
Methoxylated Chrysin and Quercetin as Potent Stimulators of Melanogenesis

 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
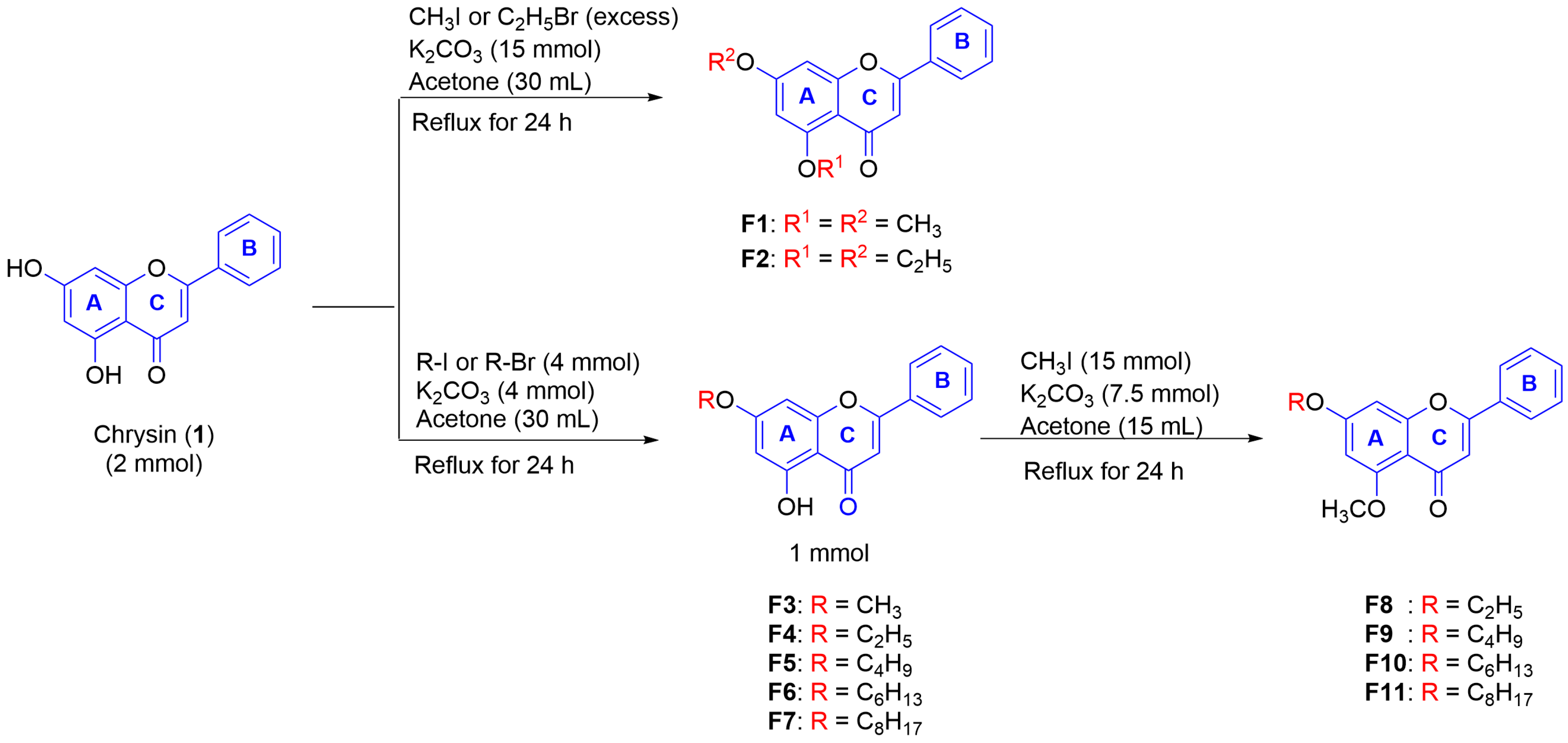
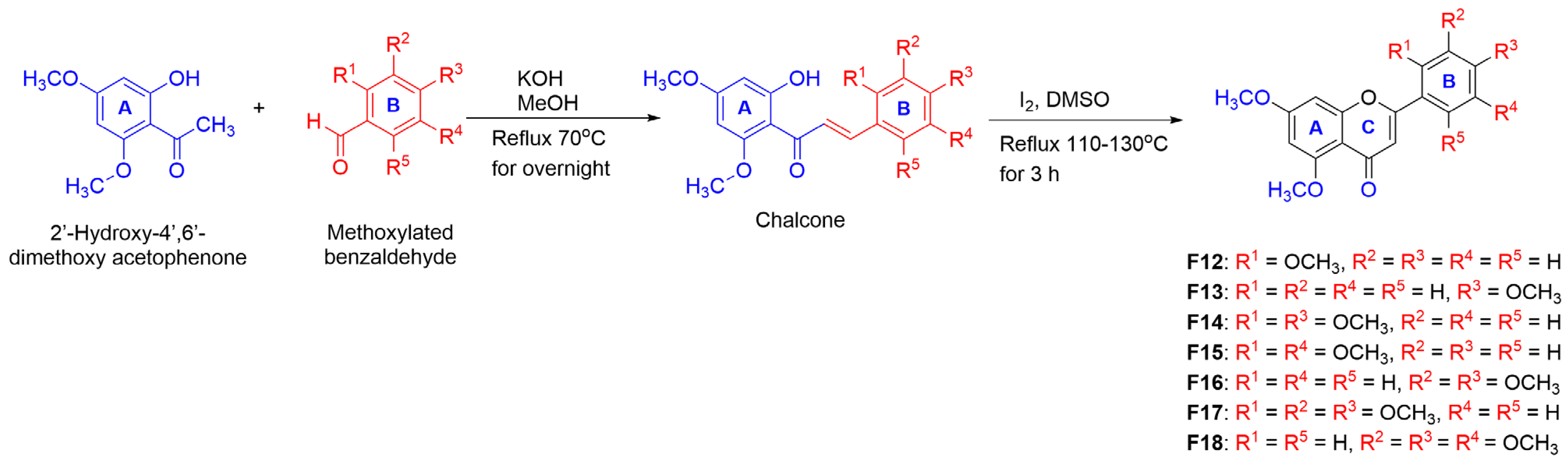
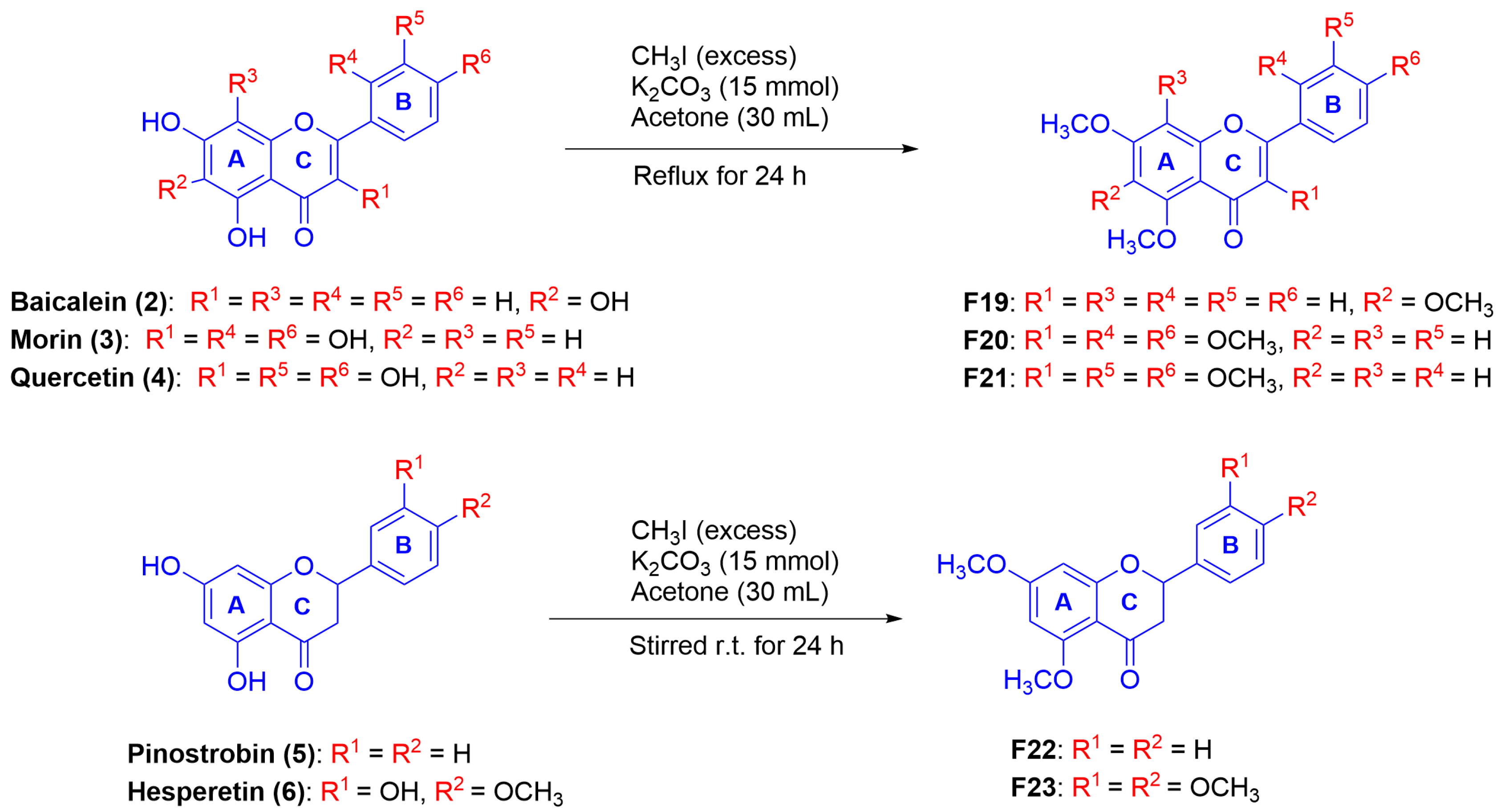
2.1. Synthesis of Mono-, Di-, and Poly-Methoxyflavonoid Derivatives
2.2. The Effects of PMFs on Melanin Production in Mouse B16F10 Melanoma Cells
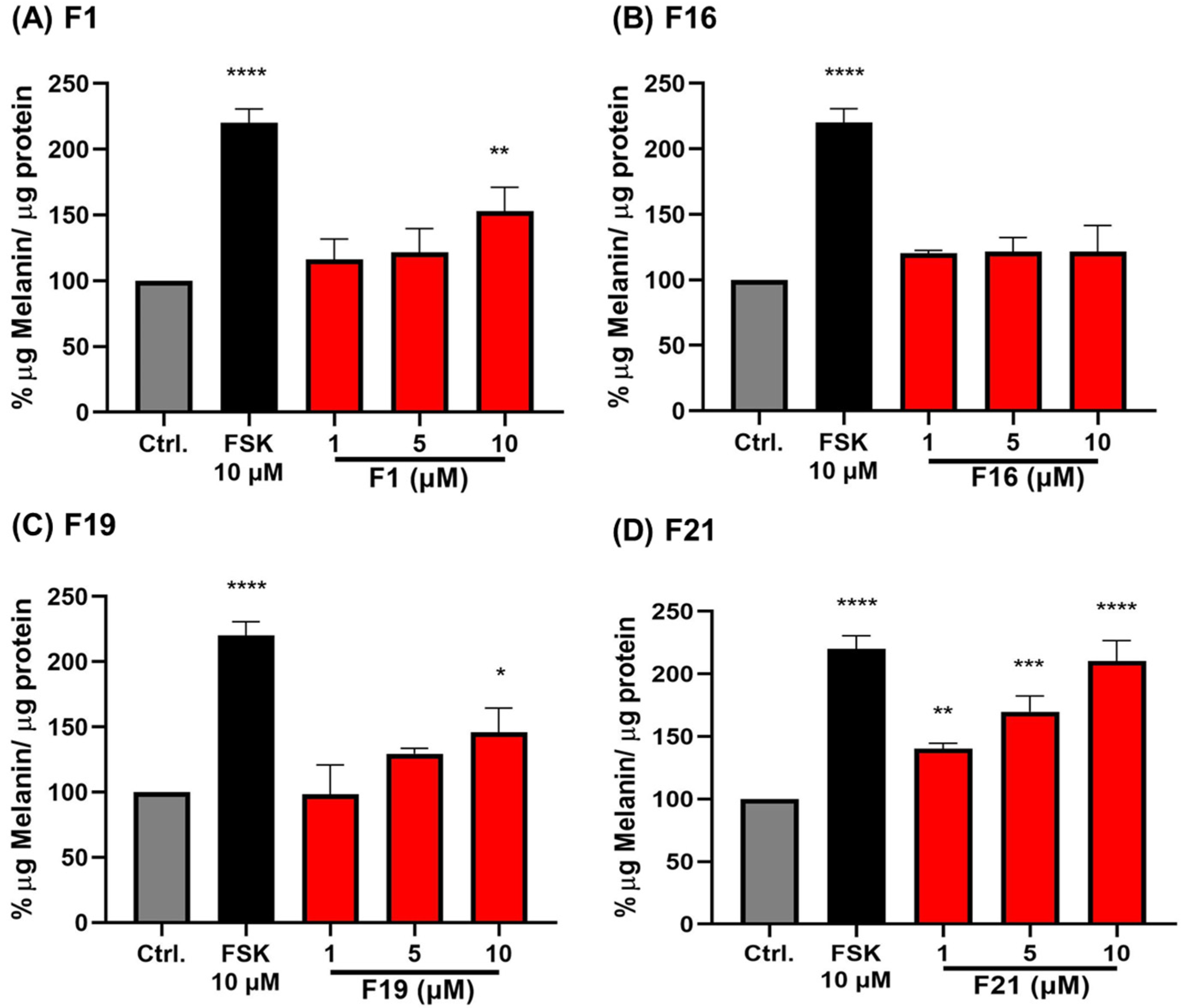
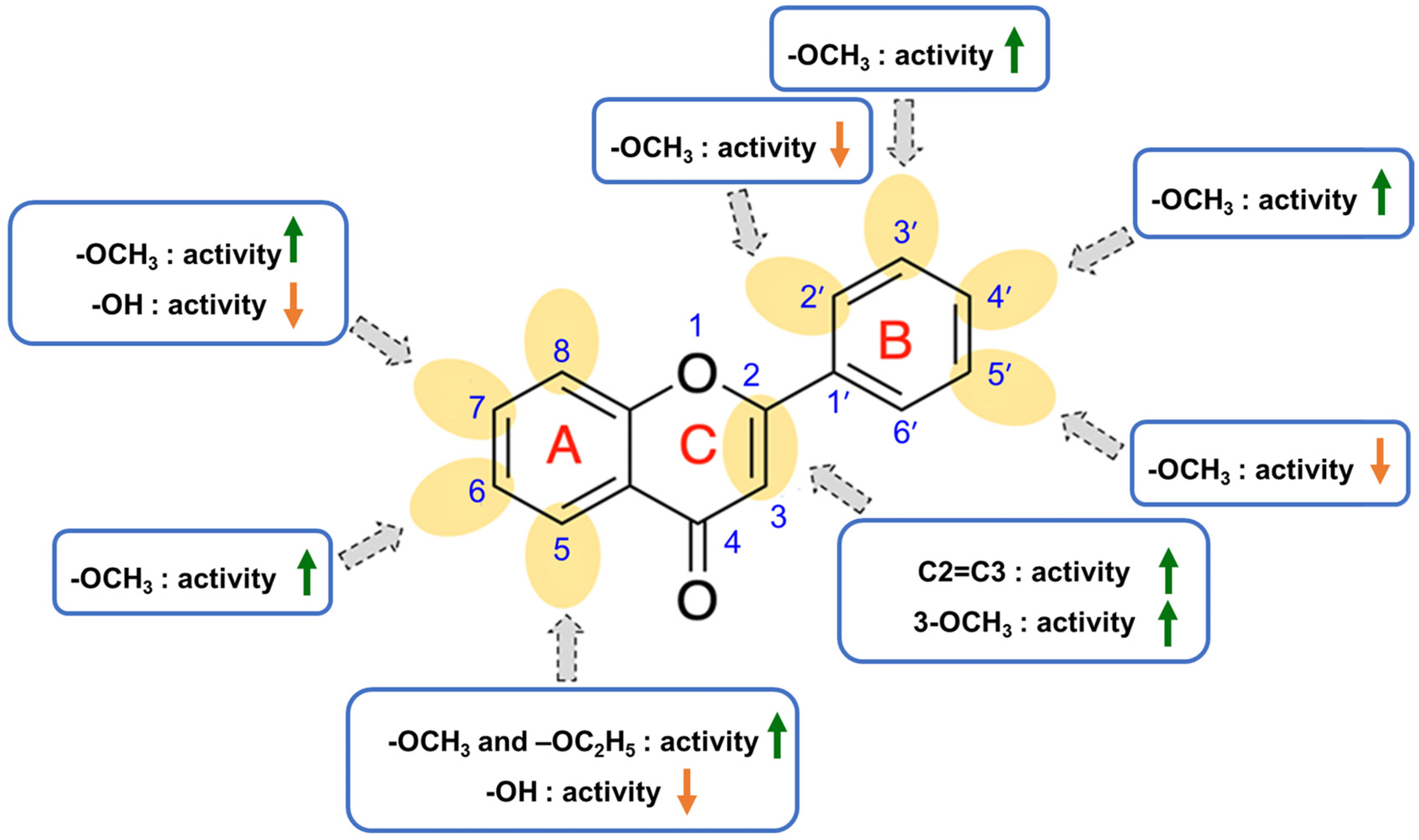
2.2.1. The Effect of Alkyl Chain Length at 7-Position of 5-OH and 5-OCH3 Chrysin (1)
2.2.2. The Effect of Methoxy Substituents on the B-Ring of 5,7-Dimethoxyflavone (F1)
2.2.3. The Effect of Fully-Methoxylated Flavonoids on B16F10 Melanin Content
2.3. The Effects of PMFs on Melanin Production in Human MNT-1 Cells
2.4. Structure-Activity Relationship of PMFs on Melanogenic Enhancing Activity
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Isolation of Natural Flavonoids
Isolation of Baicalein from Scutellaria baicalensis Roots
Isolation of Pinostrobin from Boesenbergia rotunda L. Rhizomes
3.2.2. General Procedure for the Synthesis of Flavonoid Derivatives
Synthesis of Chrysin Derivatives (F1–F15)
Synthesis of PMFs F12–F18
Synthesis of PMFs F19–F23
3.2.3. Melanogenesis Activity
Cell Culture
Cell Viability Assay
Measurement of Melanin Content
Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar]
