
Thieno[2,3-b]pyridines as a Novel Strategy Against Cervical Cancer: Mechanistic Insights and Therapeutic Potential

 , , ,
, , ,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Results
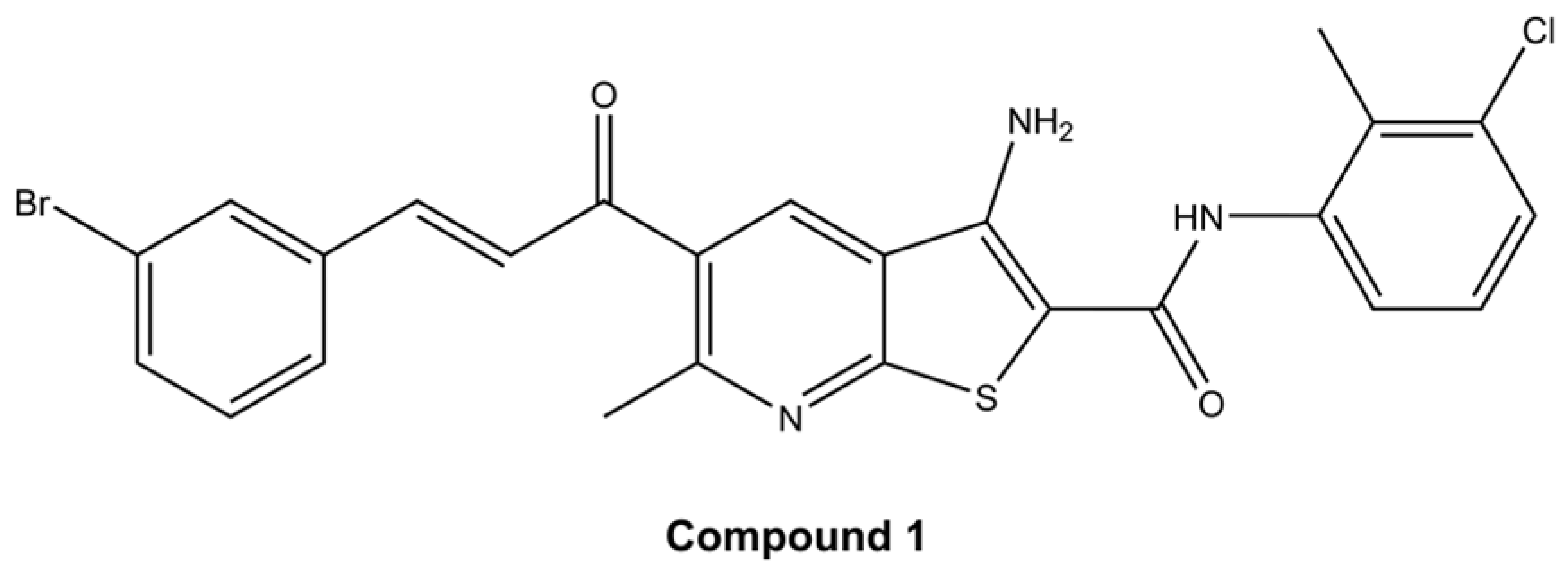
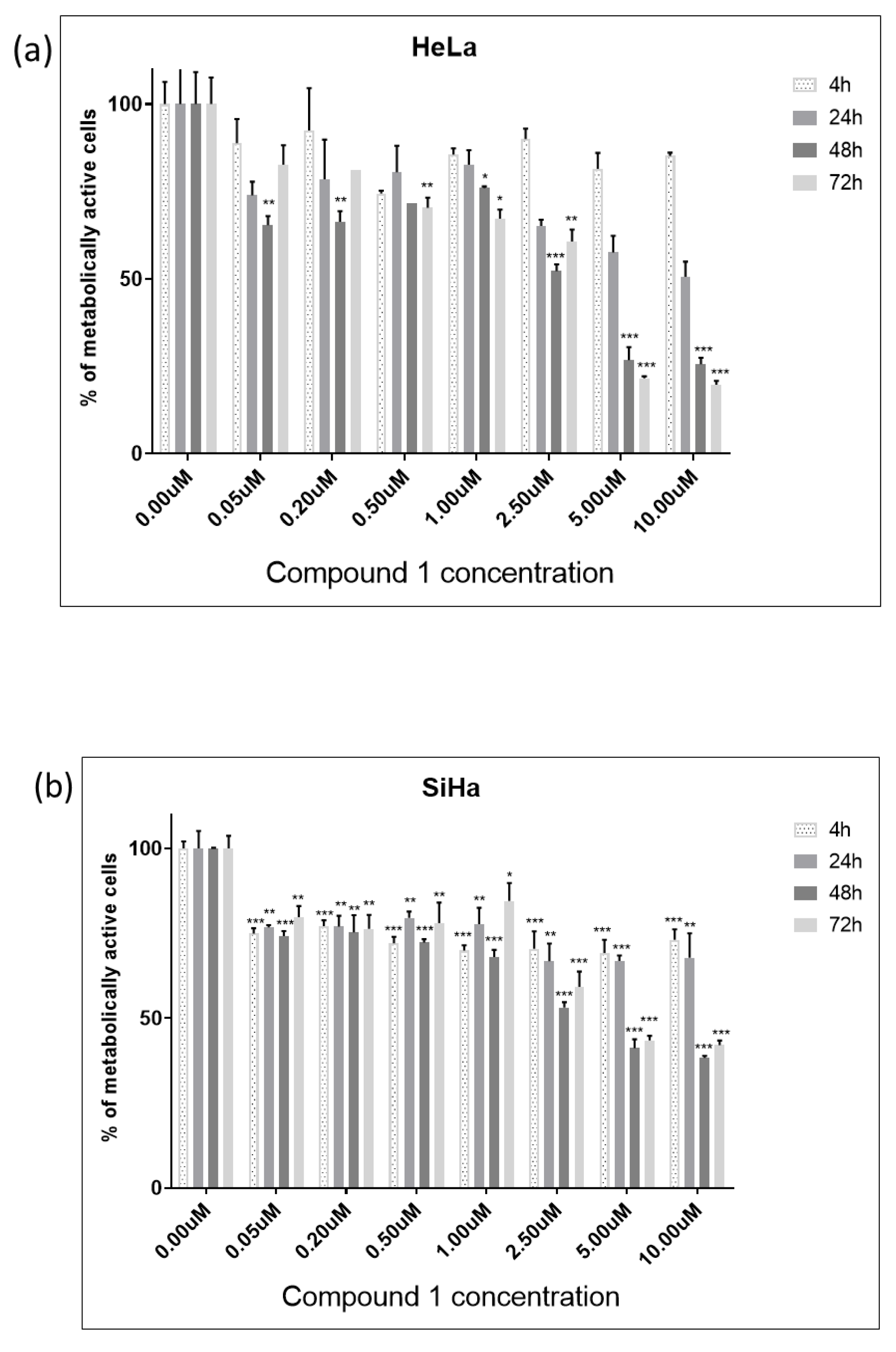
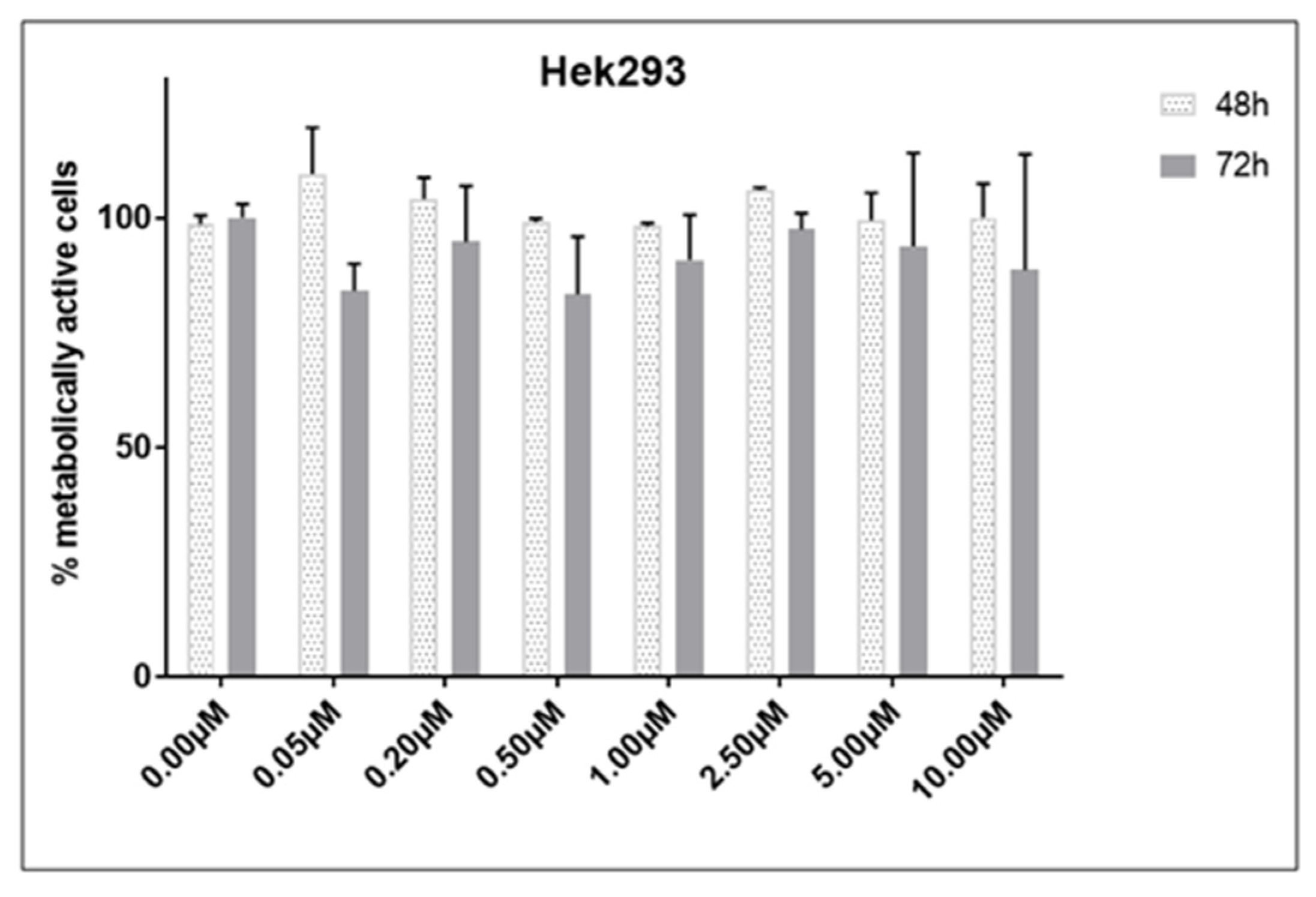
2.1. Cell Viability
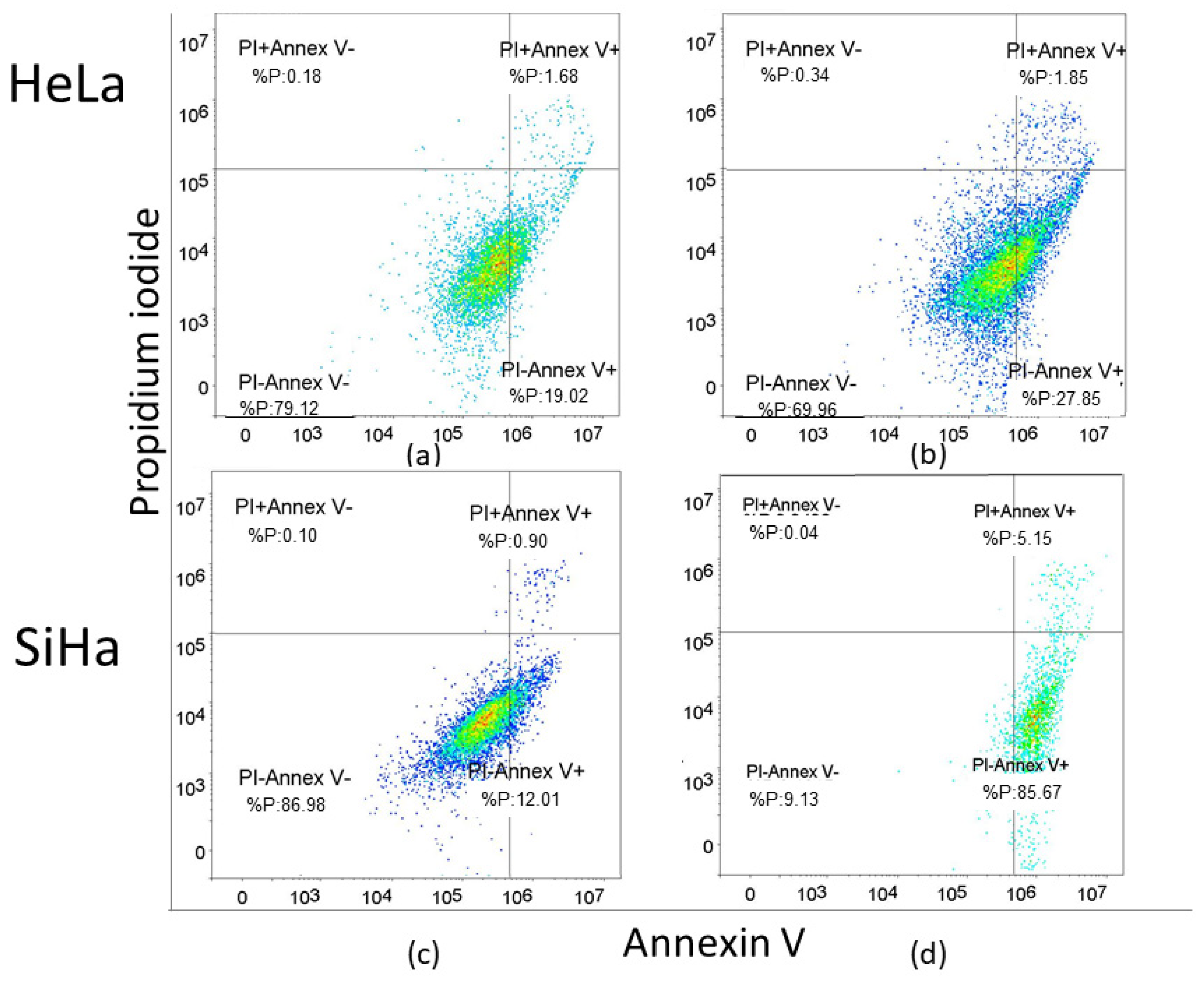
2.2. Apoptosis Rate
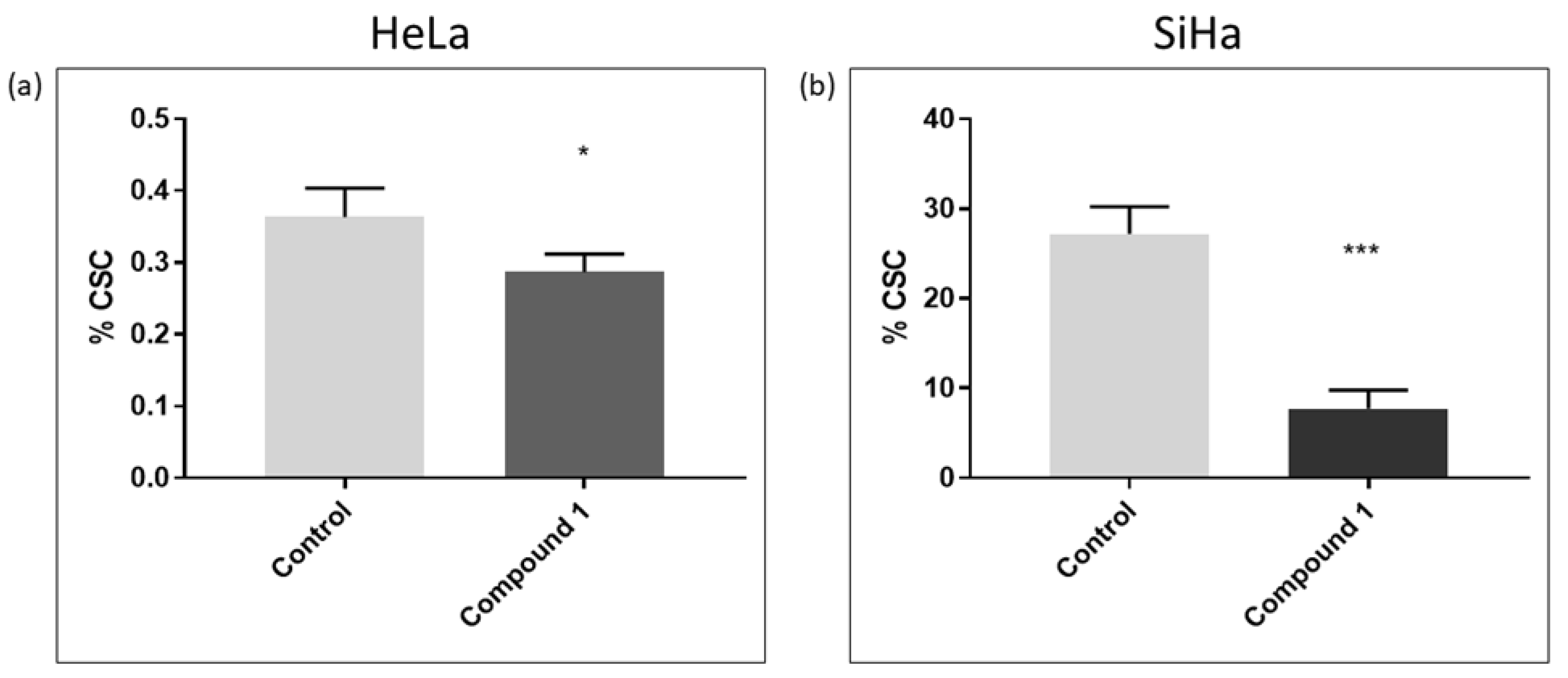
2.3. Cancer Stem Cells
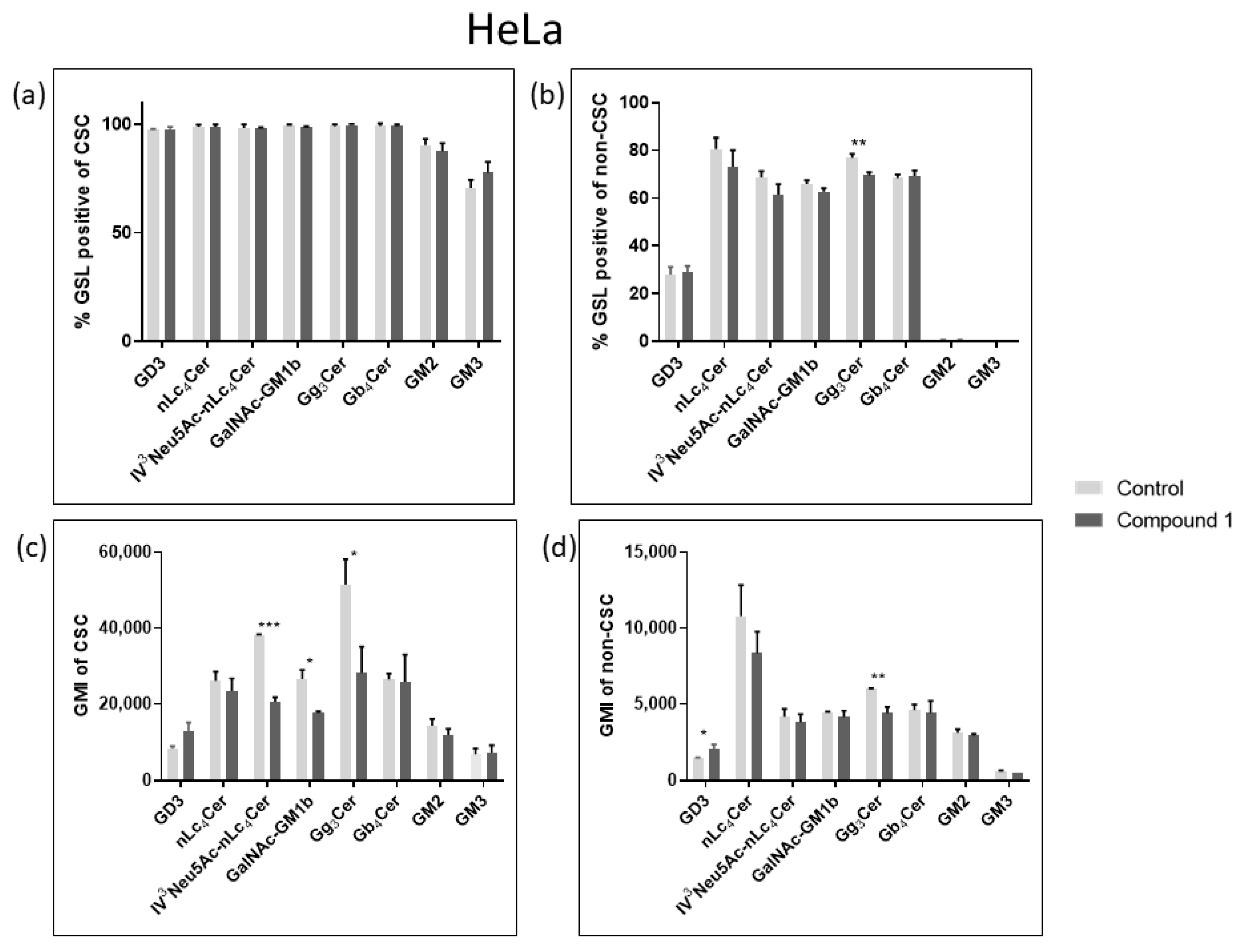
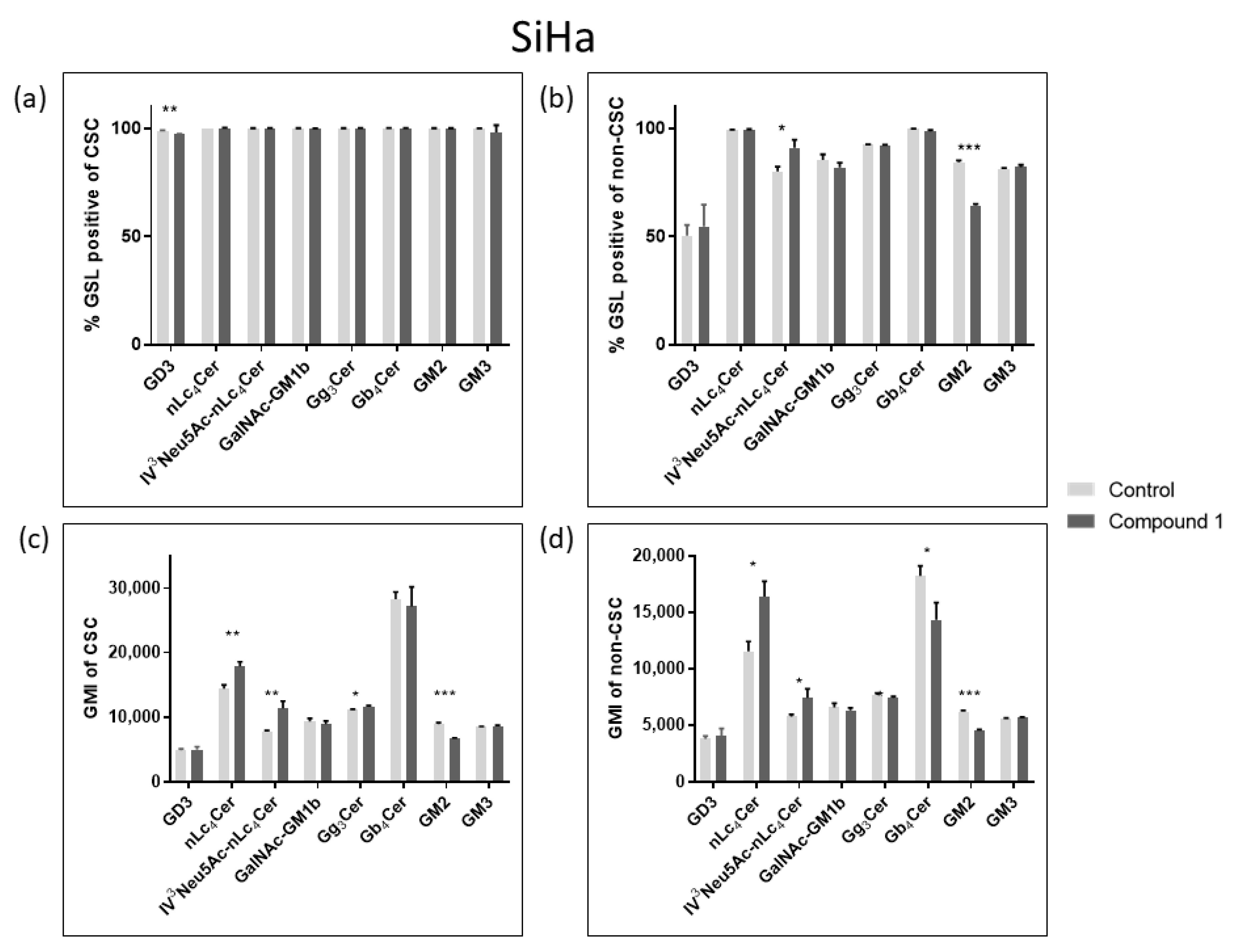
2.4. Expression of Glycosphingolipids
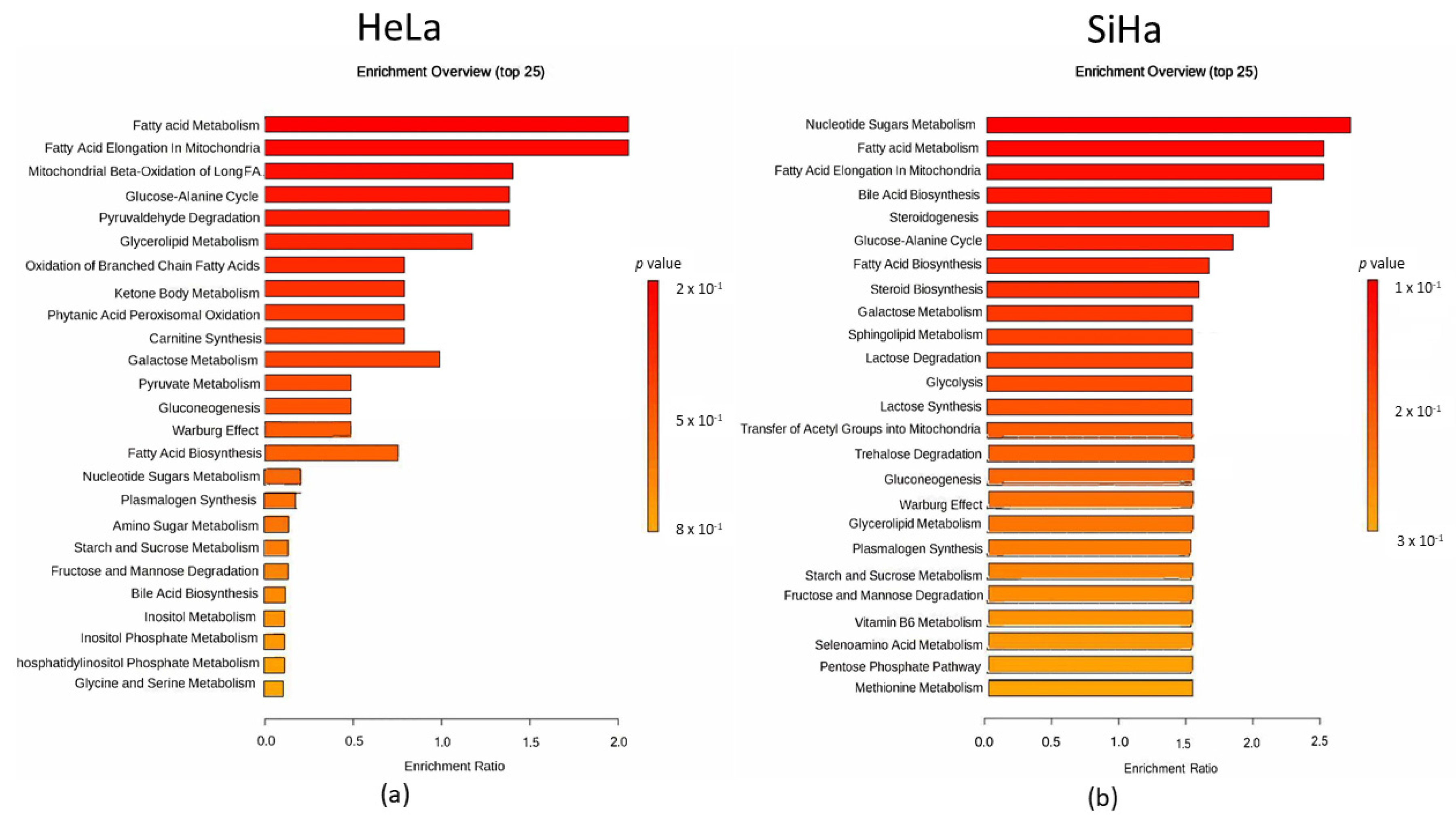
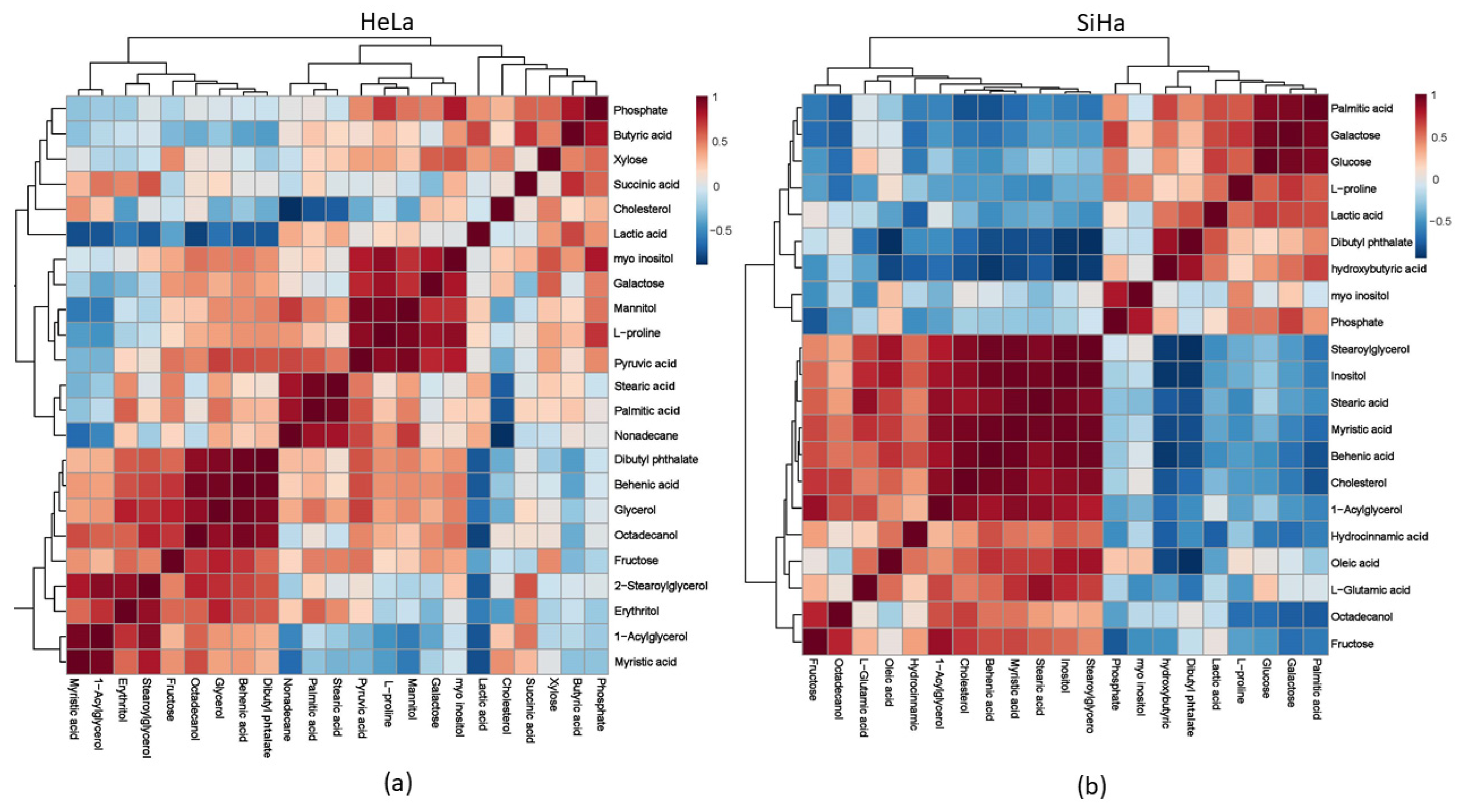
2.5. Metabolites
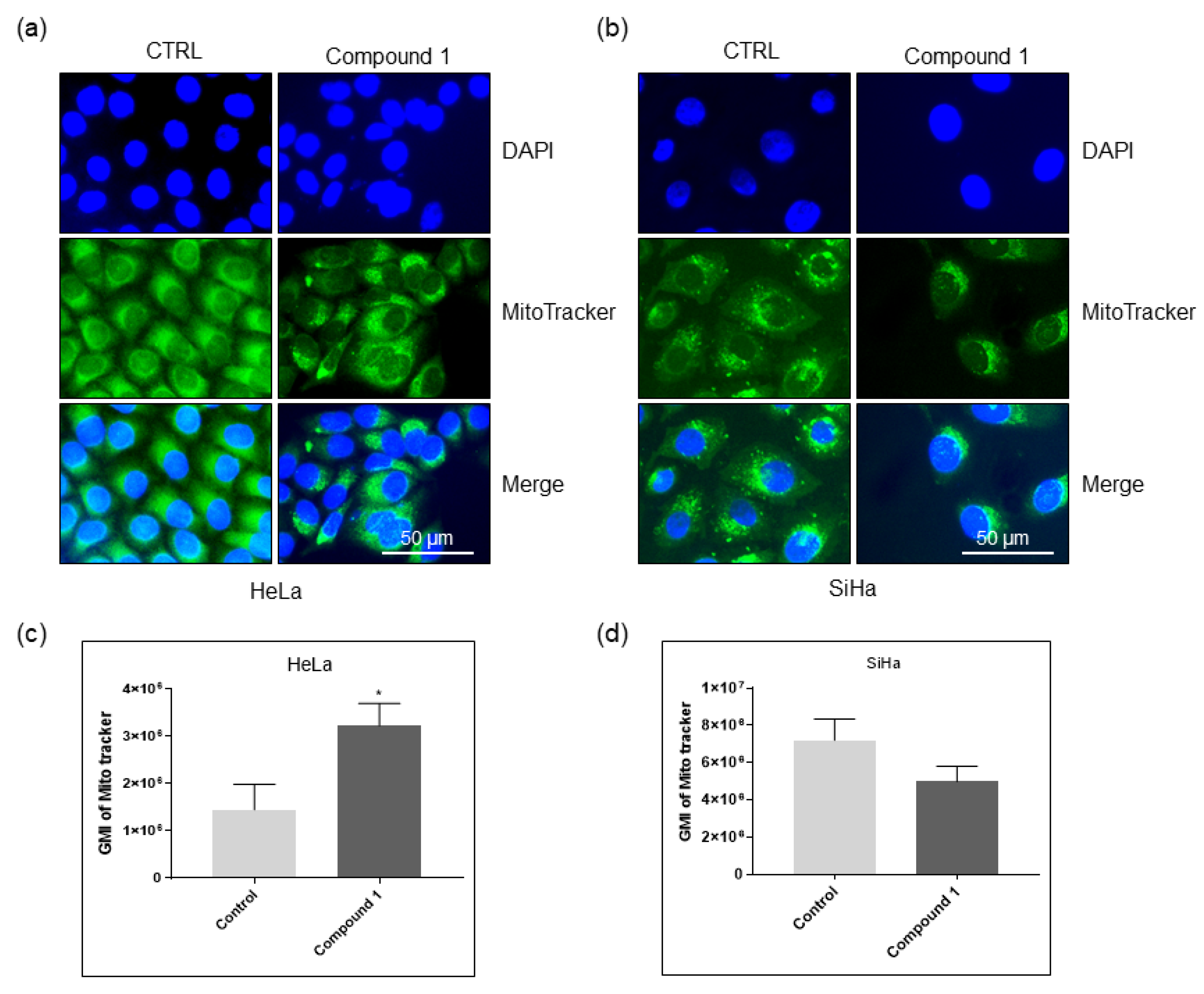
2.6. Effect on Mitochondria
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Thieno[2,3-b]pyridine Compound
4.2. Measurement of Cytotoxic Activity
4.3. Flow Cytometric Analyses
4.3.1. Apoptosis
4.3.2. Glycosphingolipid (GSL) Expression
4.3.3. Tracking of Mitochondria
4.4. Statistical Analysis
4.5. Metabolite Analyses
4.5.1. Sample Collection
4.5.2. Sample Derivatization and GC-MS Analysis
4.5.3. GC-MS Data Pre-Processing and Statistical Analysis
