
KRAS–SOS-1 Inhibition as New Pharmacological Target to Counteract Anaplastic Thyroid Carcinoma (ATC)

 ,
, 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. In Vitro Results
2.1.1. The Effect of BAY-293 on Reducing Thyroid Cancer (TC) Cell Viability
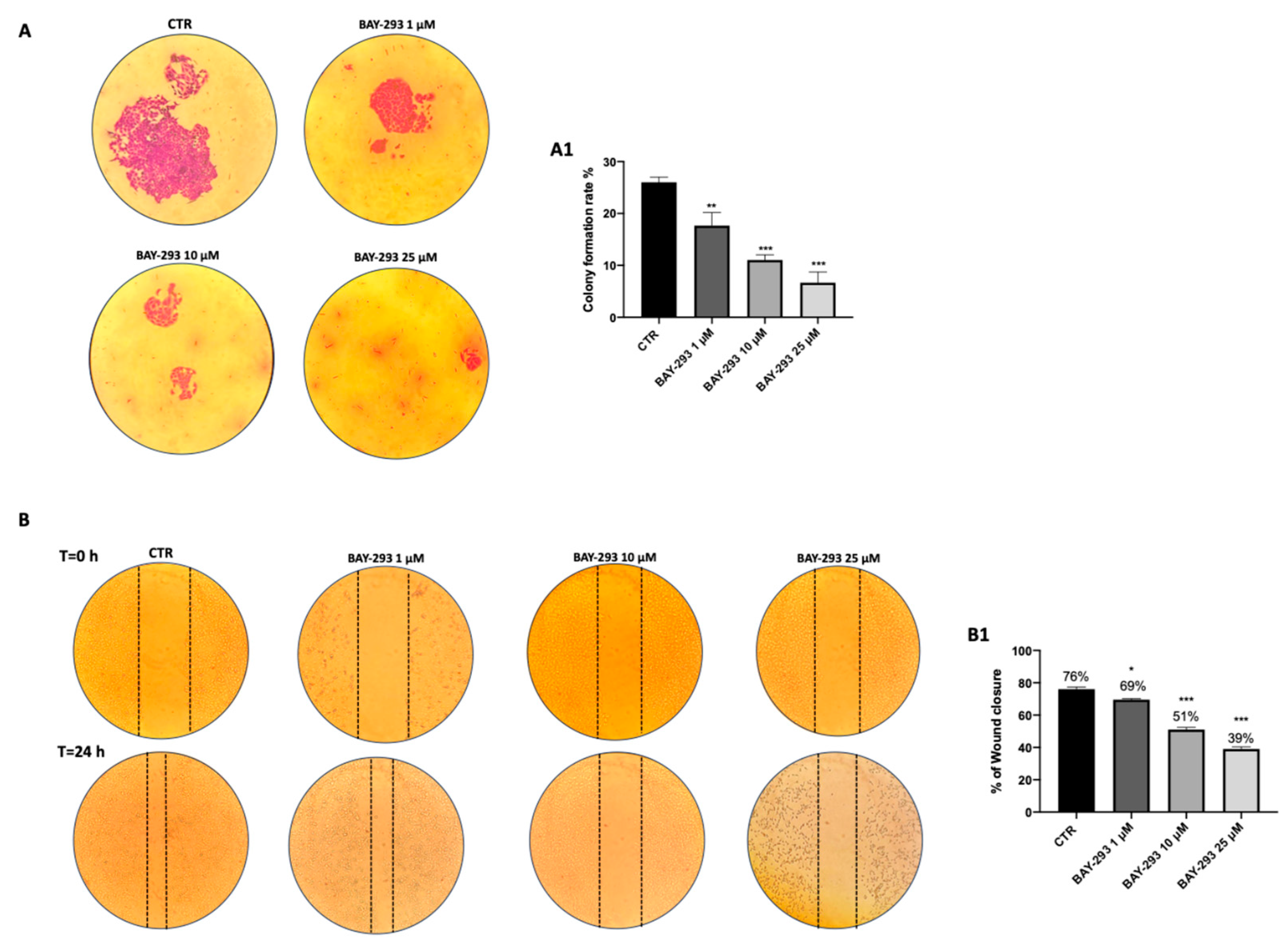
2.1.2. ATC Cell Migration and Proliferation Were Reduced by BAY-293 Treatment
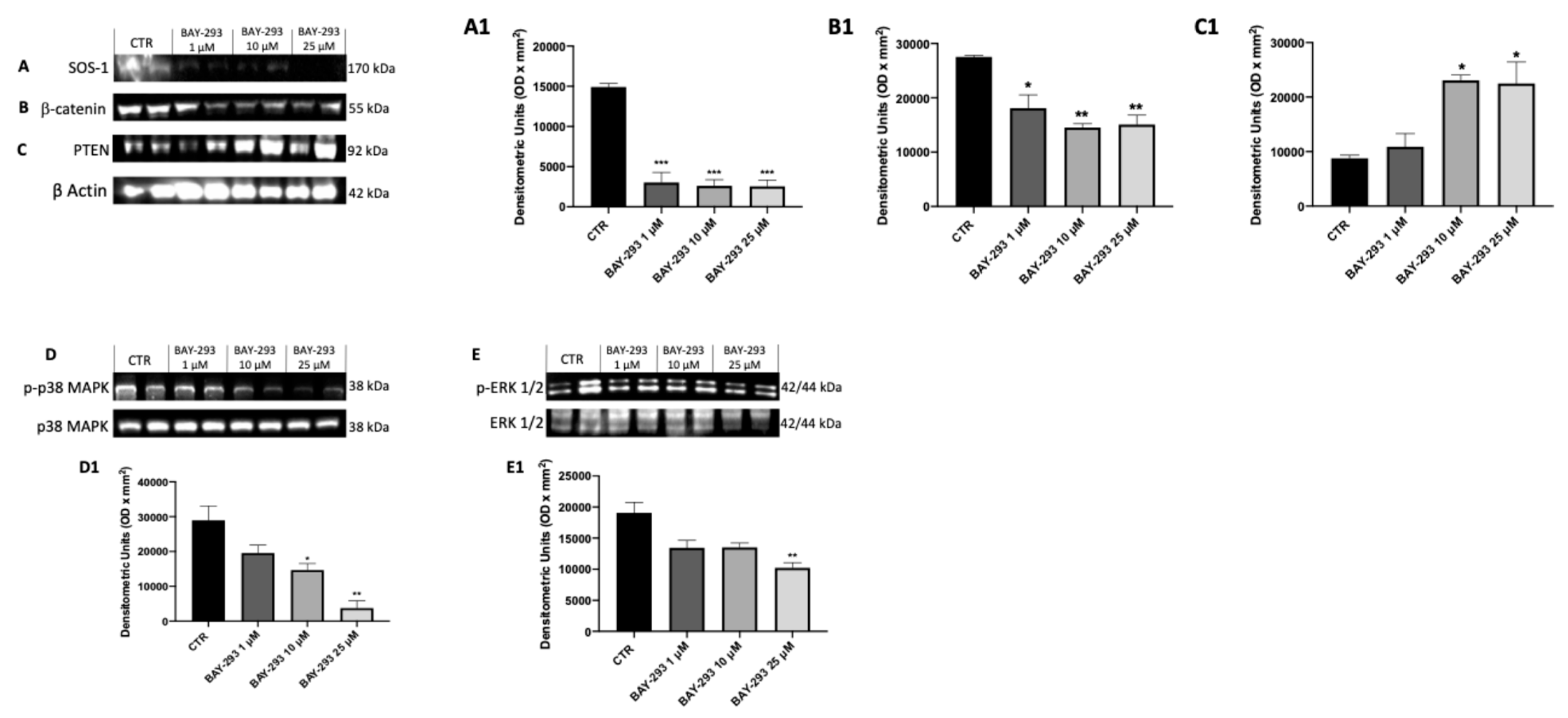
2.1.3. The Impact of BAY-293 on Modulation of the KRAS/SOS-1 Pathway
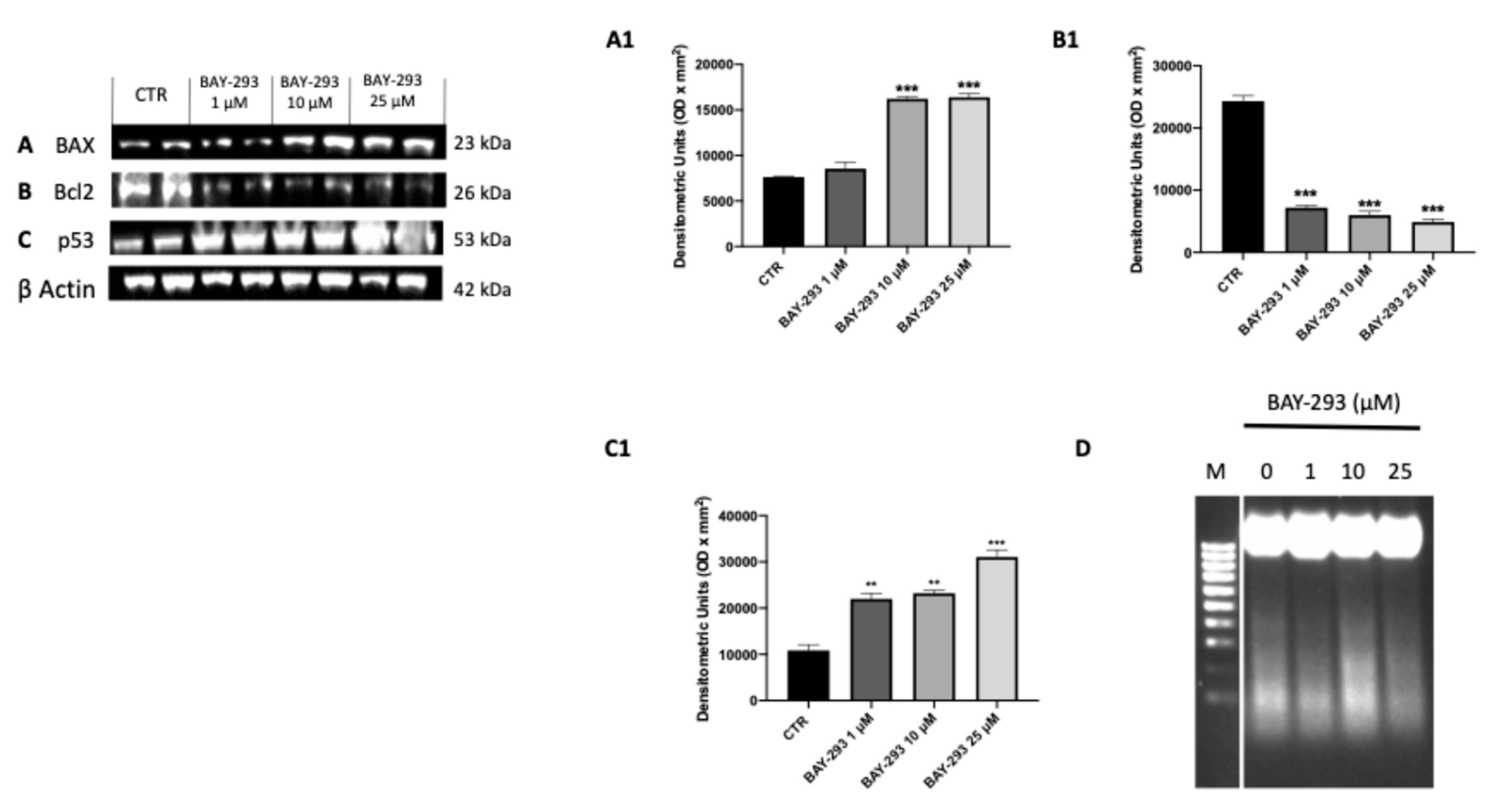
2.1.4. BAY-293 Activated ATC Cell Apoptosis
2.2. In Vivo Results
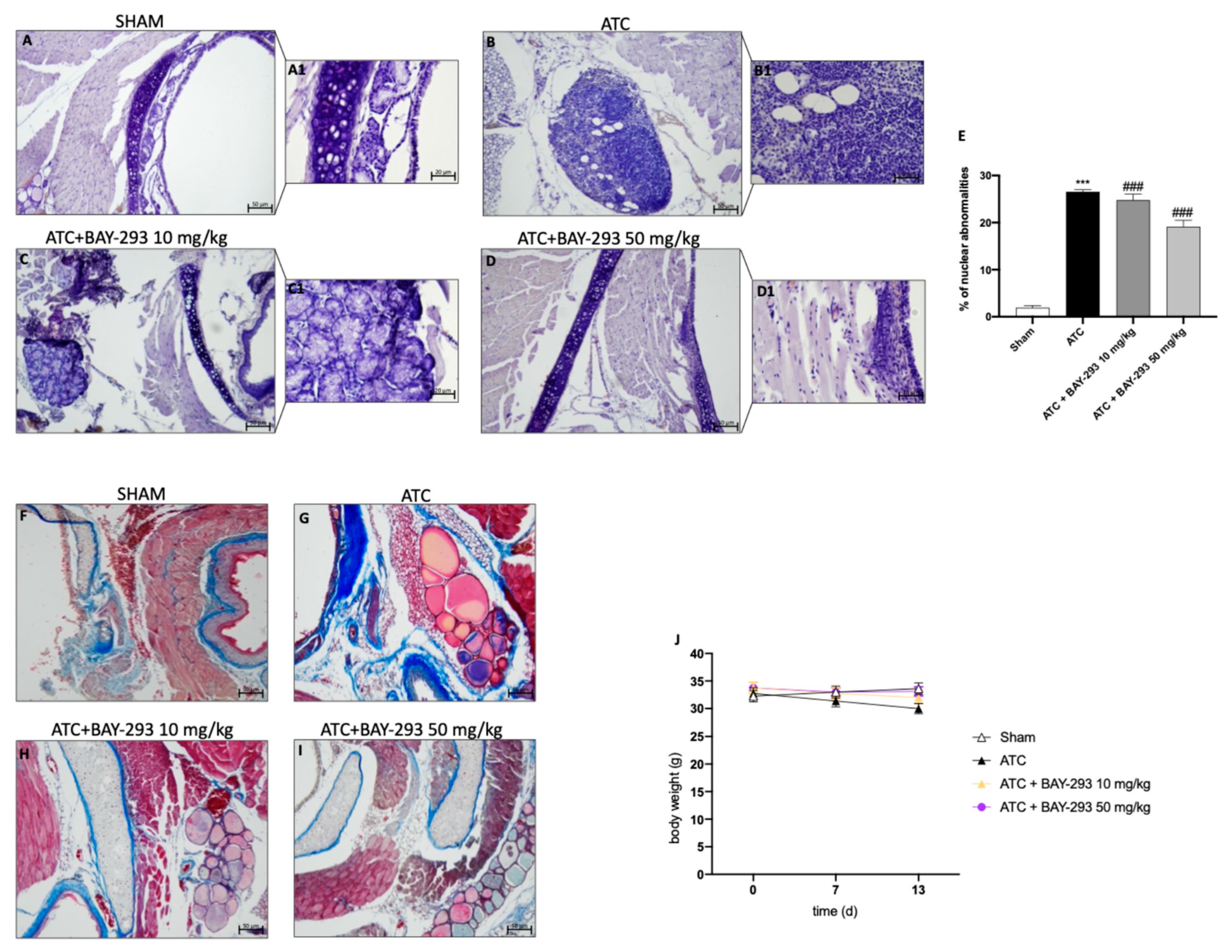
2.2.1. BAY-293 Improved Histopathological Features in an ATC Orthotopic Model
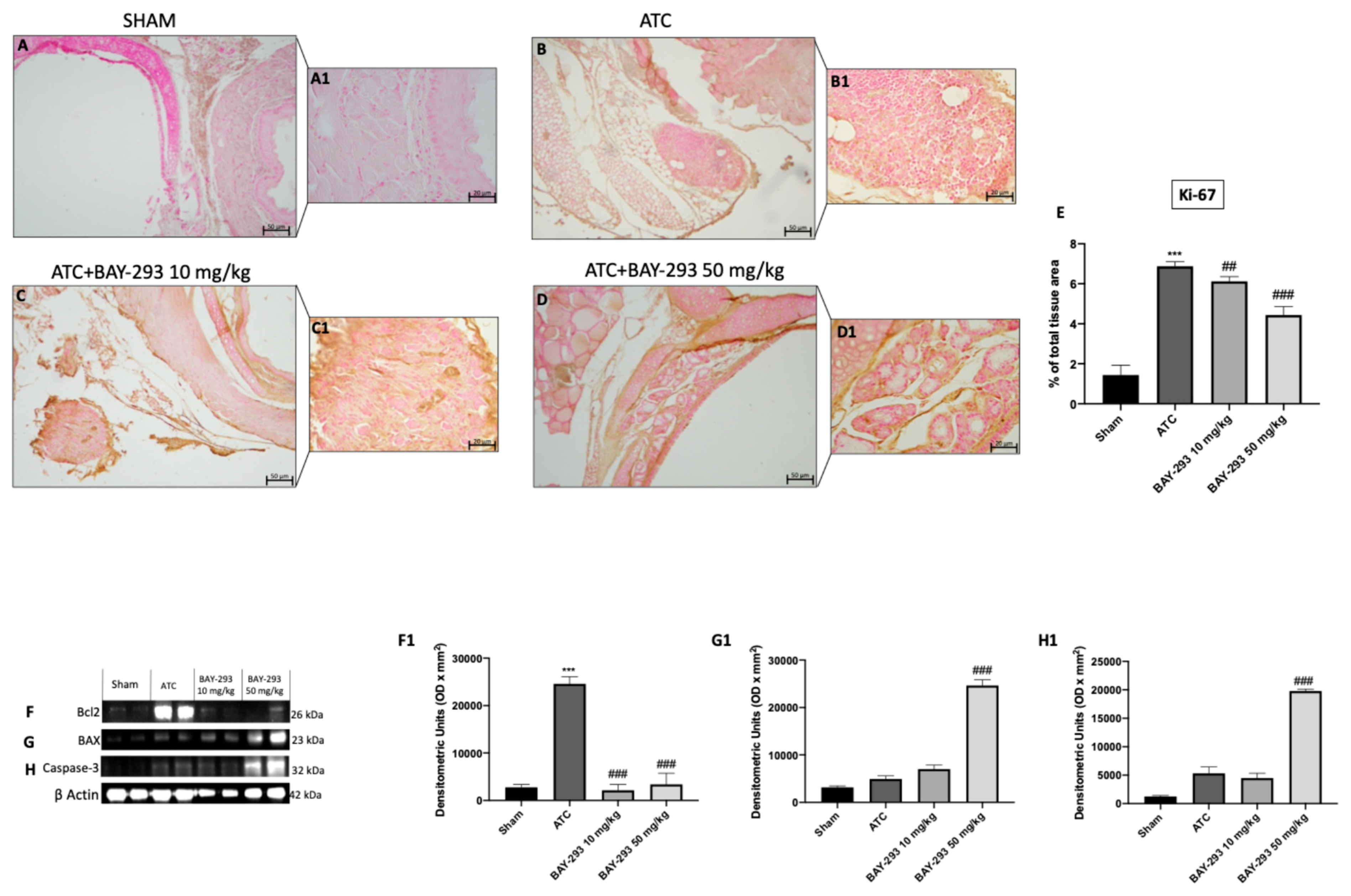
2.2.2. BAY-293 Reduced Ki-67 Expression and Increased Apoptosis in the Orthotopic Model
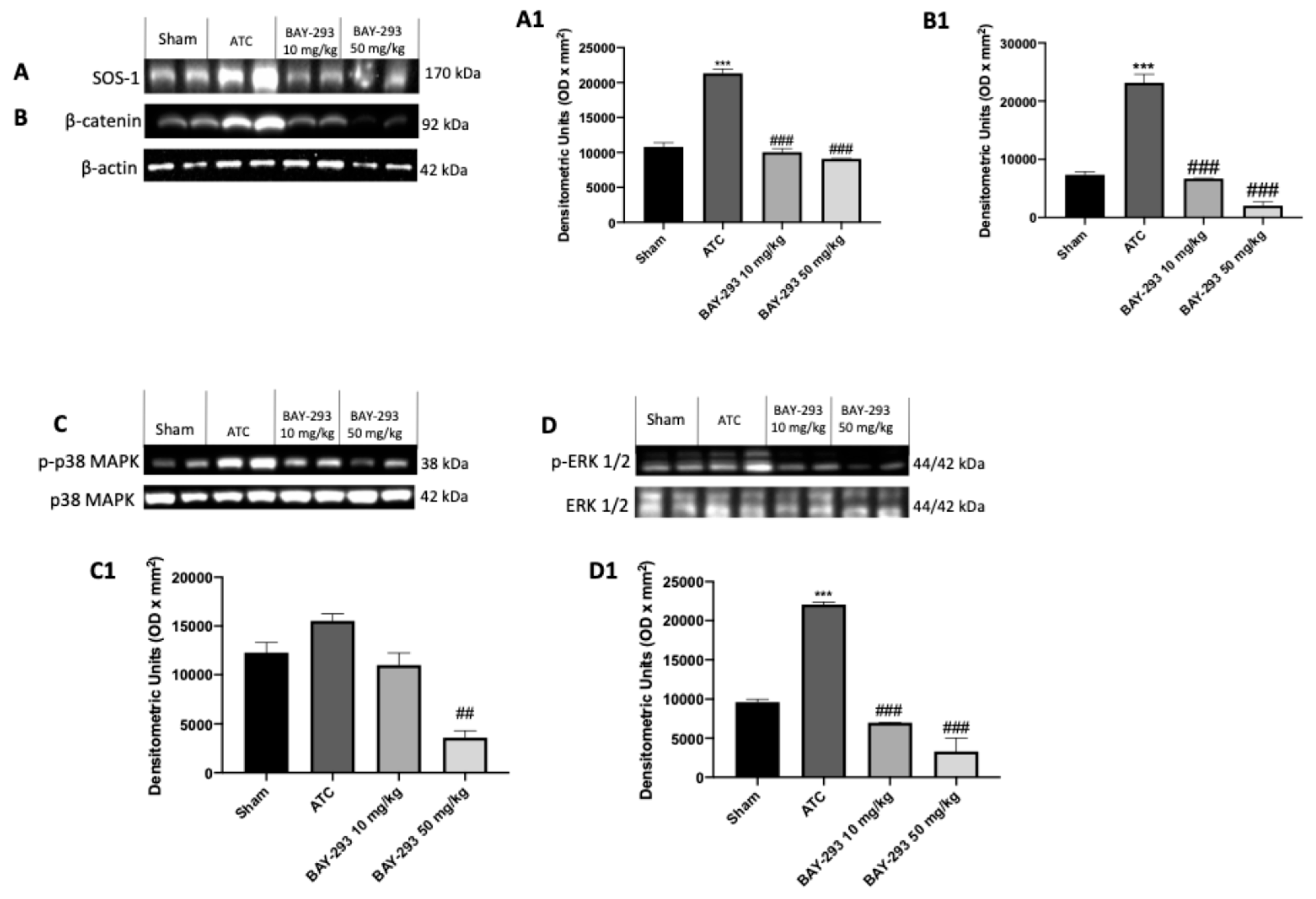
2.2.3. BAY-293 Inhibits the KRAS/SOS-1 Pathway in the Orthotopic Model
3. Discussion
4. Materials and Methods
4.1. In Vitro Studies
4.1.1. Materials
4.1.2. Cell Cultures
4.1.3. Cell Viability (MTT Assay)
4.1.4. Experimental Groups
4.1.5. Western Blot Analysis for Cell Lysates
4.1.6. Wound Healing Assay (Scratch Test)
4.1.7. Colony Formation Assay
4.1.8. DNA Fragmentation Assay
4.2. In Vivo Studies
4.2.1. Animals
4.2.2. Orthotopic Model of ATC
- SHAM group (8): intraperitoneal administration of saline;
- ATC group (8): mice that received tumor cell inoculation intraperitoneally administered with saline;
- ATC + BAY-293 10 mg/kg group (8): mice that received tumor cell inoculation intraperitoneally administered with BAY-293 at a dose of 10 mg/kg;
- ATC + BAY-293 50 mg/kg (8): mice that received tumor cell inoculation intraperitoneally administered with BAY-293 at a dose of 50 mg/kg.
4.2.3. Histological Evaluation
4.2.4. Masson’s Trichrome Staining
4.2.5. Immunohistochemistry Assay
4.2.6. Western Blot Analysis for Thyroid Samples
4.2.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATC | Anaplastic thyroid carcinoma |
