Introduction
Polycyclic aromatic hydrocarbons (PAH) are organic compounds consisting of two or more fused 5- and 6-membered rings. They are introduced into natural environment mainly from natural and anthropogenic combustion processes. Some PAH, for example benzo(a)pyrene, are of toxicological concern because they are precursors to cancer-causing metabolites [
1,
2]. The toxic response may be caused by irreversible covalent binding between PAH and the biological substrates or receptors, which results in the production of activated intermediates that can bind covalently to DNA or proteins to form adducts [
3].
Photolysis transforms numerous organic pollutants to less toxic or nontoxic products. However, at the concentrations below their aqueous solubility limits some PAH become more toxic to aquatic organisms in the presence of solar or artificial irradiations [
4,
5]. Moreover, some PAH may be bioavailable but nontoxic to aquatic organisms; however, they may be absorbed by the organisms and then photo-transformed
in vivo with lethal results [
6,
7]. The mechanism of the photo-induced toxicity involves generation of reactive oxygen species and singlet oxygen [
8] or photomodification process [
9]. Thus, photolysis constitutes a force which alters the chemical structure, physical properties, concentration, or toxicity of a PAH compound, and may affect its bioavailability. Aqueous concentrations of lightweight PAH range between 1 and 100 μg/L [
10]. 1-Aminopyrene (1-AP), an amino-PAH compound, is a major metabolite during biotransformation of 1-nitropyrene by microflora in natural environments and in the guts of animals and humans [
11]. Under UV-A irradiation, 1-AP has been shown to cause light-induced DNA single strand cleavage [
12].
Humic substances (HS) are ubiquitous in natural environments. Among them, humic acid (HA) is the fraction of HS that is not soluble in water under acidic conditions (< pH 2), but is soluble at higher pH, with typical average molecular weight of 2,000 to 3,000 [
13]. HS have a relatively high content of radicals, presumably of the semiquinone type, which are more prominent in HA than the other components. Most of the solar energy absorbed by the HS is between 300 and 500 nm. The aquatic photolysis of PAH can be influenced in several ways by the HA contained in natural waters. In one way, direct photolysis can be inhibited because competitive light absorption by colored HS occurs. In this case, the HA play the role of light filter. In another way, HS can enhance the degradation of PAH under irradiation by producing radicals or oxidizing species. Absorption of light by HS can initiate a number of photochemical processes, producing peroxy radicals and hydroxyl radicals as well as hydrated electrons, hydrogen peroxide, singlet oxygen and superoxide [
14]. These chemical species can promote redox reactions. Together with HS, they can photosensitize nonpolar organics such as PAH and possibly accelerate their degradation and detoxification. However, HS can also act as the sinks of photochemically produced transient reactants [
15].
HS can function as surfactants with the ability to bind both hydrophobic and hydrophilic materials. This function in combination with their colloidal properties, makes HA effective agents in transporting both organic and inorganic contaminants in the environment. The HS in aquatic ecosystems have been shown to be important in influencing the bioavailability, toxicity and fate of organic xenobiotics [
13]. Complexation of a variety of organic chemicals with dissolved humic substances (DHS) has been shown to affect chemical bioavailability and toxicity in addition to influencing the biodegradation and mobility of chemicals in the environment. For example, Holman
et al. reported enhancement of biodegradation of PAH with the addition of a soil HA [
16]. DHS has also been demonstrated to reduce the bioconcentration of PAH by aquatic organisms [
17]. Weinstein and Oris [
18] have shown that HA interacts with the test PAH and result in significant decreases in their bioaccumulation and photoinduced toxicity to the test organisms. Conversely, in algal bioassays, DHS have been shown to increase the toxicity of several organic contaminants [
19].
