

Identification of Novel Staphylococcus aureus Core and Accessory Virulence Patterns in Chronic Rhinosinusitis

 ,
,
Abstract
1. Introduction
2. Results
2.1. Patient Demographics
2.2. Bacterial Genome Sequencing
2.3. Staphylococcal Protein A and agr Loci and Frameshift Assessment
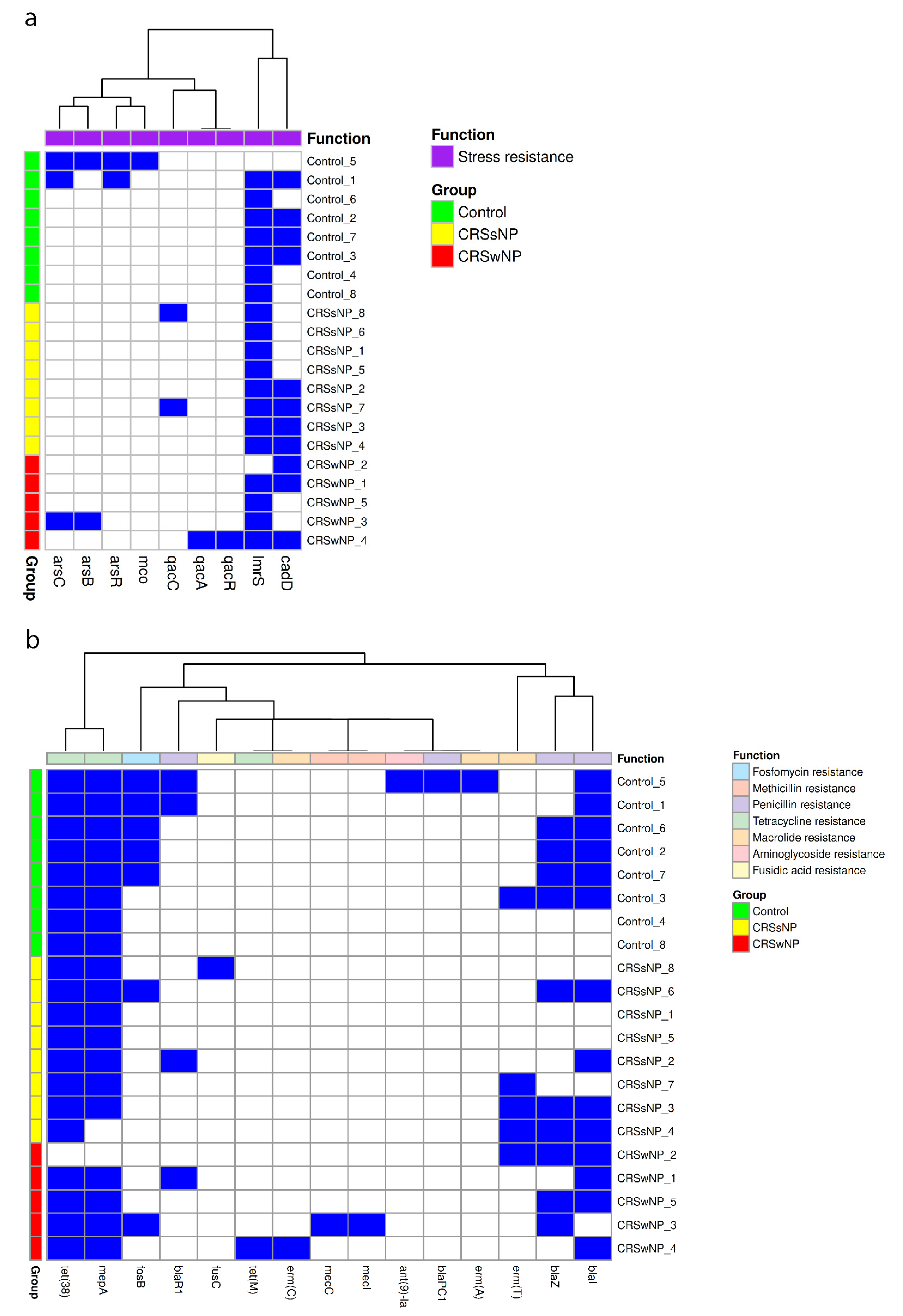
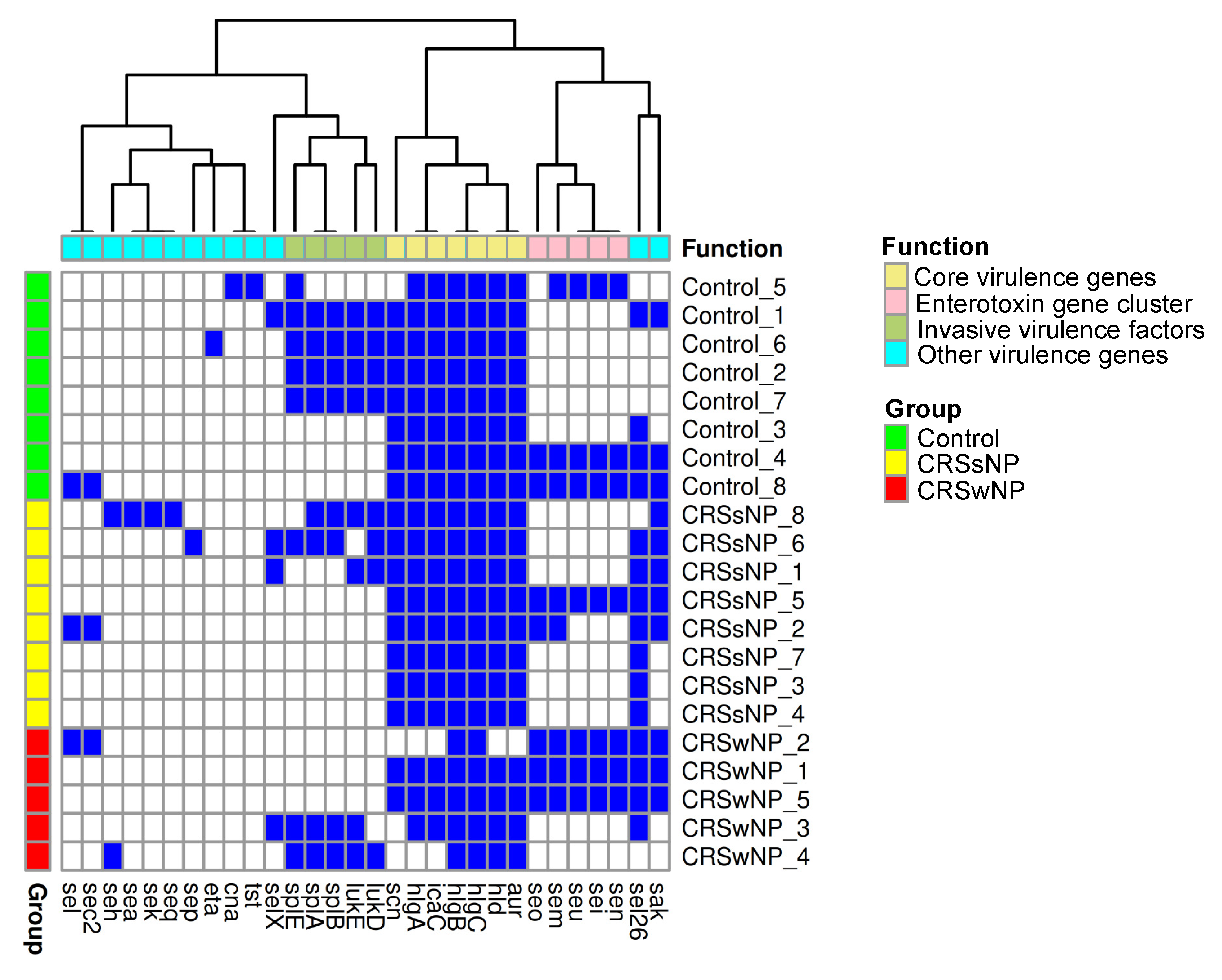
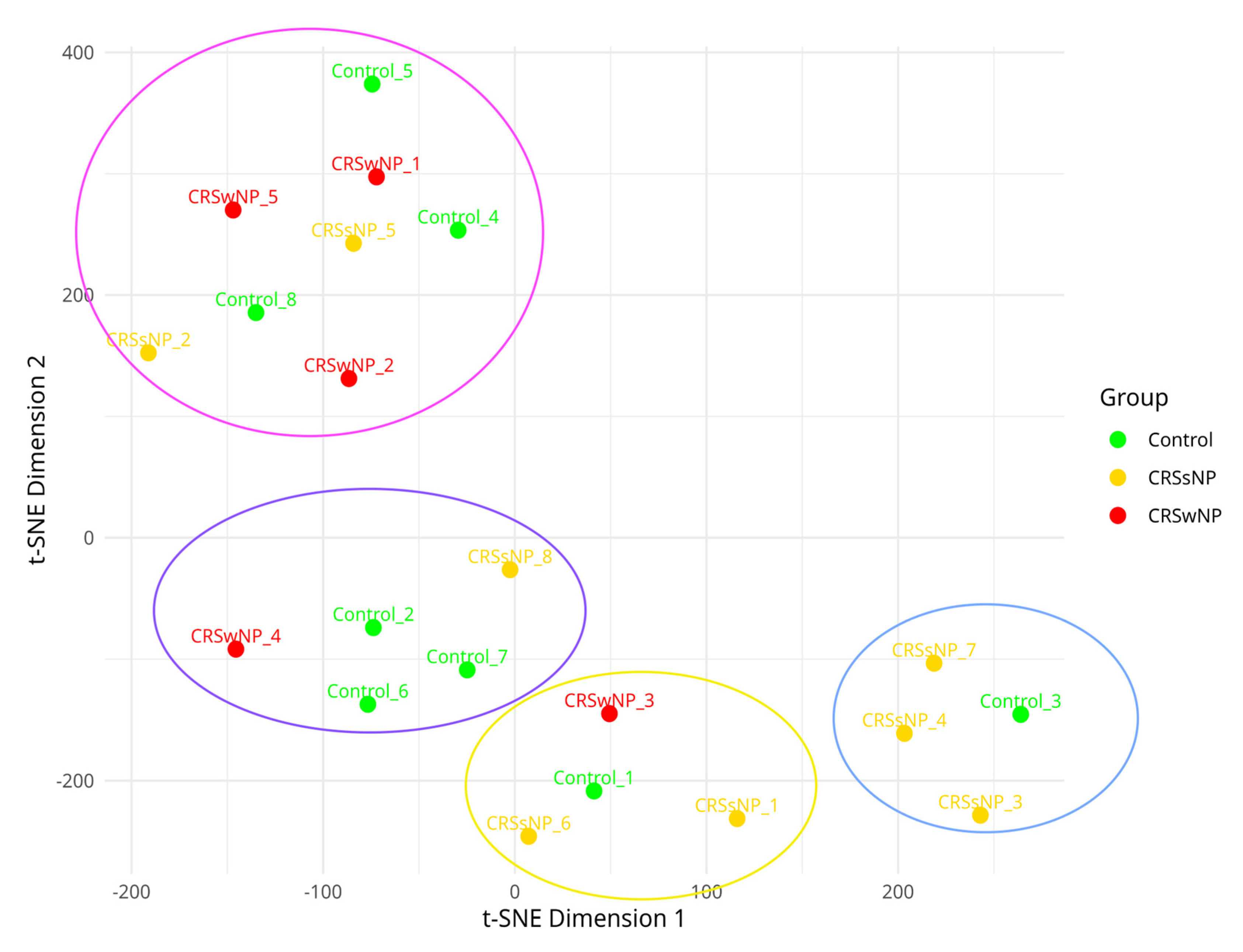
2.4. Stress, Antimicrobial Resistance, and Virulence Genes
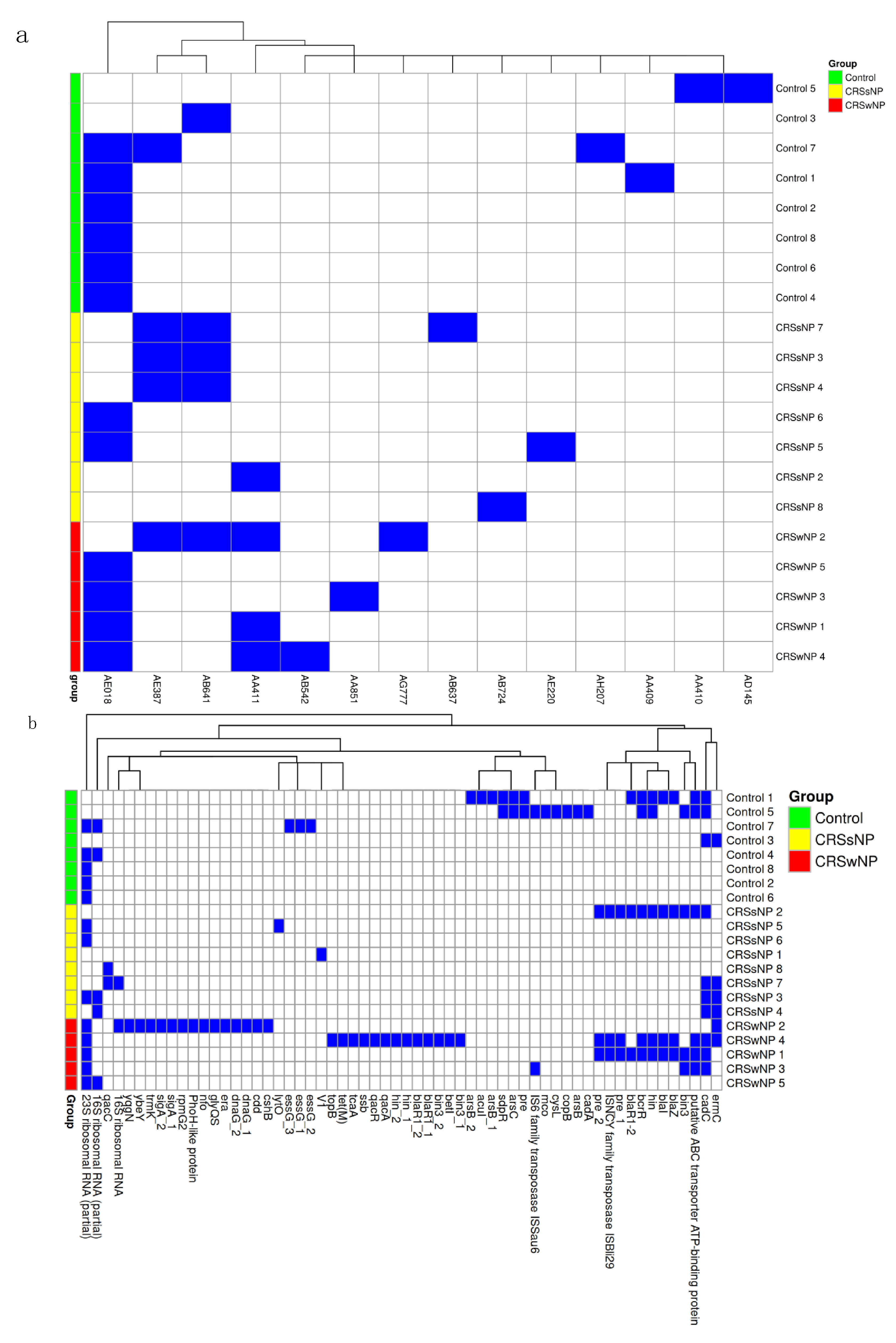
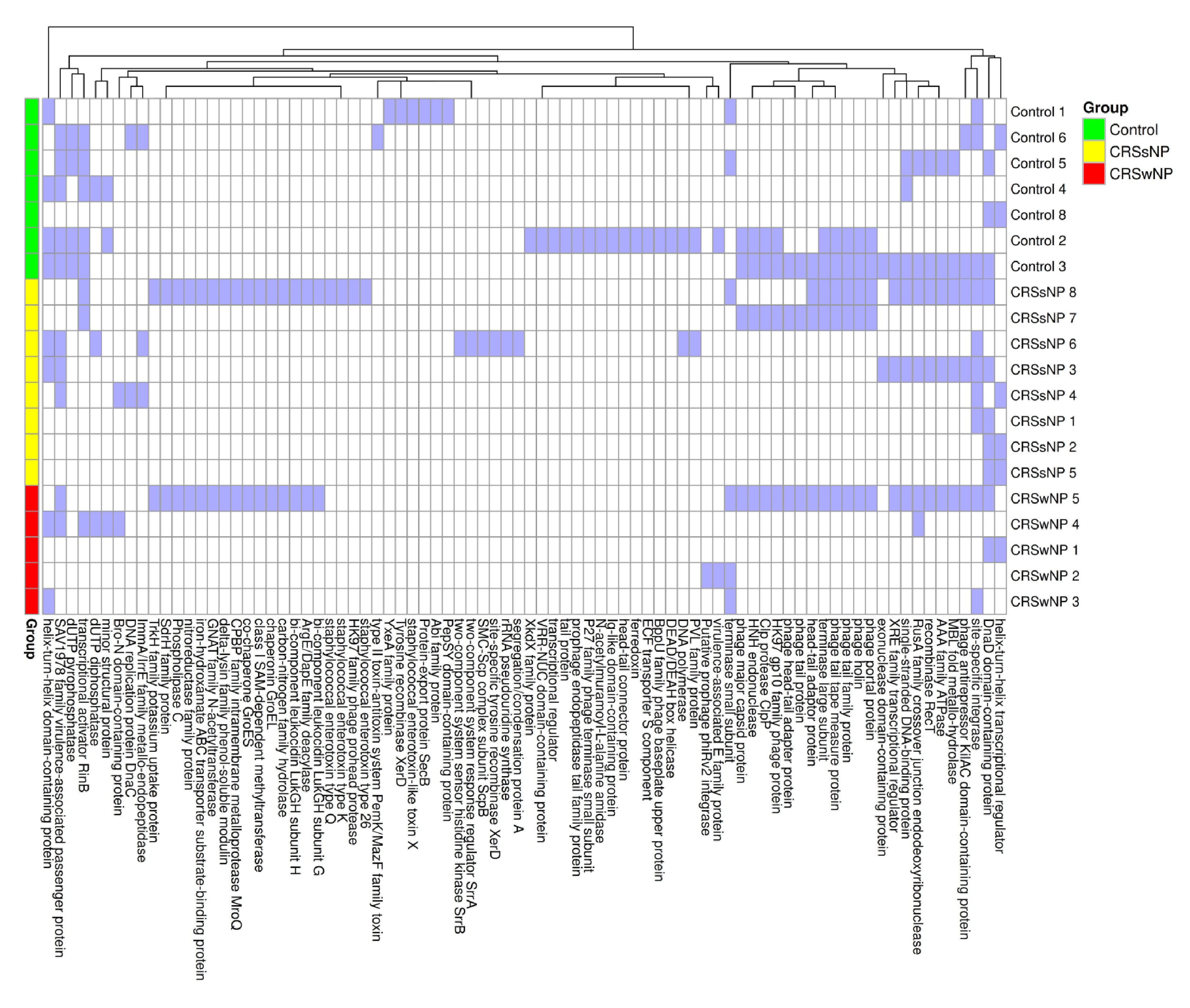
2.5. Mobile Genetic Element Identification
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Specimen Testing and Genomic Sequencing
4.3. Alignment and Assembly of the Genome
4.4. Typing of the spa and agr Loci and Detection of Frameshifts
4.5. Stress, Virulence, and Antimicrobial Resistance Genes
4.6. Plasmid Detection
4.7. Prophage Detection
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| agr | Accessory gene regulator |
| ANOVA | Analysis of Variance |
| aur | Aureolysin gene |
| blaZ | Beta-lactamase gene |
| cadD | Cadmium resistance gene |
| CRS | Chronic rhinosinusitis |
| CRSsNP | Chronic rhinosinusitis without nasal polyps |
| CRSwNP | Chronic rhinosinusitis with nasal polyps |
| DNA | Deoxyribonucleic acid |
| erm(A), erm (T), erm (C) | Erythromycin resistance methylase genes |
| fosB | Fosfomycin resistance gene |
| hlgA, hlgB, hlgC | Gamma hemolysin genes |
| HSP | High scoring pairs |
| hld | Delta-hemolysin gene |
| icaC | Biofilm-associated intercellular adhesion gene |
| lmrS | Lincomycin resistance protein of S. aureus |
| lukE, lukD | Leukocidin genes |
| MALDI-TOF | Matrix-assisted laser desorption/ionization time of flight |
| MGE | Mobile genetic element |
| MLMS | Modified Lund–Mackay Score |
| PERMANOVA | Permutational multivariate analysis of variance |
| RNA | Ribonucleic acid |
| RNAII | Second transcript of the agr operon |
| RNAIII | Third transcript of the agr operon |
| S. aureus | Staphylococcus aureus |
| SCV | Small colony variant |
| sei, sem, sen, seo, seu | Enterotoxin gene cluster |
| spa | Staphylococcal protein A gene |
| splA, splB, splE | Serine protease genes |
| tet(38), tet(M) | Tetracycline resistance genes |
| t-SNE | t-distributed stochastic neighbor embedding |
References
