
Flow Cytometry-Based Rapid Assay for Antigen Specific Antibody Relative Affinity in SRBC-Immunized Mouse Models

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
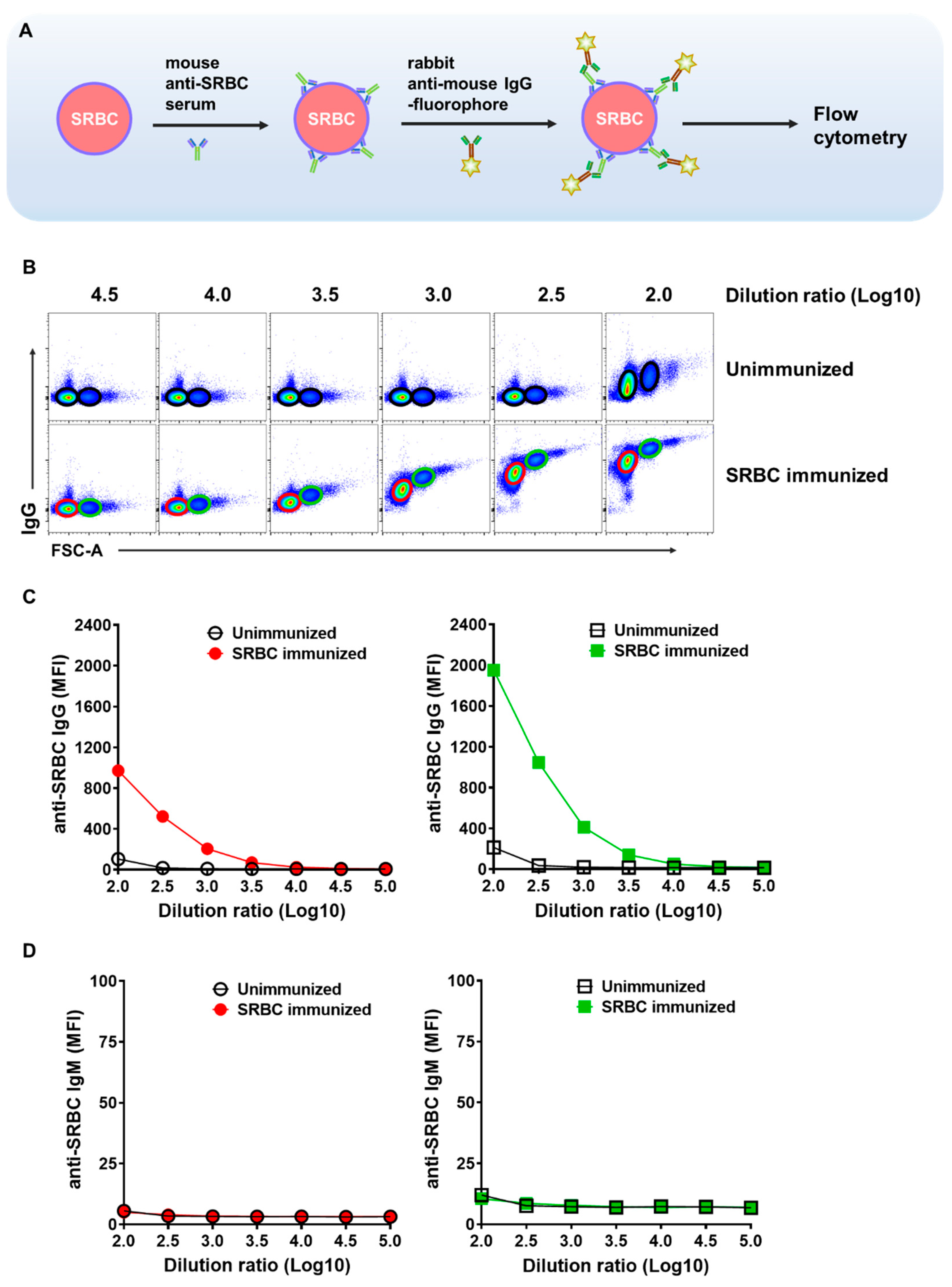
2.1. Detection of the Binding of Serum SRBC-Specific Antibodies to SRBC
2.2. Scheme of the Procedures for Assessing the Relative Affinity Levels of SRBC Antiserum
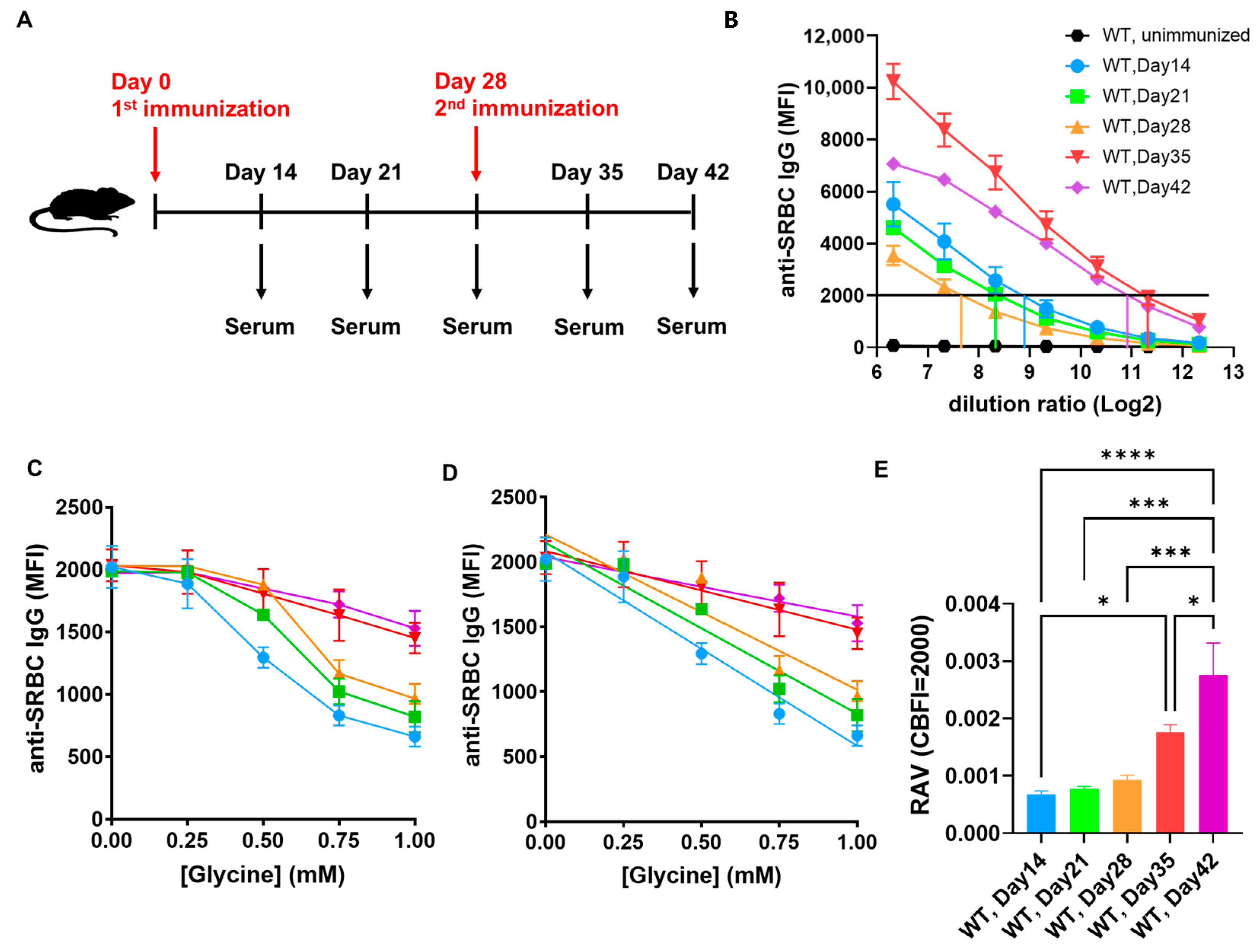
2.3. The Post-Immunizational Progressively Increased Relative Affinity Levels of SRBC-Immunized Wild-Type Mice Antisera Can Be Detected with FCRA
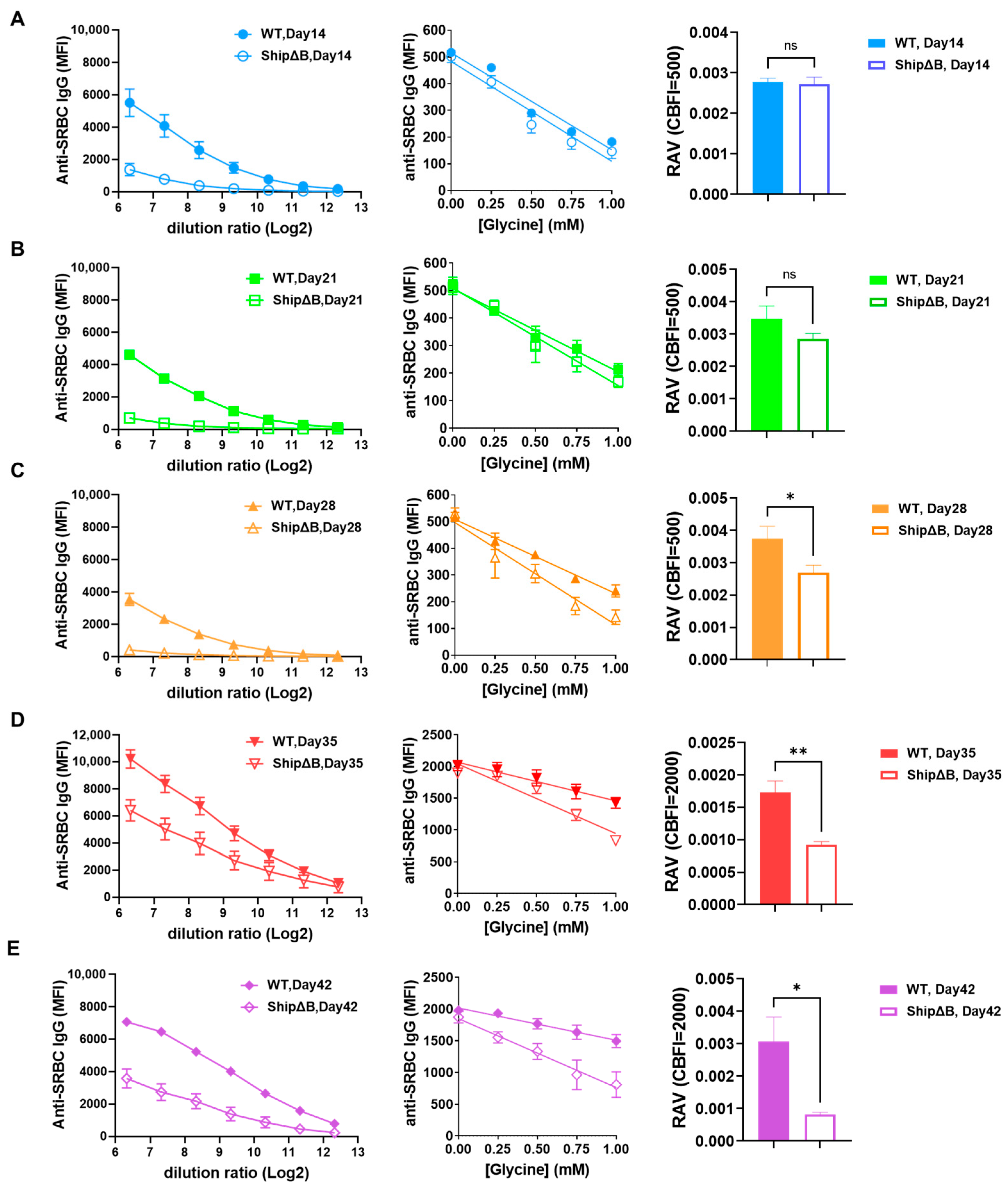
2.4. The AAM-Deficient ShipΔB Mice Antisera Has Lower Affinity Compared to That of WT Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. SRBC Immunization of the Mouse Models and Antiserum Preparation
4.3. Determination of the DF Values of the Test Samples for a CBFI
4.4. Preparation of the Serial Glycine-HCl Incubation Solutions
4.5. Determination of SRBC Antiserum Relative Affinity Level with Serial Pseudo Eluting Incubation Starting from a Pre-Defined CBFI
4.6. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lam, N.; Lee, Y.; Farber, D.L. A guide to adaptive immune memory. Nat. Rev. Immunol. 2024, 24, 810–829. [Google Scholar] [CrossRef] [PubMed]
- Ise, W.; Kurosaki, T. Plasma cell generation during T-cell-dependent immune responses. Int. Immunol. 2021, 33, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Zheng, T.; Zhou, P. T Follicular Helper Cell Subsets and the Associated Cytokine IL-21 in the Pathogenesis and Therapy of Asthma. Front. Immunol. 2019, 10, 2918. [Google Scholar] [CrossRef] [PubMed]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Annu. Rev. Immunol. 2022, 40, 413–442. [Google Scholar] [CrossRef]