4.6. Fluorescence Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.F.; Peto, J.; Meijer, C.J.L.M.; Muñoz, N. Human Papillomavirus Is a Necessary Cause of Invasive Cervical Cancer Worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Singh, G. Global Inequalities in Cervical Cancer Incidence and Mortality Are Linked to Deprivation, Low Socioeconomic Status, and Human Development. Int. J. MCH AIDS 2012, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Cibula, D.; Pötter, R.; Planchamp, F.; Avall-Lundqvist, E.; Fischerova, D.; Haie-Meder, C.; Köhler, C.; Landoni, F.; Lax, S.; Lindegaard, J.C.; et al. Correction to: The European Society of Gynaecological Oncology/European Society for Radiotherapy and Oncology/European Society of Pathology Guidelines for the Management of Patients with Cervical Cancer. Virchows Arch. 2018, 472, 937–938. [Google Scholar] [CrossRef]
- Girda, E.; Randall, L.M.; Chino, F.; Monk, B.J.; Farley, J.H.; O’Cearbhaill, R.E. Cervical Cancer Treatment Update: A Society of Gynecologic Oncology Clinical Practice Statement. Gynecol. Oncol. 2023, 179, 115–122. [Google Scholar] [CrossRef]
- Das, M.; Law, S. Role of Tumor Microenvironment in Cancer Stem Cell Chemoresistance and Recurrence. Int. J. Biochem Cell Biol. 2018, 103, 115–124. [Google Scholar] [CrossRef]
