

The Application of Non-Coding RNAs as Biomarkers, Therapies, and Novel Vaccines in Diseases
 , ,
, ,  ,
, 
Abstract
1. Introduction

{kind=link}
{kind=link}
| Types of RNA | Features and Introduction | References |
|---|---|---|
| tRNAs | These possess a cloverleaf structure, deliver the correct amino acids to the ribosome, and facilitate the formation of polypeptide chains | [8,9] |
| rRNAs | The structural components of the ribosome | [10] |
| snRNAs | A class of small RNA molecules in the nucleus of eukaryotic cells that are capable of processing pre-mRNAs | [11] |
| snoRNAs | Regulatory factors promoting rRNA maturation | [12] |
| TERCs | The RNA component of telomerase, which provides a structural scaffold for the assembly of the telomerase complex | [13,14] |
| miRNAs | With a length of approximately 19~24 nucleotides (nts), these serve as post-transcriptional regulatory factors of gene expression | [15] |
| circRNAs | These lack free ends and have covalently closed loop structures, and can function as miRNA sponges, RNA-binding protein sponges, and translation regulatory factors | [16,17] |
| lncRNAs | With a length exceeding 200 nts, these mediate the post-transcriptional control of signal transduction pathways, translation processes, and gene expression | [18] |
| siRNAs | These are primarily involved in RNA interference to regulate gene expression | [19] |
| piRNAs | These have diverse functions including gene regulation, transposon suppression, epigenetic programming, and antiviral defense, among others | [20,21] |
2. NcRNAs as Diagnostic Biomarkers
2.1. MiRNAs as Biomarkers
2.2. CircRNAs as Biomarkers
2.3. LncRNAs as Biomarkers
3. NcRNAs in Target Therapy
4. NcRNAs in Novel Vaccines
5. Discussions: Challenges and Opportunities
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, W.; Chen, R.; Droll, S.; Barber, E.; Saleh, L.; Corrigan-Cummins, M.; Trick, M.; Anastas, V.; Hawk, N.V.; Zhao, Z.; et al. MiR-181c regulates MCL1 and cell survival in GATA2 deficient cells. J. Leukoc. Biol. 2022, 111, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Liu, G. NcRNA-regulated immune response and its role in inflammatory lung diseases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L1076–L1087. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Bu, P. Non-coding RNA in cancer. Essays Biochem. 2021, 65, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Das, T.K.; Khodarkovskaya, A.; Dash, S. Non-coding RNAs and their bioengineering applications for neurological diseases. Bioengineered 2021, 12, 11675–11698. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Biela, A.; Hammermeister, A.; Kaczmarczyk, I.; Walczak, M.; Koziej, L.; Lin, T.Y.; Glatt, S. The diverse structural modes of tRNA binding and recognition. J. Biol. Chem. 2023, 299, 104966. [Google Scholar] [CrossRef]
- Mathew, B.A.; Katta, M.; Ludhiadch, A.; Singh, P.; Munshi, A. Role of tRNA-Derived Fragments in Neurological Disorders: A Review. Mol. Neurobiol. 2023, 60, 655–671. [Google Scholar] [CrossRef]
- Denisenko, O. Epigenetics of Ribosomal RNA Genes. Biochemistry 2022, 87, S103–S131. [Google Scholar] [CrossRef]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [PubMed]
- van der Werf, J.; Chin, C.V.; Fleming, N.I. SnoRNA in Cancer Progression, Metastasis and Immunotherapy Response. Biology 2021, 10, 809. [Google Scholar] [CrossRef]
- Rubtsova, M.; Dontsova, O. Human Telomerase RNA: Telomerase Component or More? Biomolecules 2020, 10, 873. [Google Scholar] [CrossRef]
- Nguyen, T.H.D.; Tam, J.; Wu, R.A.; Greber, B.J.; Toso, D.; Nogales, E.; Collins, K. Cryo-EM structure of substrate-bound human telomerase holoenzyme. Nature 2018, 557, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, biogenesis, and their evolving role in animal development and disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, present, and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Alshaer, W.; Zureigat, H.; Al Karaki, A.; Al-Kadash, A.; Gharaibeh, L.; Hatmal, M.M.; Aljabali, A.A.A.; Awidi, A. SiRNA: Mechanism of action, challenges, and therapeutic approaches. Eur. J. Pharmacol. 2021, 905, 174178. [Google Scholar] [CrossRef]
- Mani, S.R.; Juliano, C.E. Untangling the web: The diverse functions of the PIWI/piRNA pathway. Mol. Reprod. Dev. 2013, 80, 632–664. [Google Scholar] [CrossRef]
- Ku, H.Y.; Lin, H. PIWI proteins and their interactors in piRNA biogenesis, germline development and gene expression. Natl. Sci. Rev. 2014, 1, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, J.; Yu, S.; Wang, Z.; He, X.; Su, Y.; Guo, T.; Sheng, H.; Chen, J.; Zheng, Q.; et al. Extracellular Vesicles Long RNA Sequencing Reveals Abundant mRNA, circRNA, and lncRNA in Human Blood as Potential Biomarkers for Cancer Diagnosis. Clin. Chem. 2019, 65, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Arantes, L.; De Carvalho, A.C.; Melendez, M.E.; Lopes Carvalho, A. Serum, plasma and saliva biomarkers for head and neck cancer. Expert Rev. Mol. Diagn. 2018, 18, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Tian, Z.; Fan, W.; Ni, B. Circular RNA: A novel biomarker and therapeutic target for human cancers. Int. J. Med. Sci. 2019, 16, 292–301. [Google Scholar] [CrossRef]