- Bannard, O.; Cyster, J.G. Germinal centers: Programmed for affinity maturation and antibody diversification. Curr. Opin. Immunol. 2017, 45, 21–30. [Google Scholar] [CrossRef]
- Di Noia, J.M.; Neuberger, M.S. Molecular Mechanisms of Antibody Somatic Hypermutation. Annu. Rev. Biochem. 2007, 76, 1–22. [Google Scholar] [CrossRef]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef]
- Romberg, N.; Le Coz, C. Common variable immunodeficiency, cross currents, and prevailing winds. Immunol. Rev. 2024, 322, 233–243. [Google Scholar] [CrossRef]
- Akkaya, M.; Kwak, K.; Pierce, S.K. B cell memory: Building two walls of protection against pathogens. Nat. Rev. Immunol. 2020, 20, 229–238. [Google Scholar] [CrossRef]
- Roth, G.A.; Picece, V.C.T.M.; Ou, B.S.; Luo, W.; Pulendran, B.; Appel, E.A. Designing spatial and temporal control of vaccine responses. Nat. Rev. Mater. 2022, 7, 174–195. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef] [PubMed]
- Tiller, T.; Tsuiji, M.; Yurasov, S.; Velinzon, K.; Nussenzweig, M.C.; Wardemann, H. Autoreactivity in Human IgG+ Memory B Cells. Immunity 2007, 26, 205–213. [Google Scholar] [CrossRef]
- Piccoli, L.; Campo, I.; Fregni, C.S.; Rodriguez, B.M.F.; Minola, A.; Sallusto, F.; Luisetti, M.; Corti, D.; Lanzavecchia, A. Neutralization and clearance of GM-CSF by autoantibodies in pulmonary alveolar proteinosis. Nat. Commun. 2015, 6, 7375. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, D.H.; Jo, S.; Cho, M.J.; Cho, Y.R.; Lee, Y.J.; Byun, S. Immunomodulatory functional foods and their molecular mechanisms. Exp. Mol. Med. 2022, 54, 1–11. [Google Scholar] [CrossRef]
