
Total Alkaloid Extract of Nelumbinis Plumula Promoted Sleep in PCPA-Induced Insomnia Rats by Affecting Neurotransmitters and Their Receptor Activities
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
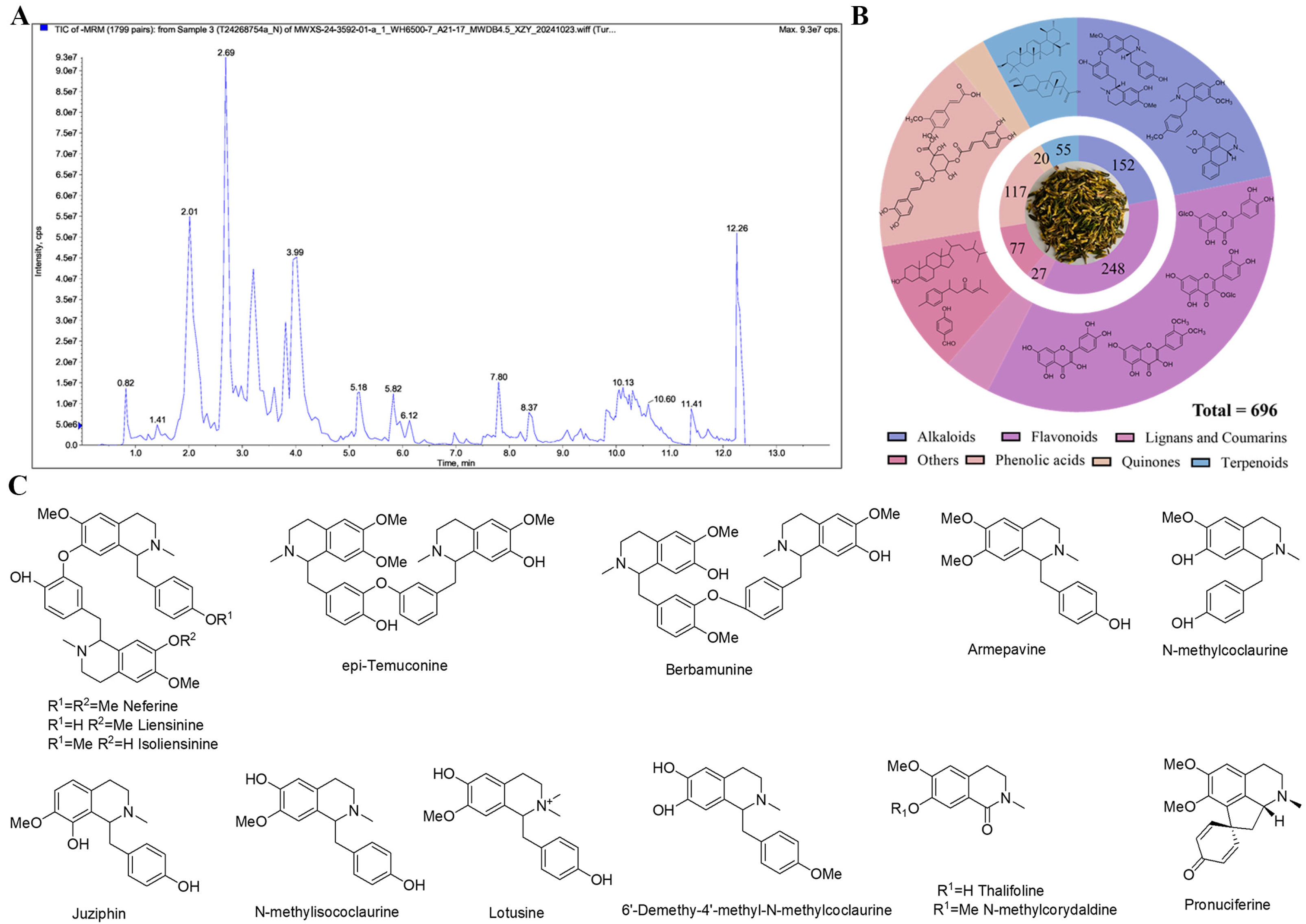
2.1. Analysis Results of Nelumbinis Plumula Extract by UPLC-MS/MS
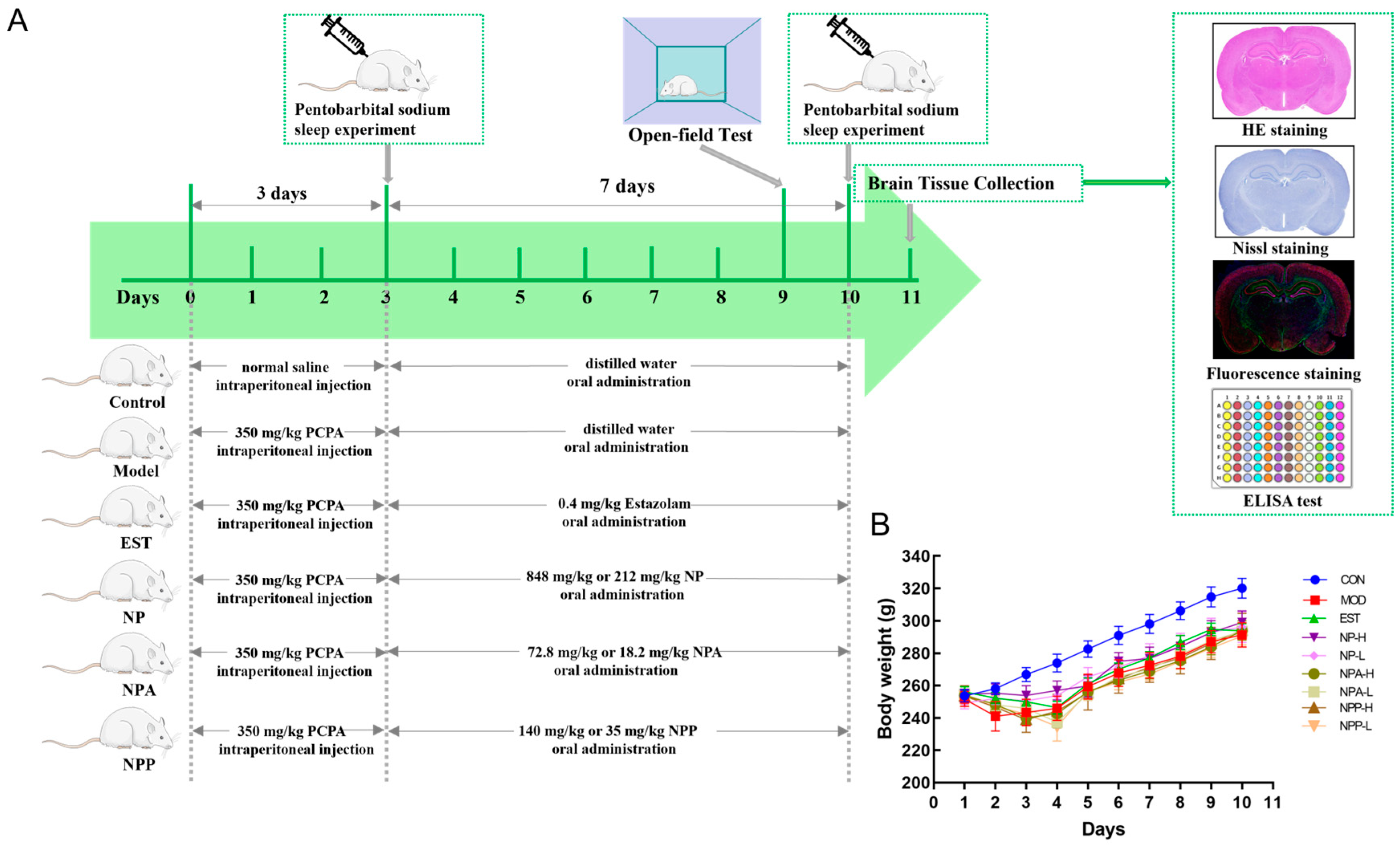
2.2. Animal Experiment and General Condition of Rats
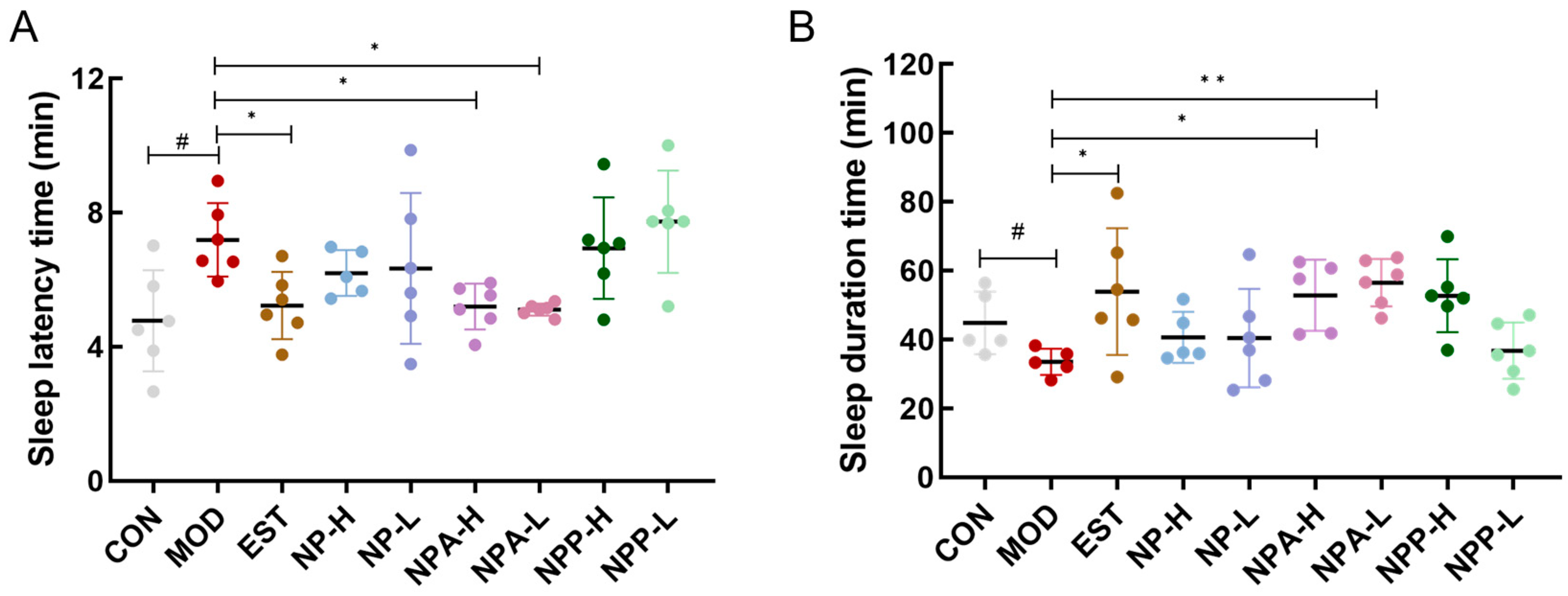
2.3. Effects of Various Extracts in PCPA-Induced Insomnia Rats by Pentobarbital Sodium Experiment
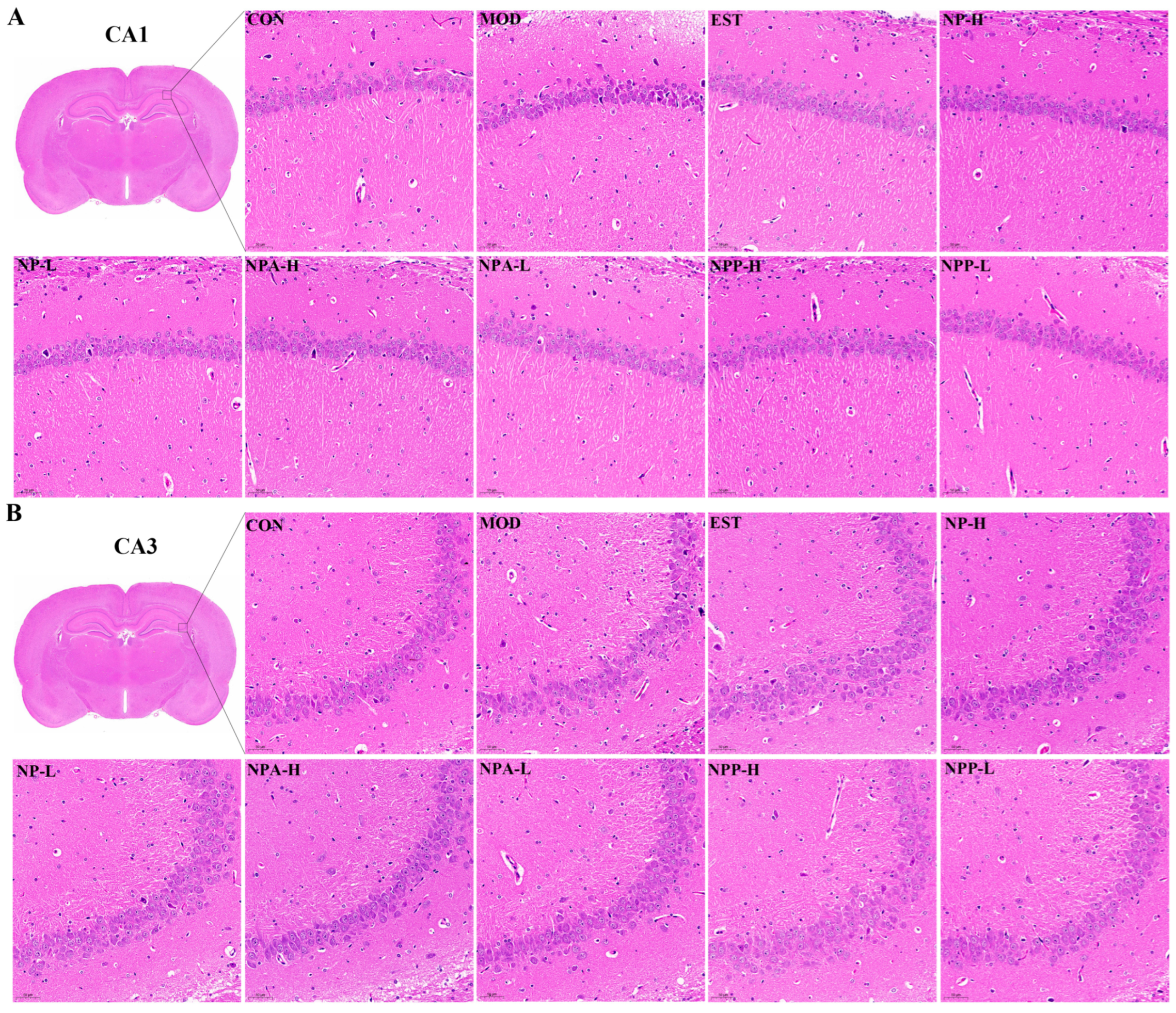
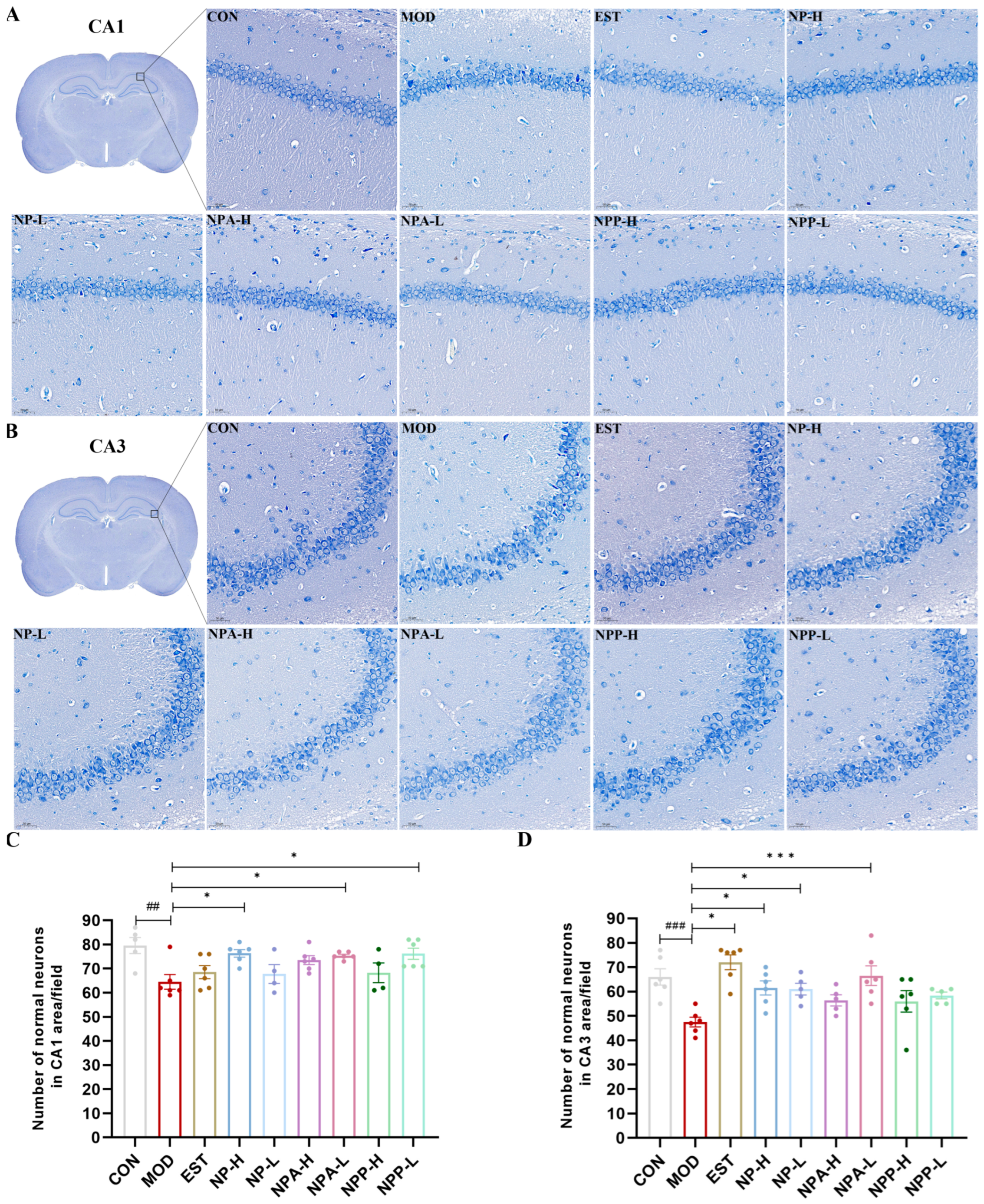
2.4. Pathological Observations of HE Staining and Nissl Staining of PCPA-Induced Insomnia Rats
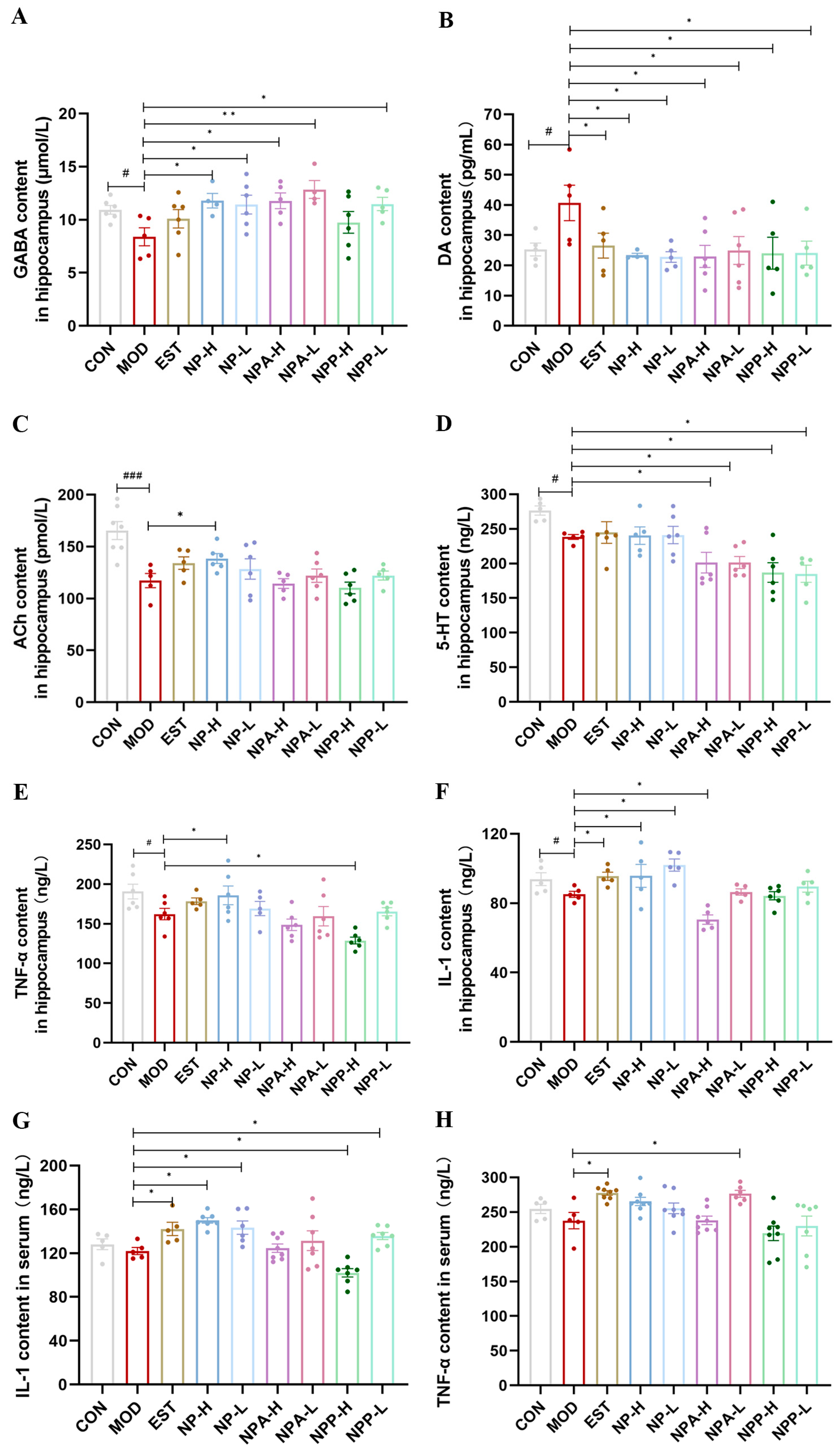
2.5. Determination of Neurotransmitters and Inflammatory Factors in Hippocampus of Rats
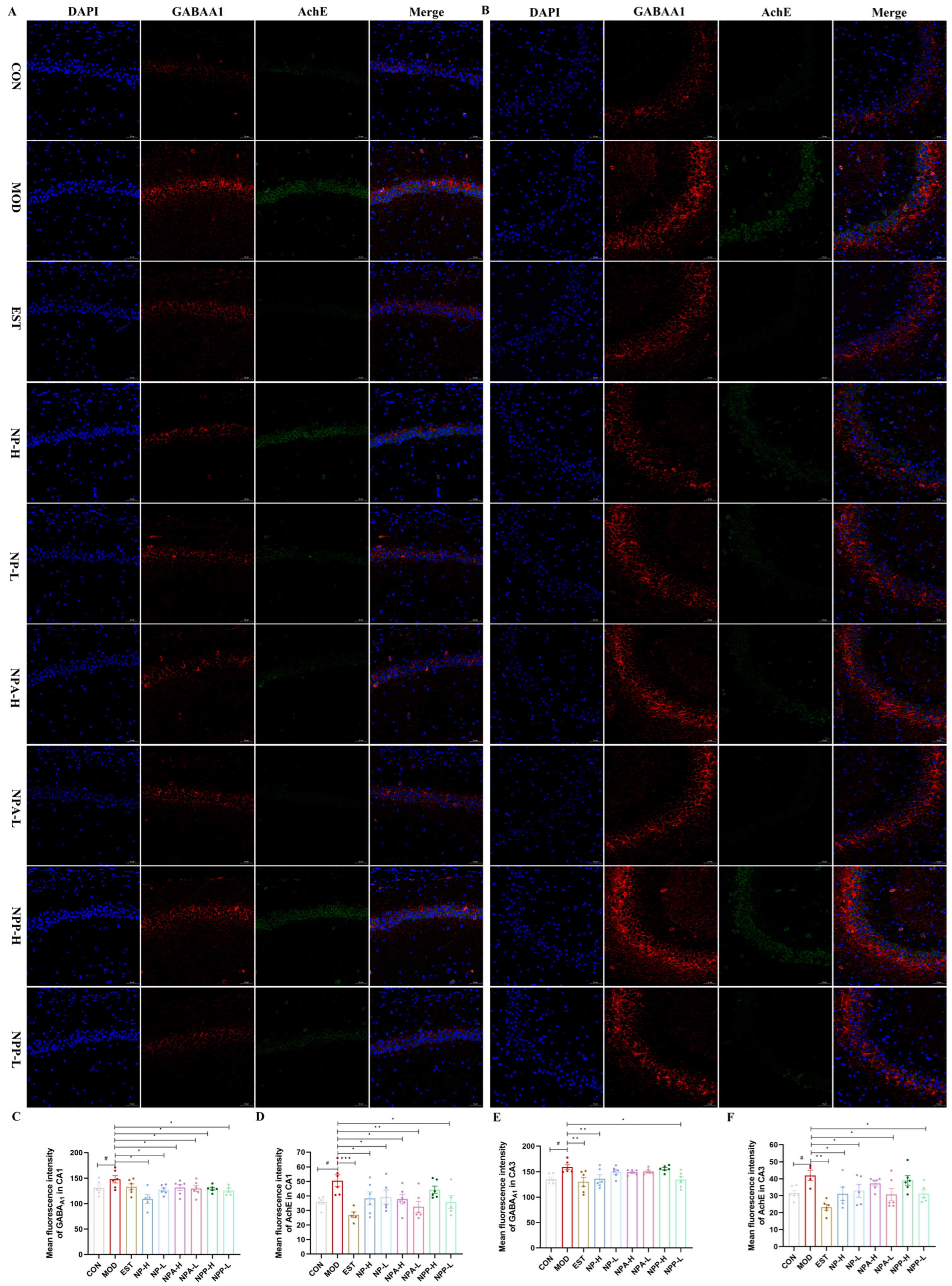
2.6. Results of Immunofluorescence Staining
3. Materials and Methods
3.1. Plant Materials