- Jin, Z.Q. MicroRNA targets and biomarker validation for diabetes-associated cardiac fibrosis. Pharmacol. Res. 2021, 174, 105941. [Google Scholar] [CrossRef]
- Felekkis, K.; Touvana, E.; Stefanou, C.; Deltas, C. MicroRNAs: A newly described class of encoded molecules that play a role in health and disease. Hippokratia 2010, 14, 236–240. [Google Scholar] [PubMed]
- Martin, E.C.; Qureshi, A.T.; Dasa, V.; Freitas, M.A.; Gimble, J.M.; Davis, T.A. MicroRNA regulation of stem cell differentiation and diseases of the bone and adipose tissue: Perspectives on miRNA biogenesis and cellular transcriptome. Biochimie 2016, 124, 98–111. [Google Scholar] [CrossRef]
- Ding, S.; Liu, G.; Jiang, H.; Fang, J. MicroRNA Determines the Fate of Intestinal Epithelial Cell Differentiation and Regulates Intestinal Diseases. Curr. Protein Pept. Sci. 2019, 20, 666–673. [Google Scholar] [CrossRef]
- He, J.F.; Luo, Y.M.; Wan, X.H.; Jiang, D. Biogenesis of MiRNA-195 and its role in biogenesis, the cell cycle, and apoptosis. J. Biochem. Mol. Toxicol. 2011, 25, 404–408. [Google Scholar] [CrossRef]
- Jing, R.; Zhong, Q.Q.; Long, T.Y.; Pan, W.; Qian, Z.X. Downregulated miRNA-26a-5p induces the apoptosis of endothelial cells in coronary heart disease by inhibiting PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4940–4947. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Agbu, P.; Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef]
- Qadir, M.I.; Bukhat, S.; Rasul, S.; Manzoor, H.; Manzoor, M. RNA therapeutics: Identification of novel targets leading to drug discovery. J. Cell. Biochem. 2020, 121, 898–929. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Ding, M.; Zhang, H.; Xu, X.; Tang, J. Molecular mechanisms and clinical applications of miR-22 in regulating malignant progression in human cancer (Review). Int. J. Oncol. 2017, 50, 345–355. [Google Scholar] [CrossRef]
- Paliwal, N.; Vashist, M.; Chauhan, M. Evaluation of miR-22 and miR-21 as diagnostic biomarkers in patients with epithelial ovarian cancer. 3 Biotech 2020, 10, 142. [Google Scholar] [CrossRef]
- Duan, Z.Y.; Bu, R.; Liang, S.; Chen, X.Z.; Zhang, C.; Zhang, Q.Y.; Li, J.J.; Chen, X.M.; Cai, G.Y. Urinary miR-185-5p is a biomarker of renal tubulointerstitial fibrosis in IgA nephropathy. Front. Immunol. 2024, 15, 1326026. [Google Scholar] [CrossRef]
- Roberts, I.S. Pathology of IgA nephropathy. Nat. Rev. Nephrol. 2014, 10, 445–454. [Google Scholar] [CrossRef]
- Koch, P.F.; Ludwig, K.; Krenzien, F.; Hillebrandt, K.H.; Schöning, W.; Pratschke, J.; Raschzok, N.; Sauer, I.M.; Moosburner, S. miRNA as potential biomarkers after liver transplantation: A systematic review. Transplant. Rev. 2024, 38, 100831. [Google Scholar] [CrossRef]
- Millán, O.; Ruiz, P.; Orts, L.; Ferré, P.; Crespo, G.; Santana, M.; Fortuna, V.; Quintairos, L.; Navasa, M.; Brunet, M. Monitoring of miR-181a-5p and miR-155-5p Plasmatic Expression as Prognostic Biomarkers for Acute and Subclinical Rejection in De Novo Adult Liver Transplant Recipients. Front. Immunol. 2019, 10, 873. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef]
- Panda, A.C. Circular RNAs Act as miRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Lu, D.; Xu, A. The interaction of circRNAs and RNA binding proteins: An important part of circRNA maintenance and function. J. Neurosci. Res. 2020, 98, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Bartoli, E.; Fra, G.P.; Carnevale Schianca, G.P. The oral glucose tolerance test (OGTT) revisited. Eur. J. Intern. Med. 2011, 22, 8–12. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, X.; Jian, D.; Hao, P.; Rao, L.; Li, M. Hsa_circ_0054633 in peripheral blood can be used as a diagnostic biomarker of pre-diabetes and type 2 diabetes mellitus. Acta Diabetol. 2017, 54, 237–245. [Google Scholar] [CrossRef]
- Kolbeinsson, H.M.; Chandana, S.; Wright, G.P.; Chung, M. Pancreatic Cancer: A Review of Current Treatment and Novel Therapies. J. Investig. Surg. 2023, 36, 2129884. [Google Scholar] [CrossRef]
- Singhi, A.D.; Koay, E.J.; Chari, S.T.; Maitra, A. Early Detection of Pancreatic Cancer: Opportunities and Challenges. Gastroenterology 2019, 156, 2024–2040. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Jun, E.; Okugawa, Y.; Toiyama, Y.; Borazanci, E.; Bolton, J.; Taketomi, A.; Kim, S.C.; Shang, D.; Von Hoff, D.; et al. A Circulating Panel of circRNA Biomarkers for the Noninvasive and Early Detection of Pancreatic Ductal Adenocarcinoma. Gastroenterology 2024, 166, 178–190.e116. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, B.; Chang, X. Emerging roles of circular RNAs in osteoporosis. J. Cell. Mol. Med. 2021, 25, 9089–9101. [Google Scholar] [CrossRef]
- Zhi, F.; Ding, Y.; Wang, R.; Yang, Y.; Luo, K.; Hua, F. Exosomal hsa_circ_0006859 is a potential biomarker for postmenopausal osteoporosis and enhances adipogenic versus osteogenic differentiation in human bone marrow mesenchymal stem cells by sponging miR-431-5p. Stem Cell Res. Ther. 2021, 12, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.K. Emerging roles of RNA processing factors in regulating long non-coding RNAs. Rna Biol. 2014, 11, 793–797. [Google Scholar] [CrossRef]
- Kanduri, C. Long noncoding RNAs: Lessons from genomic imprinting. Biochim. Biophys. Acta-Gene Regul. Mech. 2016, 1859, 102–111. [Google Scholar] [CrossRef]