- Hooda, P.; Malik, R.; Bhatia, S.; Al-Harrasi, A.; Najmi, A.; Zoghebi, K.; Halawi, M.A.; Makeen, H.A.; Mohan, S. Phytoimmunomodulators: A review of natural modulators for complex immune system. Heliyon 2024, 10, e23790. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, F.; Jalayeri, M.H.T.; Montaseri, A.; MohamedKhosroshahi, L.; Baradaran, B. Microbial natural compounds and secondary metabolites as Immunomodulators: A review. Int. J. Biol. Macromol. 2024, 278, 134778. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, Q.; Rong, C.; Wang, S.; Zhao, Z.; Liu, Y.; Xu, J. Immunomodulatory Effects of Edible and Medicinal Mushrooms and Their Bioactive Immunoregulatory Products. J. Fungi 2020, 6, 269. [Google Scholar] [CrossRef]
- Cho, S.; Lee, H.-M.; Yu, I.-S.; Choi, Y.S.; Huang, H.-Y.; Hashemifar, S.S.; Lin, L.-L.; Chen, M.-C.; Afanasiev, N.D.; Khan, A.A.; et al. Differential cell-intrinsic regulations of germinal center B and T cells by miR-146a and miR-146b. Nat. Commun. 2018, 9, 2757. [Google Scholar] [CrossRef]
- Valeri, V.; Sochon, A.; Ye, C.; Mao, X.; Lecoeuche, D.; Fillatreau, S.; Weill, J.-C.; Reynaud, C.-A.; Hao, Y. B cell intrinsic and extrinsic factors impacting memory recall responses to SRBC challenge. Front. Immunol. 2022, 13, 873886. [Google Scholar] [CrossRef]
- Sato, T.; Ikegami, I.; Yanagi, M.; Ohyu, T.; Sugaya, T.; Shirato, S.; Tanemoto, M.; Kamiya, S.; Kikuchi, K.; Kamada, Y.; et al. Interleukin 9 mediates T follicular helper cell activation to promote antibody responses. Front. Immunol. 2024, 15, 1441407. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Ogikubo, Y.; Norimatsu, M.; Sasaki, Y.; Yasuda, A.; Saegusa, J.; Tamura, Y. Effect of lipopolysaccharide (LPS) injection on the immune responses of LPS-sensitive mice. J. Vet. Med. Sci. 2004, 66, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Ladics, G.S. The Sheep Erythrocyte T-Dependent Antibody Response (TDAR). In Immunotoxicity Testing: Methods and Protocols; DeWitt, J.C., Rockwell, C.E., Bowman, C.C., Eds.; Springer: New York, NY, USA, 2018; pp. 83–94. [Google Scholar]