Since the presence of HS in surface waters has significant influence on the transport and fate of PAH in natural environment, we must consider the influences of HS in order to successfully predict the fate and effects of PAH and implement remediation strategies. As we know, the global UV-irradiation is expected to increase due to the reduced ozone layer in the stratosphere. Therefore, it is of our interest to have a better understanding of the UV-mediated chemical processes occurring in natural humic waters and the influence of these processes on the fate and effects of PAH in natural aquatic environment. The main objective of this study was to compare the effect of a HA on the toxicity of 1-AP to natural aquatic bacterial assemblages in the presence and the absence of solar irradiation. In this study two bacterial bioassays, plate counting with spread plate method and heterotrophic mineralization of
14C-glucose, were used to measure the microbial ecotoxicity of 1-AP. Spread plate counting measures viability of heterotrophic bacteria and heterotrophic mineralization reflects general metabolic activity of bacterial assemblages, respectively [
20]. SAS (Cary, NC, USA) was used to analyze the interactions between and among the various parameters (i.e., irradiation, time of exposure, HA and 1-AP) involved in the experimentation.
Materials And Methods
Reagents
All chemical reagents utilized were of analytical grade and fresh solutions as well. Standard River HA was purchased from the International Humic Substances Society (IHSS). HA stock aqueous solution was prepared at 2.5 g/L concentration. Considering the environmental concentration of lightweight PAH and its water solubility, 1-AP (Aldrich Chemical Co., Milwaukee, WI) solution was prepared at 10 μM concentration in dimethylsulfoxide (DMSO, 4%; HPLC grade, Fisher Scientific). 14C-D-glucose in ethanol solution was prepared (S.A: 265 mCi/mmol; Moravek Biochemicals, Brea, CA) and about 104 dpm was added to each sample. Fresh river water was collected from the Ross Barnett Reservoir, Ridgeland, Mississippi. Its total organic carbon content (TOC) ranged from 30 to 60 ppm. TOC was measured by using a PASTEL UV Spectrophotometer (ESCOMAN, France). The pH of the water samples ranged from 7.0 to 7.5.
Photolysis process
The photolysis process was carried out in 150-mL Erlenmeyer quartz flasks without shaking (Quartz Scientific, Fairport Harbor, Ohio). Initially, all flasks contained 50 ml of sterile fresh river water. Four groups of flasks (triplicate samples for each group) were prepared by adding HA solution to reach three different concentrations: 0, 20, and 60 ppm. To two of the four groups, 1-AP was added to reach a final concentration of 0 μM and subsequently to the other two a final concentration of 10 μM. Then after, they were incubated in a running water tub under natural sunlight outdoors at noontime during sunny days in late October 2001 at Jackson, Mississippi (320 19’ N; 900 5’ W). The average UV-A light intensity of the solar irradiation was 2.4 + 0.4 mJ/cm2.sec. Two groups, containing 1-AP, 0 μM and 10 μM, were incubated for a period of 18 minutes and subsequently the other two for a period of 90 minutes. For outdoor photolysis experiment, the flasks were suspended in an outdoor tub that contained continuous running tap water for maintaining the water temperature at 27±3oC. The surface water level in the flask was about 3 cm below the surface of the cooling water. A research radiometer (model IL 1700, International Light Inc., Newburyport, MA) was placed beside the tub to measure UV-A irradiation. The irradiation was not significantly attenuated by the cooling tap water and the incubating flasks were assumed to receive the same amount of irradiation. Simultaneously, similar treatments were assembled for darkness incubation, strictly under the same conditions. Darkness was achieved by wrapping the flasks with a sheet of aluminum foil. Therefore, the experimental control samples included HA concentration (0 ppm), 1-AP concentration (0 μM), and exposure to darkness samples.
HPLC analysis of 1-AP
The analysis was conducted with a Waters 996 HPLC system equipped with a Waters 717 plus Autosampler, a photodiode array detector, a pre-column filter (Sigma-Aldrich, USA) and a Lichrospher® 100 RP-8 column (25 cm X 4.0 mm I.D., 5 µm) (Hewlett Packard, USA). Mixtures of acetonitrile (A) and water (B) were used as the mobile phase. The following gradient elution was run: 40 to 90% A from 0 to 10 min, held at 90% A from 10 to 22 min, 90 to 40% A from 22 to 24 min. The flow rate was 1.0 mL/min. The detection wavelength was 254 nm. The temperature of the autosampler was controlled at 12oC.
Microbial in vitro ecotoxicity assay
After the exposure to light or darkness, microorganisms were inoculated into the flasks by adding 25 mL of fresh river water containing approximately 106 cfu/mL of viable heterotrophic bacteria. Flasks were incubated overnight in darkness without shaking at room temperature (25oC). After incubation, aliquots were used to perform microbiological analysis.