- D’Mello, S.D.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jung, H.; Jang, B.; Song, H.K.; Han, I.O.; Oh, E.S. D-tyrosine adds an anti-melanogenic effect to cosmetic peptides. Sci. Rep. 2020, 10, 262. [Google Scholar]
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [PubMed]
- Ramsden, C.A.; Riley, P.A. Tyrosinase: The four oxidation states of the active site and their relevance to enzymatic activation, oxidation and inactivation. Bioorg Med. Chem. 2014, 22, 2388–2395. [Google Scholar]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar]
- Sturm, R.A. Molecular genetics of human pigmentation diversity. Hum. Mol. Genet. 2009, 18, R9–R17. [Google Scholar]
- Serre, C.; Busuttil, V.; Botto, J.M. Intrinsic and extrinsic regulation of human skin melanogenesis and pigmentation. Int. J. Cosmet. Sci. 2018, 40, 328–347. [Google Scholar]
- Yamauchi, K.; Mitsunaga, T.; Batubara, I. Synthesis of quercetin glycosides and their melanogenesis stimulatory activity in B16 melanoma cells. Bioorg Med. Chem. 2014, 22, 937–944. [Google Scholar]
- Liu, C.; Nueraihemaiti, M.; Zang, D.; Edirs, S.; Zou, G.; Aisa, H.A. Quercetin 3-O-(6″-O-E-caffeoyl)-β-D-glucopyranoside, a Flavonoid Compound, Promotes Melanogenesis through the Upregulation of MAPKs and Akt/GSK3β/β-Catenin Signaling Pathways. Int. J. Mol. Sci. 2023, 24, 4780. [Google Scholar] [CrossRef] [PubMed]
- Dian, A.; Lestari, D.; Renata, P. In Silico Study of Naringenin as Melanogenesis Inducer in Vitiligo. Pharmacogn. J. 2022, 14, 847–855. [Google Scholar]
