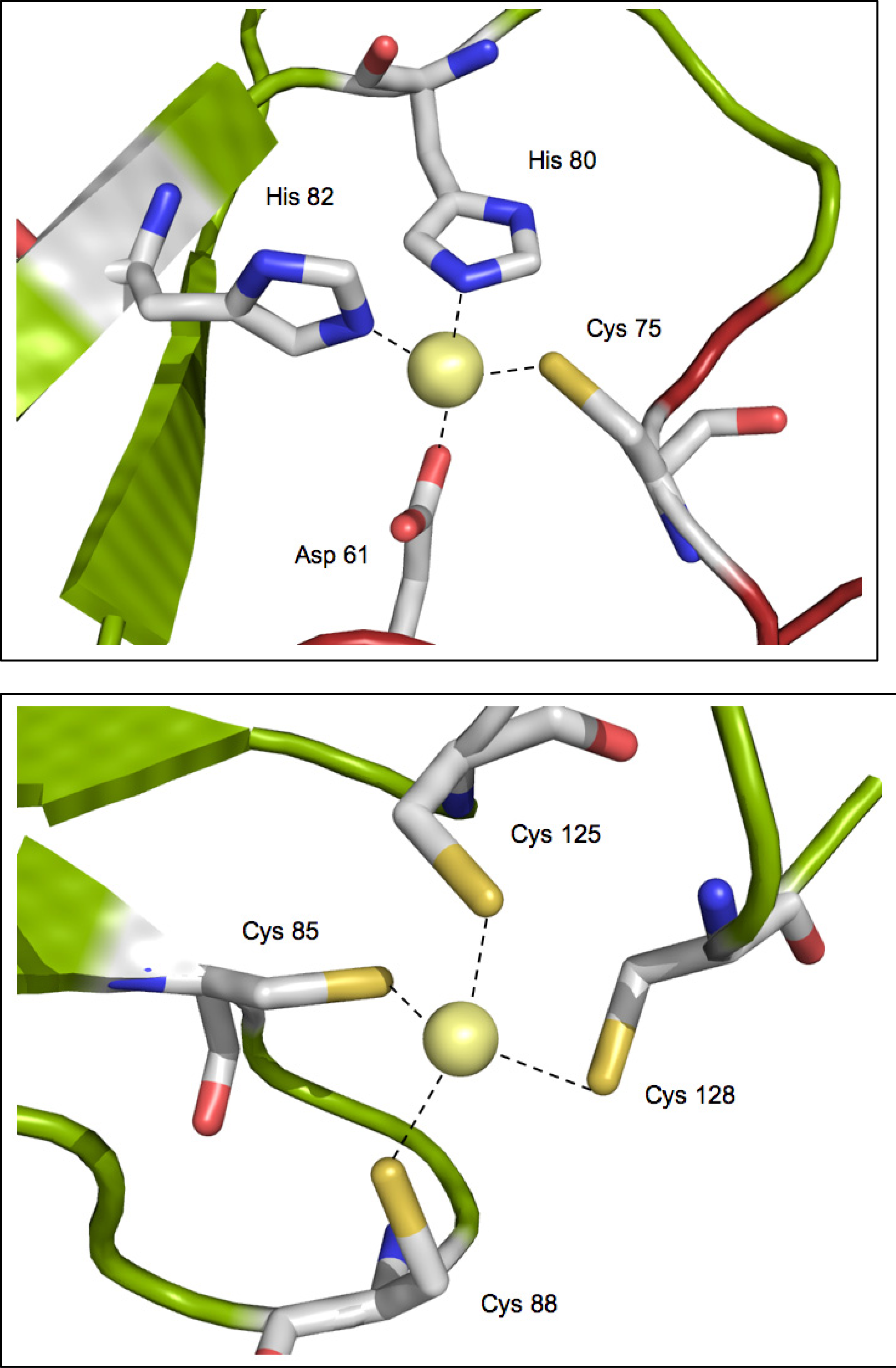
2.3. Metal binding sites and metal selectivity
3. Conclusions
Acknowledgments
References
- WHO. Report on tuberculosis: global tuberculosis control-surveillance, planning, financing. World Health Organization: Geneva, Switzerland; Volume 2008.