Microbiological analysis
The influence of humic acids on 1-AP ecotoxicity during solar photolysis process was measured with two methods: (a) microbial spread plate counting and (b) microbial mineralization of 14C-D-glucose.
(a) Viable Count of Heterotrophic Bacteria by the Spread Plate Method
Aliquots of 2 mL of all replicates were obtained to determine the heterotrophic bacteria counting with the spread plate method. Serial dilutions were carried out on each sample and cultured on nutrient agar plates. Plates were incubated in a growth chamber for 24 hours at 25
oC before counting the viable colony forming units (cfu) developed on the plates. Results are summarized in
Table 1 and depicted in
Figure 1.
(b) Heterotrophic Microbial Mineralization of 14C-D-glucose
After sampling for heterotrophic microbial counting, radiolabeled
14C-D-glucose solution (~1 μg/L) was added into each sample. Flasks were capped with silicon stoppers and incubated without shaking for 1 hour at room temperature. Heterotrophic microbial mineralization of
14C-D-glucose was terminated with injection of 0.5 mL of 2N H
2SO
4 into each flask. The
14CO
2 generated (disintegration per minute; DPM) was trapped for 12 hours into filter papers soaked with β-phenylethylamine (Sigma, Milwaukee). Radioactivity on the filter papers was measured with liquid scintillation spectrometry (Packard Instrument; model TR 1600). Results are summarized in