- Nainar, S.; Feng, C.; Spitale, R.C. Chemical Tools for Dissecting the Role of lncRNAs in Epigenetic Regulation. ACS Chem. Biol. 2016, 11, 2091–2100. [Google Scholar] [CrossRef]
- González-Moro, I.; Garcia-Etxebarria, K.; Mendoza, L.M.; Fernández-Jiménez, N.; Mentxaka, J.; Olazagoitia-Garmendia, A.; Arroyo, M.N.; Sawatani, T.; Moreno-Castro, C.; Vinci, C.; et al. LncRNA ARGI Contributes to Virus-Induced Pancreatic β Cell Inflammation Through Transcriptional Activation of IFN-Stimulated Genes. Adv. Sci. 2023, 10, e2300063. [Google Scholar] [CrossRef]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genom. Proteom. Bioinform. 2016, 14, 42–54. [Google Scholar] [CrossRef]
- Su, M.; Wang, H.; Wang, W.; Wang, Y.; Ouyang, L.; Pan, C.; Xia, L.; Cao, D.; Liao, Q. LncRNAs in DNA damage response and repair in cancer cells. Acta Biochim. Biophys. Sin. 2018, 50, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Baniahmad, A.; Taheri, M.; Rashnoo, F. A review on the role of PCA3 lncRNA in carcinogenesis with an especial focus on prostate cancer. Pathol. Res. Pract. 2022, 231, 153800. [Google Scholar] [CrossRef] [PubMed]
- Bourdoumis, A.; Papatsoris, A.G.; Chrisofos, M.; Efstathiou, E.; Skolarikos, A.; Deliveliotis, C. The novel prostate cancer antigen 3 (PCA3) biomarker. Int. Braz. J. Urol. 2010, 36, 665–668, discussion 669. [Google Scholar] [CrossRef] [PubMed]
- Hessels, D.; Schalken, J.A. The use of PCA3 in the diagnosis of prostate cancer. Nat. Rev. Urol. 2009, 6, 255–261. [Google Scholar] [CrossRef]
- Venderbos, L.D.; Luiting, H.; Hogenhout, R.; Roobol, M.J. Interaction of MRI and active surveillance in prostate cancer: Time to re-evaluate the active surveillance inclusion criteria. Urol. Oncol. Semin. Orig. Investig. 2023, 41, 82–87. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Li, Y.Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef]
- Heydari, R.; Fayazzadeh, S.; Shahrokh, S.; Shekari, F.; Farsad, F.; Meyfour, A. Plasma Extracellular Vesicle LncRNA H19 as a Potential Diagnostic Biomarker for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2024, 30, 795–807. [Google Scholar] [CrossRef]
- Chen, S.W.; Wang, P.Y.; Liu, Y.C.; Sun, L.; Zhu, J.; Zuo, S.; Ma, J.; Li, T.Y.; Zhang, J.L.; Chen, G.W.; et al. Effect of Long Noncoding RNA H19 Overexpression on Intestinal Barrier Function and Its Potential Role in the Pathogenesis of Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 2582–2592. [Google Scholar] [CrossRef]
- Benshoshan, M.; Sosnovski, K.E.; Braun, T.; Amir, A.; Anikster, Y.; Haberman Ziv, Y. P041 CDKN2B-AS1 (ANRIL) expression is decreased in Inflammatory Bowel Disease epithelia and in Celiac, and its reduction is linked with induced cells proliferation. J. Crohn’s Colitis 2021, 15, S152. [Google Scholar] [CrossRef]
- Han, J.; Li, Y.; Zhang, B.; Liu, H.; Wu, M.; Zhang, X. lncRNA TUG1 regulates ulcerative colitis through miR-142-5p/SOCS1 axis. Microb. Pathog. 2020, 143, 104139. [Google Scholar] [CrossRef]
- Lucafò, M.; Pugnetti, L.; Bramuzzo, M.; Curci, D.; Di Silvestre, A.; Marcuzzi, A.; Bergamo, A.; Martelossi, S.; Villanacci, V.; Bozzola, A.; et al. Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD. Int. J. Mol. Sci. 2019, 20, 5280. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Li, X.F.; Cheng, L.N.; Li, X.L. Long non-coding RNA CRNDE promotes cell apoptosis by suppressing miR-495 in inflammatory bowel disease. Exp. Cell Res. 2019, 382, 111484. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ahn, C.; Chun, C.H.; Jin, E.J. A long non-coding RNA, GAS5, plays a critical role in the regulation of miR-21 during osteoarthritis. J. Orthop. Res. 2014, 32, 1628–1635. [Google Scholar] [CrossRef]
- Jiang, Y.; Du, T. Relation of circulating lncRNA GAS5 and miR-21 with biochemical indexes, stenosis severity, and inflammatory cytokines in coronary heart disease patients. J. Clin. Lab. Anal. 2022, 36, e24202. [Google Scholar] [CrossRef] [PubMed]
- Samsonov, R.; Burdakov, V.; Shtam, T.; Radzhabova, Z.; Vasilyev, D.; Tsyrlina, E.; Titov, S.; Ivanov, M.; Berstein, L.; Filatov, M.; et al. Plasma exosomal miR-21 and miR-181a differentiates follicular from papillary thyroid cancer. Tumor Biol. 2016, 37, 12011–12021. [Google Scholar] [CrossRef]
- Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 2018, 9, 191. [Google Scholar] [CrossRef]
- Luo, T.; Liu, Q.; Tan, A.; Duan, L.; Jia, Y.; Nong, L.; Tang, J.; Zhou, W.; Xie, W.; Lu, Y.; et al. Mesenchymal Stem Cell-Secreted Exosome Promotes Chemoresistance in Breast Cancer via Enhancing miR-21-5p-Mediated S100A6 Expression. Mol. Ther.-Oncolyt. 2020, 19, 283–293. [Google Scholar] [CrossRef]
- Gao, M.; Yu, T.; Liu, D.; Shi, Y.; Yang, P.; Zhang, J.; Wang, J.; Liu, Y.; Zhang, X. Sepsis plasma-derived exosomal miR-1-3p induces endothelial cell dysfunction by targeting SERP1. Clin. Sci. 2021, 135, 347–365. [Google Scholar] [CrossRef]