- Zignol, M; Hosseini, MS; Wright, A; Weezenbeek, CL; Nunn, P; Watt, CJ; Williams, BG; Dye, C. Global incidence of multidrug-resistant tuberculosis. J. Infect. Dis 2006, 194, 479–485. [Google Scholar]
- Nelson, N. Metal ion transporters and homeostasis. EMBO J 1999, 18, 4361–4371. [Google Scholar]
- Blencowe, DK; Morby, AP. Zn(II) metabolism in prokaryotes. FEMS Microbiol. Rev 2003, 27, 291–311. [Google Scholar]
- Agranoff, D; Krishna, S. Metal ion transport and regulation in Mycobacterium tuberculosis. Front. Biosci 2004, 9, 2996–3006. [Google Scholar]
- Braun, V. Iron uptake mechanisms and their regulation in pathogenic bacteria. Int. J. Med. Microbiol 2001, 291, 67–79. [Google Scholar]
- Cole, ST; Brosch, R; Parkhill, J; Garnier, T; Churcher, C; Harris, D; Gordon, SV; Eiglmeier, K; Gas, S; Barry, CE, 3rd; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar]
- Rodriguez, GM; Smith, I. Mechanisms of iron regulation in mycobacteria: role in physiology and virulence. Mol. Microbiol 2003, 47, 1485–1494. [Google Scholar]
- Rodriguez, G. Control of iron metabolism in Mycobacterium tuberculosis. Trends. Microbiol 2006, 14, 320–327. [Google Scholar]
- Pohl, E; Holmes, RK; Hol, WG. Crystal structure of the iron-dependent regulator (IdeR) from Mycobacterium tuberculosis shows both metal binding sites fully occupied. J. Mol. Biol 1999, 285, 1145–1156. [Google Scholar]
- Wisedchaisri, G; Holmes, RK; Hol, WG. Crystal structure of an IdeR-DNA complex reveals a conformational change in activated IdeR for base-specific interactions. J. Mol. Biol 2004, 342, 1155–1169. [Google Scholar]
- Feese, MD; Ingason, BP; Goranson-Siekierke, J; Holmes, RK; Hol, WG. Crystal structure of the iron-dependent regulator from Mycobacterium tuberculosis at 2.0-A resolution reveals the Src homology domain 3-like fold and metal binding function of the third domain. J. Biol. Chem 2001, 276, 5959–5965. [Google Scholar]
- Hill, PJ; Cockayne, A; Landers, P; Morrissey, JA; Sims, CM; Williams, P. SirR, a novel iron-dependent repressor in Staphylococcus epidermidis. Infect. Immun 1998, 66, 4123–4129. [Google Scholar]
- Morrissey, J; Cockayne, A; Hill, P; Williams, P. Molecular cloning and analysis of a putative siderophore ABC transporter from Staphylococcus aureus. Infect. Immun 2001, 68, 6281–6288. [Google Scholar]
- Hantke, K. Regulation of ferric iron transport in Escherichia coli K12: isolation of a constitutive mutant. Mol. Gen. Genet 1981, 182, 288–292. [Google Scholar]
- Hantke, K. Iron and metal regulation in bacteria. Curr. Opin. Microbiol 2001, 4(2), 172–177. [Google Scholar]
- Vasil, ML; Ochsner, UA. The response of Pseudomonas aeruginosa to iron: genetics, biochemistry and virulence. Mol. Microbiol 1999, 34, 399–413. [Google Scholar]
- Lee, JW; Helmann, JD. Functional specialization within the Fur family of metalloregulators. Biometals 2007, 20, 485–499. [Google Scholar]
- Vasil, ML. How we learnt about iron acquisition in Pseudomonas aeruginosa: a series of very fortunate event. Biometals 2007, 20, 587–601. [Google Scholar]
- Bagg, A; Neilands, JB. Molecular mechanism of regulation of siderophore-mediated iron assimilation. Microbiol. Rev 1987, 51, 509–518. [Google Scholar]
- Jacquamet, L; Aberdam, D; Adrait, A; Hazemann, JL; Latour, JM; Michaud-Soret, I. X-ray absorption spectroscopy of a new zinc site in the fur protein from Escherichia coli. Biochemistry 1998, 37, 2564–2571. [Google Scholar]
- Althaus, EW; Outten, CE; Olson, KE; Cao, H; O'Halloran, TV. The ferric uptake regulation (Fur) repressor is a zinc metalloprotein. Biochemistry 1999, 38, 6559–6569. [Google Scholar]