| PTC | Papillary thyroid carcinoma |
| FTC | Follicular thyroid carcinoma |
| TC | Thyroid carcinoma |
| GDP | Guanosine diphosphate |
| GTP | Guanosine triphosphate |
| PI3K | Phosphoinositide 3-kinase |
| MAPK | Mitogen-activated protein kinase |
| SOS-1 | Son of sevenless 1 |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PTEN | Phosphatase and tensin homolog |
| p38 | Phosphorylated p38 mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| Bcl2 | B-cell leukemia/lymphoma 2 protein |
| BAX | Bcl2-associated X, apoptosis regulator |
| p53 | Tumor protein p53 |
References
- Deeken-Draisey, A.; Yang, G.-Y.; Gao, J.; Alexiev, B.A. Anaplastic thyroid carcinoma: An epidemiologic, histologic, immunohistochemical, and molecular single-institution study. Human. Pathol. 2018, 82, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Bychkov, A.; Agarwal, S.; Keelawat, S. Anaplastic Thyroid Carcinoma. In Thyroid FNA Cytology: Differential Diagnoses and Pitfalls; Springer: Singapore, 2024; pp. 407–425. [Google Scholar]
- Aslan, Z.A.; Granados-Garcia, M.; Luna-Ortiz, K.; Guerrero-Huerta, F.J.; Gomez-Pedraza, A.; Namendys-Silva, S.A.; Meneses-Garcia, A.; Ordonez-Mosquera, J.M. Anaplastic thyroid cancer: Multimodal treatment results. Ecancermedicalscience 2014, 8, 449. [Google Scholar] [CrossRef]
- Baloch, Z.W.; Asa, S.L.; Barletta, J.A.; Ghossein, R.A.; Juhlin, C.C.; Jung, C.K.; LiVolsi, V.A.; Papotti, M.G.; Sobrinho-Simoes, M.; Tallini, G.; et al. Overview of the 2022 WHO Classification of Thyroid Neoplasms. Endocr. Pathol. 2022, 33, 27–63. [Google Scholar] [CrossRef] [PubMed]
- Amaral, M.; Afonso, R.A.; Gaspar, M.M.; Reis, C.P. Anaplastic thyroid cancer: How far can we go? EXCLI J. 2020, 19, 800–812. [Google Scholar] [PubMed]
- Bible, K.C.; Kebebew, E.; Brierley, J.; Brito, J.P.; Cabanillas, M.E.; Clark, T.J., Jr.; Di Cristofano, A.; Foote, R.; Giordano, T.; Kasperbauer, J. 2021 American thyroid association guidelines for management of patients with anaplastic thyroid cancer: American thyroid association anaplastic thyroid cancer guidelines task force. Thyroid 2021, 31, 337–386. [Google Scholar] [CrossRef]