- Ladics, G.S. Primary Immune Response to Sheep Red Blood Cells (SRBC) as the Conventional T-Cell Dependent Antibody Response (TDAR) Test. J. Immunotoxicol. 2007, 4, 149–152. [Google Scholar] [CrossRef] [PubMed]
- García-Mendoza, D.; van den Berg, H.; van den Brink, N.W. Environmental exposure to cadmium reduces the primary antibody-mediated response of wood mice (Apodemus sylvaticus) from differentially polluted locations in the Netherlands. Environ. Pollut. 2021, 289, 117909. [Google Scholar] [CrossRef]
- Abid, S.; Khajuria, A.; Parvaiz, Q.; Sidiq, T.; Bhatia, A.; Singh, S.; Ahmad, S.; Randhawa, M.; Satti, N.; Dutt, P. Immunomodulatory studies of a bioactive fraction from the fruit of Prunus cerasus in BALB/c mice. Int. Immunopharmacol. 2012, 12, 626–634. [Google Scholar] [CrossRef]
- Van Loveren, H.; Verlaan, A.P.; Vos, J.G. An enzyme-linked immunosorbent assay of anti-sheep red blood cell antibodies of the classes M, G, and A in the rat. Int. J. Immunopharmacol. 1991, 13, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, M.; Akkaya, B.; Kim, A.S.; Miozzo, P.; Sohn, H.; Pena, M.; Roesler, A.S.; Theall, B.P.; Henke, T.; Kabat, J.; et al. Toll-like receptor 9 antagonizes antibody affinity maturation. Nat. Immunol. 2018, 19, 255–266. [Google Scholar] [CrossRef]
- Pedersen, G.K.; Wørzner, K.; Andersen, P.; Christensen, D. Vaccine Adjuvants Differentially Affect Kinetics of Antibody and Germinal Center Responses. Front. Immunol. 2020, 11, 579761. [Google Scholar] [CrossRef]
