

Hepatoprotective Activity of Nature-Derived Polyphenols Studied by Mass Spectrometry Based Multi-OMICS Approach
 , , ,
, , ,  ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
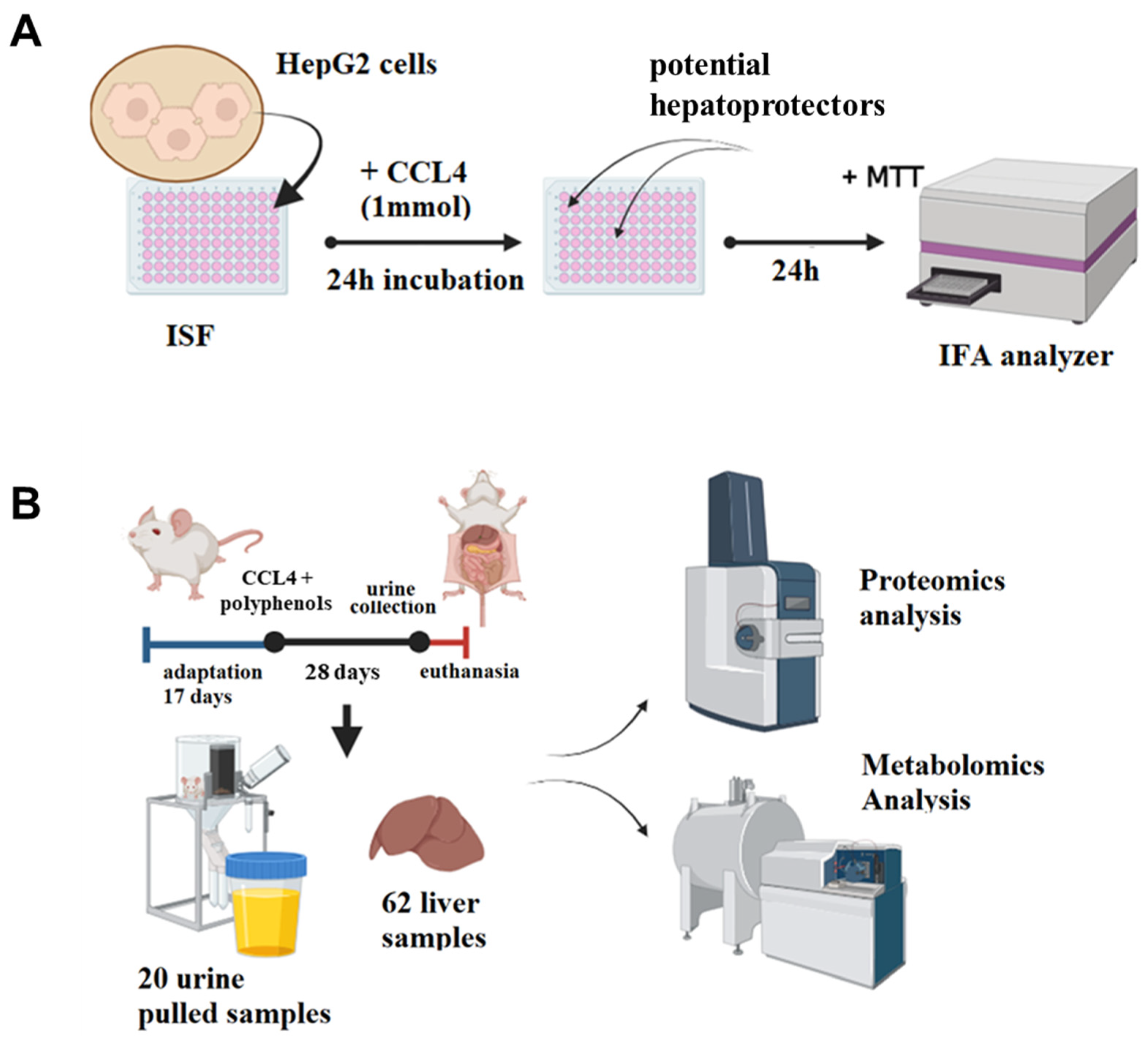