- Organista-Nava, J.; Gómez-Gómez, Y.; Garibay-Cerdenares, O.; Leyva-Vázquez, M.; Illades-Aguiar, B. Cervical Cancer Stem Cell-associated Genes: Prognostic Implications in Cervical Cancer. Oncol. Lett. 2019, 18, 7–14. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Zheng, P.-S. High Aldehyde Dehydrogenase Activity Identifies Cancer Stem Cells in Human Cervical Cancer. Oncotarget 2013, 4, 2462–2475. [Google Scholar] [CrossRef]
- Ortiz-Sánchez, E.; Santiago-López, L.; Cruz-Domínguez, V.B.; Toledo-Guzmán, M.E.; Hernández-Cueto, D.; Muñiz-Hernández, S.; Garrido, E.; De León, D.C.; García-Carrancá, A. Characterization of Cervical Cancer Stem Cell-like Cells: Phenotyping, Stemness, and Human Papilloma Virus Co-Receptor Expression. Oncotarget 2016, 7, 31943–31954. [Google Scholar] [CrossRef]
- López, J.; Poitevin, A.; Mendoza-Martínez, V.; Pérez-Plasencia, C.; García-Carrancá, A. Cancer-Initiating Cells Derived from Established Cervical Cell Lines Exhibit Stem-Cell Markers and Increased Radioresistance. BMC Cancer 2012, 12, 48. [Google Scholar] [CrossRef]
- Ammothumkandy, A.; Maliekal, T.T.; Bose, M.V.; Rajkumar, T.; Shirley, S.; Thejaswini, B.; Giri, V.G.; Krishna, S. CD66 and CD49f Expressing Cells Are Associated with Distinct Neoplastic Phenotypes and Progression in Human Cervical Cancer. Eur. J. Cancer 2016, 60, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Muraro, M.G.; Mele, V.; Däster, S.; Han, J.; Heberer, M.; Cesare