- Cosgrove, S.E.; Sakoulas, G.; Perencevich, E.N.; Schwaber, M.J.; Karchmer, A.W.; Carmeli, Y. Comparison of mortality associated with methicillin-resistant and methicillin-susceptible Staphylococcus aureus bacteremia: A meta-analysis. Clin. Infect. Dis. 2003, 36, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Vickery, T.W.; Ramakrishnan, V.R.; Suh, J.D. The Role of Staphylococcus aureus in Patients with Chronic Sinusitis and Nasal Polyposis. Curr. Allergy Asthma Rep. 2019, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- Van Zele, T.; Gevaert, P.; Watelet, J.B.; Claeys, G.; Holtappels, G.; Claeys, C.; van Cauwenberge, P.; Bachert, C. Staphylococcus aureus colonization and IgE antibody formation to enterotoxins is increased in nasal polyposis. J. Allergy Clin. Immunol. 2004, 114, 981–983. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.Y.; Miao, J.L.; Lu, H.Q.; Qi, Q.H.; Chen, X.I.; Xu, J.; Lin, Z.P.; Chen, Z.B.; Yin, M.; Cheng, L. Serum levels of specific IgE to Staphylococcus aureus enterotoxins in patients with chronic rhinosinusitis. Exp. Ther. Med. 2015, 9, 1523–1527. [Google Scholar] [CrossRef]
- Maniakas, A.; Asmar, M.H.; Renteria Flores, A.E.; Nayan, S.; Alromaih, S.; Mfuna Endam, L.; Desrosiers, M.Y. Staphylococcus aureus on Sinus Culture Is Associated With Recurrence of Chronic Rhinosinusitis After Endoscopic Sinus Surgery. Front. Cell Infect. Microbiol. 2018, 8, 150. [Google Scholar] [CrossRef]
- Watkins, K.E.; Unnikrishnan, M. Evasion of host defenses by intracellular Staphylococcus aureus. Adv. Appl. Microbiol. 2020, 112, 105–141. [Google Scholar]
- Tan, N.C.; Foreman, A.; Jardeleza, C.; Douglas, R.; Vreugde, S.; Wormald, P.J. Intracellular Staphylococcus aureus: The Trojan horse of recalcitrant chronic rhinosinusitis? Int. Forum Allergy Rhinol. 2013, 3, 261–266. [Google Scholar] [CrossRef]
- Singhal, D.; Foreman, A.; Jervis-Bardy, J.; Wormald, P.J. Staphylococcus aureus biofilms: Nemesis of endoscopic sinus surgery. Laryngoscope 2011, 121, 1578–1583. [Google Scholar] [CrossRef]
- Hayes, S.M.; Howlin, R.; Johnston, D.A.; Webb, J.S.; Clarke, S.C.; Stoodley, P.; Harries, P.G.; Wilson, S.J.; Pender, S.L.; Faust, S.N.; et al. Intracellular residency of Staphylococcus aureus within mast cells in nasal polyps: A novel observation. J. Allergy Clin. Immunol. 2015, 135, 1648–1651. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef]
