
Neural Mechanism of 5-HT4R-Mediated Memory Enhancement in Hippocampal–Prefrontal Circuits in a Mouse Model of Schizophrenia

 ,
, 
Abstract
1. Introduction
2. Results
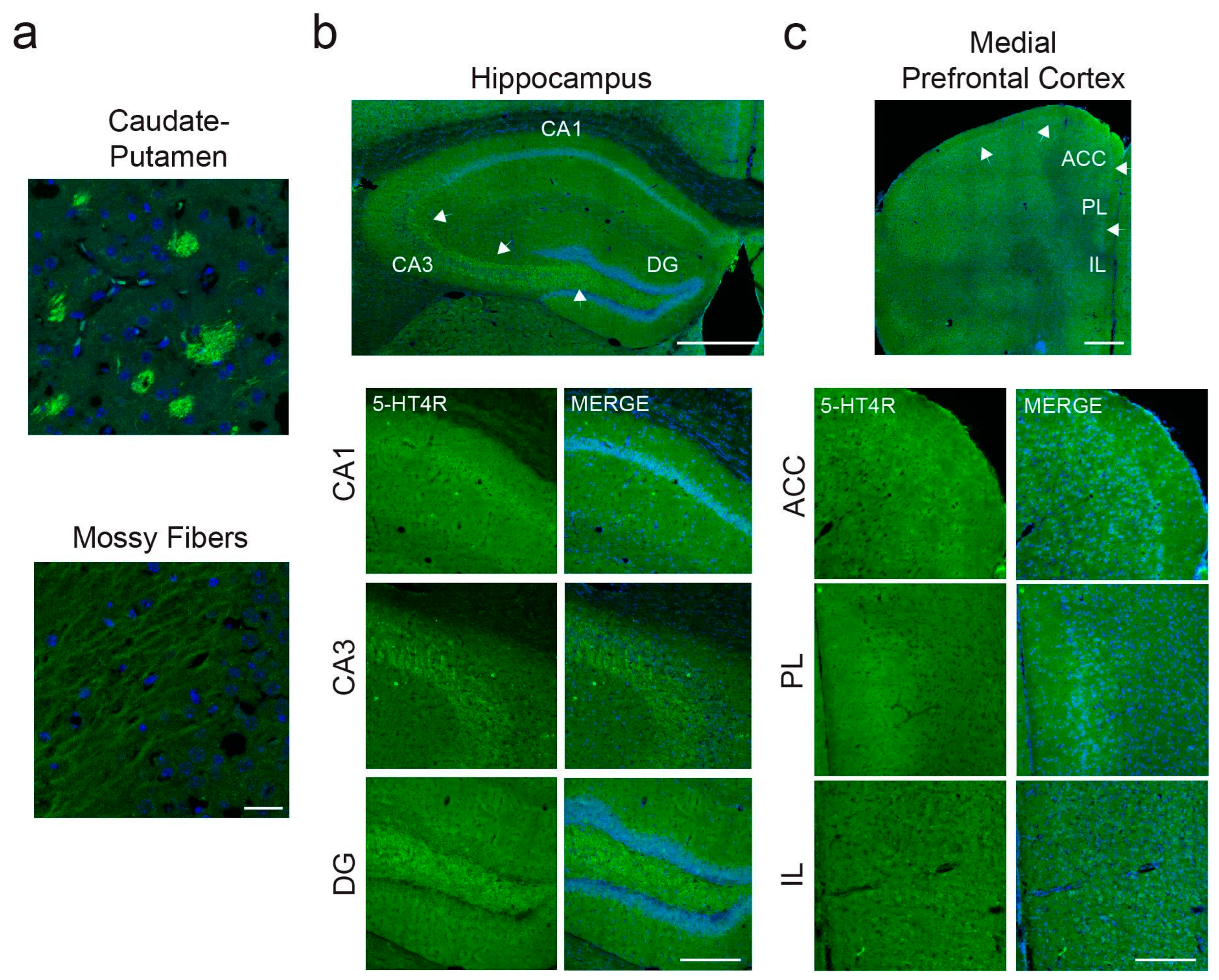
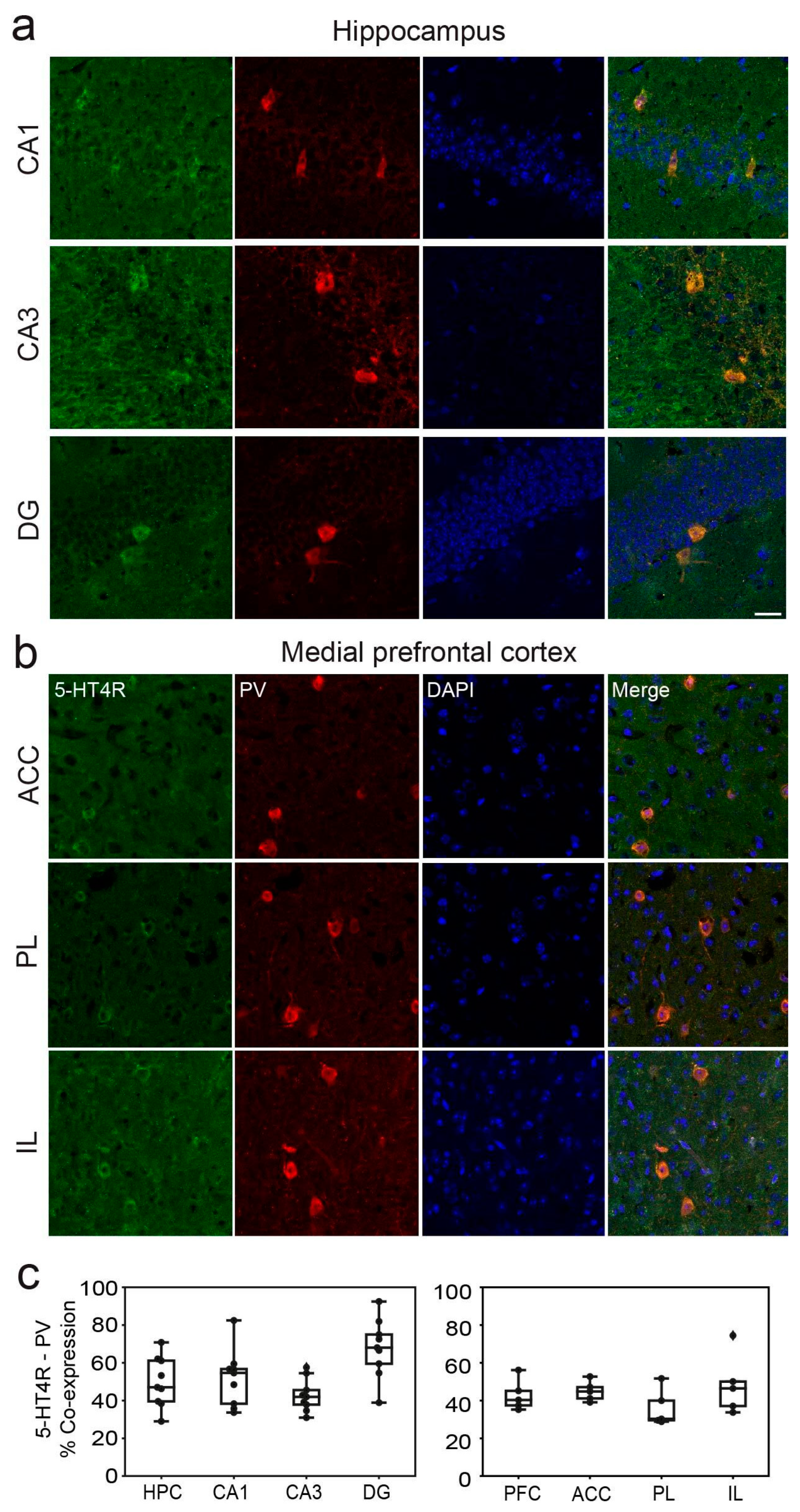
2.1. 5-HT4Rs Are Expressed by 30 to 60% of PV+ Interneurons in Mouse dHPC and mPFC and 15% of SST+ Neurons in the dHPC