3.2. Sample Preparation
3.2.1. NP Extract Preparation
3.2.2. NPA Preparation
3.2.3. NPP Preparation
3.3. Reagents
3.4. Analysis of Secondary Metabolite by UPLC-MS/MS
3.5. Animal Administration
3.6. PCPA-Induced Insomnia Animal Model
3.7. Pentobarbital Sodium Sleep Experiment
3.8. HE Staining
3.9. Nissl Staining
3.10. Enzyme-Linked Immunosorbent Assay
3.11. Immunofluorescence Staining
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TCM | Traditional Chinese medicine |
| PCPA | DL-4-chlorophenylalanine |
| GABA | Gamma-aminobutyric acid |
| DA | Dopamine |
| ACh | Acetylcholine |
| 5-HT | 5-Hydroxytryptamine |
| IL-1 | Interleukin-1 |
| TNF-α | Tumor necrosis factor-alpha |
| GABRA1 | Gamma aminobutyric acid type A alpha1 receptor |
| 5-HT1A | 5-Hydroxytryptamine 1A receptor |
| DRD2 | Dopamine D2 receptor |
| AChE | Acetylcholinesterase |
| NREM | Non-rapid-eye-movement |
| REM | Rapid-eye-movement |
| NPA | Nelumbinis Plumula total alkaloid |