- Haddad, R.I.; Bischoff, L.; Ball, D.; Bernet, V.; Blomain, E.; Busaidy, N.L.; Campbell, M.; Dickson, P.; Duh, Q.Y.; Ehya, H.; et al. Thyroid Carcinoma, Version 2.2022, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2022, 20, 925–951. [Google Scholar] [CrossRef]
- Wright, P.A.; Williams, E.D.; Lemoine, N.R.; Wynford-Thomas, D. Radiation-associated and ‘spontaneous’ human thyroid carcinomas show a different pattern of ras oncogene mutation. Oncogene 1991, 6, 471–473. [Google Scholar]
- Zhu, Z.; Gandhi, M.; Nikiforova, M.N.; Fischer, A.H.; Nikiforov, Y.E. Molecular profile and clinical-pathologic features of the follicular variant of papillary thyroid carcinoma: An unusually high prevalence of ras mutations. Am. J. Clin. Pathol. 2003, 120, 71–77. [Google Scholar] [CrossRef]
- Nikiforov, Y.E. Genetic alterations involved in the transition from well-differentiated to poorly differentiated and anaplastic thyroid carcinomas. Endocr. Pathol. 2004, 15, 319–327. [Google Scholar] [CrossRef]
- Prete, A.; Borges de Souza, P.; Censi, S.; Muzza, M.; Nucci, N.; Sponziello, M. Update on Fundamental Mechanisms of Thyroid Cancer. Front. Endocrinol. 2020, 11, 102. [Google Scholar] [CrossRef]
- Jeng, H.-H. Characterization of the Physiological Roles of Sos-WT Ras Signaling Cascade in Oncogenic Ras-Driven Tumorigenesis. Ph.D. Thesis, New York University, New York, NY, USA, 2011. [Google Scholar]
- Menyhárd, D.K.; Pálfy, G.; Orgován, Z.; Vida, I.; Keserű, G.M.; Perczel, A. Structural impact of GTP binding on downstream KRAS signaling. Chem. Sci. 2020, 11, 9272–9289. [Google Scholar] [CrossRef] [PubMed]
- Jančík, S.; Drábek, J.; Radzioch, D.; Hajdúch, M. Clinical relevance of KRAS in human cancers. BioMed Res. Int. 2010, 2010, 150960. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.R.; Rosenberg, S.C.; McCormick, F.; Malek, S. RAS-targeted therapies: Is the undruggable drugged? Nat. Rev. Drug Discov. 2020, 19, 533–552. [Google Scholar] [CrossRef]