- Niu, C.; Aisa, H.A. Upregulation of Melanogenesis and Tyrosinase Activity: Potential Agents for Vitiligo. Molecules 2017, 22, 1303. [Google Scholar] [CrossRef]
- Gan, R.; Liu, Y.; Li, H.; Xia, Y.; Guo, H.; Geng, F.; Zhuang, Q.; Li, H.; Wu, D. Natural sources, refined extraction, biosynthesis, metabolism, and bioactivities of dietary polymethoxyflavones (PMFs). Food Sci. Human Wellness 2024, 13, 27–49. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, S.; Ho, C.-T.; Huang, Q. Citrus polymethoxyflavones as regulators of metabolic homoeostasis: Recent advances for possible mechanisms. Trends Food Sci. Technol. 2021, 110, 743–753. [Google Scholar] [CrossRef]
- You, Q.; Li, D.; Ding, H.; Chen, H.; Hu, Y.; Liu, Y. Pharmacokinetics and Metabolites of 12 Bioactive Polymethoxyflavones in Rat Plasma. J. Agric. Food Chem. 2021, 69, 12705–12716. [Google Scholar] [CrossRef]
- Klinngam, W.; Rungkamoltip, P.; Thongin, S.; Joothamongkhon, J.; Khumkhrong, P.; Khongkow, M.; Namdee, K.; Tepaamorndech, S.; Chaikul, P.; Kanlayavattanakul, M.; et al. Polymethoxyflavones from Kaempferia parviflora ameliorate skin aging in primary human dermal fibroblasts and ex vivo human skin. Biomed. Pharmacother. 2022, 145, 112461. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Qu, L.Q.; Ng, J.P.L.; Zeng, W.; Yu, L.; Song, L.L.; Wong, V.K.W.; Xia, C.L.; Law, B.Y.K. Natural citrus flavanone 5-demethylnobiletin stimulates melanogenesis through the activation of cAMP/CREB pathway in B16F10 cells. Phytomedicine 2022, 98, 153941. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Lai, H.J.; Wu, A.G.; Tang, Y.; Song, L.L.; Lo, H.H.; Wong, I.N.; Wong, V.K.W.; Law, B.Y.K. Melanogenic effects of 5-demethylnobiletin on mouse model of chemical-induced vitiligo. J. Funct. Foods 2024, 112, 105962. [Google Scholar] [CrossRef]
- Na Takuathung, M.; Klinjan, P.; Koonrungsesomboon, N. A systematic review and meta-analysis of animal and human studies demonstrates the beneficial effects of Kaempferia parviflora on metabolic syndrome and erectile dysfunction. Nutr. Res. 2024, 122, 80–91. [Google Scholar] [CrossRef]
- Joothamongkhon, J.; Susantikarn, P.; Kongkachana, W.; Ketngamkum, Y.; Batthong, S.; Jomchai, N.; Yingyong, P.; Asawapirom, U.; Tangphatsornruang, S.; Paemanee, A.; et al. Quantitative analysis of methoxyflavones discriminates between the two types of Kaempferia parviflora. Phytochem. Anal. 2022, 33, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.G.; Choi, E.J.; Choi, Y.; Hwang, J.K. 5,7-Dimethoxyflavone induces melanogenesis in B16F10 melanoma cells through cAMP-dependent signalling. Exp. Dermatol. 2011, 20, 445–447. [Google Scholar] [PubMed]
- Hairani, R.; Chavasiri, W. A new series of chrysin derivatives as potent non-saccharide α-glucosidase inhibitors. Epub 2022, 19, 105301. [Google Scholar] [CrossRef]