- Shaghayegh, G.; Cooksley, C.; Ramezanpour, M.; Wormald, P.J.; Psaltis, A.J.; Vreugde, S. Chronic Rhinosinusitis, S. aureus Biofilm and Secreted Products, Inflammatory Responses, and Disease Severity. Biomedicines 2022, 10, 1362. [Google Scholar] [CrossRef] [PubMed]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Loffler, B.; Tuchscherr, L.; Niemann, S.; Peters, G. Staphylococcus aureus persistence in non-professional phagocytes. Int. J. Med. Microbiol. 2014, 304, 170–176. [Google Scholar] [CrossRef]
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef]
- Yoong, P.; Torres, V.J. The effects of Staphylococcus aureus leukotoxins on the host: Cell lysis and beyond. Curr. Opin. Microbiol. 2013, 16, 63–69. [Google Scholar] [CrossRef]
- Morinaga, N.; Kaihou, Y.; Noda, M. Purification, cloning and characterization of variant LukE-LukD with strong leukocidal activity of staphylococcal bi-component leukotoxin family. Microbiol. Immunol. 2003, 47, 81–90. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Otto, M. Phenol-soluble modulins. Int. J. Med. Microbiol. 2014, 304, 164–169. [Google Scholar] [CrossRef]
- Van Zele, T.; Gevaert, P.; Holtappels, G.; van Cauwenberge, P.; Bachert, C. Local immunoglobulin production in nasal polyposis is modulated by superantigens. Clin. Exp. Allergy 2007, 37, 1840–1847. [Google Scholar] [CrossRef]
- Hayes, S.M.; Biggs, T.C.; Goldie, S.P.; Harries, P.G.; Walls, A.F.; Allan, R.N.; Pender, S.L.F.; Salib, R.J. Staphylococcus aureus internalization in mast cells in nasal polyps: Characterization of interactions and potential mechanisms. J. Allergy Clin. Immunol. 2020, 145, 147–159. [Google Scholar] [CrossRef]
- Tan, L.; Huang, Y.; Shang, W.; Yang, Y.; Peng, H.; Hu, Z.; Wang, Y.; Rao, Y.; Hu, Q.; Rao, X.; et al. Accessory Gene Regulator (agr) Allelic Variants in Cognate Staphylococcus aureus Strain Display Similar Phenotypes. Front. Microbiol. 2022, 13, 700894. [Google Scholar] [CrossRef] [PubMed]
- Sangvik, M.; Olsen, R.S.; Olsen, K.; Simonsen, G.S.; Furberg, A.S.; Sollid, J.U. Age- and gender-associated Staphylococcus aureus spa types found among nasal carriers in a general population: The Tromso Staph and Skin Study. J. Clin. Microbiol. 2011, 49, 4213–4218. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, I.; Millon, B.; Meugnier, H.; Vandenesch, F.; Maurin, M.; Pavese, P.; Boisset, S. High prevalence of spa type t571 among methicillin-susceptible Staphylococcus aureus from bacteremic patients in a French University Hospital. PLoS ONE 2018, 13, e0204977. [Google Scholar] [CrossRef]