Spagnoli, G.; Iezzi, G. CD133+, CD166+CD44+, and CD24+CD44+ Phenotypes Fail to Reliably Identify Cell Populations with Cancer Stem Cell Functional Features in Established Human Colorectal Cancer Cell Lines. Stem Cells Transl. Med. 2012, 1, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Z.; Lin, X.G.; Hua, P.; Wang, M.; Ao, X.; Xiong, L.H.; Wu, C.; Guo, J.J. The Study of the Tumor Stem Cell Properties of CD133+CD44+ Cells in the Human Lung Adenocarcinoma Cell Line A549. Cell. Mol. Biol. 2010, 56, 1350–1358. [Google Scholar]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Yang, K.; Xia, B.; Wang, W.; Cheng, J.; Yin, M.; Xie, H.; Li, J.; Ma, L.; Yang, C.; Li, A.; et al. A Comprehensive Analysis of Metabolomics and Transcriptomics in Cervical Cancer. Sci. Rep. 2017, 7, 43353. [Google Scholar] [CrossRef]
- Nuer-Allornuvor, G.F.; Alolga, R.N.; Liang, S.; Ling, Z.; Jingjing, W.; Xu, B.-Q.; Jiangli, Z.; Ennin, V.K.; Zhou, Z.; Ying, X. GC–MS-Based Untargeted Plasma Metabolomics Identifies a 2-Biomarker Panel for Possible Diagnosis of Precancerous Cervical Intraepithelial Neoplasia Stages from Cervical Cancer. Sci. Rep. 2024, 14, 17649. [Google Scholar] [CrossRef]