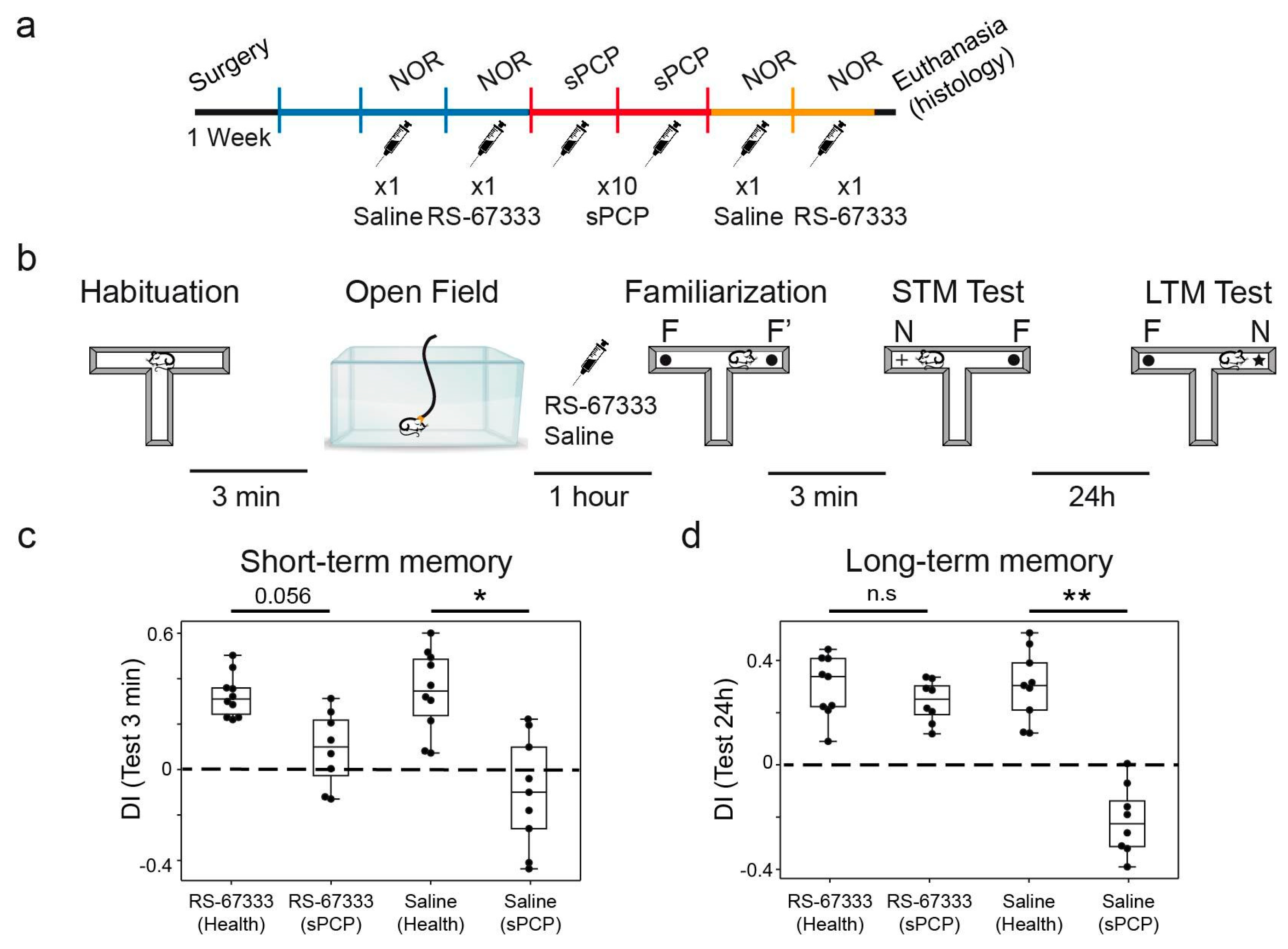
2.2. 5-HT4R Partial Agonist RS-67333 Improves Memory Performance and Reduces Anxiety in the sPCP Model of Schizophrenia
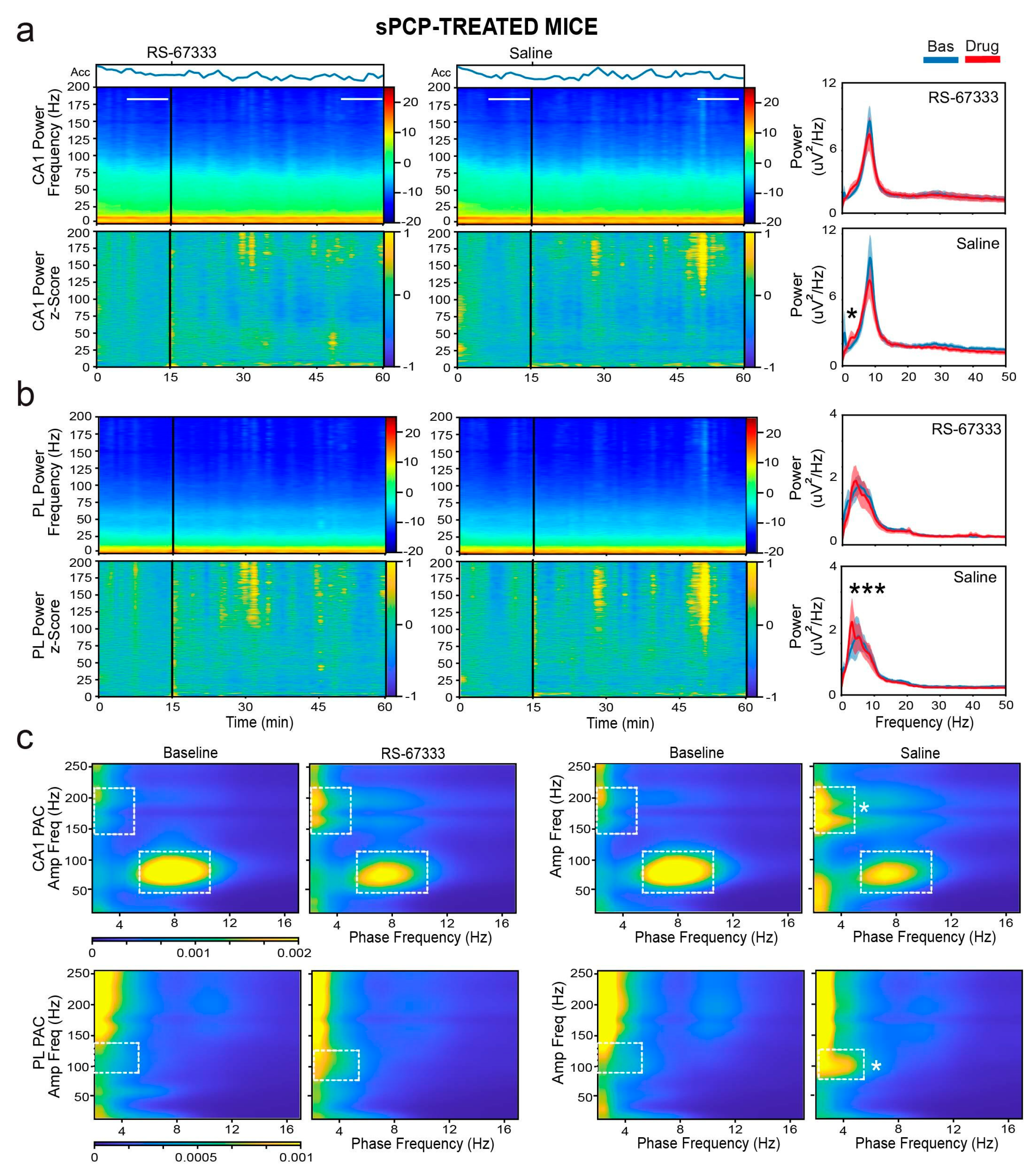
2.3. 5-HT4R Partial Agonist RS-67333 Mitigates sPCP-Induced Aberrant Delta