- Zheleznova, EE; Crosa, JH; Brennan, RG. Characterization of the DNA- and metal-binding properties of Vibrio anguillarum fur reveals conservation of a structural Zn(2+) ion. J. Bacteriol 2000, 182, 6264–6267. [Google Scholar]
- Patzer, SI; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol 1998, 28, 1199–1210. [Google Scholar]
- Ahn, BE; Cha, J; Lee, EJ; Han, AR; Thompson, CJ; Roe, JH. Nur, a nickel-responsive regulator of the Fur family, regulates superoxide dismutases and nickel transport in Streptomyces coelicolor. Mol. Microbiol 2006, 59, 1848–1858. [Google Scholar]
- Diaz-Mireles, E; Wexler, M; Sawers, G; Bellini, D; Todd, JD; Johnston, AW. The Fur-like protein Mur of Rhizobium leguminosarum is a Mn(2+)-responsive transcriptional regulator. Microbiology 2004, 150, 1447–1456. [Google Scholar]
- Masse, E; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar]
- Davis, BM; Quinones, M; Pratt, J; Ding, Y; Waldor, MK. Characterization of the small untranslated RNA RyhB and its regulon in Vibrio cholerae. J. Bacteriol 2005, 187, 4005–4014. [Google Scholar]
- Wilderman, PJ; Sowa, NA; FitzGerald, DJ; FitzGerald, PC; Gottesman, S; Ochsner, UA; Vasil, ML. Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar]
- Mellin, JR; Goswami, S; Grogan, S; Tjaden, B; Genco, CA. A novel fur-and iron-regulated small RNA, NrrF, is required for indirect fur-mediated regulation of the sdhA and sdhC genes in Neisseria meningitidis. J. Bacteriol 2007, 189, 3686–3694. [Google Scholar]
- Massé, E; Salvaila, H; Desnoyersa, G; Arguin, M. Small RNAs controlling iron metabolism. Curr. Opin. Microbiol 2007, 10, 140–145. [Google Scholar]
- Pym, AS; Domenech, P; Honore, N; Song, J; Deretic, V; Cole, ST. Regulation of catalase-peroxidase (KatG) expression, isoniazid sensitivity and virulence by furA of Mycobacterium tuberculosis. Mol. Microbiol 2001, 40, 879–889. [Google Scholar]
- Sala, C; Forti, F; Di, Florio E; Canneva, F; Milano, A; Riccardi, G; Ghisotti, D. Mycobacterium tuberculosis FurA autoregulates its own expression. J. Bacteriol 2003, 185, 5357–5362. [Google Scholar]
- Mongkolsuk, S; Helmann, JD. Regulation of inducible peroxide stress responses. Mol. Microbiol 2002, 45, 9–15. [Google Scholar]
- Lee, JW; Helmann, JD. Biochemical characterization of the structural Zn2+ site in the Bacillus subtilis peroxide sensor PerR. J. Biol. Chem 2006, 281, 23567–23578. [Google Scholar]
- Lee, JW; Helmann, JD. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature 2006, 440, 363–367. [Google Scholar]
- Marrakchi, H; Laneelle, G; Quemard, A. InhA, a target of the antituberculous drug isoniazid, is involved in a mycobacterial fatty acid elongation system, FAS-II. Microbiology 2000, 146, 289–296. [Google Scholar]
- Vilchèze, C; Jacobs, WRJ. The mechanism of isoniazid killing: clarity through the scope of genetics. Annu. Rev. Microbiol 2007, 61, 35–50. [Google Scholar]
- Timmins, GS; Deretic, V. Mechanisms of action of isoniazid. Mol. Microbiol 2006, 62, 1220–1227. [Google Scholar]
- Zhang, M; Yue, J; Yang, YP; Zhang, HM; Lei, JQ; Jin, RL; Zhang, XL; Wang, HH. Detection of mutations associated with isoniazid resistance in Mycobacterium tuberculosis isolates from China. J. Clin. Microbiol 2005, 43, 5477–5482. [Google Scholar]
- Canneva, F; Branzoni, M; Riccardi, G; Provvedi, R; Milano, A. Rv2358 and FurB: two transcriptional regulators from Mycobacterium tuberculosis which respond to zinc. J. Bacteriol 2005, 187, 5837–5840. [Google Scholar]