- Ibrahim, A.G.; Cheng, K.; Marbán, E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, Y.; Duan, C.; Zhu, K.; Huang, R.; Zhao, H.; Hintze, M.; Pu, Q.; Yuan, Z.; Lv, L.; et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics 2021, 11, 268–291. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Exploratory study on microRNA profiles from plasma-derived extracellular vesicles in Alzheimer’s disease and dementia with Lewy bodies. Transl. Neurodegener. 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, F.; Miao, P.; Yan, L.; Liu, S.; Wang, X.; Jin, Z.; Gu, Z. miR-138 Increases Depressive-Like Behaviors by Targeting SIRT1 in Hippocampus. Neuropsychiatr. Dis. Treat. 2020, 16, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Gartz, M.; Beatka, M.; Prom, M.J.; Strande, J.L.; Lawlor, M.W. Cardiomyocyte-produced miR-339-5p mediates pathology in Duchenne muscular dystrophy cardiomyopathy. Hum. Mol. Genet. 2021, 30, 2347–2361. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Yang, D.; Feng, F.; Zhu, D.; Li, X. miR-3146 induces neutrophil extracellular traps to aggravate gout flare. J. Clin. Lab. Anal. 2021, 35, e24032. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Y.H.; Yoon, C.; Huang, X.Y.; Xu, Y.; Xie, J.W.; Wang, J.B.; Lin, J.X.; Chen, Q.Y.; Cao, L.L.; et al. Circular RNA circ-RanGAP1 regulates VEGFA expression by targeting miR-877-3p to facilitate gastric cancer invasion and metastasis. Cancer Lett. 2020, 471, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Huang, Z.L.; Huang, J.; Xu, B.; Huang, X.Y.; Xu, Y.H.; Zhou, J.; Tang, Z.Y. Exosomal circRNA-100338 promotes hepatocellular carcinoma metastasis via enhancing invasiveness and angiogenesis. J. Exp. Clin. Cancer Res. 2020, 39, 20. [Google Scholar] [CrossRef]
- Pan, Z.; Zhao, R.; Li, B.; Qi, Y.; Qiu, W.; Guo, Q.; Zhang, S.; Zhao, S.; Xu, H.; Li, M.; et al. EWSR1-induced circNEIL3 promotes glioma progression and exosome-mediated macrophage immunosuppressive polarization via stabilizing IGF2BP3. Mol. Cancer 2022, 21, 16. [Google Scholar] [CrossRef]
- Sun, R.; Liu, W.; Zhao, Y.; Chen, H.; Wang, Z.; Zhang, Y.; Sun, X.; Cui, X. Exosomal circRNA as a novel potential therapeutic target for multiple myeloma-related myocardial damage. Cancer Cell Int. 2021, 21, 311. [Google Scholar] [CrossRef]
- Pan, W.; Hu, Y.; Wang, L.; Li, J. Circ_0003611 acts as a miR-885-5p sponge to aggravate the amyloid-β-induced neuronal injury in Alzheimer’s disease. Metab. Brain Dis. 2022, 37, 961–971. [Google Scholar] [CrossRef]
- Xiong, L.; Chen, L.; Wu, L.; He, W.; Chen, D.; Peng, Z.; Li, J.; Zhu, X.; Su, L.; Li, Y.; et al. Lipotoxicity-induced circGlis3 impairs beta cell function and is transmitted by exosomes to promote islet endothelial cell dysfunction. Diabetologia 2022, 65, 188–205. [Google Scholar] [CrossRef]
- Yang, J.; Cheng, M.; Gu, B.; Wang, J.; Yan, S.; Xu, D. CircRNA_09505 aggravates inflammation and joint damage in collagen-induced arthritis mice via miR-6089/AKT1/NF-κB axis. Cell Death Dis. 2020, 11, 833. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Wang, H.; Ye, L.; Shi, X.; Yan, K.; Lin, K.; Huang, Q.; Li, B.; Lin, Q.; Zhu, L.; et al. Hsa_circ_0000479 as a Novel Diagnostic Biomarker of Systemic Lupus Erythematosus. Front. Immunol. 2019, 10, 2281. [Google Scholar] [CrossRef]
- Jiang, Z.; Huang, L.; Chen, L.; Zhou, J.; Liang, B.; Bai, X.; Wu, L.; Huang, H. Circular RNA profile in Graves’ disease and potential function of hsa_circ_0090364. Endocr. Connect. 2022, 11, e220030. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, H.; Chen, Y.; Li, K.; Li, T.; Chen, J.; Zhang, B.; Guo, C.; Qing, L.; Shen, J.; et al. Exosomal long noncoding RNA HOXD-AS1 promotes prostate cancer metastasis via miR-361-5p/FOXM1 axis. Cell Death Dis. 2021, 12, 1129. [Google Scholar] [CrossRef]
- Ni, C.; Fang, Q.Q.; Chen, W.Z.; Jiang, J.X.; Jiang, Z.; Ye, J.; Zhang, T.; Yang, L.; Meng, F.B.; Xia, W.J.; et al. Breast cancer-derived exosomes transmit lncRNA SNHG16 to induce CD73+γδ1 Treg cells. Signal Transduct. Target. Ther. 2020, 5, 41. [Google Scholar] [CrossRef]
- Zheng, H.; Chen, C.; Luo, Y.; Yu, M.; He, W.; An, M.; Gao, B.; Kong, Y.; Ya, Y.; Lin, Y.; et al. Tumor-derived exosomal BCYRN1 activates WNT5A/VEGF-C/VEGFR3 feedforward loop to drive lymphatic metastasis of bladder cancer. Clin. Transl. Med. 2021, 11, e497. [Google Scholar] [CrossRef]
- Sun, L.; He, X.; Zhang, T.; Tao, G.; Wang, X. Knockdown of lnc-KCNC3-3:1 Alleviates the Development of Atherosclerosis via Downregulation of JAK1/STAT3 Signaling Pathway. Front. Cardiovasc. Med. 2021, 8, 701058. [Google Scholar] [CrossRef]
- Sato, M.; Kadomatsu, T.; Miyata, K.; Warren, J.S.; Tian, Z.; Zhu, S.; Horiguchi, H.; Makaju, A.; Bakhtina, A.; Morinaga, J.; et al. The lncRNA Caren antagonizes heart failure by inactivating DNA damage response and activating mitochondrial biogenesis. Nat. Commun. 2021, 12, 2529. [Google Scholar] [CrossRef]
- Wang, Q.; Han, C.L.; Wang, K.L.; Sui, Y.P.; Li, Z.B.; Chen, N.; Fan, S.Y.; Shimabukuro, M.; Wang, F.; Meng, F.G. Integrated analysis of exosomal lncRNA and mRNA expression profiles reveals the involvement of lnc-MKRN2-42:1 in the pathogenesis of Parkinson’s disease. CNS Neurosci. Ther. 2020, 26, 527–537. [Google Scholar] [CrossRef]
- Wang, D.; Wang, P.; Bian, X.; Xu, S.; Zhou, Q.; Zhang, Y.; Ding, M.; Han, M.; Huang, L.; Bi, J.; et al. Elevated plasma levels of exosomal BACE1-AS combined with the volume and thickness of the right entorhinal cortex may serve as a biomarker for the detection of Alzheimer’s disease. Mol. Med. Rep. 2020, 22, 227–238. [Google Scholar] [CrossRef]