| NP | Nelumbinis Plumula |
| NPP | Nelumbinis Plumula crude polysaccharide |
| NeuN | Neuronal nuclei antigen |
| GFAP | Glial fibrillary acidic protein |
| HE | Hematoxylin and Eosin staining |
| UPLC | Ultra high performance liquid chromatogram |
| ELISA | Enzyme-linked immunosorbent assay |
References
- Jansen, P.R.; Watanabe, K.; Stringer, S.; Skene, N.; Bryois, J.; Hammerschlag, A.R.; de Leeuw, C.A.; Benjamins, J.S.; Muñoz-Manchado, A.B.; Nagel, M.; et al. Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nat. Genet. 2019, 51, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Zheng, X.; Qu, L.; Zhang, H.; Yuan, H.; Hui, J.; Mi, Y.; Ma, P.; Fan, D. Ginsenoside Rg5/Rk1 ameliorated sleep via regulating the GABAergic/serotoninergic signaling pathway in rodent model. Food Funct. 2020, 11, 1245. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.B.; Mccarthy, M.J.; Chen, C.Y.; Jain, S.; Gelernter, J.; He, F.; Heeringa, S.; Kessler, R.C.; Nock, M.K.; Ripke, S.; et al. Genome-wide analysis of insomnia disorder. Mol. Psychiatry 2018, 23, 2238–2250. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B. The link between sleep disorders and anxiety and depression disorders. Chin. J. Clin. 2018, 46, 131–133. [Google Scholar]
- Zhang, S.; Zhang, Y.; Zheng, Y.; Zhu, S.; Sun, J.; Deng, Y.; Wang, Q.; Zhai, Q. Dexmedetomidine attenuates sleep deprivation-induced inhibition of hippocampal neurogenesis via VEGF-VEGFR2 signaling and inhibits neuroinflammation. Biomed. Pharmacother. 2023, 165, 115085. [Google Scholar] [CrossRef]
- Dubey, A.K.; Handu, S.S.; Mediratta, P.K. Suvorexant: The first orexin receptor antagonist to treat insomnia. J. Pharmacol. Pharmacother. 2015, 6, 118–121. [Google Scholar] [CrossRef]
- Wang, Z.J.; Liu, J.F. The molecular basis of insomnia: Implication for therapeutic approaches. Drug Dev. Res. 2016, 77, 427–436. [Google Scholar] [CrossRef]
- Lv, Y.B.; Zhou, Q.; Yan, J.X.; Luo, L.S.; Zhang, J.L. Enzymolysis peptides from Mauremys mutica plastron improve the disorder of neurotransmitter system and facilitate sleep-promoting in the PCPA-induced insomnia mice. J. Ethnopharmacol. 2021, 274, 114047. [Google Scholar] [CrossRef]
- Bao, Y.; Zhou, H.; Fu, Y.; Wang, C.; Huang, Q. Zhumian Granules improves PCPA-induced insomnia by regulating the expression level of neurotransmitters and reducing neuronal apoptosis. J. Ethnopharmacol. 2024, 327, 118048. [Google Scholar] [CrossRef]
- Du, L.; Yang, D.; Wu, L.; Mei, L.; Wu, S.; Ba, Y.; Bao, Y.; Su, R.; Song, L. Integration of Gut Microbiota, Serum Metabolomic, and Network Pharmacology to Reveal the Anti Insomnia Mechanism of Mongolian Medicine Sugemule-4 Decoction on Insomnia Model Rats. Drug Des. Dev. Ther. 2024, 18, 2617–2639. [Google Scholar] [CrossRef]
- Li, T.; Wang, W.; Guo, Q.; Li, J.; Tang, T.; Wang, Y.; Liu, D.; Yang, K.; Li, J.; Deng, K.; et al. Rosemary (Rosmarinus officinalis L.) hydrosol based on serotonergic synapse for insomnia. J. Ethnopharmacol. 