- Dunman, P.M.; Murphy, E.; Haney, S.; Palacios, D.; Tucker-Kellogg, G.; Wu, S.; Brown, E.L.; Zagursky, R.J.; Shlaes, D.; Projan, S.J. Transcription profiling-based identification of Staphylococcus aureus genes regulated by the agr and/or sarA loci. J. Bacteriol. 2001, 183, 7341–7353. [Google Scholar] [CrossRef]
- Nepal, R.; Houtak, G.; Shaghayegh, G.; Bouras, G.; Shearwin, K.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Prophages encoding human immune evasion cluster genes are enriched in Staphylococcus aureus isolated from chronic rhinosinusitis patients with nasal polyps. Microb. Genom. 2021, 7, 000726. [Google Scholar] [CrossRef]
- Houtak, G.; Bouras, G.; Nepal, R.; Shaghayegh, G.; Cooksley, C.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. The intra-host evolutionary landscape and pathoadaptation of persistent Staphylococcus aureus in chronic rhinosinusitis. Microb. Genom. 2023, 9, 001128. [Google Scholar] [CrossRef]
- Petit, R.A., III; Read, T.D. Bactopia: A Flexible Pipeline for Complete Analysis of Bacterial Genomes. mSystems 2020, 5, 10–1128. [Google Scholar] [CrossRef]
- Philpott, C.M.; Erskine, S.; Hopkins, C.; Kumar, N.; Anari, S.; Kara, N.; Sunkaraneni, S.; Ray, J.; Clark, A.; Wilson, A.; et al. Prevalence of asthma, aspirin sensitivity and allergy in chronic rhinosinusitis: Data from the UK National Chronic Rhinosinusitis Epidemiology Study. Respir. Res. 2018, 19, 129. [Google Scholar] [CrossRef]
- Jarvis, D.; Newson, R.; Lotvall, J.; Hastan, D.; Tomassen, P.; Keil, T.; Gjomarkaj, M.; Forsberg, B.; Gunnbjornsdottir, M.; Minov, J.; et al. Asthma in adults and its association with chronic rhinosinusitis: The GA2LEN survey in Europe. Allergy 2012, 67, 91–98. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58 (Suppl. S29), 1–464. [Google Scholar] [CrossRef]
- Hopkins, C.; Browne, J.P.; Slack, R.; Lund, V.; Brown, P. The Lund-Mackay staging system for chronic rhinosinusitis: How is it used and what does it predict? Otolaryngol. Head. Neck Surg. 2007, 137, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.M.; Hoehle, L.; Bergmark, R.W.; Caradonna, D.S.; Gray, S.T.; Sedaghat, A.R. Reversal of Smoking Effects on Chronic Rhinosinusitis after Smoking Cessation. Otolaryngol. Head. Neck Surg. 2017, 157, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Wagner Mackenzie, B.; Baker, J.; Douglas, R.G.; Taylor, M.W.; Biswas, K. Detection and quantification of Staphylococcus in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2019, 9, 1462–1469. [Google Scholar] [CrossRef] [PubMed]
