
A Novel Peptide-Based Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Neutralizing Antibodies Against NADC30-like PRRSV GP5 Protein


 , ,
, , 
Abstract
1. Introduction
2. Results
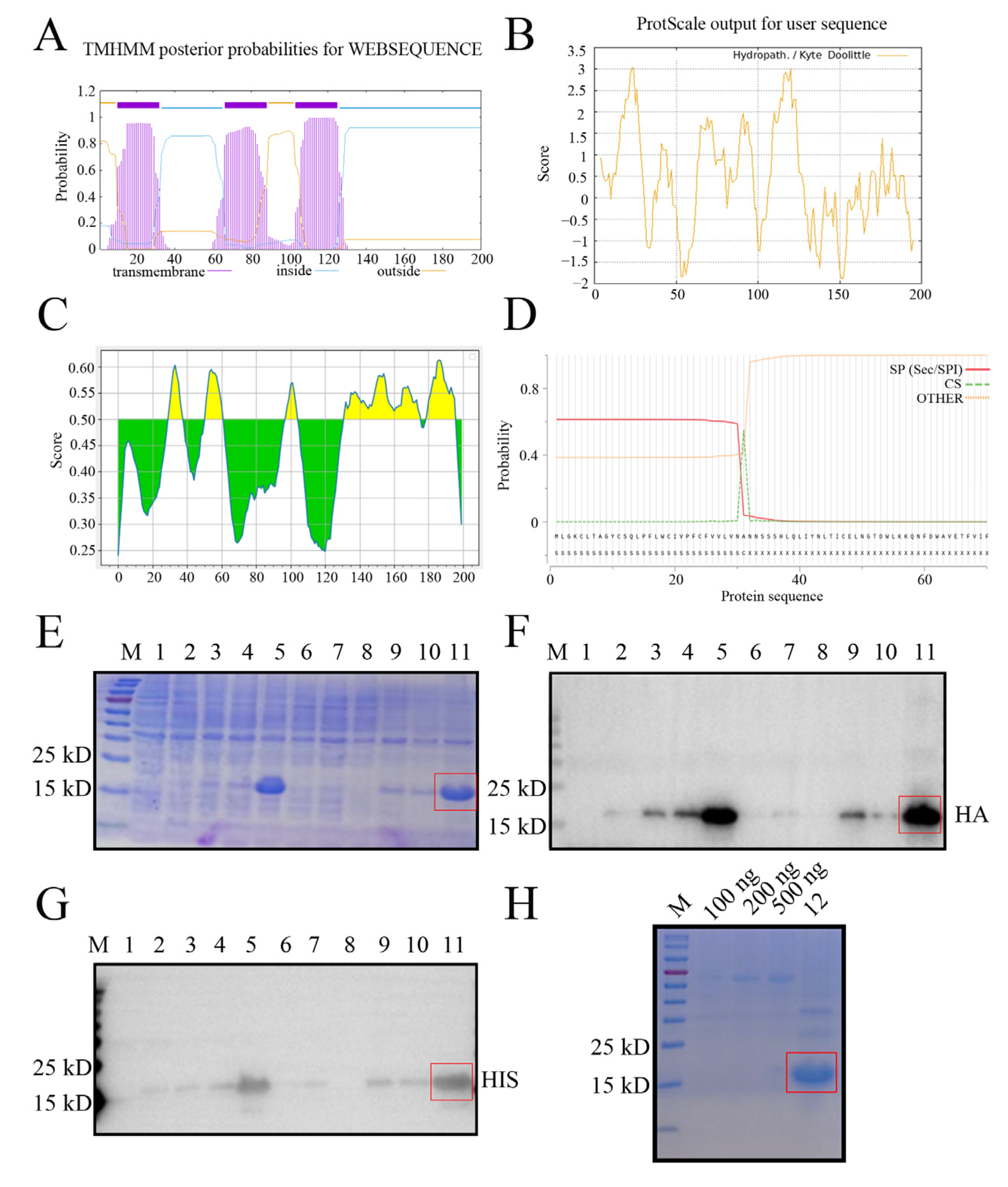
2.1. Expression and Purification of the Recombinant NADC30-like PRRSV GP5 Truncated Protein
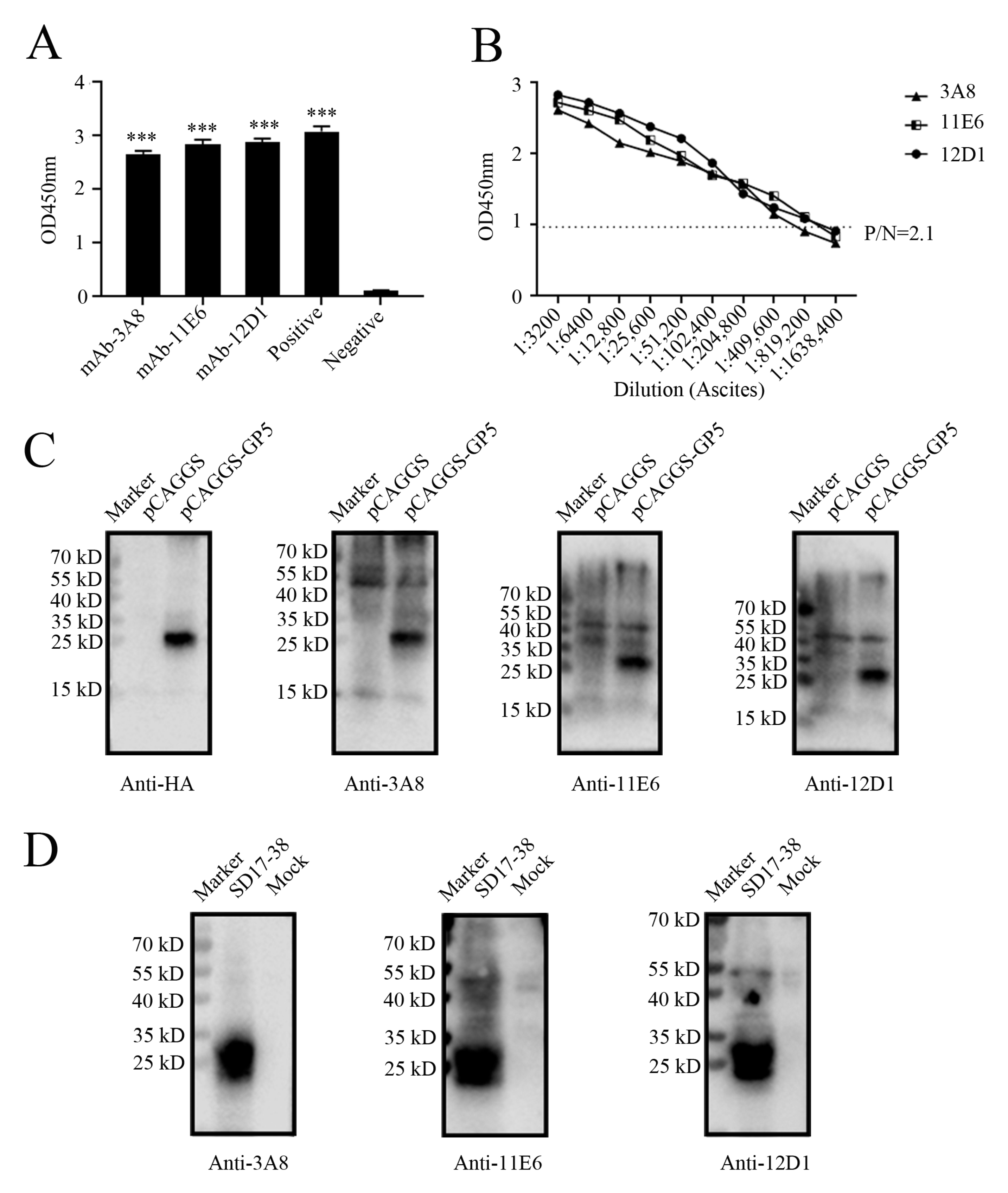
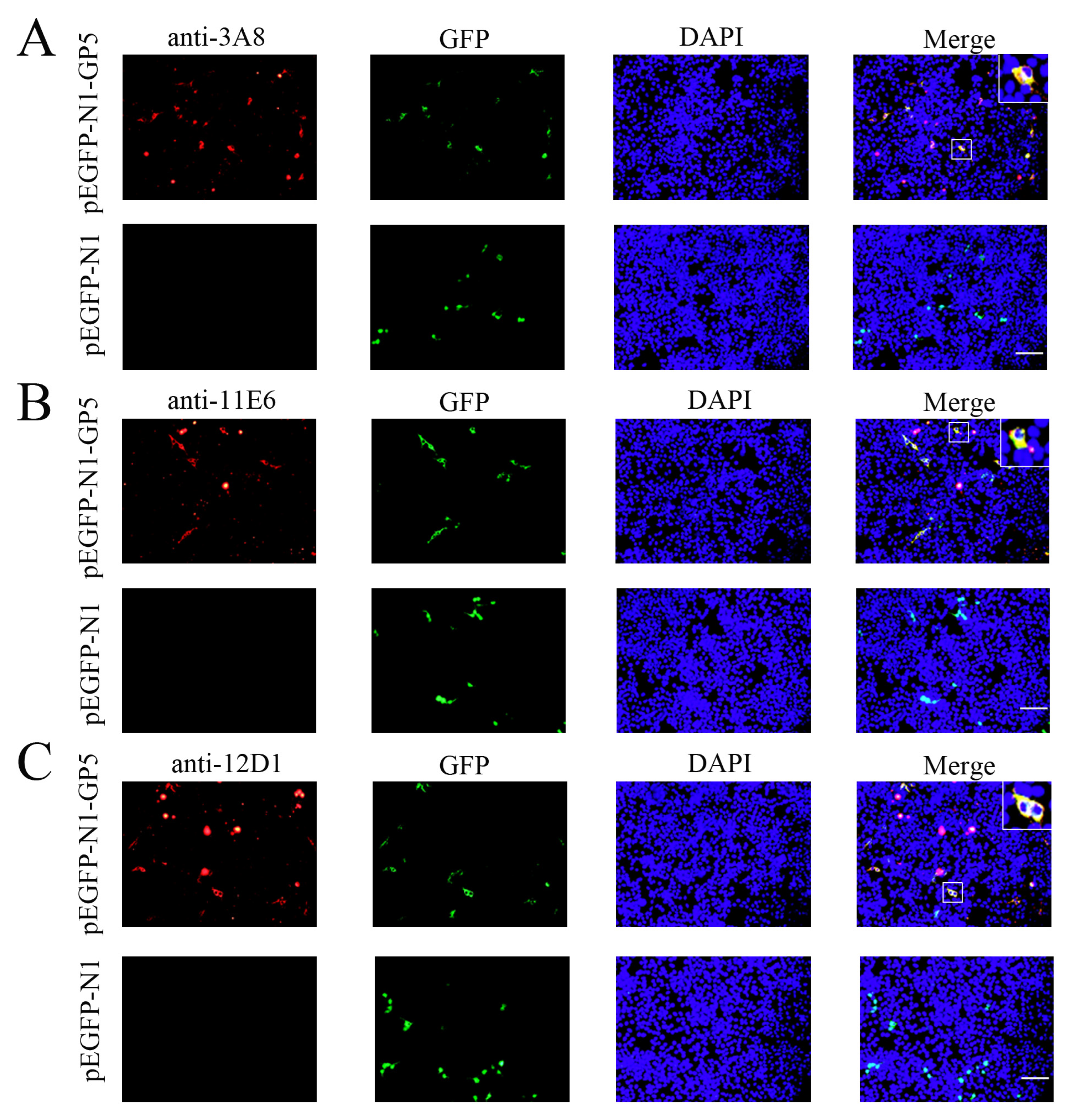
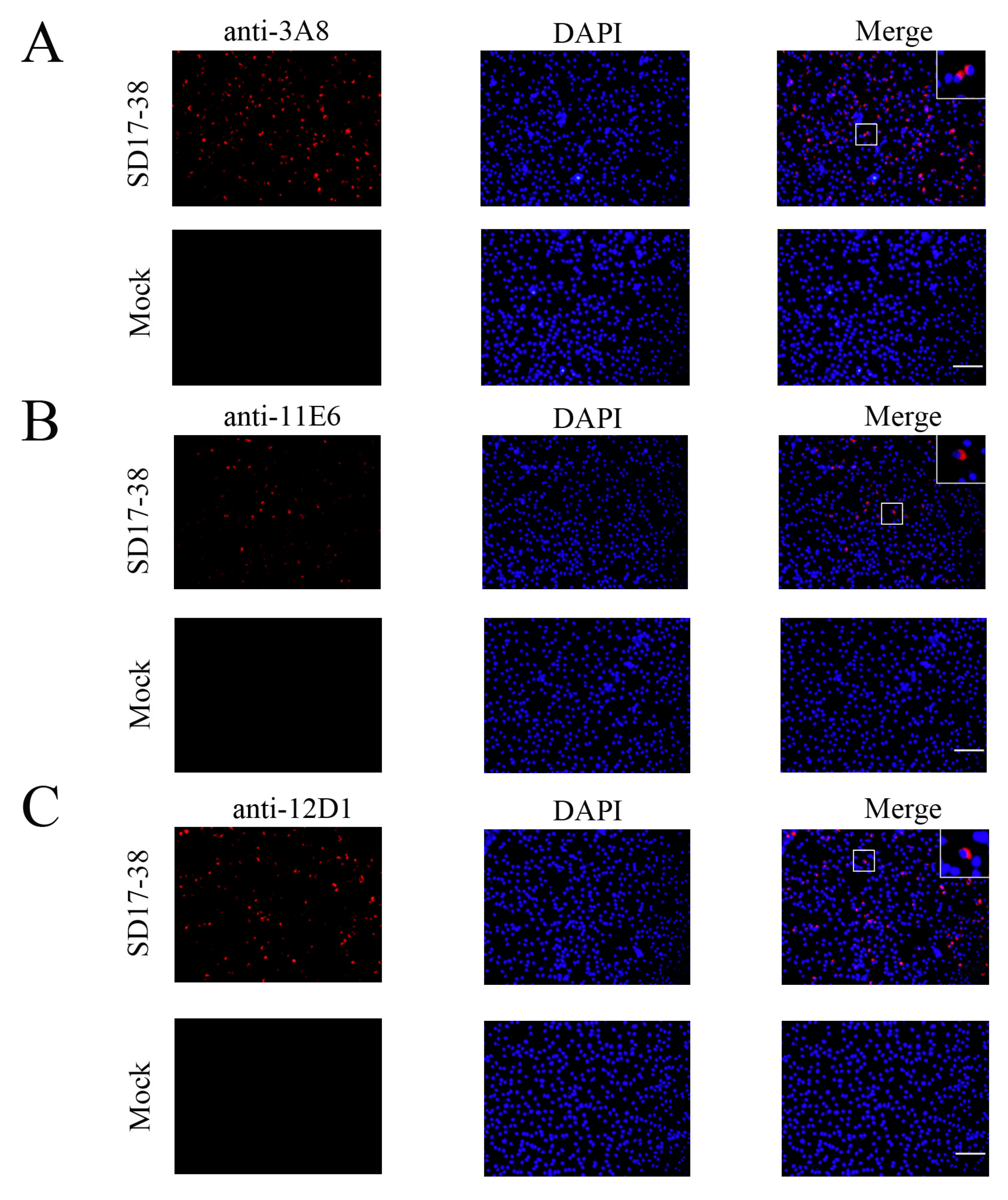
2.2. Preparation and Identification of GP5 Protein Specific mAbs
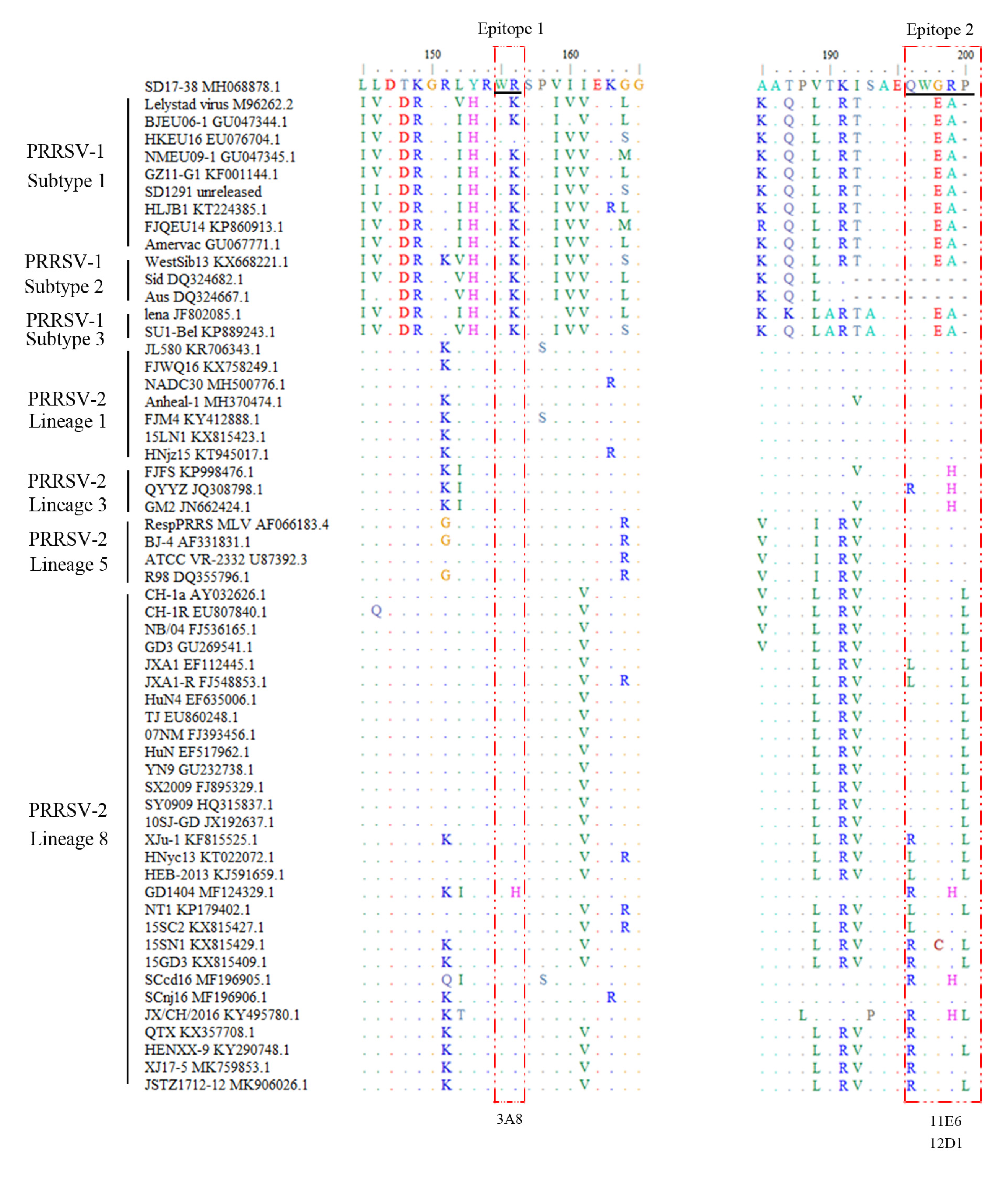
2.3. Precise Mapping and Identification of Linear B Cell Epitopes Recognized by GP5 mAbs
2.4. Conservation Analysis of the Identified Antigenic Epitopes Across Different PRRSV Strains
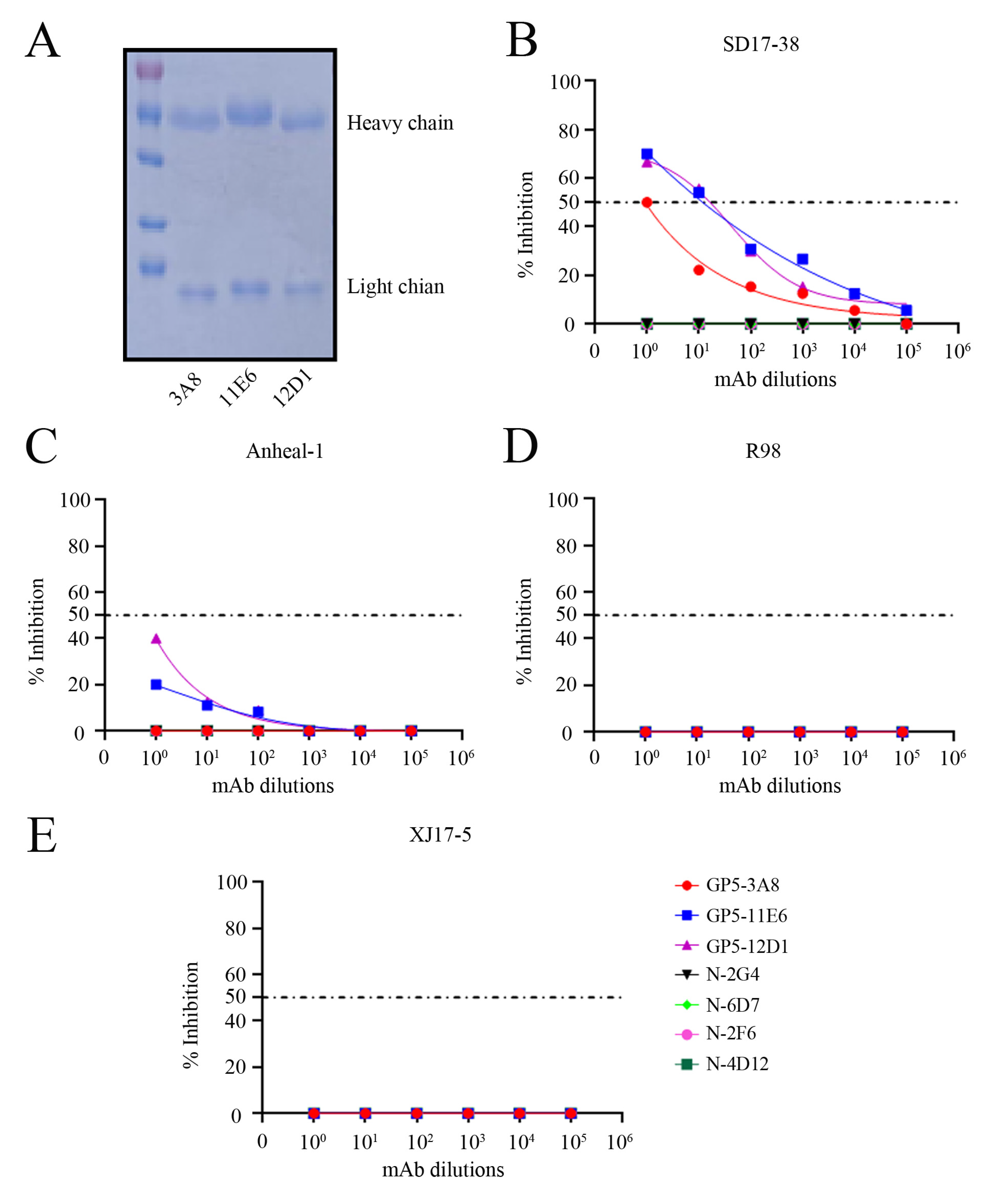
2.5. Neutralizing Activity of Three GP5 mAbs and Development of the Peptide-Based ELISA Detecting PRRSV Antibodies
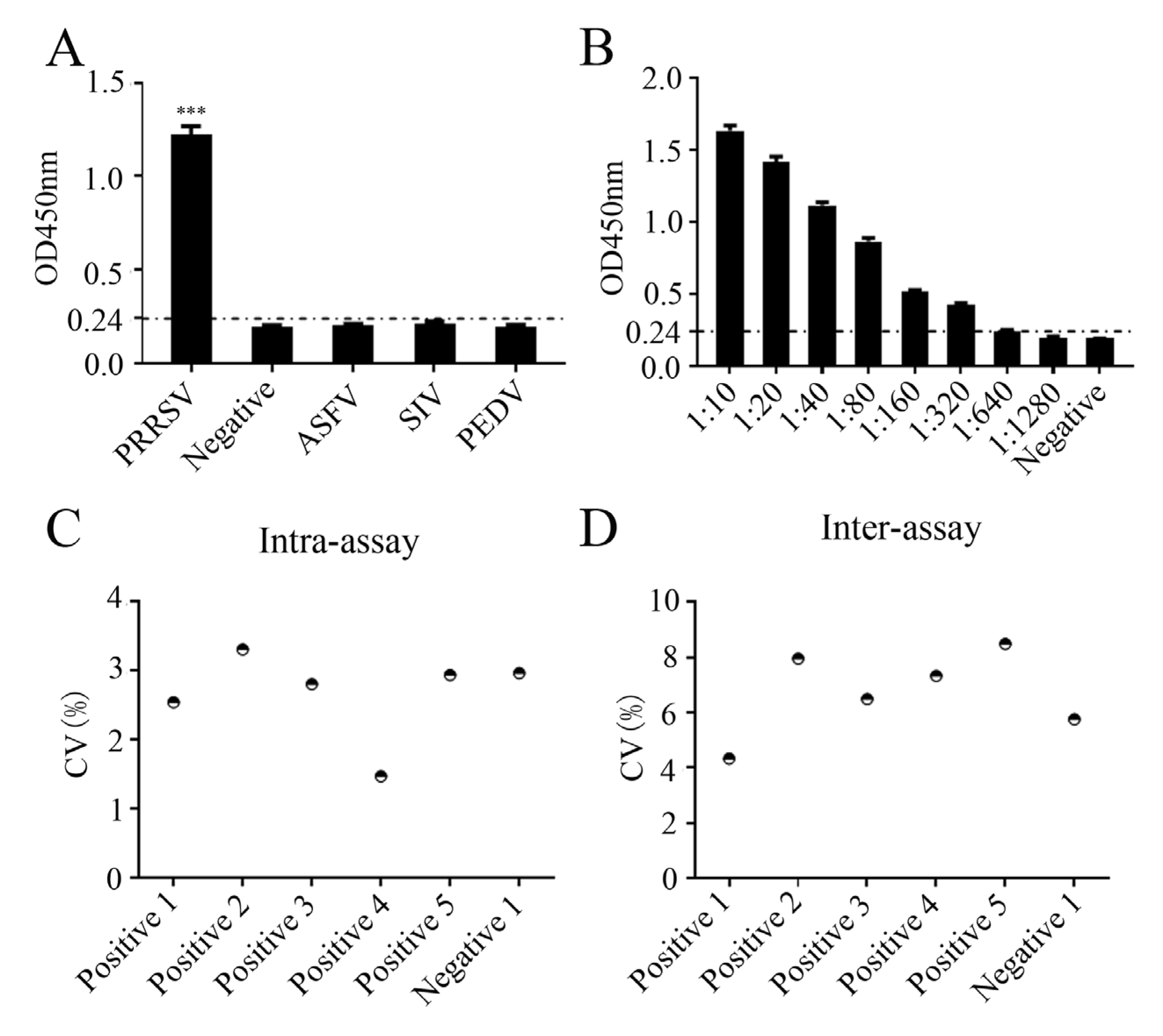
2.6. Specificity, Sensitivity, and Repeatability of the Peptide-Based ELISA
2.7. Applicability of the Peptide-Based ELISA for Detection of Clinical Samples
3. Discussion
4. Materials and Methods
4.1. Cells, Viruses, and Mice
4.2. Bioinformatics Analysis of the GP5 Protein and Construction of the Recombinant Plasmids
4.3. Expression and Purification of the Recombinant GP5 Truncation Protein
4.4. Immunization of the Mice and Monoclonal Antibody (mAb) Production and Purification
4.5. GP5 Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Western Blotting (WB)
4.7. Immunofluorescence Assay (IFA)
