
Extracellular Vesicle-Derived microRNA Crosstalk Between Equine Chondrocytes and Synoviocytes—An In Vitro Approach

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Demographics of EV-Donor and EV-Recipient Cells
2.2. EV Concentration and Size
2.3. 5-EU RNA Capture from Recipient Cells
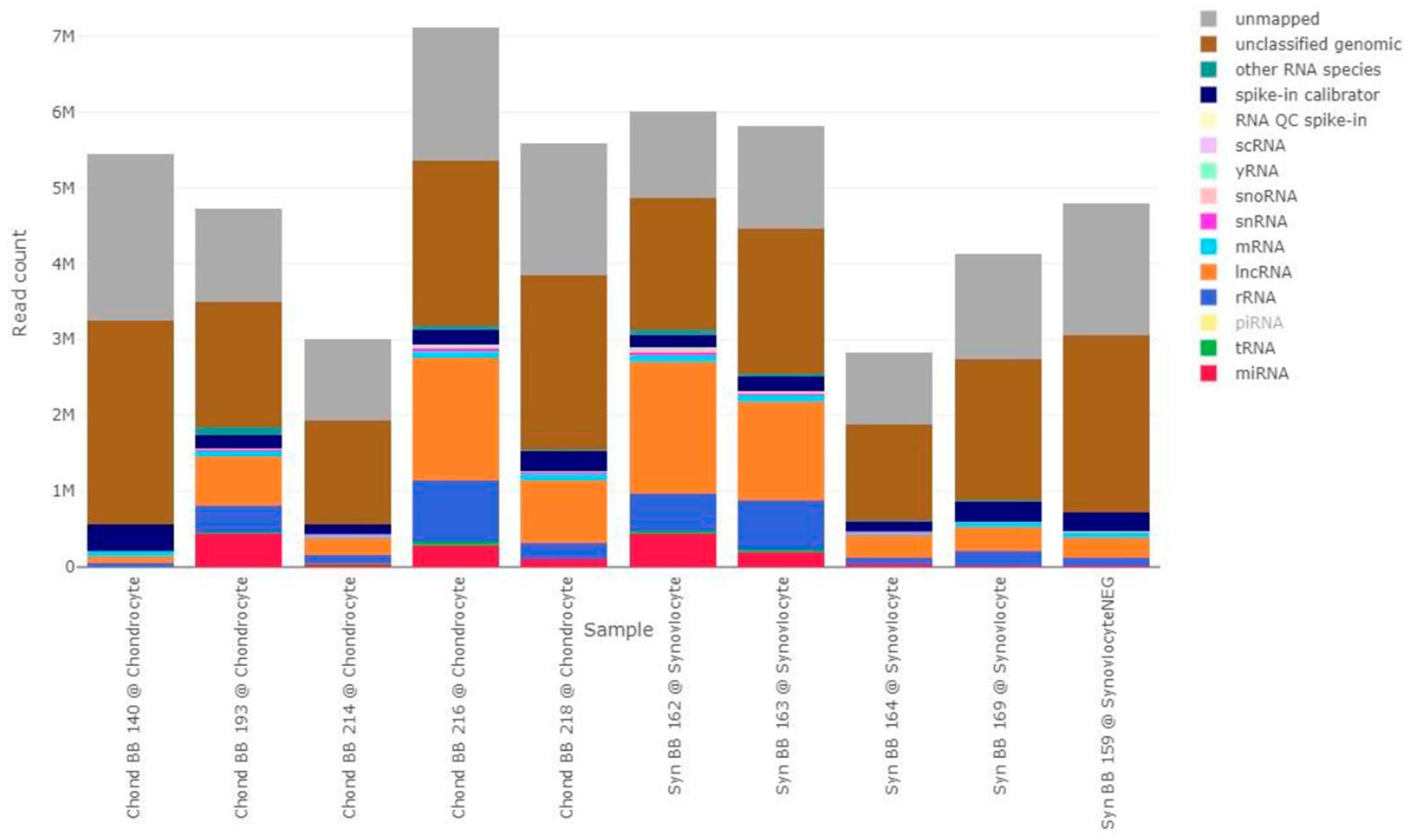
2.4. Sequencing Results
2.4.1. Data Overview
Predicted mRNA Targets of Unique miRNAs
Unsupervised Analysis
Differential Expression Analysis
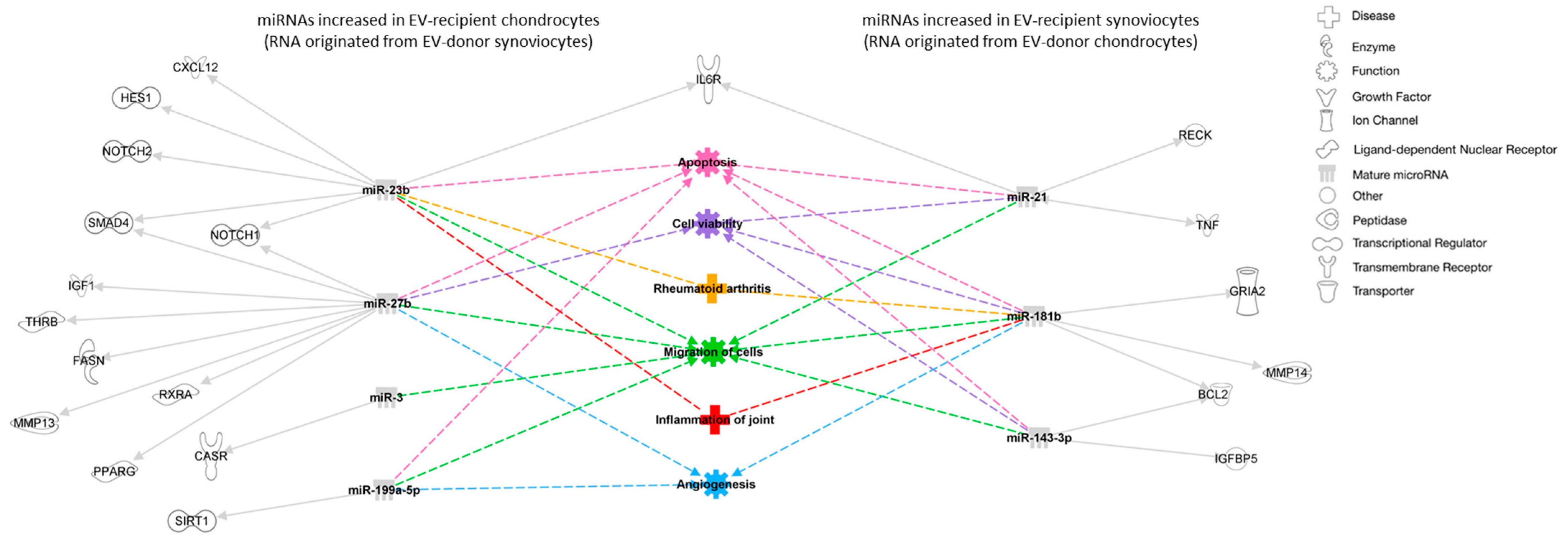
Target Prediction and Pathway Analysis of Differentially Expressed miRNAs
3. Discussion
4. Materials and Methods
4.1. Tissue Collection
4.2. Isolation of Primary Chondrocytes and Synoviocytes
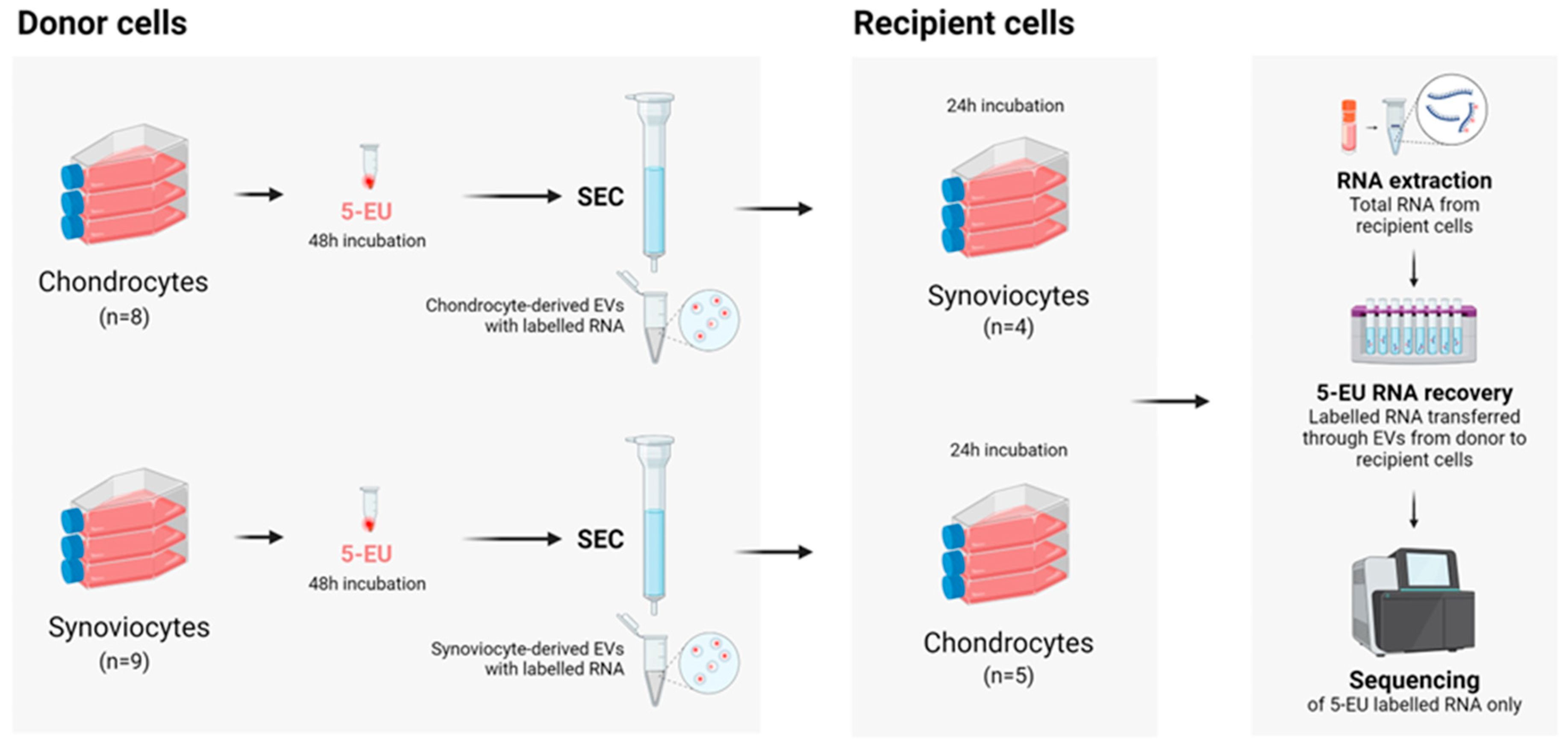
4.3. EV-Donor Cell Set Up and RNA Labelling
4.4. EV Isolation and Characterization
4.5. EV-Recipient Cell Setup and EV Incubation
4.6. RNA Extraction
4.7. Biotinylation of RNA and RNA Capture
4.8. Small RNA Sequencing
4.9. Data Analysis
4.9.1. Small RNA Sequencing Analysis
4.9.2. Target Prediction and Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-EU | 5-Ethynyl uridine |
| BCL2 | B-cell lymphoma 2 apoptosis regulator |
| CASR | Calcium-sensing receptor |
| CI | Confidence interval |
| CXCL12 | C-X-C motif chemokine ligand 12 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECM | Extracellular matrix |
| EV | Extracellular vesicle |
| FASN | Fatty acid synthase |
| FBS | Fetal bovine serum |
| FDR | False discovery rate |
| GRIA2 | Glutamate ionotropic receptor AMPA-type subunit 2 |
| HES1 | Hes family bHLH transcription factor 1 |
| IGF1 | Insulin-like growth factor 1 |
| IGFBP5 | Insulin-like growth factor-binding protein 5 |
| IL | Interleukin |
| IL6R | Interleukin 6 receptor |
| IPA | Ingenuity Pathway Analysis |
| IRS1 | Insulin receptor substrate 1 |