- Zhuo, D.; Li, X.; Guan, F. Biological Roles of Aberrantly Expressed Glycosphingolipids and Related Enzymes in Human Cancer Development and Progression. Front. Physiol. 2018, 9, 466. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Terreni, M.; Sollogoub, M.; Zhang, Y. Functional Role of Glycosphingolipids in Cancer. Curr. Med. Chem. 2021, 28, 3913–3924. [Google Scholar] [CrossRef]
- Krengel, U.; Bousquet, P.A. Molecular Recognition of Gangliosides and Their Potential for Cancer Immunotherapies. Front. Immunol. 2014, 5, 325. [Google Scholar] [CrossRef]
- Reynisson, J.; Court, W.; O’Neill, C.; Day, J.; Patterson, L.; McDonald, E.; Workman, P.; Katan, M.; Eccles, S.A. The Identification of Novel PLC-γ Inhibitors Using Virtual High Throughput Screening. Bioorg. Med. Chem. 2009, 17, 3169–3176. [Google Scholar] [CrossRef]
- Odak, Z.; Marijan, S.; Radan, M.; Pilkington, L.I.; Čikeš Botić, M.; Barker, D.; Reynisson, J.; Leung, E.; Čikeš Čulić, V. Deciphering the Interplay: Thieno[2,3-b]Pyridine’s Impact on Glycosphingolipid Expression, Cytotoxicity, Apoptosis, and Metabolomics in Ovarian Tumor Cell Lines. Int. J. Mol. Sci. 2024, 25, 6954. [Google Scholar] [CrossRef] [PubMed]
- Marijan, S.; Markotić, A.; Mastelić, A.; Režić-Mužinić, N.; Pilkington, L.I.; Reynisson, J.; Čulić, V.Č. Glycosphingolipid Expression at Breast Cancer Stem Cells after Novel Thieno[2,3-b]Pyridine Anticancer Compound Treatment. Sci. Rep. 2020, 10, 11876. [Google Scholar] [CrossRef] [PubMed]