Oscillations and Coupling in the Dorsal Hippocampus and Prelimbic Cortex
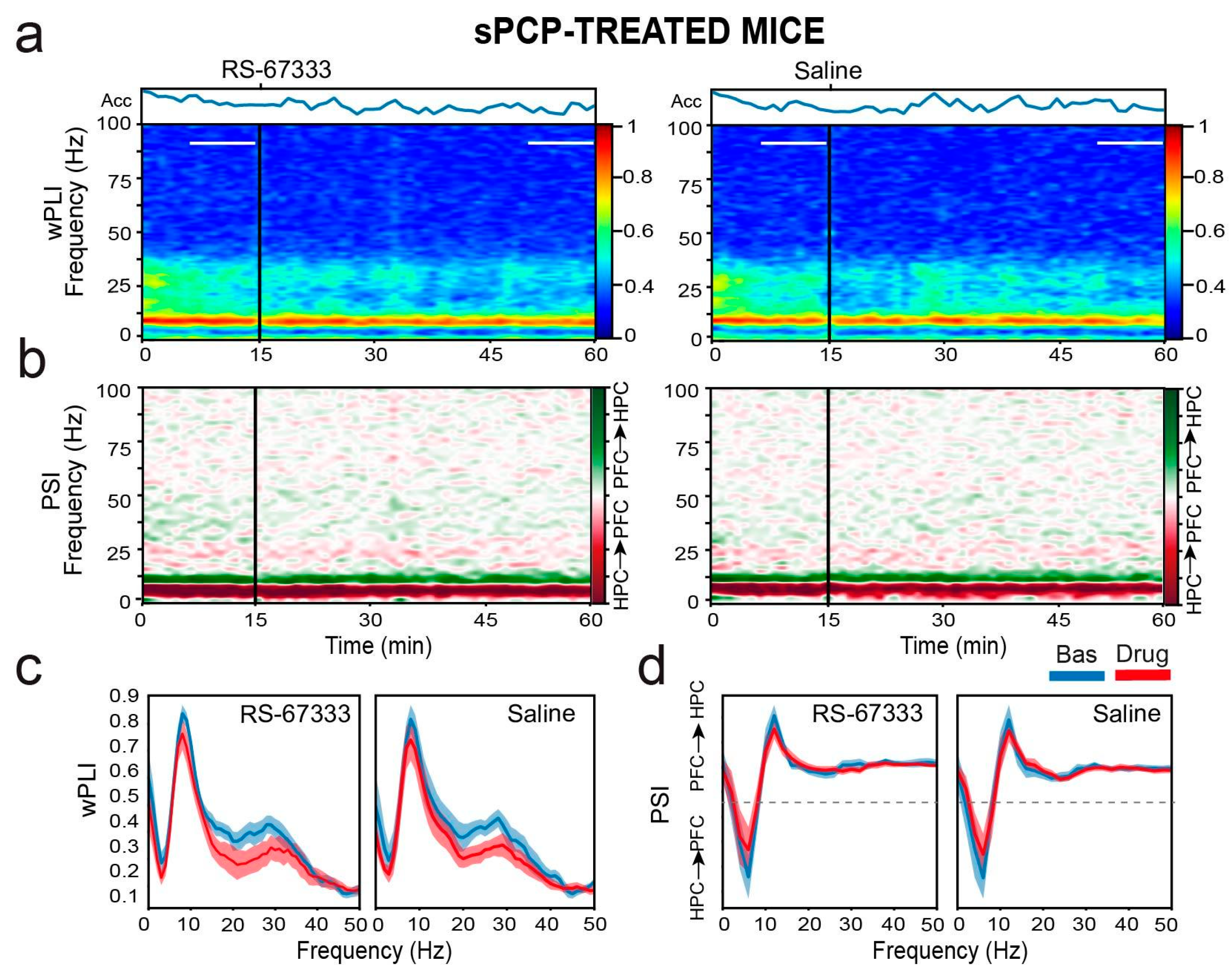
2.4. 5-HT4R Partial Agonist RS-67333 Does Not Influence Signal Directionality Between the Dorsal Hippocampus and the Prelimbic Cortex
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Immunohistochemistry
4.3. Surgical Procedures
4.4. Novel Object Recognition Test (NOR)
4.5. Pharmacology
4.6. Neurophysiological Recordings and Data Analyses
4.7. Statistical Analyses
5. Conclusions
- 5-HT4 receptors are not only expressed by pyramidal neurons in the HPC and mPFC but also by 30–60% of PV interneurons and 10–15% of SST interneurons.