| lncRNA | Long non-coding RNA |
| logFC | Log fold change |
| Max | Maximum |
| MCP | Metacarpophalangeal |
| Min | Minimum |
| MMP | Matrix metalloproteinase |
| miRNA | microRNA |
| mRNA | Messenger RNA |
| NOTCH | Notch receptor |
| OA | Osteoarthritis |
| P | Passage |
| PBS | Phosphate-buffered saline |
| PCA | Principal component analysis |
| Pen/Strep | Penicillin–streptomycin |
| piRNA | Piwi-interfering RNA |
| PPARG | Peroxisome proliferator-activated receptor gamma |
| QC | Quality control |
| RECK | Reversion-inducing cysteine-rich protein with Kazal motifs |
| RPM | Reads per million |
| rRNA | Ribosomal RNA |
| RXRA | Retinoid X receptor alpha |
| SEC | Size exclusion chromatography |
| scRNA | Small conditional RNA |
| SIRT1 | Sirtuin 1 |
| SMAD4 | Mothers against decapentaplegic homolog 4 |
| snRNA | Small nuclear RNA |
| snoRNA | Small nucleolar RNA |
| SF | Synovial fluid |
| SD | Standard deviation |
| THRB | Thyroid hormone receptor beta |
| TNF | Tumor necrosis factor |
| tRNA | Transfer RNA |
| VCAM | Vascular cell adhesion molecule 1 |
| VEGF | Vascular endothelial growth factor |
References
- Fox, A.J.S.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Krakowski, P.; Rejniak, A.; Sobczyk, J.; Karpiński, R. Cartilage Integrity: A Review of Mechanical and Frictional Properties and Repair Approaches in Osteoarthritis. Healthcare 2024, 12, 1648. [Google Scholar] [CrossRef] [PubMed]
- Khaniki, H.B.; Ghayesh, M.H.; Chin, R.; Amabili, M. Hyperelastic structures: A review on the mechanics and biomechanics. Int. J. Nonlin Mech. 2023, 148, 104275. [Google Scholar] [CrossRef]
- Tarafder, S.; Lee, C.H. Chapter 14—Synovial Joint: In Situ Regeneration of Osteochondral and Fibrocartilaginous Tissues by Homing of Endogenous Cells. In In Situ Tissue Regeneration: Host Cell Recruitment and Biomaterial Design; Lee, S.J., Yoo, J.J., Atala, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 253–273. [Google Scholar] [CrossRef]