- Tian, F.; Wang, J.; Zhang, Z.; Yang, J. LncRNA SNHG7/miR-34a-5p/SYVN1 axis plays a vital role in proliferation, apoptosis and autophagy in osteoarthritis. Biol. Res. 2020, 53, 9. [Google Scholar] [CrossRef] [PubMed]
- Kuai, X.Y.; Yu, S.Y.; Cui, X.F.; Zhao, X.J.; Mao, X.Q.; Yu, Y.; Hu, T.; Zhang, H.J.; Zhou, C.L. LncRNA LUCAT1 as a Plasma Biomarker for Assessing Disease Activity in Adult Patients with Crohn’s Disease. Gastroenterol. Res. Pract. 2021, 2021, 5557357. [Google Scholar] [CrossRef]
- Ji, X.; Sun, J.; Wang, Z. High level of lncRNA NR2F1-AS1 predict the onset and progression of diabetic retinopathy in type 2 diabetes. Exp. Eye Res. 2022, 219, 109069. [Google Scholar] [CrossRef]
- Group, V.S. A randomized controlled clinical trial of intravitreous fomivirsen for treatment of newly diagnosed peripheral cytomegalovirus retinitis in patients with AIDS. Am. J. Ophthalmol. 2002, 133, 467–474. [Google Scholar] [CrossRef]
- Geary, R.S.; Henry, S.P.; Grillone, L.R. Fomivirsen: Clinical pharmacology and potential drug interactions. Clin. Pharmacokinet. 2002, 41, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef]
- Abdelaal, A.M.; Sohal, I.S.; Iyer, S.; Sudarshan, K.; Kothandaraman, H.; Lanman, N.A.; Low, P.S.; Kasinski, A.L. A first-in-class fully modified version of miR-34a with outstanding stability, activity, and anti-tumor efficacy. Oncogene 2023, 42, 2985–2999. [Google Scholar] [CrossRef]
- Liu, N.; Kataoka, M.; Wang, Y.; Pu, L.; Dong, X.; Fu, X.; Zhang, F.; Gao, F.; Liang, T.; Pei, J.; et al. LncRNA LncHrt preserves cardiac metabolic homeostasis and heart function by modulating the LKB1-AMPK signaling pathway. Basic Res. Cardiol. 2021, 116, 48. [Google Scholar] [CrossRef]
- Gragoudas, E.S.; Adamis, A.P.; Cunningham, E.T., Jr.; Feinsod, M.; Guyer, D.R.; VEGF Inhibition Study in Ocular Neovascularization Clinical Trial Group. Pegaptanib for neovascular age-related macular degeneration. N. Engl. J. Med. 2004, 351, 2805–2816. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.D.; Raal, F.J.; Donovan, J.M.; Cromwell, W.C. Mipomersen preferentially reduces small low-density lipoprotein particle number in patients with hypercholesterolemia. J. Clin. Lipidol. 2015, 9, 201–209. [Google Scholar] [CrossRef]
- Mendell, J.R.; Rodino-Klapac, L.R.; Sahenk, Z.; Roush, K.; Bird, L.; Lowes, L.P.; Alfano, L.; Gomez, A.M.; Lewis, S.; Kota, J.; et al. Eteplirsen for the treatment of Duchenne muscular dystrophy. Ann. Neurol. 2013, 74, 637–647. [Google Scholar] [CrossRef]
- Mercuri, E.; Darras, B.T.; Chiriboga, C.A.; Day, J.W.; Campbell, C.; Connolly, A.M.; Iannaccone, S.T.; Kirschner, J.; Kuntz, N.L.; Saito, K.; et al. Nusinersen versus Sham Control in Later-Onset Spinal Muscular Atrophy. N. Engl. J. Med. 2018, 378, 625–635. [Google Scholar] [CrossRef]
- Benson, M.D.; Waddington-Cruz, M.; Berk, J.L.; Polydefkis, M.; Dyck, P.J.; Wang, A.K.; Planté-Bordeneuve, V.; Barroso, F.A.; Merlini, G.; Obici, L.; et al. Inotersen Treatment for Patients with Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M.; Sardh, E.; Ventura, P.; Peiró, P.A.; Rees, D.C.; Stölzel, U.; Bissell, D.M.; Bonkovsky, H.L.; Windyga, J.; Anderson, K.E.; et al. Phase 3 Trial of RNAi Therapeutic Givosiran for Acute Intermittent Porphyria. N. Engl. J. Med. 2020, 382, 2289–2301. [Google Scholar] [CrossRef]
- Scaglioni, D.; Catapano, F.; Ellis, M.; Torelli, S.; Chambers, D.; Feng, L.; Beck, M.; Sewry, C.; Monforte, M.; Harriman, S.; et al. The administration of antisense oligonucleotide golodirsen reduces pathological regeneration in patients with Duchenne muscular dystrophy. Acta Neuropathol. Commun. 2021, 9, 7. [Google Scholar] [CrossRef]
- Kolovou, G.; Kolovou, V.; Katsiki, N. Volanesorsen: A New Era in the Treatment of Severe Hypertriglyceridemia. J. Clin. Med. 2022, 11, 982. [Google Scholar] [CrossRef] [PubMed]
- Roshmi, R.R.; Yokota, T. Pharmacological Profile of Viltolarsen for the Treatment of Duchenne Muscular Dystrophy: A Japanese Experience. Clin. Pharmacol. 2021, 13, 235–242. [Google Scholar] [CrossRef]
- Garrelfs, S.F.; Frishberg, Y.; Hulton, S.A.; Koren, M.J.; O’Riordan, W.D.; Cochat, P.; Deschênes, G.; Shasha-Lavsky, H.; Saland, J.M.; Van’t Hoff, W.G.; et al. Lumasiran, an RNAi Therapeutic for Primary Hyperoxaluria Type 1. N. Engl. J. Med. 2021, 384, 1216–1226. [Google Scholar] [CrossRef]
- Zakeri, S.E.; Pradeep, S.P.; Kasina, V.; Laddha, A.P.; Manautou, J.E.; Bahal, R. Casimersen for the treatment of Duchenne muscular dystrophy. Trends Pharmacol. Sci. 2022, 43, 607–608. [Google Scholar] [CrossRef]
- Samuel, E.; Watford, M.; Egolum, U.O.; Ombengi, D.N.; Ling, H.; Cates, D.W. Inclisiran: A First-in-Class siRNA Therapy for Lowering Low-Density Lipoprotein Cholesterol. Ann. Pharmacother. 2023, 57, 317–324. [Google Scholar] [CrossRef]