- Rao, Y.K.; Harikishore, P.; Rao, C.V.; Gunasekar, D.; Blond, A.; Bodo, B. Flavones from Andrographis viscosula. Phytochemistry 2002, 61, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.; Tomás-Navarro, M.; Yuste, J.E.; Crupi, P.; Vallejo, F. An update on citrus polymethoxyflavones: Chemistry, metabolic fate, and relevant bioactivities. Eur. Food Res. Technol. 2024, 250, 2179–2192. [Google Scholar] [CrossRef]
- Łużny, M.; Tronina, T.; Kozłowska, E.; Kostrzewa-Susłow, E.; Janeczko, T. Biotransformation of 5,7-Methoxyflavones by Selected Entomopathogenic Filamentous Fungi. J. Agric. Food Chem. 2021, 69, 3879–3886. [Google Scholar] [CrossRef]
- Azuma, T.; Tanaka, Y.; Kikuzaki, H. Phenolic glycosides from Kaempferia parviflora. Phytochemistry 2008, 69, 2743–2748. [Google Scholar] [CrossRef]
- Jaipetch, T.; Reutrakul, V.; Tuntiwachwuttikul, P.; Santisuk, T. Flavonoids in the black rhizomes of Boesenbergia panduta. Phytochemistry 1983, 22, 625–626. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, Y.; Song, M.T.; Qian, L.L.; Liu, X.L.; Gao, R.Y.; Han, R.M.; Skibsted, L.H.; Zhang, J.P. Promotion effects of flavonoids on browning induced by enzymatic oxidation of tyrosinase: Structure-activity relationship. RSC Adv. 2021, 11, 13769–13779. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; El-Din, M.I.G.; Hritcu, L.; Eldahshan, O.A. Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021. Molecules 2021, 26, 7546. [Google Scholar] [CrossRef]