- Foreman, A.; Wormald, P.J. Different biofilms, different disease? A clinical outcomes study. Laryngoscope 2010, 120, 1701–1706. [Google Scholar] [CrossRef]
- Shopsin, B.; Mathema, B.; Alcabes, P.; Said-Salim, B.; Lina, G.; Matsuka, A.; Martinez, J.; Kreiswirth, B.N. Prevalence of agr specificity groups among Staphylococcus aureus strains colonizing children and their guardians. J. Clin. Microbiol. 2003, 41, 456–459. [Google Scholar] [CrossRef]
- Jarraud, S.; Lyon, G.J.; Figueiredo, A.M.; Lina, G.; Vandenesch, F.; Etienne, J.; Muir, T.W.; Novick, R.P. Exfoliatin-producing strains define a fourth agr specificity group in Staphylococcus aureus. J. Bacteriol. 2000, 182, 6517–6522. [Google Scholar] [CrossRef]
- Janzon, L.; Arvidson, S. The role of the delta-lysin gene (hld) in the regulation of virulence genes by the accessory gene regulator (agr) in Staphylococcus aureus. EMBO J. 1990, 9, 1391–1399. [Google Scholar] [CrossRef]
- Peterson, M.L.; Schlievert, P.M. Glycerol monolaurate inhibits the effects of Gram-positive select agents on eukaryotic cells. Biochemistry 2006, 45, 2387–2397. [Google Scholar] [CrossRef]
- Haslinger-Loffler, B.; Kahl, B.C.; Grundmeier, M.; Strangfeld, K.; Wagner, B.; Fischer, U.; Cheung, A.L.; Peters, G.; Schulze-Osthoff, K.; Sinha, B. Multiple virulence factors are required for Staphylococcus aureus-induced apoptosis in endothelial cells. Cell Microbiol. 2005, 7, 1087–1097. [Google Scholar] [CrossRef]
- Sendi, P.; Proctor, R.A. Staphylococcus aureus as an intracellular pathogen: The role of small colony variants. Trends Microbiol. 2009, 17, 54–58. [Google Scholar] [CrossRef]
- Vuong, C.; Saenz, H.L.; Gotz, F.; Otto, M. Impact of the agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis. 2000, 182, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Bardy, J.J.; Sarovich, D.S.; Price, E.P.; Steinig, E.; Tong, S.; Drilling, A.; Ou, J.; Vreugde, S.; Wormald, P.J.; Psaltis, A.J. Staphylococcus aureus from patients with chronic rhinosinusitis show minimal genetic association between polyp and non-polyp phenotypes. BMC Ear Nose Throat Disord. 2018, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Staali, L.; Colin, D.A. Bi-component HlgC/HlgB and HlgA/HlgB gamma-hemolysins from S. aureus: Modulation of Ca(2+) channels activity through a differential mechanism. Toxicon 2021, 201, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Laarman, A.J.; Ruyken, M.; Malone, C.L.; van Strijp, J.A.; Horswill, A.R.; Rooijakkers, S.H. Staphylococcus aureus metalloprotease aureolysin cleaves complement C3 to mediate immune evasion. J. Immunol. 2011, 186, 6445–6453. [Google Scholar] [CrossRef]