- Haverkate, N.A.; Leung, E.; Pilkington, L.I.; Barker, D. Tethered Aryl Groups Increase the Activity of Anti-Proliferative Thieno[2,3-b]Pyridines by Targeting a Lipophilic Region in the Active Site of PI-PLC. Pharmaceutics 2021, 13, 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhu, X.; Deng, S.; Chen, Q.; Zeng, X. Synthesis and Cytotoxic Effects on HeLa Cervical Cancer Cells of (E)-2-Aroyl-4-(4-Fluorobenzylidene)-5-Oxopyrrolidine. J. Chem. Res. 2020, 44, 576–579. [Google Scholar] [CrossRef]
- Roy, M.; Mukherjee, S. Reversal of Resistance towards Cisplatin by Curcumin in Cervical Cancer Cells. Asian Pac. J. Cancer Prev. 2014, 15, 1403–1410. [Google Scholar] [CrossRef]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN) Nomenclature of Glycolipids: Recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [CrossRef]
- Svennerholm, L. Chromatographic Separation of Human Brain Gangliosides. J. Neurochem. 1963, 10, 613–623. [Google Scholar] [CrossRef]
- Global Strategy to Accelerate the Elimination of Cervical Cancer as a Public Health Problem. Available online: https://www.who.int/publications/i/item/9789240014107 (accessed on 28 August 2024).