2024, 318 Pt B, 116984. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zhi, H.; Yang, C.; Wang, L.; Long, J.; Xiao, L.; Liang, J.; Huang, Y.; Zheng, X.; Zhao, S.; et al. Chemical composition of alkaloids of Plumula nelumbinis and their antioxidant activity from different habitats in China. Ind. Crops Prod. 2018, 125, 537–548. [Google Scholar] [CrossRef]
- Liu, M.; Jiang, Y.; Liu, R.; Liu, M.; Yi, L.; Liao, N.; Liu, S. Structural features guided “fishing” strategy to identification of flavonoids from lotus plumule in a self-built data “pool” by ultra-high performance liquid chromatography coupled with hybrid quadrupole-orbitrap high resolution mass spectrometry. J. Chromatogr. B 2019, 1124, 122–134. [Google Scholar] [CrossRef]
- Chen, S.; Li, X.; Wu, J.; Li, J.; Xiao, M.; Yang, Y.; Liu, Z.; Cheng, Y. Plumula Nelumbinis: A review of traditional uses, phytochemistry, pharmacology, pharmacokinetics and safety. J. Ethnopharmacol. 2021, 266, 113429. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, L.; Xie, J.; Chen, H.; Ou, G.; Zeng, L.; Li, Y.; Li, W.; Fan, H.; Zheng, J. Exploring the chemical composition, medicinal benefits, and antioxidant activity of Plumula nelumbinis essential oil from different habitats in China. Saudi Pharm. J. 2023, 31, 101829. [Google Scholar] [CrossRef]
- Li, Z.; Wu, W.; Liu, R.; Niu, B.; Chen, H.; Shentu, X.; Gao, H.; Chen, H. Flavonoid profiling of Plumula nelumbinis and evaluation of their anti-inflammatory effects on lipopolysaccharide-induced RAW 264.7 macrophages. eFood 2024, 5, e150. [Google Scholar] [CrossRef]
- Chen, Y.C.; Liu, Y.Y.; Chen, L.; Tang, D.M.; Zhao, Y.; Luo, X.D. Antimelanogenic Effect of Isoquinoline Alkaloids from Plumula Nelumbinis. J. Agric. Food Chem. 2023, 71, 16090–16101. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. National Pharmacopoeia Committee Pharmacopoeia; China Medical Science and Technology Press: Beijing, China, 2020; p. 285.