- Liu-Smith, F.; Meyskens, F.L. Molecular mechanisms of flavonoids in melanin synthesis and the potential for the prevention and treatment of melanoma. Mol. Nutr. Food Res. 2016, 60, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Wu, S.; He, Y.; Nian, Q.; Lei, J.; Yao, Y.; Guo, J.; Zeng, J. Plant-Derived Compounds as Promising Therapeutics for Vitiligo. Front. Pharmacol. 2021, 12, 685116. [Google Scholar] [CrossRef] [PubMed]
- Obaid, R.J.; Mughal, E.U.; Naeem, N.; Sadiq, A.; Alsantali, R.I.; Jassas, R.S.; Moussa, Z.; Ahmed, S.A. Natural and synthetic flavonoid derivatives as new potential tyrosinase inhibitors: A systematic review. RSC Adv. 2021, 11, 22159–22198. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Methylation of dietary flavones greatly improves their hepatic metabolic stability and intestinal absorption. Mol. Pharm. 2007, 4, 826–832. [Google Scholar] [CrossRef]
- Fan, M.; Ding, H.; Zhang, G.; Hu, X.; Gong, D. Relationships of dietary flavonoid structure with its tyrosinase inhibitory activity and affinity. LWT 2019, 107, 25–34. [Google Scholar] [CrossRef]
- Feng, D.; Fang, Z.; Zhang, P. The melanin inhibitory effect of plants and phytochemicals: A systematic review. Phytomedicine 2022, 107, 154449. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||
|---|---|---|---|
| Compound | R1 | R2 | % Melanin Content |
| Chrysin (1) | H | H | 141.00 ± 1.15 |