4.8. Precise Identification of the mAb Recognizing Epitopes and the Epitope Conservation Analysis
4.9. Virus Neutralization Test by the mAbs
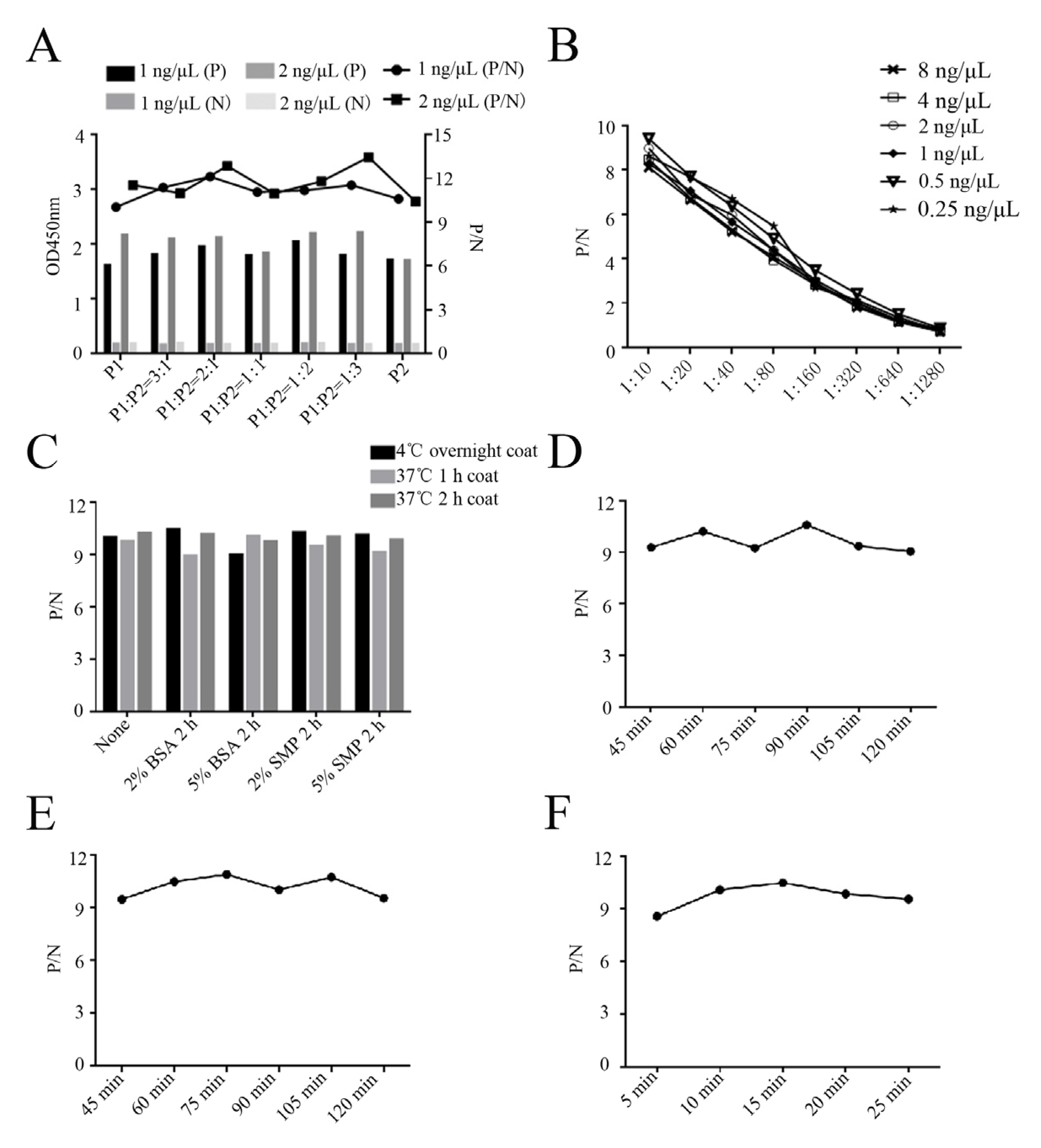
4.10. Development and Optimization of GP5 Peptide-Based ELISA
4.11. Cut-Off Values, Specificity, and Sensitivity of the Peptide-Based ELISA
4.12. Repeatability and Application of the Peptide-Based ELISA
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Brinton, M.A.; Gulyaeva, A.A.; Balasuriya, U.B.R.; Dunowska, M.; Faaberg, K.S.; Goldberg, T.; Leung, F.C.C.; Nauwynck, H.J.; Snijder, E.J.; Stadejek, T.; et al. ICTV Virus Taxonomy Profile: 2021. J. Gen. Virol. 2021, 102, 001632. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.H.; Chen, X.X.; Li, R.; Qiao, S.L.; Zhang, G.P. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Q.; Cao, Z.; Tang, Y.D.; Xia, D.; Wang, G.; Shan, H. Recent Advances in Porcine Reproductive and Respiratory Syndrome Virus NADC30-Like Research in China: Molecular Characterization, Pathogenicity, and Control. Front. Microbiol. 2021, 12, 791313. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef]
- Han, M.; Yoo, D. Engineering the PRRS virus genome: Updates and perspectives. Vet. Microbiol. 2014, 174, 279–295. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef]
- Luo, Q.; Zheng, Y.; Zhang, H.; Yang, Z.; Sha, H.; Kong, W.; Zhao, M.; Wang, N. Research Progress on Glycoprotein 5 of Porcine Reproductive and Respiratory Syndrome Virus. Animals 2023, 13, 813. [Google Scholar] [CrossRef]