- Iwanaga, T.; Shikichi, M.; Kitamura, H.; Yanase, H.; Nozawa-Inoue, K. Morphology and Functional Roles of Synoviocytes in the Joint. Arch. Histol. Cytol. 2000, 63, 17–31. [Google Scholar] [CrossRef]
- Veale, D.J.; Firestein, G.S. Synovium. In Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Firestein, G.S., Budd, R.C., Gabriel, S.E., McInnes, I.B., O’Dell, J.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1–2, pp. 20–33. [Google Scholar] [CrossRef]
- Smith, M.D. The Normal Synovium. Open Rheumatol. J. 2011, 5, 100–106. [Google Scholar] [CrossRef]
- Sellam, J.; Berenbaum, F. The Role of Synovitis in Pathophysiology and Clinical Symptoms of Osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- O’Grady, T.; Njock, M.-S.; Lion, M.; Bruyr, J.; Mariavelle, E.; Galvan, B.; Boeckx, A.; Struman, I.; Dequiedt, F. Sorting and Packaging of RNA into Extracellular Vesicles Shape Intracellular Transcript Levels. BMC Biol. 2022, 20, 72. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange Between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Kato, T.; Miyaki, S.; Ishitobi, H.; Nakamura, Y.; Nakasa, T.; Lotz, M.K.; Ochi, M. Exosomes from IL-1β Stimulated Synovial Fibroblasts Induce Osteoarthritic Changes in Articular Chondrocytes. Arthritis Res. Ther. 2014, 16, r163. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuang, Y.; Fang, L.; Yuan, C.; Wang, X.; Lin, K. Breakthrough of Extracellular Vesicles in Pathogenesis, Diagnosis and Treatment of Osteoarthritis. Bioact. Mater. 2022, 22, 423–452. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Yan, Y.; Li, R.; Dai, H.; Xu, J. Extracellular Vesicles from M1-Polarized Macrophages Promote Inflammation in the Temporomandibular Joint via MiR-1246 Activation of the Wnt/β-Catenin Pathway. Ann. N. Y. Acad. Sci. 2021, 1503, 48–59. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, H.; Zhao, F.; Liu, M.; Wang, F.; Kang, M.; He, W.; Lv, Z. Exosomal Circ-BRWD1 Contributes to Osteoarthritis Development through the Modulation of MiR-1277/TRAF6 Axis. Arthritis Res. Ther. 2021, 23, 159. [Google Scholar] [CrossRef]
- Wu, X.; Crawford, R.; Xiao, Y.; Mao, X.; Prasadam, I. Osteoarthritic Subchondral Bone Release Exosomes That Promote Cartilage Degeneration. Cells 2021, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, J.; Guo, B.; Liang, C.; Dang, L.; Lu, C.; He, X.; Cheung, H.Y.-S.; Xu, L.; Lu, C.; et al. Osteoclast-Derived Exosomal MiR-214-3p Inhibits Osteoblastic Bone Formation. Nat. Commun. 2016, 7, 10872. [Google Scholar] [CrossRef] [PubMed]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E.; Fuller, C.J.; Hurtig, M.; Cruz, A. The OARSI Histopathology Initiative—Recommendations for Histological Assessments of Osteoarthritis in the Horse. Osteoarthr. Cartil. 2010, 18, S93–S105. [Google Scholar] [CrossRef]
- Hu, C.; Wang, J. Chapter Five—Method for Enzyme Design with Genetically Encoded Unnatural Amino Acids. In Methods Enzymology; Pecoraro, V.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 580, pp. 109–133. [Google Scholar] [CrossRef]
- Probert, C.; Dottorini, T.; Speakman, A.; Hunt, S.; Nafee, T.; Fazeli, A.; Wood, S.; Brown, J.E.; James, V. Communication of Prostate Cancer Cells with Bone Cells via Extracellular Vesicle RNA; a Potential Mechanism of Metastasis. Oncogene 2018, 38, 1751–1763. [Google Scholar] [CrossRef]
- ThermoFisher Scientific. Click-IT® Nascent RNA Capture Kit Product Information. 2013. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/mp10365.pdf (accessed on 15 February 2025).