- Kessler, D.; Gerlach, D.; Kraut, N.; McConnell, D.B. Targeting son of sevenless 1: The pacemaker of KRAS. Curr. Opin. Chem. Biol. 2021, 62, 109–118. [Google Scholar] [CrossRef]
- Fedele, C.; Li, S.; Teng, K.W.; Foster, C.J.R.; Peng, D.; Ran, H.; Mita, P.; Geer, M.J.; Hattori, T.; Koide, A.; et al. SHP2 inhibition diminishes KRASG12C cycling and promotes tumor microenvironment remodeling. J. Exp. Med. 2021, 218, e20201414. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Fan, Y.; Wang, X.; Wang, J.; Yang, H.; Fan, W.; Tang, C. Design, synthesis and biological evaluation of quinazoline SOS1 inhibitors. Bioorg. Med. Chem. Lett. 2023, 88, 129265. [Google Scholar] [CrossRef]
- Plangger, A.; Rath, B.; Stickler, S.; Hochmair, M.; Lang, C.; Weigl, L.; Funovics, M.; Hamilton, G. Cytotoxicity of combinations of the pan-KRAS SOS1 inhibitor BAY-293 against pancreatic cancer cell lines. Discov. Oncol. 2022, 13, 84. [Google Scholar] [CrossRef]
- Wang, C.X.; Wang, T.T.; Zhang, K.D.; Li, M.Y.; Shen, Q.C.; Lu, S.Y.; Zhang, J. Pan-KRAS inhibitors suppress proliferation through feedback regulation in pancreatic ductal adenocarcinoma. Acta Pharmacol. Sin. 2022, 43, 2696–2708. [Google Scholar] [CrossRef]
- Sewell, W.; Reeb, A.; Lin, R.Y. An orthotopic mouse model of anaplastic thyroid carcinoma. J. Vis. Exp. 2013, e50097. [Google Scholar] [CrossRef]
- Nucera, C.; Nehs, M.A.; Mekel, M.; Zhang, X.; Hodin, R.; Lawler, J.; Nose, V.; Parangi, S. A novel orthotopic mouse model of human anaplastic thyroid carcinoma. Thyroid 2009, 19, 1077–1084. [Google Scholar] [CrossRef]
- Plangger, A.; Rath, B.; Hochmair, M.; Funovics, M.; Hamilton, G. Cytotoxicity of combinations of the pan-KRAS inhibitor BAY-293 against primary non-small lung cancer cells. Transl. Oncol. 2021, 14, 101230. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, H.N.; Jeong, M.S.; Jang, S.B. Oncogenic KRAS: Signaling and drug resistance. Cancers 2021, 13, 5599. [Google Scholar] [CrossRef] [PubMed]