- Francica, J.R.; Sheng, Z.; Zhang, Z.; Nishimura, Y.; Shingai, M.; Ramesh, A.; Keele, B.F.; Schmidt, S.D.; Flynn, B.J.; Darko, S.; et al. Analysis of immunoglobulin transcripts and hypermutation following SHIVAD8 infection and protein-plus-adjuvant immunization. Nat. Commun. 2015, 6, 6565. [Google Scholar] [CrossRef]
- Ladics, G.S. Use of SRBC antibody responses for immunotoxicity testing. Methods 2007, 41, 9–19. [Google Scholar] [CrossRef]
- Koganei, A.; Tsuchiya, T.; Samura, K.; Nishikibe, M. Use of Whole Sheep Red Blood Cells in ELISA to Assess Immunosuppression In Vivo. J. Immunotoxicol. 2007, 4, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Taliaferro, W.H.; Taliaferro, L.G.; Pizzi, A.K. Avidity and inter cellular transfer of hemolysin. J. Infect. Dis. 1959, 105, 197–221. [Google Scholar] [CrossRef] [PubMed]
- DeHeer, D.H.; Edgington, T.S. Estimation of relative antibody affinity at the level of the antibody-secreting cell during maturation of the immune response. Cell. Immunol. 1975, 18, 466–475. [Google Scholar] [CrossRef]
- DeLisi, C. Hemolytic plaque inhibition: The physical chemical limits on its use as an affinity assay. J. Immunol. 1976, 117, 2249–2257. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Telen, M.J. Red blood cell surface adhesion molecules: Their possible roles in normal human physiology and disease. Semin. Hematol. 2000, 37, 130–142. [Google Scholar] [CrossRef]
- Murphy, K.; Weaver, C. Janeway’s Immunobiology, 9th ed.; Garland Science: New York, NY, USA, 2017; p. 476. [Google Scholar]
- Zhang, W.; Zhang, H.; Liu, S.; Xia, F.; Kang, Z.; Zhang, Y.; Liu, Y.; Xiao, H.; Chen, L.; Huang, C.; et al. Excessive CD11c(+)Tbet(+) B cells promote aberrant T(FH) differentiation and affinity-based germinal center selection in lupus. Proc. Natl. Acad. Sci. USA 2019, 116, 18550–18560. [Google Scholar] [CrossRef]