- Maciag, A; Dainese, E; Rodriguez, GM; Milano, A; Provvedi, R; Pasca, MR; Smith, I; Palu, G; Riccardi, G; Manganelli, R. Global analysis of Mycobacterium tuberculosis Zur (FurB) regulon. J. Bacteriol 2006, 189, 730–740. [Google Scholar]
- Lucarelli, D; Russo, S; Garman, E; Milano, A; Meyer-Klaucke, W; Pohl, E. Crystal structure and function of the zinc uptake regulator FurB from Mycobacterium tuberculosis. J. Biol. Chem 2007, 282, 9914–9922. [Google Scholar]
- Panina, E; Mironov, A; Gelfand, M. Comparative genomics of bacterial zinc regulons: enhanced ion transport, pathogenesis, and rearrangement of ribosomal proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 9912–9917. [Google Scholar]
- Larkin, M; Blackshields, G; Brown, N; Chenna, R; McGettigan, P; McWilliam, H; Valentin, F; Wallace, I; Wilm, A; Lopez, R; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- Traoré, DA; El Ghazouani, A; Ilango, S; Dupuy, J; Jacquamet, L; Ferrer, JL; Caux-Thang, C; Duarte, V; Latour, JM. Crystal structure of the apo-PerR-Zn protein from Bacillus subtilis. Mol. Microbiol 2006, 61, 1211–1219. [Google Scholar]
- Pohl, E; Haller, JC; Mijovilovich, A; Meyer-Klaucke, W; Garman, E; Vasil, ML. Architecture of a protein central to iron homeostasis: crystal structure and spectroscopic analysis of the ferric uptake regulator. Mol. Microbiol 2003, 47, 903–915. [Google Scholar]
- Pecqueu, L; D'Autréaux, B; Dupuy, J; Nicolet, Y; Jacquamet, L; Brutscher, B; Michaud-Soret, I; Bersch, B. Structural changes of Escherichia coli ferric uptake regulator during metal-dependent dimerization and activation explored by NMR and X-ray crystallography. J. Biol. Chem 2006, 281, 21286–21295. [Google Scholar]
- Pohl, E; Holmes, RK; Hol, WG. Motion of the DNA-binding domain with respect to the core of the diphtheria toxin repressor (DtxR) revealed in the crystal structures of apo- and holo-DtxR. J. Biol. Chem 1998, 273, 22420–22427. [Google Scholar]
- .
- Harding, MM. Geometry of metal-ligand interactions in proteins. Acta Crystallogr. D. Biol. Crystallogr 2001, 57, 401–411. [Google Scholar]
- Auld, DS. Zinc coordination sphere in biochemical zinc sites. Biometals 2001, 14, 271–313. [Google Scholar]
- Garman, EF; Grime, GW. Elemental analysis of proteins by microPIXE. Prog. Biophys. Mol. Biol 2005, 89, 173–205. [Google Scholar]
- Outten, CE; Tobin, DA; Penner-Hahn, JE; O'Halloran, TV. Characterization of the metal receptor sites in Escherichia coli Zur, an ultrasensitive zinc(II) metalloregulatory protein. Biochemistry 2001, 40, 10417–10423. [Google Scholar]

© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lucarelli, D.; Vasil, M.L.; Meyer-Klaucke, W.; Pohl, E. The Metal-Dependent Regulators FurA and FurB from Mycobacterium Tuberculosis. Int. J. Mol. Sci. 2008, 9, 1548-1560. https://doi.org/10.3390/ijms9081548
Lucarelli D, Vasil ML, Meyer-Klaucke W, Pohl E. The Metal-Dependent Regulators FurA and FurB from Mycobacterium Tuberculosis. International Journal of Molecular Sciences. 2008; 9(8):1548-1560. https://doi.org/10.3390/ijms9081548
Chicago/Turabian StyleLucarelli, Debora, Michael L. Vasil, Wolfram Meyer-Klaucke, and Ehmke Pohl. 2008. "The Metal-Dependent Regulators FurA and FurB from Mycobacterium Tuberculosis" International Journal of Molecular Sciences 9, no. 8: 1548-1560. https://doi.org/10.3390/ijms9081548
APA StyleLucarelli, D., Vasil, M. L., Meyer-Klaucke, W., & Pohl, E. (2008). The Metal-Dependent Regulators FurA and FurB from Mycobacterium Tuberculosis. International Journal of Molecular Sciences, 9(8), 1548-1560. https://doi.org/10.3390/ijms9081548