Table 2 and depicted in
Figure 2.
Table 1.
Colony forming units per milliliter (cfu/mL)x104 (±STD) of viable heterotrophic bacteria after exposure to 1-AP in sunlight and darkness for 18 and 90 minutes in the presence or in the absence of HA.
Table 1.
Colony forming units per milliliter (cfu/mL)x104 (±STD) of viable heterotrophic bacteria after exposure to 1-AP in sunlight and darkness for 18 and 90 minutes in the presence or in the absence of HA.
| Conditions: | cfu/mL x 104 (± STD) |
| HA/1-AP/TC* | Light | Darkness |
| 0-0-18 | 3.0 | ± 2.5 | 5.7 | ± 4.0 |
| 20-0-18 | 8.3 | ± 6.3 | 7.5 | ± 2.0 |
| 60-0-18 | 22.5 | ± 0.8 | 22.2 | ±18.4 |
| 0-10-18 | 11.3 | ± 2.6 | 13.0 | ± 4.0 |
| 20-10-18 | 6.7 | ± 1.8 | 14.1 | ± 2.4 |
| 60-10-18 | 10.0 | ± 3.9 | 16.1 | ±13.5 |
| 0-0-90 | 6.8 | ± 4.3 | 6.1 | ± 3.0 |
| 20-0-90 | 13.3 | ± 2.0 | 12.3 | ± 3.8 |
| 60-0-90 | 19.4 | ±13.0 | 20.4 | ±14.0 |
| 0-10-90 | 4.1 | ± 0.7 | 5.0 | ± 3.4 |
| 20-10-90 | 4.5 | ± 1.1 | 5.5 | ± 2.4 |
| 60-10-90 | 12.0 | ± 2.3 | 13.3 | ± 2.9 |
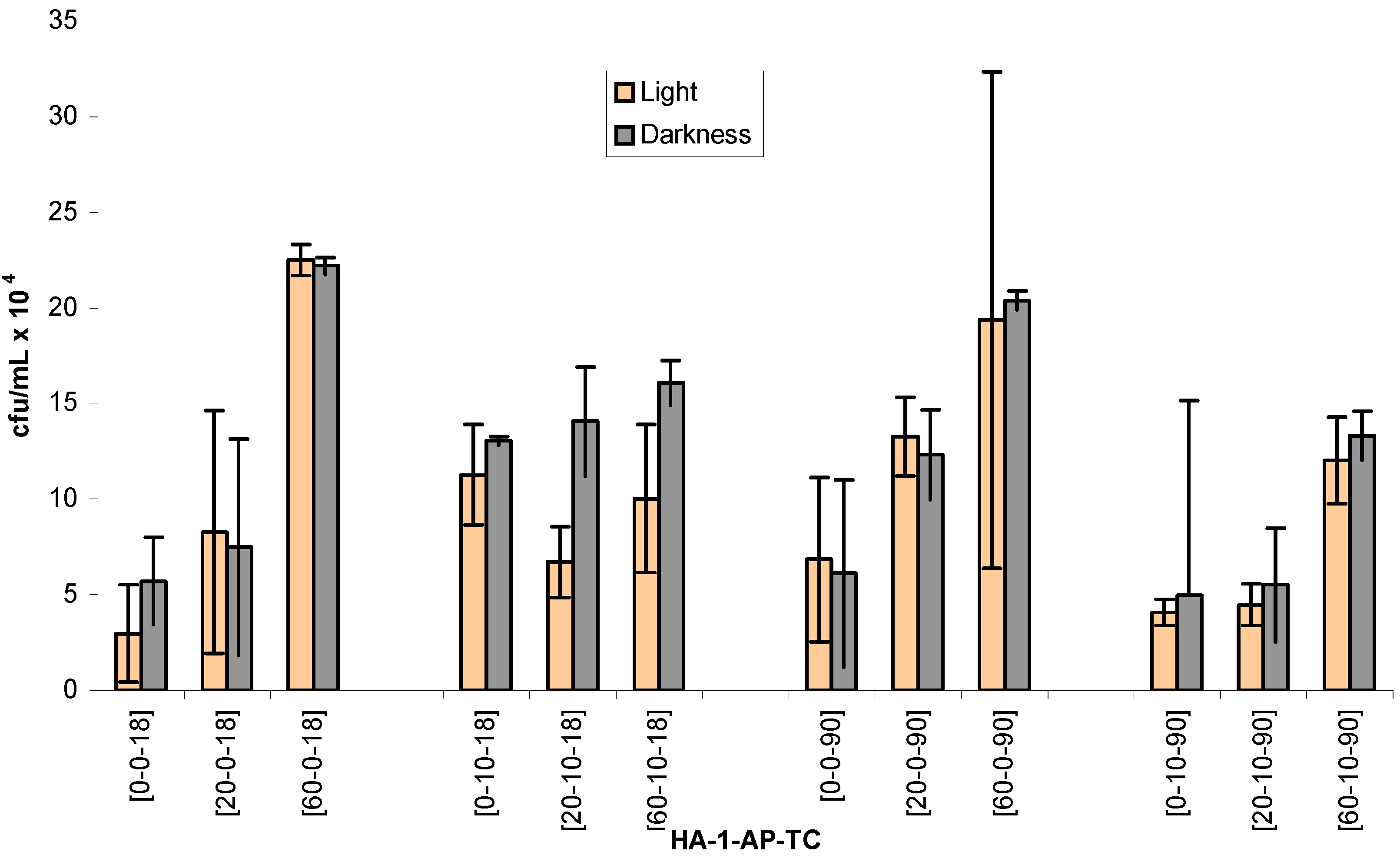
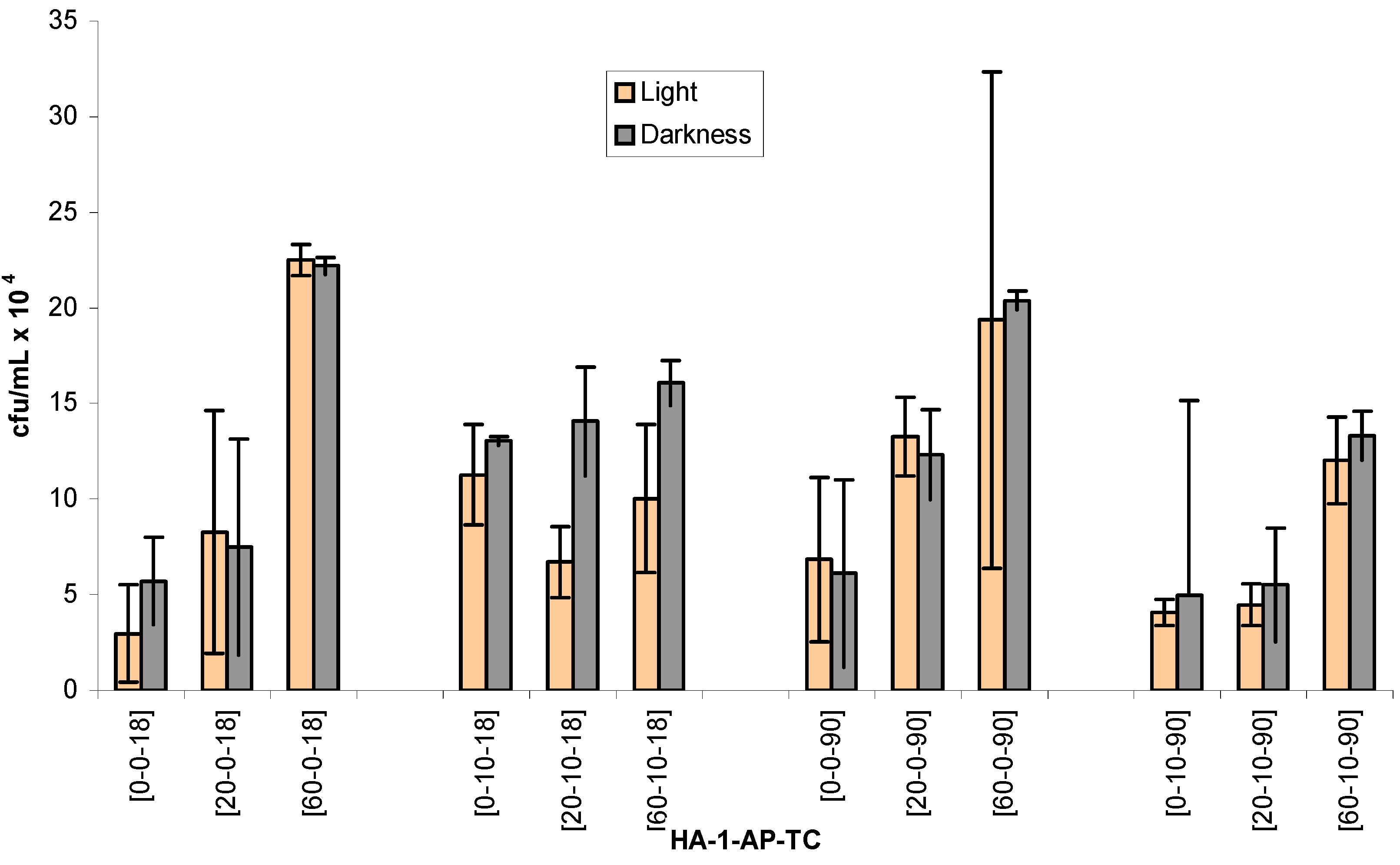
Figure 1.
Number of viable heterotrophic bacteria [colony forming units per mililiter (cfu/mL) (±STD)] after exposure to1-AP in sunlight or darkness, in the absence and in the presence of HA, for 18 and 90 minutes. Units for x-axis labels: HA (ppm); 1-AP (μM); TC (minutes).
Figure 1.