| F1 | CH3 | CH3 | 267.47 ± 1.30 **** |
| F2 | C2H5 | C2H5 | 151.66 ± 2.81 * |
| F3 | H | CH3 | 113.61 ± 1.69 |
| F4 | H | C2H5 | 138.44 ± 1.18 |
| F8 | CH3 | C2H5 | 163.39 ± 1.52 ** |
| F9 | CH3 | C4H9 | 179.15 ± 0.45 **** |
| F10 | CH3 | C6H13 | 160.19 ± 2.75 ** |
| F11 | CH3 | C8H17 | 88.77 ± 0.57 |
| Forskolin (Positive control) | 291.94 ± 0.82 **** | ||
| No compound added (Control) | 100 ± 00 | ||
 | ||||||
|---|---|---|---|---|---|---|
| Compounds | R1 | R2 | R3 | R4 | R5 | % Melanin Content |
| F1 | H | H | H | H | H | 267.47 ± 1.30 **** |
| F12 | OCH3 | H | H | H | H | 173.43 ± 2.14 *** |
| F13 | H | H | OCH3 | H | H | 196.33 ± 1.82 **** |
| F14 | OCH3 | H | OCH3 | H | H | 143.41 ± 1.68 |
| F15 | OCH3 | H | H | OCH3 | H | 114.25 ± 2.93 |
| F16 | H | OCH3 | OCH3 | H | H | 241.47 ± 3.37 **** |
| F17 | OCH3 | OCH3 | OCH3 | H | H | 193.74 ± 3.69 **** |
| F18 | H | OCH3 | OCH3 | OCH3 | H | 72.68 ± 0.97 |
| Forskolin (positive control) | 291.94 ± 0.82 **** | |||||
| No compound added (control) | 100 ± 00 | |||||
 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Compounds | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | R10 | % Melanin Content |
| F1 | H | OCH3 | H | OCH3 | H | H | H | H | H | H | 267.47 ± 1.30 **** |
| F13 | H | OCH3 | H | OCH3 | H | H | H | OCH3 | H | H | 196.33 ± 1.82 **** |
| F19 | H | OCH3 | OCH3 | OCH3 | H | H | H | H | H | H | 211.22 ± 1.23 **** |