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Tabak, S.; Schreiber-Avissar, S.; Beit-Yannai, E. Extracellular Vesicles Have Variable Dose-Dependent Effects on Cultured Draining Cells in the Eye. J. Cell Mol. Med. 2018, 22, 1992–2000. [Google Scholar] [CrossRef]
- Goldring, M.B.; Culley, K.L.; Wondimu, E.; Otero, M. Chapter 3—Cartilage and Chondrocytes. In Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Firestein, G.S., Budd, R.C., Gabriel, S.E., McInnes, I.B., O’Dell, J.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, pp. 34–59.e3. [Google Scholar] [CrossRef]
- Wu, X.; Fan, X.; Crawford, R.; Xiao, Y.; Prasadam, I. The Metabolic Landscape in Osteoarthritis. Aging Dis. 2022, 13, 1166–1182. [Google Scholar] [CrossRef]
- Adair, T.H.; Gay, W.J.; Montani, J.P. Growth Regulation of the Vascular System: Evidence for a Metabolic Hypothesis. Am. J. Physiol. 1990, 259, R393–R404. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Kalbfleisch, T.S.; Rice, E.S.; DePriest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; O′Connell, B.L.; Fiddes, I.T.; Vershinina, A.O.; Saremi, N.F.; et al. Improved Reference Genome for the Domestic Horse Increases Assembly Contiguity and Composition. Comm. Biol. 2018, 1, 197. [Google Scholar] [CrossRef]
- Brennecke, P.; Anders, S.; Kim, J.K.; Kołodziejczyk, A.A.; Zhang, X.; Proserpio, V.; Baying, B.; Benes, V.; Teichmann, S.A.; Marioni, J.C.; et al. Accounting for Technical Noise in Single-Cell RNA-Seq Experiments. Nat. Methods 2013, 10, 1093–1095. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, P.S.; Gilbert, S.J.; Tarn, J.; Charlton, S.; Skelton, A.J.; Barter, M.J.; Duance, V.C.; Young, D.A.; Blain, E.J. Regulation of MicroRNA-221, -222, -21 and -27 in Articular Cartilage Subjected to Abnormal Compressive Forces. J. Physiol. 2021, 599, 143–155. [Google Scholar] [CrossRef]
- Dunn, W.; DuRaine, G.; Reddi, A.H. Profiling MicroRNA Expression in Bovine Articular Cartilage and Implications for Mechano transduction. Arthritis Rheum. 2009, 60, 2333–2339. [Google Scholar] [CrossRef]
- Shang, X.; Böker, K.O.; Taheri, S.; Lehmann, W.; Schilling, A.F. Extracellular Vesicles Allow Epigenetic Mechanotransduction between Chondrocytes and Osteoblasts. Int. J. Mol. Sci. 2021, 22, 13282. [Google Scholar] [CrossRef]
- Song, J.; Jin, E.H.; Kim, D.; Kim, K.Y.; Chun, C.H.; Jin, E.J. MicroRNA-222 Regulates MMP-13 via Targeting HDAC-4 during Osteoarthritis Pathogenesis. BBA Clin. 2015, 3, 79–89. [Google Scholar] [CrossRef]
- Zheng, X.; Zhao, F.-C.; Pang, Y.; Li, D.-Y.; Yao, S.-C.; Sun, S.-S.; Guo, K.-J. Downregulation of MiR-221-3p Contributes to IL-1β-Induced Cartilage Degradation by Directly Targeting the SDF1/CXCR4 Signaling Pathway. J. Mol. Med. 2017, 95, 615–627. [Google Scholar] [CrossRef]
- Anderson, J.R.; Jacobsen, S.; Walters, M.; Bundgaard, L.; Diendorfer, A.; Hackl, M.; Clarke, E.J.; James, V.; Peffers, M.J. Small Non-Coding RNA Landscape of Extracellular Vesicles from a Post-Traumatic Model of Equine Osteoarthritis. Front. Vet. Sci. 2022, 9, 901269. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.-N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. MiR-100-5p-Abundant Exosomes Derived from Infrapatellar Fat Pad MSCs Protect Articular Cartilage and Ameliorate Gait Abnormalities via Inhibition of MTOR in Osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Pan, Y.; Ma, J.; Miao, X.; Qi, X.; Zhou, H.; Jia, L. MiR-26a and MiR-26b Mediate Osteoarthritis Progression by Targeting FUT4 via NF-ΚB Signaling Pathway. Int. J. Biochem. Cell Biol. 2018, 94, 79–88. [Google Scholar] [CrossRef]