Number of viable heterotrophic bacteria [colony forming units per mililiter (cfu/mL) (±STD)] after exposure to1-AP in sunlight or darkness, in the absence and in the presence of HA, for 18 and 90 minutes. Units for x-axis labels: HA (ppm); 1-AP (μM); TC (minutes).
Statistical analysis
To detect the interactions between and among different parameters in the experiment, statistical analysis of results was conducted by factorial arrangement of treatments in a complete randomized design using General Linear Model by SAS. LSMEANS were used to separate means or combination of means. Difference in experimental data between two treatment groups was determined with student-t-test (p< 0.05).
Results and Discussion
Viable Count of Heterotrophic Bacteria
The data of the colony forming units (cfu/mL) of viable heterotrophic bacteria under different treatment structures are reported in
Table 1 and
Figure 1. According to chemical analysis with HPLC in preliminary experiments, 50% and all of 1-AP parent compound was photochemically transformed after 18 and 90 minutes, respectively. Therefore, time frames of 19 minutes and 90 minutes were chosen to compare the time-dependent toxicity effect.
Table 2.
Percent of 14C-D-glucose mineralized ( ± STD) by heterotrophic bacteria after exposure to 1-AP in sunlight and darkness for 18 and 90 minutes, in the presence or in the absence of HA, for 18 and 90 minutes.