- Nie, T.; Heo, Y.A.; Shirley, M. Vutrisiran: A Review in Polyneuropathy of Hereditary Transthyretin-Mediated Amyloidosis. Drugs 2023, 83, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.A.; Langman, C.; Cochat, P.; Lieske, J.C.; Moochhala, S.H.; Hamamoto, S.; Satoh, H.; Mourani, C.; Ariceta, G.; Torres, A.; et al. PHYOX2: A pivotal randomized study of nedosiran in primary hyperoxaluria type 1 or 2. Kidney Int. 2023, 103, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Coelho, T.; Marques, W., Jr.; Dasgupta, N.R.; Chao, C.C.; Parman, Y.; França, M.C., Jr.; Guo, Y.C.; Wixner, J.; Ro, L.S.; Calandra, C.R.; et al. Eplontersen for Hereditary Transthyretin Amyloidosis With Polyneuropathy. JAMA 2023, 330, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.M.; Cudkowicz, M.E.; Genge, A.; Shaw, P.J.; Sobue, G.; Bucelli, R.C.; Chiò, A.; Van Damme, P.; Ludolph, A.C.; Glass, J.D.; et al. Trial of Antisense Oligonucleotide Tofersen for SOD1 ALS. N. Engl. J. Med. 2022, 387, 1099–1110. [Google Scholar] [CrossRef]
- Bergmark, B.A.; Marston, N.A.; Prohaska, T.A.; Alexander, V.J.; Zimerman, A.; Moura, F.A.; Murphy, S.A.; Goodrich, E.L.; Zhang, S.; Gaudet, D.; et al. Olezarsen for Hypertriglyceridemia in Patients at High Cardiovascular Risk. N. Engl. J. Med. 2024, 390, 1770–1780. [Google Scholar] [CrossRef]
- Zhou, W.; Jiang, L.; Liao, S.; Wu, F.; Yang, G.; Hou, L.; Liu, L.; Pan, X.; Jia, W.; Zhang, Y. Vaccines’ New Era-RNA Vaccine. Viruses 2023, 15, 1760. [Google Scholar] [CrossRef]
- He, Q.; Gao, H.; Tan, D.; Zhang, H.; Wang, J.Z. mRNA cancer vaccines: Advances, trends and challenges. Acta Pharm. Sin. B 2022, 12, 2969–2989. [Google Scholar] [CrossRef]
- Vishweshwaraiah, Y.L.; Dokholyan, N.V. mRNA vaccines for cancer immunotherapy. Front. Immunol. 2022, 13, 1029069. [Google Scholar] [CrossRef]
- Niu, D.; Wu, Y.; Lian, J. Circular RNA vaccine in disease prevention and treatment. Signal Transduct. Target. Ther. 2023, 8, 341. [Google Scholar] [CrossRef] [PubMed]
- Wesselhoeft, R.A.; Kowalski, P.S.; Parker-Hale, F.C.; Huang, Y.; Bisaria, N.; Anderson, D.G. RNA Circularization Diminishes Immunogenicity and Can Extend Translation Duration In Vivo. Mol. Cell 2019, 74, 508–520.e504. [Google Scholar] [CrossRef]
- Kameda, S.; Ohno, H.; Saito, H. Synthetic circular RNA switches and circuits that control protein expression in mammalian cells. Nucleic Acids Res. 2023, 51, e24. [Google Scholar] [CrossRef]
- Usman, N.; Cedergren, R. Exploiting the chemical synthesis of RNA. Trends Biochem. Sci. 1992, 17, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Rong, M.; He, B.; McAllister, W.T.; Durbin, R.K. Promoter specificity determinants of T7 RNA polymerase. Proc. Natl. Acad. Sci. USA 1998, 95, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Appel, B. In vitro circularization of RNA. Rna Biol. 2017, 14, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, N.I.; Ashirbekova, D.T.; Dolinnaya, N.G.; Shabarova, Z.A. Chemical reactions within DNA duplexes cyanogen bromide as an effective oligodeoxyribonucleotide coupling agent. FEBS Lett. 1988, 232, 153–155. [Google Scholar] [CrossRef]
- Petkovic, S.; Müller, S. RNA circularization strategies in vivo and in vitro. Nucleic Acids Res. 2015, 43, 2454–2465. [Google Scholar] [CrossRef]
- Bai, Y.; Liu, D.; He, Q.; Liu, J.; Mao, Q.; Liang, Z. Research progress on circular RNA vaccines. Front. Immunol. 2022, 13, 1091797. [Google Scholar] [CrossRef]
- Xie, J.; Ye, F.; Deng, X.; Tang, Y.; Liang, J.Y.; Huang, X.; Sun, Y.; Tang, H.; Lei, J.; Zheng, S.; et al. Circular RNA: A promising new star of vaccine. J. Transl. Intern. Med. 2023, 11, 372–381. [Google Scholar] [CrossRef]
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P.H. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982, 1, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.M.; Wolf, E.D.; Wu, R.; Sanford, J.C. High-velocity microprojectiles for delivering nucleic acids into living cells. 1987. Biotechnology 1992, 24, 384–386. [Google Scholar] [PubMed]
- Sun, X.; Zeng, L.; Huang, Y. Transcutaneous delivery of DNA/mRNA for cancer therapeutic vaccination. J. Gene Med. 2019, 21, e3089. [Google Scholar] [CrossRef] [PubMed]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef]
- Cullis, P.R.; Felgner, P.L. The 60-year evolution of lipid nanoparticles for nucleic acid delivery. Nat. Rev. Drug Discov. 2024, 23, 709–722. [Google Scholar] [CrossRef]
- Laumont, C.M.; Vincent, K.; Hesnard, L.; Audemard, É.; Bonneil, É.; Laverdure, J.P.; Gendron, P.; Courcelles, M.; Hardy, M.P.; Côté, C.; et al. Noncoding regions are the main source of targetable tumor-specific antigens. Sci. Transl. Med. 2018, 10, eaau5516. [Google Scholar] [CrossRef]
- Chong, C.; Coukos, G.; Bassani-Sternberg, M. Identification of tumor antigens with immunopeptidomics. Nat. Biotechnol. 2022, 40, 175–188. [Google Scholar] [CrossRef]
- Huang, D.; Zhu, X.; Ye, S.; Zhang, J.; Liao, J.; Zhang, N.; Zeng, X.; Wang, J.; Yang, B.; Zhang, Y.; et al. Tumour circular RNAs elicit anti-tumour immunity by encoding cryptic peptides. Nature 2024, 625, 593–602. [Google Scholar] [CrossRef]
- Qu, L.; Yi, Z.; Shen, Y.; Lin, L.; Chen, F.; Xu, Y.; Wu, Z.; Tang, H.; Zhang, X.; Tian, F.; et al. Circular RNA vaccines against SARS-CoV-2 and emerging variants. Cell 2022, 185, 1728–1744.e1716. [Google Scholar] [CrossRef]
- Li, H.; Peng, K.; Yang, K.; Ma, W.; Qi, S.; Yu, X.; He, J.; Lin, X.; Yu, G. Circular RNA cancer vaccines drive immunity in hard-to-treat malignancies. Theranostics 2022, 12, 6422–6436. [Google Scholar] [CrossRef]