- The 5-HT4R partial agonist RS-67333 improves memory performance and reduces anxiety in the sPCP model of schizophrenia.
- RS-67333 attenuates sPCP-induced increases in delta oscillations and associated cross-frequency coupling in both the CA1 area of the HPC and the PL mPFC without affecting the circuit’s connectivity.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meneses, A. Stimulation of 5-HT1A, 5-HT1B, 5-HT2A/2C, 5-HT3 and 5-HT4 Receptors or 5-HT Uptake Inhibition: Short- and Long-Term Memory. Behav. Brain Res. 2007, 184, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Karayol, R.; Medrihan, L.; Warner-Schmidt, J.L.; Fait, B.W.; Rao, M.N.; Holzner, E.B.; Greengard, P.; Heintz, N.; Schmidt, E.F. Serotonin Receptor 4 in the Hippocampus Modulates Mood and Anxiety. Mol. Psychiatry 2021, 26, 2334–2349. [Google Scholar] [CrossRef]
- Hagena, H.; Manahan-Vaughan, D. The Serotonergic 5-HT4 Receptor: A Unique Modulator of Hippocampal Synaptic Information Processing and Cognition. Neurobiol. Learn. Mem. 2017, 138, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Haahr, M.E.; Fisher, P.; Holst, K.; Madsen, K.; Jensen, C.G.; Marner, L.; Lehel, S.; Baaré, W.; Knudsen, G.; Hasselbalch, S. The 5-HT4 Receptor Levels in Hippocampus Correlates Inversely with Memory Test Performance in Humans. Hum. Brain Mapp. 2013, 34, 3066–3074. [Google Scholar] [CrossRef]
- King, M.V.; Marsden, C.A.; Fone, K.C.F. A Role for the 5-HT(1A), 5-HT4 and 5-HT6 Receptors in Learning and Memory. Trends Pharmacol. Sci. 2008, 29, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Pehrson, A.L.; Roberts, D.; Khawaja, A.; McNair, R. The Role of Serotonin Neurotransmission in Rapid Antidepressant Actions. Psychopharmacology 2022, 239, 1823–1838. [Google Scholar] [CrossRef]
- Ishii, T.; Kinoshita, K.-I.; Muroi, Y. Serotonin 5-HT4 Receptor Agonists Improve Facilitation of Contextual Fear Extinction in an MPTP-Induced Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5340. [Google Scholar] [CrossRef]
- Sgambato, V. The Serotonin 4 Receptor Subtype: A Target of Particular Interest, Especially for Brain Disorders. Int. J. Mol. Sci. 2024, 25, 5245. [Google Scholar] [CrossRef]
- Quiedeville, A.; Boulouard, M.; Hamidouche, K.; Costa-Aze, V.D.S.; Nee, G.; Rochais, C.; Dallemagne, P.; Fabis, F.; Freret, T.; Bouet, V. Chronic Activation of 5-HT4 Receptors or Blockade of 5-HT6 Receptors Improve Memory Performances. Behav. Brain Res. 2015, 293, 10–17. [Google Scholar] [CrossRef]
- Roux, C.M.; Leger, M.; Freret, T. Memory Disorders Related to Hippocampal Function: The Interest of 5-HT4Rs Targeting. Int. J. Mol. Sci. 2021, 22, 12082. [Google Scholar] [CrossRef]
- Lucas, G.; Rymar, V.V.; Du, J.; Mnie-Filali, O.; Bisgaard, C.; Manta, S.; Lambas-Senas, L.; Wiborg, O.; Haddjeri, N.; Piñeyro, G.; et al. Serotonin(4) (5-HT(4)) Receptor Agonists Are Putative Antidepressants with a Rapid Onset of Action. Neuron 2007, 55, 712–725. [Google Scholar] [CrossRef]
- Lamirault, L.; Simon, H. Enhancement of Place and Object Recognition Memory in Young Adult and Old Rats by RS 67333, a Partial Agonist of 5-HT4 Receptors. Neuropharmacology 2001, 41, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Restivo, L.; Roman, F.; Dumuis, A.; Bockaert, J.; Marchetti, E.; Ammassari-Teule, M. The Promnesic Effect of G-Protein-Coupled 5-HT4 Receptors Activation Is Mediated by a Potentiation of Learning-Induced Spine Growth in the Mouse Hippocampus. Neuropsychopharmacology 2008, 33, 2427–2434. [Google Scholar] [CrossRef]
- Kemp, A.; Manahan-Vaughan, D. The 5-Hydroxytryptamine4 Receptor Exhibits Frequency-Dependent Properties in Synaptic Plasticity and Behavioural Metaplasticity in the Hippocampal CA1 Region in Vivo. Cereb. Cortex 2005, 15, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Peñas-Cazorla, R.; Vilaró, M.T. Serotonin 5-HT4 Receptors and Forebrain Cholinergic System: Receptor Expression in Identified Cell Populations. Brain Struct. Funct. 2015, 220, 3413–3434. [Google Scholar] [CrossRef] [PubMed]