Table 2.
Percent of 14C-D-glucose mineralized ( ± STD) by heterotrophic bacteria after exposure to 1-AP in sunlight and darkness for 18 and 90 minutes, in the presence or in the absence of HA, for 18 and 90 minutes.
| Conditions: | % DPM (± STD) |
| HA/1-AP/TC* | Light | Darkness |
| 0-0-18 | 16.3 | ± 0.2 | 29.5 | ± 2.3 |
| 20-0-18 | 20.6 | ± 0.5 | 21.4 | ± 5.6 |
| 60-0-18 | 24.4 | ± 1.9 | 21.8 | ± 0.5 |
| 0-10-18 | 13.73 | ± 0.04 | 6.3 | ± 0.2 |
| 20-10-18 | 15.7 | ± 2.9 | 28.7 | ± 2.8 |
| 60-10-18 | 6.6 | ± 1.7 | 19.02 | ± 1.17 |
| 0-0-90 | 26.6 | ± 1.1 | 16.6 | ± 4.9 |
| 20-0-90 | 28.4 | ± 1.6 | 27.1 | ± 2.4 |
| 60-0-90 | 27.8 | ± 2.5 | 28.08 | ± 0.47 |
| 0-10-90 | 10.3 | ± 2.3 | 17.4 | ±10.2 |
| 20-10-90 | 15.06 | ± 1.83 | 20.8 | ± 3.0 |
| 60-10-90 | 6.3 | ± 0.89 | 13.07 | ± 1.28 |
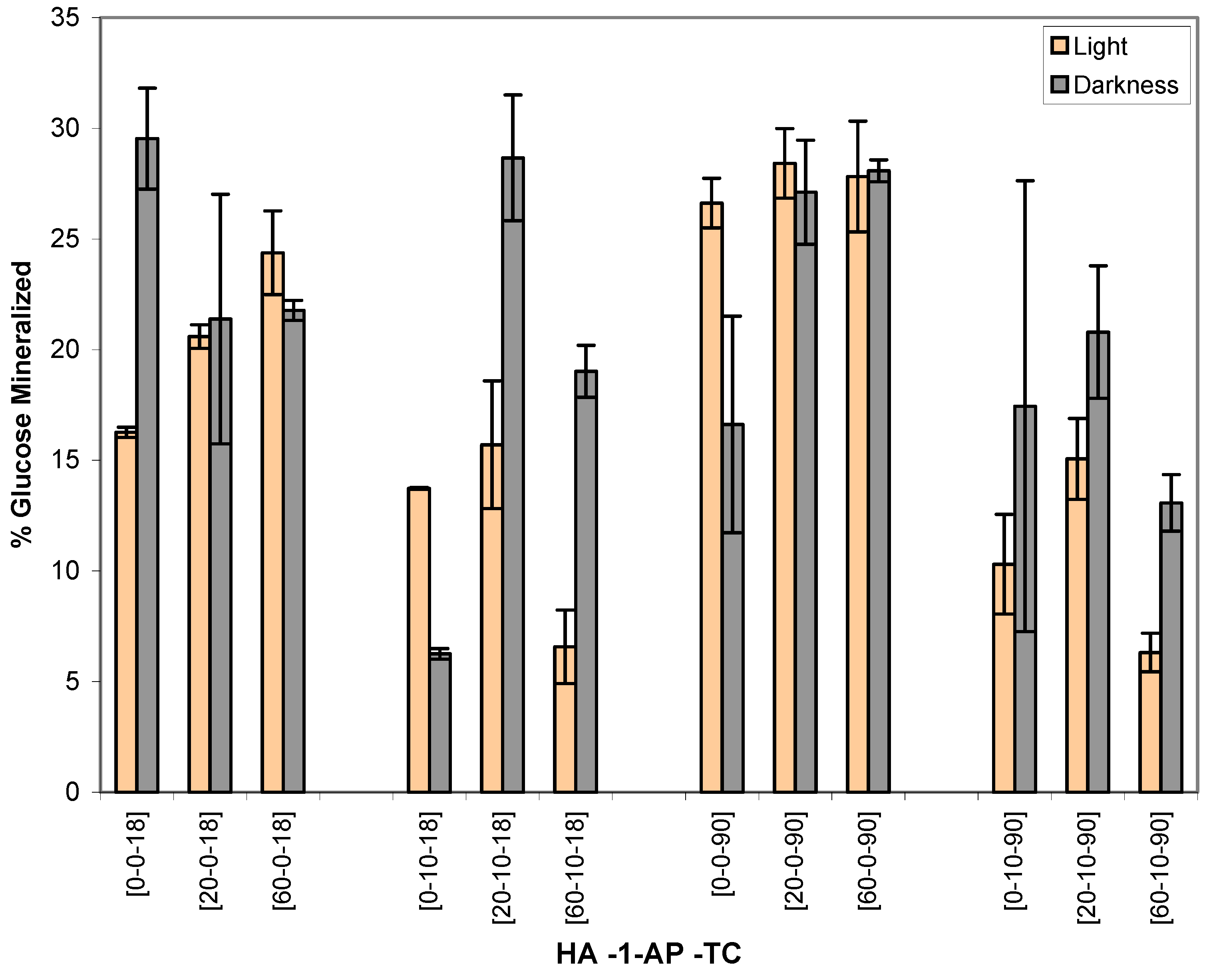
Figure 2.