- Walsh, D.A.; Pearson, C.I. Angiogenesis in Pathogenesis of Inflammatory Joint and Lung Diseases. Arthritis Res. 2001, 3, 147–153. [Google Scholar] [CrossRef]
- Mapp, P.I.; Walsh, D.A. Mechanisms and Targets of Angiogenesis and Nerve Growth in Osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 390–398. [Google Scholar] [CrossRef]
- Hashimoto, S.; Creighton-Achermann, L.; Takahashi, K.; Amiel, D.; Coutts, R.D.; Lotz, M. Development and Regulation of Osteophyte Formation during Experimental Osteoarthritis. Osteoarthr. Cartil. 2002, 10, 180–187. [Google Scholar] [CrossRef]
- Bonnet, C.S.; Walsh, D.A. Osteoarthritis, Angiogenesis and Inflammation. Rheumatology 2005, 44, 7–16. [Google Scholar] [CrossRef]
- Ogata, N.; Chikazu, D.; Kubota, N.; Terauchi, Y.; Tobe, K.; Azuma, Y.; Ohta, T.; Kadowaki, T.; Nakamura, K.; Kawaguchi, H. Insulin Receptor Substrate-1 in Osteoblast Is Indispensable for Maintaining Bone Turnover. J. Clin. Investig. 2000, 105, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.A.; Campbell, I.K.; O’Donnell, K.L.; Wicks, I.P. Vascular Cell Adhesion Molecule-1 (VCAM-1) Blockade in Collagen-Induced Arthritis Reduces Joint Involvement and Alters B Cell Trafficking. Clin. Exp. Immunol. 2002, 128, 44–51. [Google Scholar] [CrossRef]
- Nagao, M.; Hamilton, J.L.; Kc, R.; Berendsen, A.D.; Duan, X.; Cheong, C.W.; Li, X.; Im, H.J.; Olsen, B.R. Vascular Endothelial Growth Factor in Cartilage Development and Osteoarthritis. Sci. Rep. 2017, 7, 13027. [Google Scholar] [CrossRef]
- Gaballah, A.; Hussein, N.A.; Risk, M.; Elsawy, N.; Elabasiry, S. Correlation between Synovial Vascular Endothelial Growth Factor, Clinical, Functional and Radiological Manifestations in Knee Osteoarthritis. Egypt. Rheumatol. 2016, 38, 29–34. [Google Scholar] [CrossRef]
- Bertoni, L.; Jacquet-Guibon, S.; Branly, T.; Legendre, F.; Desancé, M.; Mespoulhes, C.; Melin, M.; Hartmann, D.-J.; Schmutz, A.; Denoix, J.-M.; et al. An Experimentally Induced Osteoarthritis Model in Horses Performed on Both Metacarpophalangeal and Metatarsophalangeal Joints: Technical, Clinical, Imaging, Biochemical, Macroscopic and Microscopic Characterization. PLoS ONE 2020, 15, e0235251. [Google Scholar] [CrossRef]
- Jasiński, T.; Turek, B.; Kaczorowski, M.; Brehm, W.; Skierbiszewska, K.; Bonecka, J.; Domino, M. Equine Models of Temporomandibular Joint Osteoarthritis: A Review of Feasibility, Biomarkers, and Molecular Signaling. Biomedicines 2024, 12, 542. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.J.; Hollinshead, F.; Goodrich, L.R. Extracellular Vesicles in The Treatment and Prevention of Osteoarthritis: Can Horses Help Us Translate This Therapy to Humans? Extracell. Vesicles Circ. Nucl. Acids 2023, 4, 151–169. [Google Scholar] [CrossRef]