- Waeber, C.; Sebben, M.; Nieoullon, A.; Bockaert, J.; Dumuis, A. Regional Distribution and Ontogeny of 5-HT4 Binding Sites in Rodent Brain. Neuropharmacology 1994, 33, 527–541. [Google Scholar] [CrossRef]
- Varnäs, K.; Halldin, C.; Pike, V.W.; Hall, H. Distribution of 5-HT4 Receptors in the Postmortem Human Brain—an Autoradiographic Study Using [125I]SB 207710. Eur. Neuropsychopharmacol. 2003, 13, 228–234. [Google Scholar] [CrossRef]
- Vilaró, M.T.; Cortés, R.; Mengod, G. Serotonin 5-HT4 Receptors and Their mRNAs in Rat and Guinea Pig Brain: Distribution and Effects of Neurotoxic Lesions. J. Comp. Neurol. 2005, 484, 418–439. [Google Scholar] [CrossRef]
- Reynolds, G.P.; Mason, S.L.; Meldrum, A.; De Keczer, S.; Parnes, H.; Eglen, R.M.; Wong, E.H. 5-Hydroxytryptamine (5-HT)4 Receptors in Post Mortem Human Brain Tissue: Distribution, Pharmacology and Effects of Neurodegenerative Diseases. Br. J. Pharmacol. 1995, 114, 993–998. [Google Scholar] [CrossRef]
- Marner, L.; Gillings, N.; Madsen, K.; Erritzoe, D.; Baaré, W.F.; Svarer, C.; Hasselbalch, S.G.; Knudsen, G.M. Brain Imaging of Serotonin 4 Receptors in Humans with [11C]SB207145-PET. Neuroimage 2010, 50, 855–861. [Google Scholar] [CrossRef]
- Celada, P.; Puig, M.V.; Artigas, F. Serotonin Modulation of Cortical Neurons and Networks. Front. Integr. Neurosci. 2013, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Gulledge, A.T. Serotonin and Prefrontal Cortex Function: Neurons, Networks, and Circuits. Mol. Neurobiol. 2011, 44, 449–464. [Google Scholar] [CrossRef]
- Leiser, S.C.; Li, Y.; Pehrson, A.L.; Dale, E.; Smagin, G.; Sanchez, C. Serotonergic Regulation of Prefrontal Cortical Circuitries Involved in Cognitive Processing: A Review of Individual 5-HT Receptor Mechanisms and Concerted Effects of 5-HT Receptors Exemplified by the Multimodal Antidepressant Vortioxetine. ACS Chem. Neurosci. 2015, 6, 970–986. [Google Scholar] [CrossRef] [PubMed]
- Alemany-González, M.; Gener, T.; Nebot, P.; Vilademunt, M.; Dierssen, M.; Puig, M.V. Prefrontal-Hippocampal Functional Connectivity Encodes Recognition Memory and Is Impaired in Intellectual Disability. Proc. Natl. Acad. Sci. USA 2020, 117, 11788–11798. [Google Scholar] [CrossRef]
- Sigurdsson, T.; Duvarci, S. Hippocampal-Prefrontal Interactions in Cognition, Behavior and Psychiatric Disease. Front. Syst. Neurosci. 2016, 9, 190. [Google Scholar] [CrossRef]
- Delgado-Sallent, C.; Gener, T.; Nebot, P.; López-Cabezón, C.; Puig, M.V. Neural Substrates of Cognitive Impairment in a NMDAR Hypofunction Mouse Model of Schizophrenia and Partial Rescue by Risperidone. Front. Cell. Neurosci. 2023, 17, 1152248. [Google Scholar] [CrossRef] [PubMed]
- Gener, T.; Campo, A.T.; Alemany-González, M.; Nebot, P.; Delgado-Sallent, C.; Chanovas, J.; Puig, M.V. Serotonin 5-HT1A, 5-HT2A and Dopamine D2 Receptors Strongly Influence Prefronto-Hippocampal Neural Networks in Alert Mice: Contribution to the Actions of Risperidone. Neuropharmacology 2019, 158, 107743. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Gener, T.; Hidalgo-Nieves, S.; López-Cabezón, C. Neural Substrates of Psychosis and Cognitive Impairment in Mouse Models of Schizophrenia; IntechOpen: London, UK, 2025; ISBN 978-1-83634-412-4. [Google Scholar]
- Cardin, J.A.; Carlén, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L.-H.; Moore, C.I. Driving Fast-Spiking Cells Induces Gamma Rhythm and Controls Sensory Responses. Nature 2009, 459, 663–667. [Google Scholar] [CrossRef]