Amount of 14C-D-glucose mineralized by heterotrophic bacteria (% ± STD) after exposure to1-AP in sunlight or darkness, in the absence and in the presence of HA, for 18 and 90 minutes. Units for x-axis labels: HA (ppm); 1-AP (μM); TC (minutes).
Figure 2.
Amount of 14C-D-glucose mineralized by heterotrophic bacteria (% ± STD) after exposure to1-AP in sunlight or darkness, in the absence and in the presence of HA, for 18 and 90 minutes. Units for x-axis labels: HA (ppm); 1-AP (μM); TC (minutes).
SAS analysis of viability counts indicates that HA*1-AP is the only significant interaction (p < 0.05) between the various treatments. Large deviation between the replicates of some groups limits the numbers of significant interactions among different treatment structures. However, there are some trends in the data sets reflecting the effect of individual parameter in this experiment. Treatment with HA only (in the absence of 1-AP) under both light and dark incubations generally cause enhancement of microbial viable count. More enhancements were observed under light than under dark incubations. Also, the degree of enhancement increases with the HA concentration. In addition, numerically for light and 18 minutes incubation with 20 and 60 ppm of HA, the enhancement was 2.8 and 7.6 times relative to the blank control (i.e. 0 ppm of HA) respectively. For dark incubations with 20 and 60 ppm, numbers were increased by 1.3 and 3.9 times respectively.
For incubation under light for 90 minutes, exposure to 20 and 60 ppm of HA enhanced viability counts by 1.1, and 3.0 times, respectively. For the similar samples exposing to darkness treatment, viability count was increased by 1.1 and 2.7 times respectively. Therefore, extent of enhancement was higher for the 18 minutes group than for the 90 minutes group. Perhaps there were more large nutrient substrates for microbes at the time point of 18 minutes. After 90 minutes the substrates were either photochemically degraded or physically adsorbed, and consequently became less effective in promoting the viable numbers. Nutrient agar is a selective microbial growth medium; therefore, it may fail to satisfy certain microorganisms that demanded other nutrient ingredients. Enhancing solubility of organic matters by HA has been reported to increase substrate bioavailability to microorganisms. Increase in bacterial viable count in the groups containing HA could occur as the result of this factor [
16].
By comparing to the relative 1-AP control groups, microbial number generally decreased when both HA and 1-AP were present. For example, cfu number decreases from (22.5 ± 0.8) x 10
4 to (10.0 ± 3.9) x 10
4 when comparison is made between the groups 60-10-18 and 60-0-18 in light. The higher inhibition was observed under two conditions: the longer incubation time (i.e., 90 minutes) and under light incubation (
Table 1,
Figure 1). It suggests that either 1-AP photoproducts caused the inhibition or the simultaneous presence of HA and photoproducts produced the toxic effects [20-10-18 vs. 20-0-18; 60-10-18 vs. 60-0-18]. In one of our relevant studies with atrazine, plate count data indicated that reactive chemical species produced by sensitized photolysis could cause the cytotoxicity to the test bacterial assemblages in natural water [
21]. In addition, microbial inhibition was also observed at 0 ppm of HA of 90 minutes groups [0-0-90 vs. 0-10-90]. Therefore, 1-AP photoproducts alone could also cause microbial toxicity [
22].