- Schubert, L.; Mariko, M.L.; Clerc, J.; Huillard, O.; Groussin, L. MAPK Pathway Inhibitors in Thyroid Cancer: Preclinical and Clinical Data. Cancers 2023, 15, 710. [Google Scholar] [CrossRef]
- Jeong, W.J.; Ro, E.J.; Choi, K.Y. Interaction between Wnt/beta-catenin and RAS-ERK pathways and an anti-cancer strategy via degradations of beta-catenin and RAS by targeting the Wnt/beta-catenin pathway. NPJ Precis. Oncol. 2018, 2, 5. [Google Scholar] [CrossRef]
- Xu, L.L.; Li, C.C.; An, L.Y.; Dai, Z.; Chen, X.Y.; You, Q.D.; Hu, C.; Di, B. Selective apoptosis-inducing activity of synthetic hydrocarbon-stapled SOS1 helix with d-amino acids in H358 cancer cells expressing KRAS(G12C). Eur. J. Med. Chem. 2020, 185, 111844. [Google Scholar] [CrossRef]
- Piersma, B.; Hayward, M.-K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef]
- Silver Karcioglu, A.; Iwata, A.J.; Pusztaszeri, M.; Abdelhamid Ahmed, A.H.; Randolph, G.W. The American Thyroid Association (ATA) integrates molecular testing into its framework for managing patients with anaplastic thyroid carcinoma (ATC): Update on the 2021 ATA ATC guidelines. Cancer Cytopathol. 2022, 130, 174–180. [Google Scholar] [CrossRef]
- Tallini, G.; Garcia-Rostan, G.; Herrero, A.; Zelterman, D.; Viale, G.; Bosari, S.; Carcangiu, M.L. Downregulation of p27KIP1 and Ki67/Mib1 labeling index support the classification of thyroid carcinoma into prognostically relevant categories. Am. J. Surg. Pathol. 1999, 23, 678–685. [Google Scholar] [CrossRef]
- Are, C.; Shaha, A.R. Anaplastic thyroid carcinoma: Biology, pathogenesis, prognostic factors, and treatment approaches. Ann. Surg. Oncol. 2006, 13, 453–464. [Google Scholar] [CrossRef]
- Ito, K.i.; Hanamura, T.; Murayama, K.; Okada, T.; Watanabe, T.; Harada, M.; Ito, T.; Koyama, H.; Kanai, T.; Maeno, K. Multimodality therapeutic outcomes in anaplastic thyroid carcinoma: Improved survival in subgroups of patients with localized primary tumors. Head Neck 2012, 34, 230–237. [Google Scholar] [CrossRef]