- Brisson, M.; Kim, J.J.; Canfell, K.; Drolet, M.; Gingras, G.; Burger, E.A.; Martin, D.; Simms, K.T.; Bénard, É.; Boily, M.-C.; et al. Impact of HPV Vaccination and Cervical Screening on Cervical Cancer Elimination: A Comparative Modelling Analysis in 78 Low-Income and Lower-Middle-Income Countries. Lancet 2020, 395, 575–590. [Google Scholar] [CrossRef]
- Poolkerd, S.; Leelahakorn, S.; Manusirivithaya, S.; Tangjitgamol, S.; Thavaramara, T.; Sukwattana, P.; Pataradule, K. Survival Rate of Recurrent Cervical Cancer Patients. J. Med. Assoc. Thai. 2006, 89, 275–282. [Google Scholar]
- Holthaus, D.; Rogmans, C.; Gursinski, I.; Quevedo-Olmos, A.; Ehsani, M.; Mangler, M.; Flörkemeier, I.; Weimer, J.P.; Meyer, T.F.; Maass, N.; et al. Inhibition of ADAM17 Increases the Cytotoxic Effect of Cisplatin in Cervical Spheroids and Organoids. Front. Oncol. 2024, 14, 1432239. [Google Scholar] [CrossRef]
- Takeuchi, S. Biology and Treatment of Cervical Adenocarcinoma. Chin. J. Cancer Res. 2016, 28, 254–262. [Google Scholar] [CrossRef]
- Yang, H.; Huang, S.G.; Dong, M.; Wang, X.; He, J.; Su, H.; Liu, C.; Zhu, Y.; Wei, L.; Liu, Z. Efficacy and Safety of Bevacizumab in Neoadjuvant and Concurrent Chemoradiotherapy for Refractory Cervical Cancer Patients. Biomol. Biomed. 2024, 24, 1586. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, B.; Iddawela, M. Bevacizumab in Advanced Cervical Cancer: Issues and Challenges for Low- and Middle-Income Countries. J. Glob. Oncol. 2017, 3, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Pervan, M.; Marijan, S.; Markotić, A.; Pilkington, L.I.; Haverkate, N.A.; Barker, D.; Reynisson, J.; Meić, L.; Radan, M.; Čikeš Čulić, V. Novel Thieno [2,3-b]Pyridine Anticancer Compound Lowers Cancer Stem Cell Fraction Inducing Shift of Lipid to Glucose Metabolism. Int. J. Mol. Sci. 2022, 23, 11457. [Google Scholar] [CrossRef] [PubMed]
- Mastelic, A.; Cikes Culic, V.; Rezic Muzinic, N.; Vuica-Ross, M.; Barker, D.; Leung, E.; Reynisson, J.; Markotic, A. Glycophenotype of Breast and Prostate Cancer Stem Cells Treated with Thieno[2,3-b]Pyridine Anticancer Compound. Drug Des. Devel. Ther. 2017, 11, 759–769. [Google Scholar] [CrossRef]
- Shoemaker, R.H. The NCI60 Human Tumour Cell Line Anticancer Drug Screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef]
- Arabshahi, H.J.; Leung, E.; Barker, D.; Reynisson, J. The Development of Thieno[2,3-b]Pyridine Analogues as Anticancer Agents Applying in Silico Methods. Med. Chem. Commun. 2014, 5, 186. [Google Scholar] [CrossRef]
- Reynisson, J.; Jaiswal, J.K.; Barker, D.; D’mello, S.A.N.; Denny, W.A.; Baguley, B.C.; Leung, E.Y. Evidence That Phospholipase C Is Involved in the Antitumour Action of NSC768313, a New Thieno[2,3-b]Pyridine Derivative. Cancer Cell Int 2016, 16, 18. [Google Scholar] [CrossRef]
- Wang, J.; Luo, C.; Shan, C.; You, Q.; Lu, J.; Elf, S.; Zhou, Y.; Wen, Y.; Vinkenborg, J.L.; Fan, J.; et al. Inhibition of Human Copper Trafficking by a Small Molecule Significantly Attenuates Cancer Cell Proliferation. Nature Chem 2015, 7, 968–979. [Google Scholar] [CrossRef]
- Arabshahi, H.J.; Van Rensburg, M.; Pilkington, L.I.; Jeon, C.Y.; Song, M.; Gridel, L.-M.; Leung, E.; Barker, D.; Vuica-Ross, M.; Volcho, K.P.; et al. A Synthesis, in Silico, in Vitro and in Vivo Study of Thieno[2,3-b]Pyridine Anticancer Analogues. Med. Chem. Commun. 2015, 6, 1987–1997. [Google Scholar] [CrossRef]
- Romagnoli, R.; Baraldi, P.G.; Kimatrai Salvador, M.; Preti, D.; Aghazadeh Tabrizi, M.; Bassetto, M.; Brancale, A.; Hamel, E.; Castagliuolo, I.; Bortolozzi, R.; et al. Synthesis and Biological Evaluation of 2-(Alkoxycarbonyl)-3-Anilinobenzo[b]Thiophenes and Thieno[2,3-b]Pyridines as New Potent Anticancer Agents. J. Med. Chem. 2013, 56, 2606–2618. [Google Scholar] [CrossRef] [PubMed]
- Eurtivong, C.; Semenov, V.; Semenova, M.; Konyushkin, L.; Atamanenko, O.; Reynisson, J.; Kiselyov, A. 3-Amino-Thieno[2,3-b]Pyridines as Microtubule-Destabilising Agents: Molecular Modelling and Biological Evaluation in the Sea Urchin Embryo and Human Cancer Cells. Bioorganic Med. Chem. 2017, 25, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Jaakola, V.-P.; Lane, J.R.; Lin, J.; IJzerman, A.P.; Yeager, M.; Kufareva, I.; Stevens, R.C.; Abagyan, R. Structure-Based Discovery of Novel Chemotypes for Adenosine A2A Receptor Antagonists. J. Med. Chem. 2010, 53, 1799–1809. [Google Scholar] [CrossRef] [PubMed]
- Proschak, E.; Stark, H.; Merk, D. Polypharmacology by Design: A Medicinal Chemist’s Perspective on Multitargeting Compounds. J. Med. Chem. 2019, 62, 420–444. [Google Scholar] [CrossRef]
- Patel, V.A.; Longacre, A.; Hsiao, K.; Fan, H.; Meng, F.; Mitchell, J.E.; Rauch, J.; Ucker, D.S.; Levine, J.S. Apoptotic Cells, at All Stages of the Death Process, Trigger Characteristic Signaling Events That Are Divergent from and Dominant over Those Triggered by Necrotic Cells: Implications for the Delayed Clearance Model of Autoimmunity. J. Biol. Chem. 2006, 281, 4663–4670. [Google Scholar] [CrossRef]
- Gutiérrez, G.; Mendoza, C.; Montaño, L.F.; López-Marure, R. Ceramide Induces Early and Late Apoptosis in Human Papilloma Virus+ Cervical Cancer Cells by Inhibiting Reactive Oxygen Species Decay, Diminishing the Intracellular Concentration of Glutathione and Increasing Nuclear Factor-κB Translocation. Anticancer Drugs 2007, 18, 149–159. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Why Therapeutic Response May Not Prolong the Life of a Cancer Patient: Selection for Oncogenic Resistance. Cell Cycle 2005, 4, 1693–1698. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem Cells, Cancer, and Cancer Stem Cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef]
- Di Fiore, R.; Suleiman, S.; Drago-Ferrante, R.; Subbannayya, Y.; Pentimalli, F.; Giordano, A.; Calleja-Agius, J. Cancer Stem Cells and Their Possible Implications in Cervical Cancer: A Short Review. Int. J. Mol. Sci. 2022, 23, 5167. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Yu, R.K. The Expression and Functions of Glycoconjugates in Neural Stem Cells. Glycobiology 2007, 17, 57R–74R. [Google Scholar] [CrossRef]
- Lopez, P.H.; Schnaar, R.L. Gangliosides in Cell Recognition and Membrane Protein Regulation. Curr. Opin. Struct. Biol. 2009, 19, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Fredman, P.; Hedberg, K.; Brezicka, T. Gangliosides as Therapeutic Targets for Cancer. BioDrugs. 2003, 17, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Kabayama, K.; Sato, T.; Saito, K.; Loberto, N.; Prinetti, A.; Sonnino, S.; Kinjo, M.; Igarashi, Y.; Inokuchi, J. Dissociation of the Insulin Receptor and Caveolin-1 Complex by Ganglioside GM3 in the State of Insulin Resistance. Proc. Natl. Acad. Sci. USA 2007, 104, 13678–13683. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.G.; Schlessinger, J.; Hakomori, S. Ganglioside-Mediated Modulation of Cell Growth. Specific Effects of GM3 on Tyrosine Phosphorylation of the Epidermal Growth Factor Receptor. J. Biol. Chem. 1986, 261, 2434–2440. [Google Scholar] [CrossRef]