- Cumpelik, A.; Heja, D.; Hu, Y.; Varano, G.; Ordikhani, F.; Roberto, M.P.; He, Z.; Homann, D.; Lira, S.A.; Dominguez-Sola, D.; et al. Dynamic regulation of B cell complement signaling is integral to germinal center responses. Nat. Immunol. 2021, 22, 757–768. [Google Scholar] [CrossRef]
- Ye, B.H.; Cattoretti, G.; Shen, Q.; Zhang, J.; Hawe, N.; de Waard, R.; Leung, C.; Nouri-Shirazi, M.; Orazi, A.; Chaganti, R.; et al. The BCL-6 proto-oncogene controls germinal-centre formation and Th2-type inflammation. Nat. Genet. 1997, 16, 161–170. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Jiang, Y.; Liu, S.; He, Q.; Han, C.; Su, D.; Ma, H.; Guo, X.; Zhang, Y.; Li, F.; et al. Flow Cytometry-Based Rapid Assay for Antigen Specific Antibody Relative Affinity in SRBC-Immunized Mouse Models. Int. J. Mol. Sci. 2025, 26, 3664. https://doi.org/10.3390/ijms26083664
Sun C, Jiang Y, Liu S, He Q, Han C, Su D, Ma H, Guo X, Zhang Y, Li F, et al. Flow Cytometry-Based Rapid Assay for Antigen Specific Antibody Relative Affinity in SRBC-Immunized Mouse Models. International Journal of Molecular Sciences. 2025; 26(8):3664. https://doi.org/10.3390/ijms26083664
Chicago/Turabian StyleSun, Chunli, Yuan Jiang, Shujun Liu, Qilin He, Chengyao Han, Dai Su, Hao Ma, Xingyu Guo, Yan Zhang, Fubin Li, and et al. 2025. "Flow Cytometry-Based Rapid Assay for Antigen Specific Antibody Relative Affinity in SRBC-Immunized Mouse Models" International Journal of Molecular Sciences 26, no. 8: 3664. https://doi.org/10.3390/ijms26083664
APA StyleSun, C., Jiang, Y., Liu, S., He, Q., Han, C., Su, D., Ma, H., Guo, X., Zhang, Y., Li, F., & Zhang, H. (2025). Flow Cytometry-Based Rapid Assay for Antigen Specific Antibody Relative Affinity in SRBC-Immunized Mouse Models. International Journal of Molecular Sciences, 26(8), 3664. https://doi.org/10.3390/ijms26083664