- Johnson, C.R.; Griggs, T.F.; Gnanandarajah, J.; Murtaugh, M.P. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J. Gen. Virol. 2011, 92, 1107–1116. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Z.W.; Xu, T.; Zhou, Y.C.; Li, J.L.; Deng, H.D.; Li, F.Q.; Xu, L.; Sun, X.A.; Zhu, L. Molecular Characterization of the Nsp2 and ORF5s of PRRSV Strains in Sichuan China during 2012–2020. Animals 2022, 12, 3309. [Google Scholar] [CrossRef] [PubMed]
- Do, V.T.; Dao, H.T.; Hahn, T.W. Generation of a cold-adapted PRRSV with a nucleotide substitution in the ORF5 and numerous mutations in the hypervariable region of NSP2. J. Vet. Sci. 2020, 21, e85. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.W.; Liu, X.; Bai, J.; Gao, Y.N.; Song, Z.B.; Nauwynck, H.; Wang, X.W.; Yang, Y.Q.; Jiang, P. Glyceraldehyde-3-Phosphate Dehydrogenase Restricted in Cytoplasmic Location by Viral GP5 Facilitates Porcine Reproductive and Respiratory Syndrome Virus Replication via Its Glycolytic Activity. J. Virol. 2021, 95, e0021021. [Google Scholar] [CrossRef]
- Popescu, L.N.; Trible, B.R.; Chen, N.H.; Rowland, R.R.R. GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef]
- Pan, J.; Zeng, M.; Zhao, M.; Huang, L. Research Progress on the detection methods of porcine reproductive and respiratory syndrome virus. Front. Microbiol. 2023, 14, 1097905. [Google Scholar] [CrossRef]
- Lopez, O.J.; Osorio, F.A. Role of neutralizing antibodies in PRRSV protective immunity. Vet. Immunol. Immunopathol. 2004, 102, 155–163. [Google Scholar] [CrossRef]
- Su, J.; Zhou, L.; He, B.; Zhang, X.; Ge, X.; Han, J.; Guo, X.; Yang, H. Nsp2 and GP5-M of Porcine Reproductive and Respiratory Syndrome Virus Contribute to Targets for Neutralizing Antibodies. Virol. Sin. 2019, 34, 631–640. [Google Scholar] [CrossRef]