- Miljan, E.A.; Meuillet, E.J.; Mania-Farnell, B.; George, D.; Yamamoto, H.; Simon, H.-G.; Bremer, E.G. Interaction of the Extracellular Domain of the Epidermal Growth Factor Receptor with Gangliosides. J. Biol. Chem. 2002, 277, 10108–10113. [Google Scholar] [CrossRef]
- Yoshida, S.; Fukumoto, S.; Kawaguchi, H.; Sato, S.; Ueda, R.; Furukawa, K. Ganglioside G(D2) in Small Cell Lung Cancer Cell Lines: Enhancement of Cell Proliferation and Mediation of Apoptosis. Cancer Res. 2001, 61, 4244–4252. [Google Scholar]
- Liang, Y.-J.; Ding, Y.; Levery, S.B.; Lobaton, M.; Handa, K.; Hakomori, S. Differential Expression Profiles of Glycosphingolipids in Human Breast Cancer Stem Cells vs. Cancer Non-Stem Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 4968–4973. [Google Scholar] [CrossRef]
- Patel, S.; Ahmed, S. Emerging Field of Metabolomics: Big Promise for Cancer Biomarker Identification and Drug Discovery. J. Pharm. Biomed. Anal. 2015, 107, 63–74. [Google Scholar] [CrossRef]
- Xue, L.-L.; Chen, H.-H.; Jiang, J.-G. Implications of Glycerol Metabolism for Lipid Production. Prog. Lipid Res. 2017, 68, 12–25. [Google Scholar] [CrossRef]
- Zweytick, D.; Athenstaedt, K.; Daum, G. Intracellular Lipid Particles of Eukaryotic Cells. Biochim. Biophys. Acta 2000, 1469, 101–120. [Google Scholar] [CrossRef]
- Bell, R.M.; Coleman, R.A. Enzymes of Glycerolipid Synthesis in Eukaryotes. Annu. Rev. Biochem. 1980, 49, 459–487. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.-G.; Filipescu, A.; Vladareanu, R.; Petca, A. The Importance of Metabolic Factors in Endometrial Cancer: Evaluating the Utility of the Triglyceride-to-Glycemia Index and Triglyceride-to-High-Density Lipoprotein Ratio As Biomarkers. Cureus 2024, 16, e62099. [Google Scholar] [CrossRef] [PubMed]
- Rosato, V.; Zucchetto, A.; Bosetti, C.; Dal Maso, L.; Montella, M.; Pelucchi, C.; Negri, E.; Franceschi, S.; La Vecchia, C. Metabolic Syndrome and Endometrial Cancer Risk. Ann. Oncol. 2011, 22, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Zhao, J.; Li, W.; Wang, X.; Gao, Y.; Wang, Z.; Hu, S.; Cai, J. Hypertension and Hyperglycaemia Are Positively Correlated with Local Invasion of Early Cervical Cancer. Front. Endocrinol. 2023, 14, 1280060. [Google Scholar] [CrossRef]
- Shen, T.; Sun, S.; Li, W.; Wang, X.; Gao, Y.; Yang, Q.; Cai, J. Association between Body Mass Index and Lymph Node Metastasis among Women with Cervical Cancer: A Systematic Review and Network Meta-Analysis. Arch. Gynecol. Obstet. 2024, 310, 1289–1301. [Google Scholar] [CrossRef]
- Atilla-Gokcumen, G.E.; Muro, E.; Relat-Goberna, J.; Sasse, S.; Bedigian, A.; Coughlin, M.L.; Garcia-Manyes, S.; Eggert, U.S. Dividing Cells Regulate Their Lipid Composition and Localization. Cell 2014, 156, 428–439. [Google Scholar] [CrossRef]
- Li, N.; Sancak, Y.; Frasor, J.; Atilla-Gokcumen, G.E. A Protective Role for Triacylglycerols during Apoptosis. Biochemistry 2018, 57, 72–80. [Google Scholar] [CrossRef]
- Pappa, K.I.; Daskalakis, G.; Anagnou, N.P. Metabolic Rewiring Is Associated with HPV-Specific Profiles in Cervical Cancer Cell Lines. Sci. Rep. 2021, 11, 17718. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of Plasmalogen Lipids in Health and Disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Meisen, I.; Peter-Katalinić, J.; Müthing, J. Direct Analysis of Silica Gel Extracts from Immunostained Glycosphingolipids by Nanoelectrospray Ionization Quadrupole Time-of-Flight Mass Spectrometry. Anal. Chem. 2004, 76, 2248–2255. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolite | HeLa | SiHa | ||
|---|---|---|---|---|---|
| p-Value | Fold Change | p-Value | Fold Change | ||
| 1 | Fructose | 0.0933 | 8.6667 | 0.1157 | 0.36235 |
| 2 | Erythritol | 0.1157 | 2.5974 | - | - |
| 3 | Glycerol | 0.0439 * | 2.177 | - | - |
| 4 | Myristic acid | 0.3537 | 2.0952 | 0.2924 | 0.68703 |
| 5 | Nonadecane | 0.7063 | 1.3947 | - | - |
| 6 | 1-Acylglycerol | 0.4000 | 1.2654 | 0.1458 | 0.51757 |
| 7 | 2-Stearoylglycerophosphoglycerol | 0.8888 | 1.2517 | 1.000 | 0.79479 |
| 8 | Xylose | 0.3941 | 1.213 | - | - |
| 9 | Stearic acid | 0.2794 | 1.1812 | 0.4050 | 0.71185 |
| 10 | Octadecanol | 0.1409 | 1.172 | 0.0003 * | 0.7916 |
| 11 | Pyruvic acid | 0.2835 | 1.1719 | - | - |
| 12 | Butyric acid | 0.8099 | 1.1667 | - | - |
| 13 | Myo inositol | 0.6579 | 1.1543 | 0.5506 | 1.0463 |
| 14 | Behenic acid | 0.1401 | 1.1304 | 0.1642 | 0.70783 |
| 15 | Dibutyl phthalate | 0.2612 | 1.1286 | 0.9625 | 0.99585 |
| 16 | Palmitic acid | 0.1696 | 1.1197 | 0.1150 | 1.2907 |
| 17 | Lactic acid | 0.5214 | 0.91443 | 0.8880 | 1.0147 |
| 18 | Mannitol | 0.6779 | 1.0656 | - | - |
| 19 | Galactose | 0.6995 | 1.0627 | 0.0953 | 1.1795 |
| 20 | Succinic acid | 0.4778 | 1.0625 | - | - |
| 21 | Cholesterol | 0.7865 | 0.94372 | 0.1637 | 0.6011 |
| 22 | L-Proline | 0.6972 | 1.0479 | 0.1044 | 1.4 |
| 23 | Phosphate | 0.7821 | 1.0199 | 0.2566 | 1.2162 |
| 24 | Glucose | - | - | 0.1841 | 1.336 |
| 25 | Oleic acid | - | - | 0.6295 | 1.3 |
| 26 | Inositol | - | - | 1.000 | 0.79592 |
| 27 | L-Glutamic acid | - | - | 0.7788 | 0.84283 |
| 28 | 3-Hydroxybutyric acid | - | - | 0.7458 | 1.064 |
| 29 | Hydrocinnamic acid | - | - | 0.8995 | 0.92891 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čikeš Botić, M.; Marijan, S.; Radan, M.; Novak, I.; Glumac, M.; Pilkington, L.I.; Odak, Z.; Barker, D.; Reynisson, J.; Čikeš Čulić, V. Thieno[2,3-b]pyridines as a Novel Strategy Against Cervical Cancer: Mechanistic Insights and Therapeutic Potential. Int. J. Mol. Sci. 2025, 26, 2651. https://doi.org/10.3390/ijms26062651
Čikeš Botić M, Marijan S, Radan M, Novak I, Glumac M, Pilkington LI, Odak Z, Barker D, Reynisson J, Čikeš Čulić V. Thieno[2,3-b]pyridines as a Novel Strategy Against Cervical Cancer: Mechanistic Insights and Therapeutic Potential. International Journal of Molecular Sciences. 2025; 26(6):2651. https://doi.org/10.3390/ijms26062651
Chicago/Turabian StyleČikeš Botić, Monika, Sandra Marijan, Mila Radan, Ivana Novak, Mateo Glumac, Lisa I. Pilkington, Zdravko Odak, David Barker, Jóhannes Reynisson, and Vedrana Čikeš Čulić. 2025. "Thieno[2,3-b]pyridines as a Novel Strategy Against Cervical Cancer: Mechanistic Insights and Therapeutic Potential" International Journal of Molecular Sciences 26, no. 6: 2651. https://doi.org/10.3390/ijms26062651
APA StyleČikeš Botić, M., Marijan, S., Radan, M., Novak, I., Glumac, M., Pilkington, L. I., Odak, Z., Barker, D., Reynisson, J., & Čikeš Čulić, V. (2025). Thieno[2,3-b]pyridines as a Novel Strategy Against Cervical Cancer: Mechanistic Insights and Therapeutic Potential. International Journal of Molecular Sciences, 26(6), 2651. https://doi.org/10.3390/ijms26062651