- Jo, K.; Kim, S.; Hong, K.B.; Suh, H.J. Nelumbo nucifera promotes non-rapid eye movement sleep by regulating GABAergic receptors in rat model. J. Ethnopharmacol. 2021, 26, 113511. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, A.; Lentini, G.; Rapacchietta, S.; Cinnirella, P.; Elia, M.; Ferri, R.; Bruni, O. The role of supplements and over-the-counter products to improve sleep in children: A systematic review. Int. J. Mol. Sci. 2023, 24, 7821. [Google Scholar] [CrossRef]
- Kim, S.; Jo, K.; Hong, K.B.; Han, S.H.; Suh, H.J. GABA and l-theanine mixture decreases sleep latency and improves NREM sleep. Pharm. Biol. 2019, 57, 65–73. [Google Scholar] [CrossRef]
- Liu, Y.M.; Li, J.C.; Gu, Y.F.; Qiu, R.H.; Huang, J.Y.; Xue, R.; Li, S.; Zhang, Y.; Zhang, K.; Zhang, Y.Z. Cannabidiol exerts sedative and hypnotic effects in normal and insomnia model mice through activation of 5-HT1A receptor. Neurochem. Res. 2024, 49, 1150–1165. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wu, T.; Jin, J.; Si, Y.; Wang, Y.; Ding, X.; Guo, T.; Wei, W. Periostracum Cicadae extract and N-acetyldopamine regulate the sleep-related neurotransmitters in PCPA-induced insomnia rats. Molecules 2024, 29, 3638. [Google Scholar] [CrossRef]
- Sayson, L.V.; Campomayor, N.B.; Ortiz, D.M.; Lee, H.J.; Balataria, S.; Park, S.; Lim, J.; Kang, H.; Kim, H.J.; Kim, M. Extracts of Prunella vulgaris enhanced pentobarbital-induced sleeping behavior in mice potentially via adenosine A2A receptor activity. Planta Med. 2024, 90, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Omond, S.E.T.; Hale, M.W.; Lesku, J.A. Neurotransmitters of sleep and wakefulness in flatworms. Sleep 2022, 45, zsac053. [Google Scholar] [CrossRef] [PubMed]
- Gott, J.A.; Stücker, S.; Kanske, P.; Haaker, J.; Dresler, M. Acetylcholine and metacognition during sleep. Conscious. Cogn. 2024, 117, 103608. [Google Scholar] [CrossRef]
- Irwin, M.R. Sleep and inflammation: Partners in sickness and in health. Nat. Rev. Immunol. 2019, 19, 702–715. [Google Scholar] [CrossRef]
- Zielinski, M.R.; Gibbons, A.J. Neuroinflammation, sleep, and circadian rhythms. Front. Cell. Infect. Microbiol. 2022, 12, 853096. [Google Scholar] [CrossRef]
- Duan, W.; Zhang, Y.P.; Hou, Z.; Huang, C.; Zhu, H.; Zhang, C.Q.; Yin, Q. Novel insights into NeuN: From neuronal marker to splicing regulator. Mol. Neurobiol. 2016, 53, 1637–1647. [Google Scholar] [CrossRef]
- Santamaría, G.; Rengifo, A.C.; Torres-Fernández, O. NeuN distribution in brain structures of normal and Zika-infected suckling mice. J. Mol. Histol. 2023, 54, 245–253. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, J.; Hou, Y.; Shi, X.; Liu, K. Prediction of clinical progression in nervous system diseases: Plasma glial fibrillary acidic protein (GFAP). Eur. J. Med. Res. 2024, 29, 51. [Google Scholar] [CrossRef]
- Bian, Z.; Zhang, W.; Tang, J.; Fei, Q.; Hu, M.; Chen, X.; Su, L.; Fei, C.; Ji, D.; Mao, C.; et al. Mechanisms underlying the action of Ziziphi spinosae semen in the treatment of insomnia: A study involving network pharmacology and experimental validation. Front. Pharmacol. 2021, 12, 752211. [Google Scholar] [CrossRef] [PubMed]
- Bruni, O.; Ferini-Strambi, L.; Giacomoni, E.; Pellegrino, P. Herbal remedies and their possible effect on the GABAergic system and sleep. Nutrients 2021, 13, 530. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yu, Q.; Liang, W. The regulation of dopaminergic nervous system on sleep-awakeness and cognition. Prog. Mod. Biomed. 2014, 14, 7172–7174. [Google Scholar]
- Zhao, K.; Qi, L.; Li, Q.; Wang, Y.; Qian, C.; Shi, Z. Self-absorbing multilayer skin-like composite with Phyllostachys nigra polysaccharides promotes wound healing. Adv. Compos. Hybrid Mater. 2024, 7, 225. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; Wang, D.; Li, H.; Tian, H.; Wang, Z.; Feng, S. Total Alkaloid Extract of Nelumbinis Plumula Promoted Sleep in PCPA-Induced Insomnia Rats by Affecting Neurotransmitters and Their Receptor Activities. Int. J. Mol. Sci. 2025, 26, 3684. https://doi.org/10.3390/ijms26083684
Wei W, Wang D, Li H, Tian H, Wang Z, Feng S. Total Alkaloid Extract of Nelumbinis Plumula Promoted Sleep in PCPA-Induced Insomnia Rats by Affecting Neurotransmitters and Their Receptor Activities. International Journal of Molecular Sciences. 2025; 26(8):3684. https://doi.org/10.3390/ijms26083684
Chicago/Turabian StyleWei, Wenjun, Dongge Wang, Hangying Li, Hongyu Tian, Zhilei Wang, and Suxiang Feng. 2025. "Total Alkaloid Extract of Nelumbinis Plumula Promoted Sleep in PCPA-Induced Insomnia Rats by Affecting Neurotransmitters and Their Receptor Activities" International Journal of Molecular Sciences 26, no. 8: 3684. https://doi.org/10.3390/ijms26083684
APA StyleWei, W., Wang, D., Li, H., Tian, H., Wang, Z., & Feng, S. (2025). Total Alkaloid Extract of Nelumbinis Plumula Promoted Sleep in PCPA-Induced Insomnia Rats by Affecting Neurotransmitters and Their Receptor Activities. International Journal of Molecular Sciences, 26(8), 3684. https://doi.org/10.3390/ijms26083684