- Atkin, K.E.; MacDonald, S.J.; Brentnall, A.S.; Potts, J.R.; Thomas, G.H. A different path: Revealing the function of staphylococcal proteins in biofilm formation. FEBS Lett. 2014, 588, 1869–1872. [Google Scholar] [CrossRef]
- Sedarat, Z.; Taylor-Robinson, A.W. Biofilm Formation by Pathogenic Bacteria: Applying a Staphylococcus aureus Model to Appraise Potential Targets for Therapeutic Intervention. Pathogens 2022, 11, 388. [Google Scholar] [CrossRef]
- Alonzo, F., III; Benson, M.A.; Chen, J.; Novick, R.P.; Shopsin, B.; Torres, V.J. Staphylococcus aureus leucocidin ED contributes to systemic infection by targeting neutrophils and promoting bacterial growth in vivo. Mol. Microbiol. 2012, 83, 423–435. [Google Scholar] [CrossRef]
- Dasari, P.; Nordengrun, M.; Vilhena, C.; Steil, L.; Abdurrahman, G.; Surmann, K.; Dhople, V.; Lahrberg, J.; Bachert, C.; Skerka, C.; et al. The Protease SplB of Staphylococcus aureus Targets Host Complement Components and Inhibits Complement-Mediated Bacterial Opsonophagocytosis. J. Bacteriol. 2022, 204, e0018421. [Google Scholar] [CrossRef]
- Fischer, A.J.; Kilgore, S.H.; Singh, S.B.; Allen, P.D.; Hansen, A.R.; Limoli, D.H.; Schlievert, P.M. High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates. Genes 2019, 10, 1036. [Google Scholar] [CrossRef]
- Song, L.; Florea, L.; Langmead, B. Lighter: Fast and memory-efficient sequencing error correction without counting. Genome Biol. 2014, 15, 509. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 2021, 7, 000685. [Google Scholar]
- Harmsen, D.; Claus, H.; Witte, W.; Rothganger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, 10–1128. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation For Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps, R package version 1.0.12; Github: Tallinn, Estonia, 2018. [Google Scholar]
- Krijthe, J.H. T-Distributed Stochastic Neighbor Embedding Using a Barnes-Hut Implementation, R package version 0.17; Github Leiden University: Leiden, The Netherlands, 2015. [Google Scholar]