- Li, Z.J.; Ti, J.F.; Li, F.; Wang, Z.Y.; Diao, Y.R. Expression of GP5 protein of porcine reproductive and respiratory syndrome virus and establishment of indirect ELISA for antibody detection. Chin. J. Prev. Vet. Med. 2018, 40, 24–28. [Google Scholar]
- Hou, H.F.; Liu, X.W.; Bai, J.; Song, Z.B.; Jiang, P. Establishment and application of indirect ELISA for detection of GP5 protein antibody of porcine reproductive and respiratory syndrome virus. Chin. Vet. Sci. 2018, 48, 1086–1092. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, H.; He, J.H.; Wu, J.Y.; Shang, Y.J.; Liu, X.T. Indirect ELISA with recombinant GP5 for detecting antibodies to porcine reproductive and respiratory syndrome virus. Virol. Sin. 2011, 26, 61–66. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, K.; Zhang, J.; He, P.; Zhang, P.; Deng, D.; Chi, C.; Jiang, S.; Zheng, W.; Chen, N.; et al. A nucleocapsid monoclonal antibody based sandwich ELISA for the general detection of both PRRSV-2 and PRRSV-1. Vet. Microbiol. 2025, 302, 110399. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Gao, X.; Wu, Y.; Wang, F.; Lai, M.; Zheng, J.; Qiu, Y.; He, Y.; Liang, X.; Yuan, K.; et al. Genomic and pathogenicity analysis of two novel highly pathogenic recombinant NADC30-like PRRSV strains in China, in 2023. Microbiol. Spectr. 2024, 12, e0036824. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.Y.; Chang, T.; Wang, D.C.; Zhang, H.L.; Liu, H.Z.; Huang, X.Y.; Tian, Z.J.; Tian, X.X.; Liu, D.; An, T.Q.; et al. Genomic surveillance and evolutionary dynamics of type 2 porcine reproductive and respiratory syndrome virus in China spanning the African swine fever outbreak. Virus Evol. 2024, 10, veae016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Feng, Y.; Martin, D.P.; Chen, J.; Ma, S.; Xia, P.; Zhang, G. Genetic diversity and phylogenetic analysis of the ORF5 gene of PRRSV from central China. Res. Vet. Sci. 2017, 115, 226–234. [Google Scholar] [CrossRef]
- Tian, H.; Cheng, Y.; Wu, J.Y.; He, J.H.; Shang, Y.J.; Liu, X.T. Application of GP5 protein to develop monoclonal antibody against porcine reproductive and respiratory syndrome virus. Virol. Sin. 2011, 26, 267–272. [Google Scholar] [CrossRef]
- Plagemann, P.G. The primary GP5 neutralization epitope of North American isolates of porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2004, 102, 263–275. [Google Scholar] [CrossRef]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef]
- de Lima, M.; Pattnaik, A.K.; Flores, E.F.; Osorio, F.A. Serologic marker candidates identified among B-cell linear epitopes of Nsp2 and structural proteins of a North American strain of porcine reproductive and respiratory syndrome virus. Virology 2006, 353, 410–421. [Google Scholar] [CrossRef]
- Liu, D.; Chen, Y. Epitope screening and vaccine molecule design of PRRSV GP3 and GP5 protein based on immunoinformatics. J. Cell Mol. Med. 2024, 28, e18103. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Yu, H.; Tian, Z.J.; Liu, J.X.; An, T.Q.; Peng, J.M.; Li, G.X.; Jiang, Y.F.; Cai, X.H.; Xue, Q.; et al. Monoclonal antibodies and conserved antigenic epitopes in the C terminus of GP5 protein of the North American type porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2009, 138, 1–10. [Google Scholar] [CrossRef]
- Fan, B.; Liu, X.; Bai, J.; Zhang, T.; Zhang, Q.; Jiang, P. The amino acid residues at 102 and 104 in GP5 of porcine reproductive and respiratory syndrome virus regulate viral neutralization susceptibility to the porcine serum neutralizing antibody. Virus Res. 2015, 204, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Gu, J.M.; Lei, L.C.; Feng, X.; Sun, C.J.; Han, W.Y. Construction and induced neutralizing antibody level by recombinant phage expression carboxy terminal of GP5 protein from porcine reproductive and respiratory syndrome virus. Chin. J. Vet. Sci. 2019, 39, 830–834. [Google Scholar]
- Graham, B.S.; Gilman, M.S.A.; McLellan, J.S. Structure-Based Vaccine Antigen Design. Annu. Rev. Med. 2019, 70, 91–104. [Google Scholar] [CrossRef]
- Ren, D.; Zhang, X.; Zhang, W.; Lian, M.; Meng, X.; Li, T.; Xie, Q.; Shao, H.; Wan, Z.; Qin, A.; et al. A peptide-based ELISA for detection of antibodies against novel goose astrovirus type 1. J. Virol. Methods 2023, 312, 114646. [Google Scholar] [CrossRef]
- Shao, H.; Li, J.; Zhang, J.; Zhang, Q.; Ma, L.; Lu, J.; Li, T.; Xie, Q.; Wan, Z.; Qin, A.; et al. Research Note: A novel peptide-based ELISA for efficient detection of antibody against chicken infectious anemia virus. Poult. Sci. 2023, 102, 102284. [Google Scholar] [CrossRef]