- Ljubas, J.; Ovesen, T.; Rusan, M. A systematic review of phase II targeted therapy clinical trials in anaplastic thyroid cancer. Cancers 2019, 11, 943. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Ghossein, R. Genomic landscape of poorly differentiated and anaplastic thyroid carcinoma. Endocr. Pathol. 2016, 27, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, N.R.; Mayall, E.S.; Wyllie, F.S.; Williams, E.D.; Goyns, M.; Stringer, B.; Wynford-Thomas, D. High frequency of ras oncogene activation in all stages of human thyroid tumorigenesis. Oncogene 1989, 4, 159–164. [Google Scholar] [PubMed]
- Suarez, H.G. Genetic alterations in human epithelial thyroid tumours. Clin. Endocrinol. 1998, 48, 531–546. [Google Scholar] [CrossRef]
- Stringer, B.M.; Rowson, J.M.; Parkar, M.H.; Seid, J.M.; Hearn, P.R.; Wynford-Thomas, D.; Ingemansson, S.; Woodhouse, N.; Goyns, M.H. Detection of the H-RAS oncogene in human thyroid anaplastic carcinomas. Experientia 1989, 45, 372–376. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Tiacci, E.; Trifonov, V.; Schiavoni, G.; Holmes, A.; Kern, W.; Martelli, M.P.; Pucciarini, A.; Bigerna, B.; Pacini, R.; Wells, V.A.; et al. BRAF mutations in hairy-cell leukemia. N. Engl. J. Med. 2011, 364, 2305–2315. [Google Scholar] [CrossRef]
- Ullah, R.; Yin, Q.; Snell, A.H.; Wan, L. RAF-MEK-ERK pathway in cancer evolution and treatment. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 123–154. [Google Scholar]
- De Luca, A.; Maiello, M.R.; D’Alessio, A.; Pergameno, M.; Normanno, N. The RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: Role in cancer pathogenesis and implications for therapeutic approaches. Expert Opin. Ther. Targets 2012, 16, S17–S27. [Google Scholar] [CrossRef]
- Davies, E.J.; Marsh Durban, V.; Meniel, V.; Williams, G.T.; Clarke, A.R. PTEN loss and KRAS activation leads to the formation of serrated adenomas and metastatic carcinoma in the mouse intestine. J. Pathol. 2014, 233, 27–38. [Google Scholar] [CrossRef]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef]