Heterotrophic Microbial Mineralization of D-glucose
SAS analysis indicates that the effect on microbial heterotrophic mineralization activity of D-glucose is significant by the following combination of treatments:
| 1-AP*PP | 1-AP*TC | PP*TC |
| HA*1-AP*PP | HA*1-AP*TC | HA*1-AP*TC*PP |
Table 2 shows the % of heterotrophic microbial mineralization of
14C-D glucose under the same treatment conditions as those in
Table 1. Although two-way, three-way, and four-way interactions were seen in this analysis set, cautious interpretation of the data is needed as microbial heterotrophic activity can be affected by a variety of factors such as toxicity, substrate competition, interaction between the substrates, and bioavailability of the substrate [
20]. Some of these results show similar pattern to viability count. In most cases, HA alone increased microbial activity under light and dark exposures. The greater increases were observed with the shorter exposure time and the higher HA concentration (
Table 2,
Figure 2). For example, in the groups of 20-0-18 (light), 60-0-18 (light), 20-0-90 (dark) and 60-0-90 (dark), HA caused enhancement of microbial activity as indicated by the higher % of mineralization of
14C-D-glucose.
Inhibition on microbial heterotrophic activity was observed in the groups exposing to HA in darkness for 18 minutes [20-0-18 vs. 0-0-18,
Table 2]. However, viability count was actually enhanced after exposure to HA (
Table 1). Therefore, we speculate that the inhibition was possibly caused by the decrease of glucose availability by HA binding instead of toxicity factor.
By comparing to the corresponding control group (e.g., 60-0-18 vs. 60-10-18) with student-t-test, microbial heterotrophic activity was significantly inhibited in the groups receiving 10 μM of 1-AP in the presence of HA (20-60 ppm) in light and in darkness. The extent of inhibition was more significant for the light exposure groups. Okinaka
et al. [
22] reported that 1-AP induces its direct-acting mutagenicity in the Ames/
Salmonella strain TA98 plating assay after exposing to an artificial source of near UV light. In our laboratory with Mutatox℘ Test the mutagenicity response of 1-AP increased after exposure to solar irradiation. 1-Nitropyrene, a known potent mammalian mutagenic compound [
23] was detected in the mixture of 1-AP photoproducts [unpublished data]. Our finding was consistent with the previous report by Okinaka
et al. [
22]. Exposure to HA in light and darkness, however, does not necessarily inhibit bacterial viability at the HA concentration range administered. Therefore, inhibition on microbial activity could have been caused by the combination of several factors including modification of glucose availability by HA, toxicity of 1-AP photoproducts and increased bioavailability of 1-AP and its photoproducts by HA.
In summary, treatment with HA only under both light and dark incubations generally cause enhancement of microbial viable count. More enhancements were observed under light than under dark incubations. In most cases, HA alone increased microbial heterotrophic activity under light and dark exposures. The greater increases were observed with the shorter exposure time and the higher HA concentrations. Inhibition on microbial heterotrophic activity was observed in the groups exposing to HA in darkness for 18 minutes. However, the inhibition was possibly caused by the decrease of glucose availability instead of toxicity factor. Microbial heterotrophic activity was significantly inhibited in the groups receiving 1-AP in the presence of HA in light and in darkness. The extent of inhibition was more significant for the light exposure groups. The inhibition could have been caused by the combination of several factors including modification of glucose availability by HA, toxicity of 1-AP photoproducts and increased bioavailability of 1-AP and its photoproducts by HA.
Although photoinduced toxicity of PAH could be caused by the dynamic interactions between light irradiation, PAH and target cellular materials, this study focused on the effect of 1-AP photoproducts after two time frames. Chemical characterization of HA could vary across sites; therefore, the results observed in this study might not be the same if different HA sources were used. Identification of 1-AP photoproducts is underway in our laboratories. It is clear more work is needed to determine the complicated interactions among multiple parameters involved in the study and further define the realistic hazards of PAH released into natural aquatic environment.


{kind=link}
{kind=link}