- Wickham, H. Ggplot2; Springer Science + Business Media, LLC: New York, NY, USA, 2016. [Google Scholar]
- Oksanen, J.S.G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.S.P.; Stevens, M.; Szoecs, E.; Wagner, H.; Barbour, M.; et al. Vegan: Community Ecology Package, R package version 2.6-10; CRAN University of Oulu: Oulu, Finland, 2024. [Google Scholar]
- Robertson, J.; Nash, J.H.E. MOB-suite: Software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 2018, 4, e000206. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.; Bessonov, K.; Schonfeld, J.; Nash, J.H.E. Universal whole-sequence-based plasmid typing and its utility to prediction of host range and epidemiological surveillance. Microb. Genom. 2020, 6, e000435. [Google Scholar] [CrossRef]
- Akhter, S.; Aziz, R.K.; Edwards, R.A. PhiSpy: A novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies. Nucleic Acids Res. 2012, 40, e126. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Type | Control | CRSsNP | CRSwNP | p-Value |
|---|---|---|---|---|
| Number of subjects | 30 (38.9%) | 20 (25.9%) | 27 (35.1%) | - |
| Gender (M/F) | 15:15 | 9:11 | 16:11 | 0.603 † |
| Mean age (SD) | 53.5 (16.6) | 57.2 (16.7) | 51.4 (15.9) | 0.447 ‡ |
| Clinical review/procedure | Clinical review 6 Biopsy 4 Septoplasty/PETS 13 Limited FESS 7 Full house FESS 0 | Clinical review 7 Biopsy 0 Septoplasty/PETS 1 Limited FESS 6 Full house FESS 6 | Clinical review 5 Biopsy 0 Septoplasty/PETS 0 Limited FESS 2 Full house FESS 20 | <0.001 † |
| Antibiotic use in the past month | 2 (6.67%) | 1 (5.0%) | 5 (18.5%) | 0.225 † |
| Steroid use in the past month | 2 (6.67%) | 0 | 6 (22.2%) | 0.033 † |
| S. aureus culture | 8 (26.6%) | 8 (40%) | 5 (18.5%) | 0.262 † |
| Airborne allergies | 8 (26.6%) | 3 (15%) | 8 (29.6%) | 0.490 † |
| Asthma | 4 (13.3%) | 4 (20%) | 15 (55.5%) | 0.001 † |
| Ex-smokers | 5 (16.7%) | 5 (25%) | 7 (25.9%) | 0.628 † |
| Current Smokers | 4 (13.3%) | 0 | 0 | 0.039 † |
| Modified Lund–Mackay Score | 3.6 | 7.9 | 13.5 | <0.001 ‡ |
| Culture of S. aureus | No Growth | Growth | p-Value |
|---|---|---|---|
| Number of subjects | 56 (73%) | 21 (27%) | - |
| Gender (M/F) | 31:25 | 9:12 | 0.328 † |
| Age (SD) | 49.3 (15.6) | 51.0 (18.5) | 0.981 ‡ |
| Clinical review/procedure | Clinical review 11 Biopsy 2 Septoplasty/PETS 9 Limited FESS 14 Full House FESS 20 | Clinical review 7 Biopsy 2 Septoplasty/PETS 5 Limited FESS 0 Full House FESS 7 | 0.090 † |
| Antibiotic use in the past month | 7 (12.5%) | 1 (4.8%) | 0.322 † |