- Anderson, J.R.; Johnson, E.; Jenkins, R.; Jacobsen, S.; Green, D.; Walter, M.; Bundgaard, L.; Hausmans, B.A.C.; van der Akker, G.; Welting, T.J.M.; et al. Multi-Omic Temporal Landscape of Plasma and Synovial Fluid-Derived Extracellular Vesicles Using an Experimental Model of Equine Osteoarthritis. Int. J. Mol. Sci. 2023, 24, 14888. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Ughetto, S.; Mahjoum, S.; Nair, A.V.; Breakefield, X.O. Uptake, Functionality, and Re-Release of Extracellular Vesicle-Encapsulated Cargo. Cell Rep. 2022, 39, 110651. [Google Scholar] [CrossRef]
- Peffers, M.J.; Milner, P.I.; Tew, S.R.; Clegg, P.D. Regulation of SOX9 in Normal and Osteoarthritic Equine Articular Chondrocytes by Hyperosmotic Loading. Osteoarthr. Cartil. 2010, 18, 1502–1508. [Google Scholar] [CrossRef]
- Morgan, R.E.; Clegg, P.D.; Hunt, J.A.; Innes, J.F.; Tew, S.R. Interaction with Macrophages Attenuates Equine Fibroblast-like Synoviocyte ADAMTS5 (Aggrecanase-2) Gene Expression Following Inflammatory Stimulation. J. Orthop. Res. 2018, 36, 2178–2185. [Google Scholar] [CrossRef]
- Diendorfer, A.; Khamina, K.; Pultar, M.; Hackl, M. MiND (MiRNA NGS Discovery Pipeline): A Small RNA-Seq Analysis Pipeline and Report Generator for MicroRNA Biomarker Discovery Studies. F1000Research 2022, 11, 233. [Google Scholar] [CrossRef]
- Khamina, K.; Diendorfer, A.B.; Skalicky, S.; Weigl, M.; Pultar, M.; Krammer, T.L.; Fournier, C.A.; Schofield, A.L.; Otto, C.; Smith, A.T.; et al. A MicroRNA Next-Generation-Sequencing Discovery Assay (MiND) for Genome-Scale Analysis and Absolute Quantitation of Circulating MicroRNA Biomarkers. Int. J. Mol. Sci. 2022, 23, 1226. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 26 January 2025).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Friedländer, M.R.; MacKowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. MiRDeep2 Accurately Identifies Known and Hundreds of Novel MicroRNA Genes in Seven Animal Clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S. The MicroRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef]
- The RNAcentral Consortium. RNAcentral: A Hub of Information for Non-Coding RNA Sequences. Nucleic Acids Res. 2019, 47, D221–D229. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | EV-Donor Cells | EV-Recipient Cells | ||
|---|---|---|---|---|
| Chondrocytes (N = 8) | Synoviocytes (N = 9) | Chondrocytes (N = 5) | Synoviocytes (N = 4) | |
| Age, years | ||||
| Mean (SD) | 6.1 (2.6) | 5.8 (2.3) | 6.2 (4.4) | 5.5 (2.1) |
| 95% CI | 4.0–8.3 | 4.0–7.5 | 0.7–11.7 | 2.2–8.8 |
| Min; Max | 3; 10 | 3; 10 | 3; 14 | 3; 8 |
| p-value | 0.899 1 | 0.810 1 | ||
| Sex, n (%) | ||||
| Female | 3 (60.0) 2 | 5 (62.5) 3 | 0 (0.0) 4 | 3 (75.0) |
| Male | 2 (40.0) 2 | 3 (37.5) 3 | 1 (100.0) 4 | 1 (25.0) |
| p-value | 0.928 5 | – | ||
| Joint macroscopic score (0–9) 6, n (%) | ||||
| Scores | ||||
| 0 | 3 (37.5) | 2 (22.2) | 1 (20.0) | 0 (0.0) |
| 1 | 4 (50.0) | 1 (11.1) | 0 (0.0) | 1 (25.0) |
| 2 | 1 (12.5) | 5 (55.6) | 1 (20.0) | 2 (50.0) |
| 3 | 0 (0.0) | 1 (11.1) | 3 (60.0) | 1 (25.0) |
| ≥4 | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Mean (SD) | 0.8 (0.7) | 1.6 (1.0) | 2.2 (1.3) | 2.0 (0.8) |
| 95% CI | 0.2–1.3 | 0.8–2.3 | 0.6–3.8 | 0.7–3.3 |
| Min; Max | 0; 2 | 0; 3 | 0; 3 | 1; 3 |
| p-value | 0.092 7 | 0.635 1 | ||
| EV Characteristics | Chondrocytes (N = 8) | Synoviocytes (N = 9) |
|---|---|---|
| EV concentration, particles/mL | ||
| Mean (SD) | 1.7 × 109 (1.0 × 109) | 8.2 × 108 (9.6 × 108) |