- Ju, C.; Zhou, M.; Du, D.; Wang, C.; Yao, J.; Li, H.; Luo, Y.; He, F.; He, J. EIF4A3-mediated circ_0042881 activates the RAS pathway via miR-217/SOS1 axis to facilitate breast cancer progression. Cell Death Dis. 2023, 14, 559. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tang, X.; Wang, Z.; Feng, F.; Xu, C.; Zhao, Q.; Wu, Y.; Sun, H.; Chen, Y. Inhibition of Son of Sevenless Homologue 1 (SOS1): Promising therapeutic treatment for KRAS-mutant cancers. Eur. J. Med. Chem. 2023, 261, 115828. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, L.; Mannino, D.; Casili, G.; Repici, A.; Crupi, L.; Paterniti, I.; Esposito, E.; Campolo, M. Prolyl oligopeptidase inhibition ameliorates experimental pulmonary fibrosis both in vivo and in vitro. Respir. Res. 2023, 24, 211. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Diao, H.; Lin, Z.; Gao, J.; Zhao, Y.; Zhang, W.; Wang, Q.; Lin, J.; Zhang, D.; Jin, Y.; et al. Benzyl Isothiocyanate Induces Apoptosis and Inhibits Tumor Growth in Canine Mammary Carcinoma via Downregulation of the Cyclin B1/Cdk1 Pathway. Front. Vet. Sci. 2020, 7, 580530. [Google Scholar] [CrossRef]
- Ohnishi, K.; Ota, I.; Yane, K.; Takahashi, A.; Yuki, K.; Emoto, M.; Hosoi, H.; Ohnishi, T. Glycerol as a chemical chaperone enhances radiation-induced apoptosis in anaplastic thyroid carcinoma cells. Mol. Cancer 2002, 1, 4. [Google Scholar] [CrossRef]
- Nucera, C.; Nehs, M.A.; Nagarkatti, S.S.; Sadow, P.M.; Mekel, M.; Fischer, A.H.; Lin, P.S.; Bollag, G.E.; Lawler, J.; Hodin, R.A.; et al. Targeting BRAFV600E with PLX4720 displays potent antimigratory and anti-invasive activity in preclinical models of human thyroid cancer. Oncologist 2011, 16, 296–309. [Google Scholar] [CrossRef]
- Fanfone, D.; Stanicki, D.; Nonclercq, D.; Port, M.; Vander Elst, L.; Laurent, S.; Muller, R.N.; Saussez, S.; Burtea, C. Molecular Imaging of Galectin-1 Expression as a Biomarker of Papillary Thyroid Cancer by Using Peptide-Functionalized Imaging Probes. Biology 2020, 9, 53. [Google Scholar] [CrossRef]