| Steroid use in the past month | 7 (12.5%) | 1 (4.8%) | 0.322 † |
| Airborne allergies | 14 (25%) | 5 (23.8%) | 0.914 † |
| Asthma | 14 (25%) | 9 (42.9%) | 0.127 † |
| Ex-smokers | 11 (19.6%) | 6 (28.5%) | 0.423 † |
| Current smokers | 3 (5.4%) | 1 (4.8%) | 0.904 † |
| Modified Lund Mackay Score | 7.5 | 12.0 | 0.038 ‡ |
| Isolate | agr Locus Type | spa Type | Frameshifts Mutations in the agr Operon |
|---|---|---|---|
| Control 1 | I | t008 | None |
| Control 2 | II | t084 | None |
| Control 3 | I | t1451 | None |
| Control 4 | I | t065 | None |
| Control 5 | III | t338 | None |
| Control 6 | II | t346 | None |
| Control 7 | II | t084 | None |
| Control 8 | I | t230 | None |
| CRSsNP 1 | I | t1236 | None |
| CRSsNP 2 | I | t340 | None |
| CRSsNP 3 | I | t571 | None |
| CRSsNP 4 | I | t571 | None |
| CRSsNP 5 | I | t065 | None |
| CRSsNP 6 | I | t008 | None |
| CRSsNP 7 | I | u | None |
| CRSsNP 8 | III | T127 | None |
| CRSwNP 1 | I | T015 | None |
| CRSwNP 2 | I | T015 | Absent hld gene |
| CRSwNP 3 | II | T6292 | Frameshift insertion at position 28 in the hld gene |
| CRSwNP 4 | III | T127 | None |
| CRSwNP 5 | I | T18906 | None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldie, S.P.; Lau, L.C.; Jones, H.A.S.; Harries, P.G.; Walls, A.F.; Salib, R.J. Identification of Novel Staphylococcus aureus Core and Accessory Virulence Patterns in Chronic Rhinosinusitis. Int. J. Mol. Sci. 2025, 26, 3711. https://doi.org/10.3390/ijms26083711
Goldie SP, Lau LC, Jones HAS, Harries PG, Walls AF, Salib RJ. Identification of Novel Staphylococcus aureus Core and Accessory Virulence Patterns in Chronic Rhinosinusitis. International Journal of Molecular Sciences. 2025; 26(8):3711. https://doi.org/10.3390/ijms26083711
Chicago/Turabian StyleGoldie, Simon P., Laurie C. Lau, Huw A. S. Jones, Philip G. Harries, Andrew F. Walls, and Rami J. Salib. 2025. "Identification of Novel Staphylococcus aureus Core and Accessory Virulence Patterns in Chronic Rhinosinusitis" International Journal of Molecular Sciences 26, no. 8: 3711. https://doi.org/10.3390/ijms26083711
APA StyleGoldie, S. P., Lau, L. C., Jones, H. A. S., Harries, P. G., Walls, A. F., & Salib, R. J. (2025). Identification of Novel Staphylococcus aureus Core and Accessory Virulence Patterns in Chronic Rhinosinusitis. International Journal of Molecular Sciences, 26(8), 3711. https://doi.org/10.3390/ijms26083711