- Hoseini, M.S.; Higashikubo, B.; Cho, F.S.; Chang, A.H.; Clemente-Perez, A.; Lew, I.; Ciesielska, A.; Stryker, M.P.; Paz, J.T.; Francisco, S.; et al. Gamma Rhythms and Visual Information in Mouse V1 Specifically Modulated by Somatostatin+ Neurons in Reticular Thalamus. Elife 2021, 10, e61437. [Google Scholar] [CrossRef]
- Antonoudiou, P.; Tan, Y.L.; Kontou, G.; Upton, A.L.; Mann, E.O. Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations. J. Neurosci. 2020, 40, 7668–7687. [Google Scholar] [CrossRef]
- Castañé, A.; Santana, N.; Artigas, F. PCP-Based Mice Models of Schizophrenia: Differential Behavioral, Neurochemical and Cellular Effects of Acute and Subchronic Treatments. Psychopharmacology 2015, 232, 4085–4097. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, H.Y.; Rajagopal, L.; Huang, M.; Oyamada, Y.; Kwon, S.; Horiguchi, M. Translating the N-Methyl-d-Aspartate Receptor Antagonist Model of Schizophrenia to Treatments for Cognitive Impairment in Schizophrenia. Int. J. Neuropsychopharmacol. 2013, 16, 2181–2194. [Google Scholar] [CrossRef] [PubMed]
- Rebholz, H.; Friedman, E.; Castello, J. Alterations of Expression of the Serotonin 5-HT4 Receptor in Brain Disorders. Int. J. Mol. Sci. 2018, 19, 3581. [Google Scholar] [CrossRef]
- Horiguchi, M.; Huang, M.; Meltzer, H.Y. Interaction of mGlu2/3 Agonism with Clozapine and Lurasidone to Restore Novel Object Recognition in Subchronic Phencyclidine-Treated Rats. Psychopharmacology 2011, 217, 13–24. [Google Scholar] [CrossRef]
- Warburton, E.C.; Brown, M. Neural Circuitry for Rat Recognition Memory. Behav. Brain Res. 2015, 285, 131–139. [Google Scholar] [CrossRef]
- Alemany-González, M.; Vilademunt, M.; Gener, T.; Puig, M.V. Postnatal Environmental Enrichment Enhances Memory through Distinct Neural Mechanisms in Healthy and Trisomic Female Mice. Neurobiol. Dis. 2022, 173, 105841. [Google Scholar] [CrossRef] [PubMed]
- Scheffer-Teixeira, R.; Tort, A.B.L. Unveiling Fast Field Oscillations through Comodulation. eNeuro 2017, 4, ENEURO.0079-17.2017. [Google Scholar] [CrossRef]
- Delgado-Sallent, C.; Nebot, P.; Gener, T.; Fath, A.B.; Timplalexi, M.; Puig, M.V. Atypical, but Not Typical, Antipsychotic Drugs Reduce Hypersynchronized Prefrontal-Hippocampal Circuits during Psychosis-like States in Mice: Contribution of 5-HT2A and 5-HT1A Receptors. Cereb. Cortex 2022, 32, 3472–3487. [Google Scholar] [CrossRef]
- Nolte, G.; Ziehe, A.; Nikulin, V.V.; Schlögl, A.; Krämer, N.; Brismar, T.; Müller, K.-R. Robustly Estimating the Flow Direction of Information in Complex Physical Systems. Phys. Rev. Lett. 2008, 100, 234101. [Google Scholar] [CrossRef]
- Wawra, M.; Fidzinski, P.; Heinemann, U.; Mody, I.; Behr, J. 5-HT4-Receptors Modulate Induction of Long-Term Depression but Not Potentiation at Hippocampal Output Synapses in Acute Rat Brain Slices. PLoS ONE 2014, 9, e88085. [Google Scholar] [CrossRef]
- Marchetti, E.; Chaillan, F.A.; Dumuis, A.; Bockaert, J.; Soumireu-Mourat, B.; Roman, F.S. Modulation of Memory Processes and Cellular Excitability in the Dentate Gyrus of Freely Moving Rats by a 5-HT4 Receptors Partial Agonist, and an Antagonist. Neuropharmacology 2004, 47, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Siekmeier, P.J.; Stufflebeam, S.M. Patterns of Spontaneous Magnetoencephalographic Activity in Patients with Schizophrenia. J. Clin. Neurophysiol. 2010, 27, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.J.; Kopell, N.J.; Traub, R.D.; Whittington, M.A. Aberrant Network Activity in Schizophrenia. Trends Neurosci. 2017, 40, 371–382. [Google Scholar] [CrossRef]