| 95% CI | 8.5 × 108–1.5 × 109 | 5.3 × 108–1.1 × 109 |
| Min; Max | 7.7 × 107; 3.0 × 109 | 7.9 × 107; 2.6 × 109 |
| p-value | 0.115 1 | |
| EV size, nm | ||
| Mean (SD) | 186.3 (92.8) | 190.6 (71.7) |
| 95% CI | 157.2–215.4 | 168.9–212.3 |
| Min; Max | 133.0; 274.8 | 136.3; 292.5 |
| p-value | 0.815 1 | |
| 5-EU-Labelled RNA | Experimental Samples | Control Samples | ||
|---|---|---|---|---|
| EV-Recipient Chondrocytes (N = 5) | EV-Recipient Synoviocytes (N = 4) | EV-Recipient Chondrocytes (N = 1) | EV-Recipient Synoviocytes (N = 1) | |
| RNA concentration, ng/µL | ||||
| Mean (SD) | 43.1 (5.6) | 62.8 (14.1) | 0.9 (0.0) | 1.8 (0.0) |
| 95% CI | 36.2–50.0 | 40.5–85.2 | – | – |
| Min; Max | 33.6; 48.4 | 54.9; 83.9 | – | – |
| p-value | 0.016 1 | – | ||
| miRNA | logFC | p-Value | FDR | Significance |
|---|---|---|---|---|
| eca-miR-27b | −2.6 | <0.001 | <0.001 | Increased in EV-recipient chondrocytes (RNA originated from EV-donor synoviocytes) |
| eca-miR-23b | −1.7 | 0.002 | 0.033 | Increased in EV-recipient chondrocytes (RNA originated from EV-donor synoviocytes) |
| eca-miR-143 | 2.6 | 0.002 | 0.033 | Increased in EV-recipient synoviocytes RNA originated from EV-donor chondrocytes) |
| eca-miR-31 | −4.9 | 0.003 | 0.033 | Increased in EV-recipient chondrocytes (RNA originated from EV-donor synoviocytes) |
| eca-miR-21 | 0.8 | 0.013 | 0.110 | Increased in EV-recipient synoviocytes (RNA originated from EV-donor chondrocytes) |
| eca-miR-181a | 2.2 | 0.015 | 0.110 | Increased in EV-recipient synoviocytes (RNA originated from EV-donor chondrocytes) |
| eca-miR-191a | −0.6 | 0.016 | 0.110 | Increased in EV-recipient chondrocytes (RNA originated from EV-donor synoviocytes) |
| eca-miR-181b | 1.8 | 0.018 | 0.110 | Increased in EV-recipient synoviocyte (RNA originated from EV-donor chondrocytes) |
| eca-miR-199a-5p | −0.6 | 0.027 | 0.145 | Increased in EV-recipient chondrocytes (RNA originated from EV-donor synoviocytes) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castanheira, C.I.G.D.; Anderson, J.R.; Clarke, E.J.; Hackl, M.; James, V.; Clegg, P.D.; Peffers, M.J. Extracellular Vesicle-Derived microRNA Crosstalk Between Equine Chondrocytes and Synoviocytes—An In Vitro Approach. Int. J. Mol. Sci. 2025, 26, 3353. https://doi.org/10.3390/ijms26073353
Castanheira CIGD, Anderson JR, Clarke EJ, Hackl M, James V, Clegg PD, Peffers MJ. Extracellular Vesicle-Derived microRNA Crosstalk Between Equine Chondrocytes and Synoviocytes—An In Vitro Approach. International Journal of Molecular Sciences. 2025; 26(7):3353. https://doi.org/10.3390/ijms26073353
Chicago/Turabian StyleCastanheira, Catarina I. G. D., James R. Anderson, Emily J. Clarke, Matthias Hackl, Victoria James, Peter D. Clegg, and Mandy J. Peffers. 2025. "Extracellular Vesicle-Derived microRNA Crosstalk Between Equine Chondrocytes and Synoviocytes—An In Vitro Approach" International Journal of Molecular Sciences 26, no. 7: 3353. https://doi.org/10.3390/ijms26073353
APA StyleCastanheira, C. I. G. D., Anderson, J. R., Clarke, E. J., Hackl, M., James, V., Clegg, P. D., & Peffers, M. J. (2025). Extracellular Vesicle-Derived microRNA Crosstalk Between Equine Chondrocytes and Synoviocytes—An In Vitro Approach. International Journal of Molecular Sciences, 26(7), 3353. https://doi.org/10.3390/ijms26073353