- Xu, Y.; Ye, M.; Sun, S.; Cao, Q.; Luo, J.; Wang, Y.; Zheng, W.; Meurens, F.; Chen, N.; Zhu, J. CD163-Expressing Porcine Macrophages Support NADC30-like and NADC34-like PRRSV Infections. Viruses 2022, 14, 2056. [Google Scholar] [CrossRef]
- Chen, N.; Ye, M.; Li, S.; Huang, Y.; Zhou, R.; Yu, X.; Tian, K.; Zhu, J. Emergence of a novel highly pathogenic recombinant virus from three lineages of porcine reproductive and respiratory syndrome virus 2 in China 2017. Transbound Emerg. Dis. 2018, 65, 1775–1785. [Google Scholar] [CrossRef]
- Chen, N.; Ye, M.; Huang, Y.; Li, S.; Xiao, Y.; Li, X.; Li, S.; Li, X.; Yu, X.; Tian, K.; et al. Identification of Two Porcine Reproductive and Respiratory Syndrome Virus Variants Sharing High Genomic Homology but with Distinct Virulence. Viruses 2019, 11, 875. [Google Scholar] [CrossRef]
- Li, C.; Qiu, M.; Li, S.B.; Sun, Z.; Huang, Z.T.; Qi, W.H.; Qiu, Y.J.; Li, J.X.; Feng, B.H.; Zhao, D.S.; et al. Metagenomic and Pathogenic Assessments Identify a Pathogenic Porcine Reproductive and Respiratory Syndrome Virus 1 with New Deletions from Adult Slaughter Pig in 2022. Transbound. Emerg. Dis. 2023, 2023, 1–12. [Google Scholar] [CrossRef]
- Chen, N.; Liu, Q.; Qiao, M.; Deng, X.; Chen, X.; Sun, M. Whole genome characterization of a novel porcine reproductive and respiratory syndrome virus 1 isolate: Genetic evidence for recombination between Amervac vaccine and circulating strains in mainland China. Infect. Genet. Evol. 2017, 54, 308–313. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Peptide-Based ELISA | IDEXX | Coincidence Rate (%) | ||