- Rosas-Arellano, A.; Villalobos-González, J.B.; Palma-Tirado, L.; Beltrán, F.A.; Cárabez-Trejo, A.; Missirlis, F.; Castro, M.A. A Simple Solution for Antibody Signal Enhancement in Immunofluorescence and Triple Immunogold Assays. Histochem. Cell Biol. 2016, 146, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Gulinello, M.; Mitchell, H.A.; Chang, Q.; Timothy O’brien, W.; Zhou, Z.; Abel, T.; Wang, L.; Corbin, J.G.; Veeraragavan, S.; Samaco, R.C.; et al. Rigor and Reproducibility in Rodent Behavioral Research. Neurobiol. Learn. Mem. 2019, 165, 106780. [Google Scholar] [CrossRef]
- Tort, A.B.L.; Kramer, M.A.; Thorn, C.; Gibson, D.J.; Kubota, Y.; Graybiel, A.M.; Kopell, N.J. Dynamic Cross-Frequency Couplings of Local Field Potential Oscillations in Rat Striatum and Hippocampus during Performance of a T-Maze Task. Proc. Natl. Acad. Sci. USA 2008, 105, 20517–20522. [Google Scholar] [CrossRef]
- Hardmeier, M.; Hatz, F.; Bousleiman, H.; Schindler, C.; Stam, C.J.; Fuhr, P. Reproducibility of Functional Connectivity and Graph Measures Based on the Phase Lag Index (PLI) and Weighted Phase Lag Index (wPLI) Derived from High Resolution EEG. PLoS ONE 2014, 9, e108648. [Google Scholar] [CrossRef]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase Lag Index: Assessment of Functional Connectivity from Multi Channel EEG and MEG with Diminished Bias from Common Sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An Improved Index of Phase-Synchronization for Electrophysiological Data in the Presence of Volume-Conduction, Noise and Sample-Size Bias. Neuroimage 2011, 55, 1548–1565. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub- Area | Sex | Number of Animals | Mean PV+ Count per Section | % Co- Expression | Number of Animals | Mean SST+ Count per Section | % Co- Expression |
|---|---|---|---|---|---|---|---|
| CA1 | Female | 5 | 28 ± 3 | 53 ± 10 | 2 | 14 ± 6 | 25 ± 14 |
| Male | 4 | 23 ± 4 | 50 ± 6 | 3 | 16 ± 3 | 10 ± 7 | |
| All | 9 | 26 ± 3 | 52 ± 6 | 5 | 15 ± 3 | 16 ± 7 | |
| CA3 | Female | 5 | 19 ± 2 | 45 ± 8 | 2 | 14 ± 9 | 21 ± 15 |
| Male | 4 | 22 ± 3 | 40 ± 5 | 3 | 7 ± 2 | 16 ± 11 | |
| All | 9 | 20 ± 2 | 43 ± 5 | 5 | 10 ± 4 | 18 ± 9 | |
| DG | Female | 5 | 4 ± 1 | 70 ± 14 | 2 | 14 ± 9 | 8 ± 9 |
| Male | 4 | 8 ± 1 | 65 ± 10 | 3 | 4 ± 1 | 4 ± 6 | |
| All | 9 | 6 ± 1 | 68 ± 9 | 5 | 8 ± 4 | 6 ± 5 | |
| ACC | Female | 4 | 65 ± 22 | 43 ± 9 | 3 | 37 ± 9 | 7 ± 5 |
| Male | 2 | 51 ± 17 | 43 ± 9 | 3 | 29 ± 6 | 4 ± 3 | |
| All | 6 | 60 ± 14 | 43 ± 7 | 6 | 33 ± 5 | 6 ± 3 | |
| PL | Female | 4 | 43 ± 14 | 34 ± 10 | 3 | 18 ± 5 | 10 ± 7 |
| Male | 2 | 37 ± 9 | 29 ± 4 | 3 | 17 ± 6 | 4 ± 4 | |
| All | 6 | 41 ± 8 | 32 ± 6 | 6 | 18 ± 4 | 7 ± 4 | |
| IL | Female | 4 | 37 ± 11 | 43 ± 14 | 3 | 18 ± 7 | 18 ± 10 |
| Male | 2 | 46 ± 12 | 42 ± 8 | 3 | 17 ± 3 | 6 ± 3 | |
| All | 6 | 40 ± 8 | 43 ± 8 | 6 | 17 ± 4 | 12 ± 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gener, T.; Hidalgo-Nieves, S.; López-Cabezón, C.; Puig, M.V. Neural Mechanism of 5-HT4R-Mediated Memory Enhancement in Hippocampal–Prefrontal Circuits in a Mouse Model of Schizophrenia. Int. J. Mol. Sci. 2025, 26, 3659. https://doi.org/10.3390/ijms26083659
Gener T, Hidalgo-Nieves S, López-Cabezón C, Puig MV. Neural Mechanism of 5-HT4R-Mediated Memory Enhancement in Hippocampal–Prefrontal Circuits in a Mouse Model of Schizophrenia. International Journal of Molecular Sciences. 2025; 26(8):3659. https://doi.org/10.3390/ijms26083659
Chicago/Turabian StyleGener, Thomas, Sara Hidalgo-Nieves, Cristina López-Cabezón, and Maria Victoria Puig. 2025. "Neural Mechanism of 5-HT4R-Mediated Memory Enhancement in Hippocampal–Prefrontal Circuits in a Mouse Model of Schizophrenia" International Journal of Molecular Sciences 26, no. 8: 3659. https://doi.org/10.3390/ijms26083659
APA StyleGener, T., Hidalgo-Nieves, S., López-Cabezón, C., & Puig, M. V. (2025). Neural Mechanism of 5-HT4R-Mediated Memory Enhancement in Hippocampal–Prefrontal Circuits in a Mouse Model of Schizophrenia. International Journal of Molecular Sciences, 26(8), 3659. https://doi.org/10.3390/ijms26083659