- Seephetdee, C.; Bhukhai, K.; Buasri, N.; Leelukkanaveera, P.; Lerdwattanasombat, P.; Manopwisedjaroen, S.; Phueakphud, N.; Kuhaudomlarp, S.; Olmedillas, E.; Saphire, E.O.; et al. A circular mRNA vaccine prototype producing VFLIP-X spike confers a broad neutralization of SARS-CoV-2 variants by mouse sera. Antivir. Res. 2022, 204, 105370. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhu, J.; Sun, J.; Chen, Y.; Du, Y.; Tan, Y.; Wu, L.; Zhai, M.; Wei, L.; Li, N.; et al. Intratumoral delivered novel circular mRNA encoding cytokines for immune modulation and cancer therapy. Mol. Ther. Nucleic Acids 2022, 30, 184–197. [Google Scholar] [CrossRef]
- Zhou, J.; Ye, T.; Yang, Y.; Li, E.; Zhang, K.; Wang, Y.; Chen, S.; Hu, J.; Zhang, K.; Liu, F.; et al. Circular RNA vaccines against monkeypox virus provide potent protection against vaccinia virus infection in mice. Mol. Ther. 2024, 32, 1779–1789. [Google Scholar] [CrossRef]
- Wang, F.; Cai, G.; Wang, Y.; Zhuang, Q.; Cai, Z.; Li, Y.; Gao, S.; Li, F.; Zhang, C.; Zhao, B.; et al. Circular RNA-based neoantigen vaccine for hepatocellular carcinoma immunotherapy. MedComm 2024, 5, e667. [Google Scholar] [CrossRef]
- Wan, J.; Wang, Z.; Wang, L.; Wu, L.; Zhang, C.; Zhou, M.; Fu, Z.F.; Zhao, L. Circular RNA vaccines with long-term lymph node-targeting delivery stability after lyophilization induce potent and persistent immune responses. MBio 2024, 15, e0177523. [Google Scholar] [CrossRef]
- Xu, S.; Xu, Y.; Solek, N.C.; Chen, J.; Gong, F.; Varley, A.J.; Golubovic, A.; Pan, A.; Dong, S.; Zheng, G.; et al. Tumor-Tailored Ionizable Lipid Nanoparticles Facilitate IL-12 Circular RNA Delivery for Enhanced Lung Cancer Immunotherapy. Adv. Mater. 2024, 36, e2400307. [Google Scholar] [CrossRef]
- Yue, X.; Zhong, C.; Cao, R.; Liu, S.; Qin, Z.; Liu, L.; Zhai, Y.; Luo, W.; Lian, Y.; Zhang, M.; et al. CircRNA based multivalent neuraminidase vaccine induces broad protection against influenza viruses in mice. Npj Vaccines 2024, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Z.; Li, X.; Wu, W.; Jiang, H.; Zheng, Y.; Zhou, J.; Ye, X.; Lu, J.; Wang, W.; et al. A single-dose circular RNA vaccine prevents Zika virus infection without enhancing dengue severity in mice. Nat. Commun. 2024, 15, 8932. [Google Scholar] [CrossRef]
- Jenike, A.E.; Halushka, M.K. miR-21: A non-specific biomarker of all maladies. Biomark. Res. 2021, 9, 18. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Bjartell, A.; Chinnaiyan, A.M.; Jenster, G.; Nam, R.K.; Rubin, M.A.; Schalken, J.A. ETS gene fusions in prostate cancer: From discovery to daily clinical practice. Eur. Urol. 2009, 56, 275–286. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, L.; Wang, Z. PCA3 and TMPRSS2-ERG gene fusions as diagnostic biomarkers for prostate cancer. Chin. J. Cancer Res. 2016, 28, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Warli, S.M.; Warli, M.H.; Prapiska, F.F. PCA3 and TMPRSS2: ERG Urine Level as Diagnostic Biomarker of Prostate Cancer. Res. Rep. Urol. 2023, 15, 149–155. [Google Scholar] [CrossRef] [PubMed]
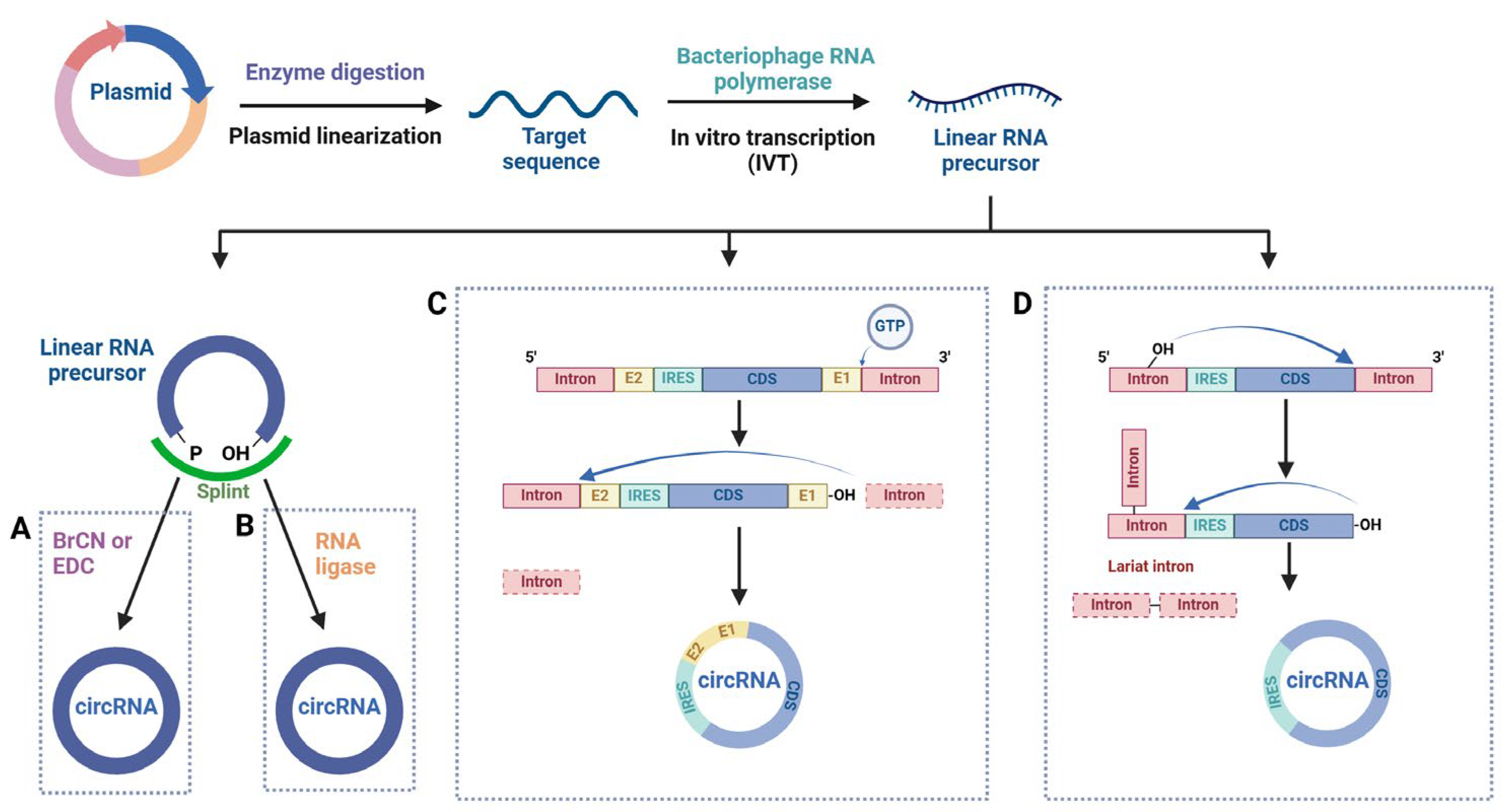
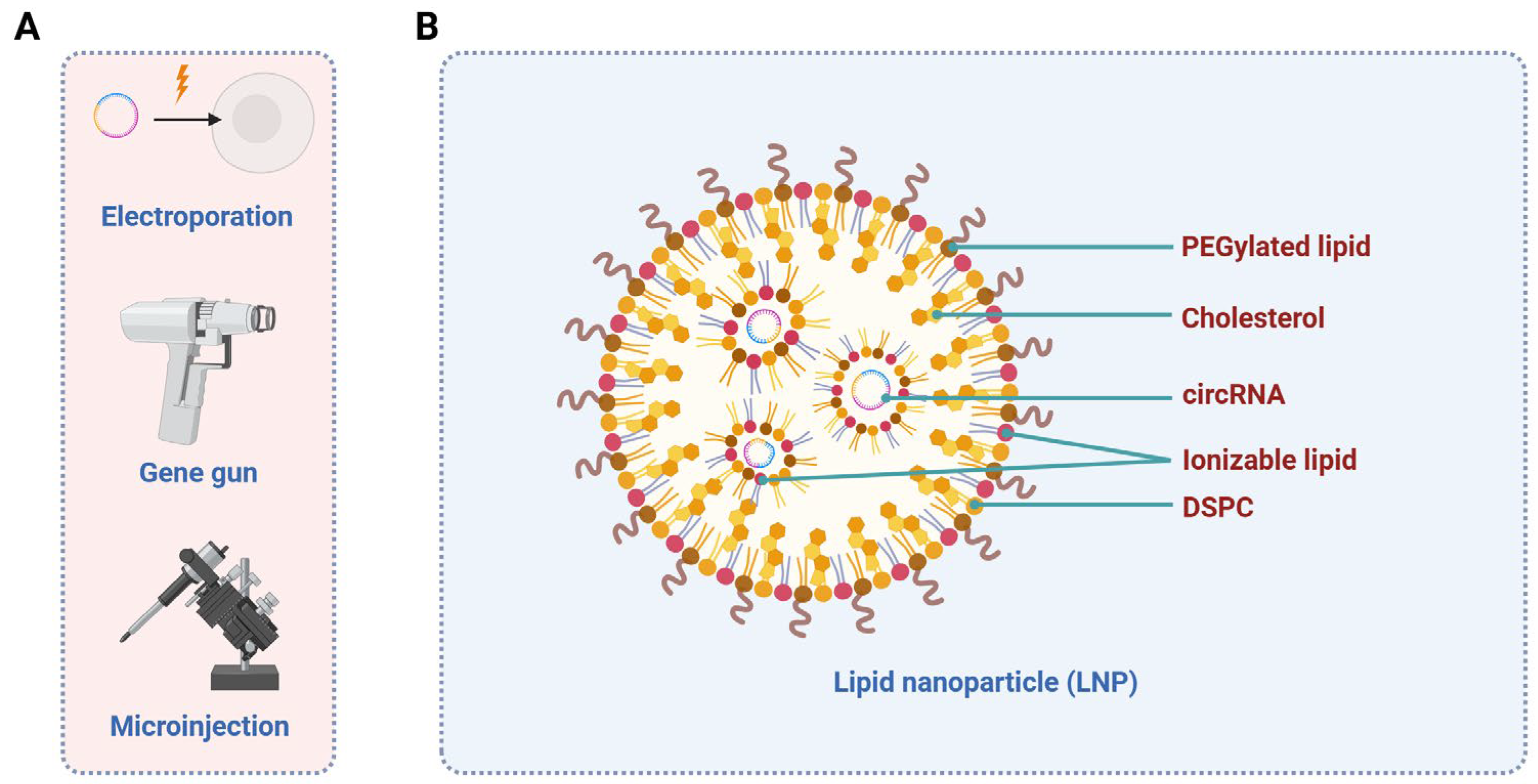
| Related Diseases | Biofluid | Potential Biomarkers | Target Genes/Pathways/Mechanistic Approaches | References |
|---|---|---|---|---|
| Thyroid Cancer | Plasma | miR-21 and miR-181a-5p | N/A | [75] |
| Liver Cancer | CCM and serum | miR-1247-3p | B4GALT3 | [76] |
| Breast Cancer | MSC CCM | miR-21-5p | S100A6 | [77] |
| Sepsis | Plasma | miR-1-3p | SERP1 | [78] |
| Heart Failure | Plasma | miR-146a | IRAK-1, TRAF6, NOX-4 SMAD4, and TGF-β | [79] |
| Myocardial Infarction | Cardiac telocyte CCM | miR-21-5p | CDIP1 | [80] |
| Alzheimer’s Disease | Plasma | miR-451a and miR-21-5p | N/A | [81] |
| Depressive Disorder | Cortical neuron CCM | miR-138 | SIRT1 | [82] |
| Duchenne Muscular Dystrophy | DMD cardiomyocytes CCM | miR-339-5p | MDM2, GSK3A and MAP2K3 | [83] |
| Gout | Plasma | miR-3146 | Mediates NETs formation | [84] |
| Gastric Cancer | Plasma | circ-RanGAP1 | miR-877-3p/VEGFA axis | [85] |
| Hepatocellular Carcinoma | HCC CCM | circRNA-100338 | N/A | [86] |
| Glioma | GBM CCM | circNEIL3 | Stabilizing IGF2BP3 | [87] |
| Myeloma-Related Myocardial Damage | Serum | circ-G042080 | miR-4268/TLR4 axis | [88] |
| Alzheimer’s Disease | Serum | circ_0003611 | miR-885-5p/KREMEN1 axis | [89] |
| Type 2 Diabetes | Serum | circGlis3 | Regulates GMEB1 degradation and HSP27 phosphorylation | [90] |
| Rheumatoid Arthritis | Plasma | circRNA_09505 | miR-6089/AKT1/NF-κB axis | [91] |
| Systemic Lupus Erythematosus | Plasma | hsa_circ_0000479 | Metabolic and the Wnt signaling pathway | [92] |
| Graves’ Disease | Plasma | hsa_circ_0090364 | hsa-miR-378a-3p/IL-6ST/IL21R axis | [93] |
| Prostate Cancer | Serum | HOXD-AS1 | miR-361-5p/FOXM1 axis | [94] |
| Breast Cancer | Serum | SNHG16 | miR-16–5p/SMAD5 axis | [95] |
| Bladder Cancer | Urine | lncBCYRN1 | Activates WNT5A/VEGF-C/VEGFR3 feedforward loop | [96] |
| Atherosclerosis | HUVEC CCM | lnc-KCNC3-3:1 | JAK1/STAT3 signaling pathway | [97] |
| Heart Failure | Plasma | lncRNA-NRF | N/A | [98] |
| Parkinson’s Disease | Plasma | lnc-MKRN2-42:1 | N/A | [99] |
| Alzheimer’s Disease | Plasma | BACE1-AS | N/A | [100] |
| Osteoarthritis | OA CCM | SNHG7 | miR-34a-5p/SYVN1 axis | [101] |