| F20 | OCH3 | OCH3 | H | OCH3 | H | OCH3 | H | OCH3 | H | H | 201.58 ± 1.14 **** |
| F21 | OCH3 | OCH3 | H | OCH3 | H | H | OCH3 | OCH3 | H | H | 289.71 ± 1.86 **** |
| F22 | 2H | OCH3 | H | OCH3 | H | H | H | H | H | H | 142.34 ± 1.85 |
| F23 | 2H | OCH3 | H | OCH3 | H | H | OCH3 | OCH3 | H | H | 112.01 ± 1.17 |
| Forskolin | 291.94 ± 0.82 **** | ||||||||||
| Control | 100 ± 00 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poungcho, P.; Hairani, R.; Chaotham, C.; De-Eknamkul, W.; Chavasiri, W. Methoxylated Chrysin and Quercetin as Potent Stimulators of Melanogenesis. Int. J. Mol. Sci. 2025, 26, 3281. https://doi.org/10.3390/ijms26073281
Poungcho P, Hairani R, Chaotham C, De-Eknamkul W, Chavasiri W. Methoxylated Chrysin and Quercetin as Potent Stimulators of Melanogenesis. International Journal of Molecular Sciences. 2025; 26(7):3281. https://doi.org/10.3390/ijms26073281
Chicago/Turabian StylePoungcho, Pattara, Rita Hairani, Chatchai Chaotham, Wanchai De-Eknamkul, and Warinthorn Chavasiri. 2025. "Methoxylated Chrysin and Quercetin as Potent Stimulators of Melanogenesis" International Journal of Molecular Sciences 26, no. 7: 3281. https://doi.org/10.3390/ijms26073281
APA StylePoungcho, P., Hairani, R., Chaotham, C., De-Eknamkul, W., & Chavasiri, W. (2025). Methoxylated Chrysin and Quercetin as Potent Stimulators of Melanogenesis. International Journal of Molecular Sciences, 26(7), 3281. https://doi.org/10.3390/ijms26073281