- Scuderi, S.A.; Filippone, A.; Basilotta, R.; Mannino, D.; Casili, G.; Capra, A.P.; Chisari, G.; Colarossi, L.; Sava, S.; Campolo, M. GSK343, an inhibitor of enhancer of zeste homolog 2, reduces glioblastoma progression through inflammatory process modulation: Focus on canonical and non-canonical NF-ΚB/IκBα Pathways. Int. J. Mol. Sci. 2022, 23, 13915. [Google Scholar] [CrossRef]
- Fallahi, P.; Ferrari, S.M.; Elia, G.; Ragusa, F.; Patrizio, A.; Paparo, S.R.; Marone, G.; Galdiero, M.R.; Guglielmi, G.; Foddis, R.; et al. Primary cell cultures for the personalized therapy in aggressive thyroid cancer of follicular origin. Semin. Cancer Biol. 2022, 79, 203–216. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, D.; Basilotta, R.; De Luca, F.; Casili, G.; Esposito, E.; Paterniti, I. KRAS–SOS-1 Inhibition as New Pharmacological Target to Counteract Anaplastic Thyroid Carcinoma (ATC). Int. J. Mol. Sci. 2025, 26, 2579. https://doi.org/10.3390/ijms26062579
Mannino D, Basilotta R, De Luca F, Casili G, Esposito E, Paterniti I. KRAS–SOS-1 Inhibition as New Pharmacological Target to Counteract Anaplastic Thyroid Carcinoma (ATC). International Journal of Molecular Sciences. 2025; 26(6):2579. https://doi.org/10.3390/ijms26062579
Chicago/Turabian StyleMannino, Deborah, Rossella Basilotta, Fabiola De Luca, Giovanna Casili, Emanuela Esposito, and Irene Paterniti. 2025. "KRAS–SOS-1 Inhibition as New Pharmacological Target to Counteract Anaplastic Thyroid Carcinoma (ATC)" International Journal of Molecular Sciences 26, no. 6: 2579. https://doi.org/10.3390/ijms26062579
APA StyleMannino, D., Basilotta, R., De Luca, F., Casili, G., Esposito, E., & Paterniti, I. (2025). KRAS–SOS-1 Inhibition as New Pharmacological Target to Counteract Anaplastic Thyroid Carcinoma (ATC). International Journal of Molecular Sciences, 26(6), 2579. https://doi.org/10.3390/ijms26062579