| Crohn’s Disease | Plasma | LUCAT1 | NA | [102] |
| Diabetic Retinopathy in Type 2 Diabetes | Plasma | NR1F1-AS2 | Moderates EndMT | [103] |
| Drugs | Types | Route of Administration | Target Organ | Mechanism of Action | Related Diseases | FDA and/or EMA Approval Year | References |
|---|---|---|---|---|---|---|---|
| Fomivirsen | ASO | IVT | Eye | Targeting and silencing the mRNA of CMV IE2 protein | CMV retinitis in immunocompromised patients | FDA (1998) EMA (1999) | [104] |
| Pegaptanib | Phosphate oligonucleotide aptamer | IVT | Eye | Inhibiting VEGF-165 | nAMD | FDA (2004) | [111] |
| Mipomersen | ASO | SC | Liver | Targeting and silencing the mRNA of apolipoprotein B to reduce LDL levels | HoFH | FDA (2013) EMA (2012) | [112] |
| Eteplirsen | ASO | IV | Muscle | Targeting and splicing the pre-mRNA of defective DMD dystrophin protein | DMD | FDA (2016) | [113] |
| Nusinersen | ASO | ITH | Central nervous system | Targeting and splicing the pre-mRNA of defective SMN protein | SMA | FDA (2016) EMA (2017) | [114] |
| Patisiran | siRNA | IV | Liver | Targeting and silencing the mRNA of TTR protein to prevent the production of TTR protein | ATTR amyloidosis | FDA (2018) | [106] |
| Inotersen | ASO | SC | Liver | Targeting and silencing the mRNA of TTR protein to prevent the production of TTR protein | ATTR amyloidosis | FDA (2018) EMA (2018) | [115] |
| Givosiran | siRNA | SC | Liver | Targeting and silencing the mRNA of ALAS1 to reduce ALAS1 levels | AHP | FDA (2019) EMA (2020) | [116] |
| Golodirsen | ASO | IV | Muscle | Targeting the splicing of DMD pre-mRNA (exon 53 skipping) | DMD | FDA (2019) | [117] |
| Volanesorsen | ASO | SC | Liver | Targeting and silencing the mRNA of APOC3 to reduce triglyceride production | FCS | EMA (2019) | [118] |
| Viltolarsen | ASO | IV | Muscle | Targeting the splicing of DMD pre-mRNA (exon 53 skipping) | DMD | FDA (2020) | [119] |
| Lumasiran | siRNA | SC | Liver | Targeting and silencing the mRNA of HAO1 to reduce GO levels | PH1 | FDA (2020) EMA (2020) | [120] |
| Casimersen | ASO | IV | Muscle | Targeting the splicing of DMD pre-mRNA (exon 45 skipping) | DMD | FDA (2021) | [121] |
| Inclisiran | siRNA | SC | Liver | Targeting PCSK9 to inhibit its synthesis and reduce LDL-C levels | ASCVD | FDA (2021) EMA (2020) | [122] |
| Vutrisiran | siRNA | SC | Liver | Targeting and silencing the mRNA of TTR protein to prevent the production of TTR protein | ATTR amyloidosis | FDA (2022) EMA (2022) | [123] |
| Nedosiran | siRNA | SC | Liver | Targeting liver LDH mRNA to reduce the expression of LDH | PH1 | FDA (2023) | [124] |
| Eplontersen | ASO | SC | Liver | Targeting and silencing the mRNA of TTR protein to prevent the production of TTR protein | ATTR amyloidosis | FDA (2023) | [125] |
| Tofersen | ASO | ITH | Muscle | Targeting SOD1 mRNA to reduce the synthesis of SOD1 protein | SOD1-ALS | FDA (2023) EMA (2023) | [126] |
| Olezarsen | ASO | SC | Liver | Reducing hepatic synthesis of apolipoprotein C-III to lower plasma triglyceride levels | FCS | FDA (2024) | [127] |
| Vaccines | Cyclization Strategy | Delivery | Antigen | References |
|---|---|---|---|---|
| circRNARBD | Ribozymatic autocatalysis | LNP | SARS-CoV-2 RBD antigen | [149] |
| VFLIP-X | T4 RNA ligase | LNP | SARS-CoV-2 spiking protein | [151] |
| circRNAOVA-luc-LNP | Ribozymatic autocatalysis | LNP | OVA [257-264]-luciferase | [150] |
| CircRNA encoding cytokines | Ribozymatic autocatalysis | LNP | Active IL-15\IL-12\GM-CSF\IFN-α 2b | [152] |
| cirA29L, cirA35R, cirB6R, and cirM1R | Ribozymatic autocatalysis | LNP | MPXV proteins A29L, A35R, B6R, and M1R | [153] |
| circRNA3×PTPN2 | Ribozymatic autocatalysis | LNP | PTPN2 | [154] |
| circRNA-G | Ribozymatic autocatalysis | LNP | Glycoproteins of the RABV vaccine strain SAD-L16 | [155] |
| IL12-circRNA | Ribozymatic autocatalysis | LNP | IL-12 | [156] |
| circRNA-NA | Ribozymatic autocatalysis | LNP | NA | [157] |
| EDIII-Fc circRNA | Ribozymatic autocatalysis | LNP | Dimeric EDIII-Fc fusion | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.-X.; Li, H.; Cheng, Z.-H.; Sun, H.-Y.; Huang, J.-P.; Li, Z.-P.; Li, X.-X.; Hu, Z.-G.; Wang, J. The Application of Non-Coding RNAs as Biomarkers, Therapies, and Novel Vaccines in Diseases. Int. J. Mol. Sci. 2025, 26, 3055. https://doi.org/10.3390/ijms26073055
Yang L-X, Li H, Cheng Z-H, Sun H-Y, Huang J-P, Li Z-P, Li X-X, Hu Z-G, Wang J. The Application of Non-Coding RNAs as Biomarkers, Therapies, and Novel Vaccines in Diseases. International Journal of Molecular Sciences. 2025; 26(7):3055. https://doi.org/10.3390/ijms26073055
Chicago/Turabian StyleYang, Lu-Xuan, Hui Li, Zhi-Hui Cheng, He-Yue Sun, Jie-Ping Huang, Zhi-Peng Li, Xin-Xin Li, Zhi-Gang Hu, and Jian Wang. 2025. "The Application of Non-Coding RNAs as Biomarkers, Therapies, and Novel Vaccines in Diseases" International Journal of Molecular Sciences 26, no. 7: 3055. https://doi.org/10.3390/ijms26073055
APA StyleYang, L.-X., Li, H., Cheng, Z.-H., Sun, H.-Y., Huang, J.-P., Li, Z.-P., Li, X.-X., Hu, Z.-G., & Wang, J. (2025). The Application of Non-Coding RNAs as Biomarkers, Therapies, and Novel Vaccines in Diseases. International Journal of Molecular Sciences, 26(7), 3055. https://doi.org/10.3390/ijms26073055