|---|---|---|---|---|---|
| Positive | Negative | Positive | Negative | ||
| Sera (81) | 19 (1 *) | 62 (13 *) | 31 (13 #) | 50 (1 #) | 82.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Zhang, K.; Zhang, J.; Zhang, P.; He, P.; Deng, D.; Jiang, S.; Zheng, W.; Chen, N.; Bai, J.; et al. A Novel Peptide-Based Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Neutralizing Antibodies Against NADC30-like PRRSV GP5 Protein. Int. J. Mol. Sci. 2025, 26, 2619. https://doi.org/10.3390/ijms26062619
Sun S, Zhang K, Zhang J, Zhang P, He P, Deng D, Jiang S, Zheng W, Chen N, Bai J, et al. A Novel Peptide-Based Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Neutralizing Antibodies Against NADC30-like PRRSV GP5 Protein. International Journal of Molecular Sciences. 2025; 26(6):2619. https://doi.org/10.3390/ijms26062619
Chicago/Turabian StyleSun, Shaohua, Kaili Zhang, Jiajia Zhang, Pingping Zhang, Ping He, Dafu Deng, Sen Jiang, Wanglong Zheng, Nanhua Chen, Jianfa Bai, and et al. 2025. "A Novel Peptide-Based Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Neutralizing Antibodies Against NADC30-like PRRSV GP5 Protein" International Journal of Molecular Sciences 26, no. 6: 2619. https://doi.org/10.3390/ijms26062619
APA StyleSun, S., Zhang, K., Zhang, J., Zhang, P., He, P., Deng, D., Jiang, S., Zheng, W., Chen, N., Bai, J., & Zhu, J. (2025). A Novel Peptide-Based Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Neutralizing Antibodies Against NADC30-like PRRSV GP5 Protein. International Journal of Molecular Sciences, 26(6), 2619. https://doi.org/10.3390/ijms26062619