2.1. Overview of the Study Pipeline
2.2. Hepatoprotective Effect In Vitro
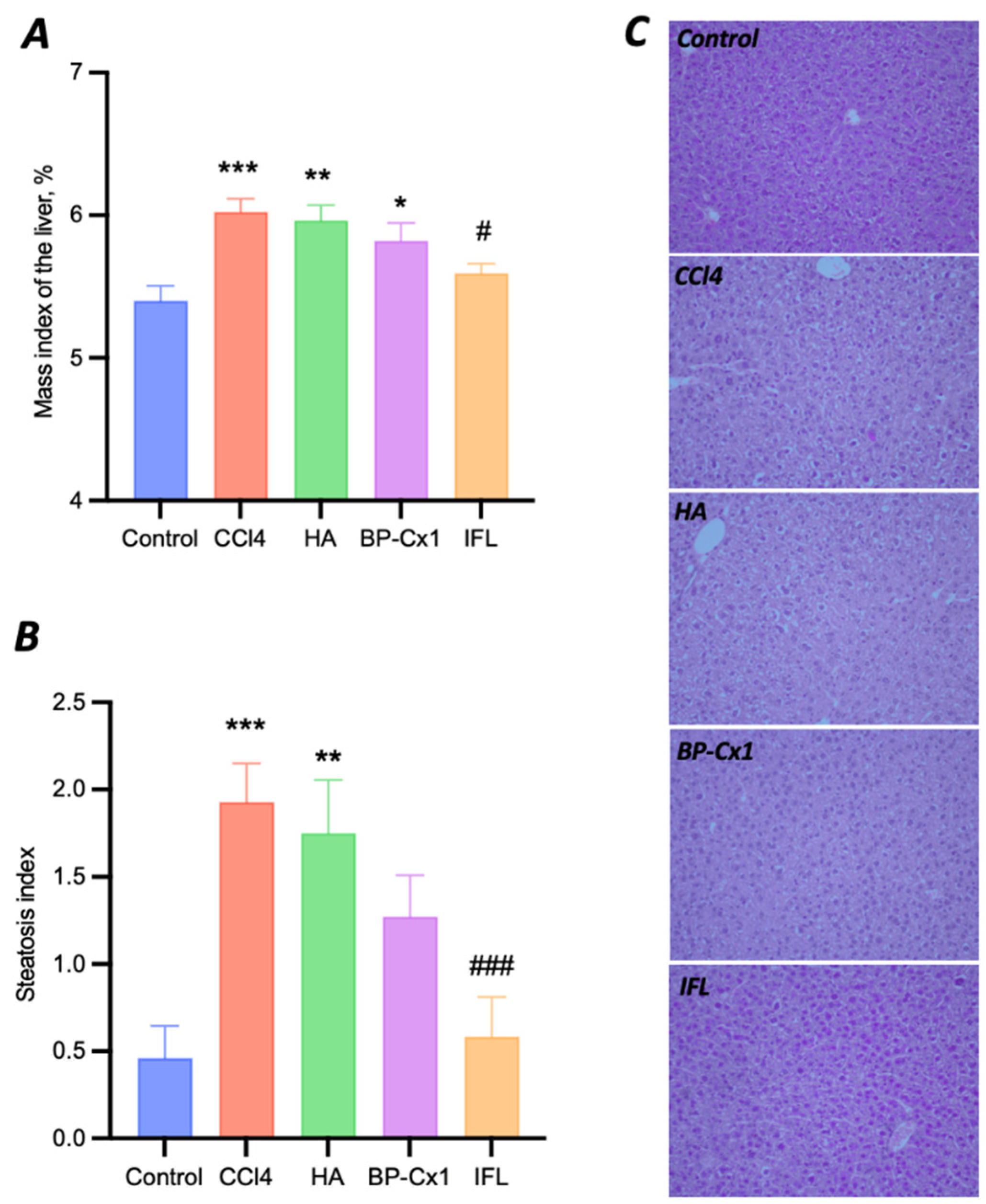
2.3. Hepatoprotective Effects In Vivo
2.4. Results of Proteomic Analysis
2.4.1. Urine Proteome Fraction Analysis
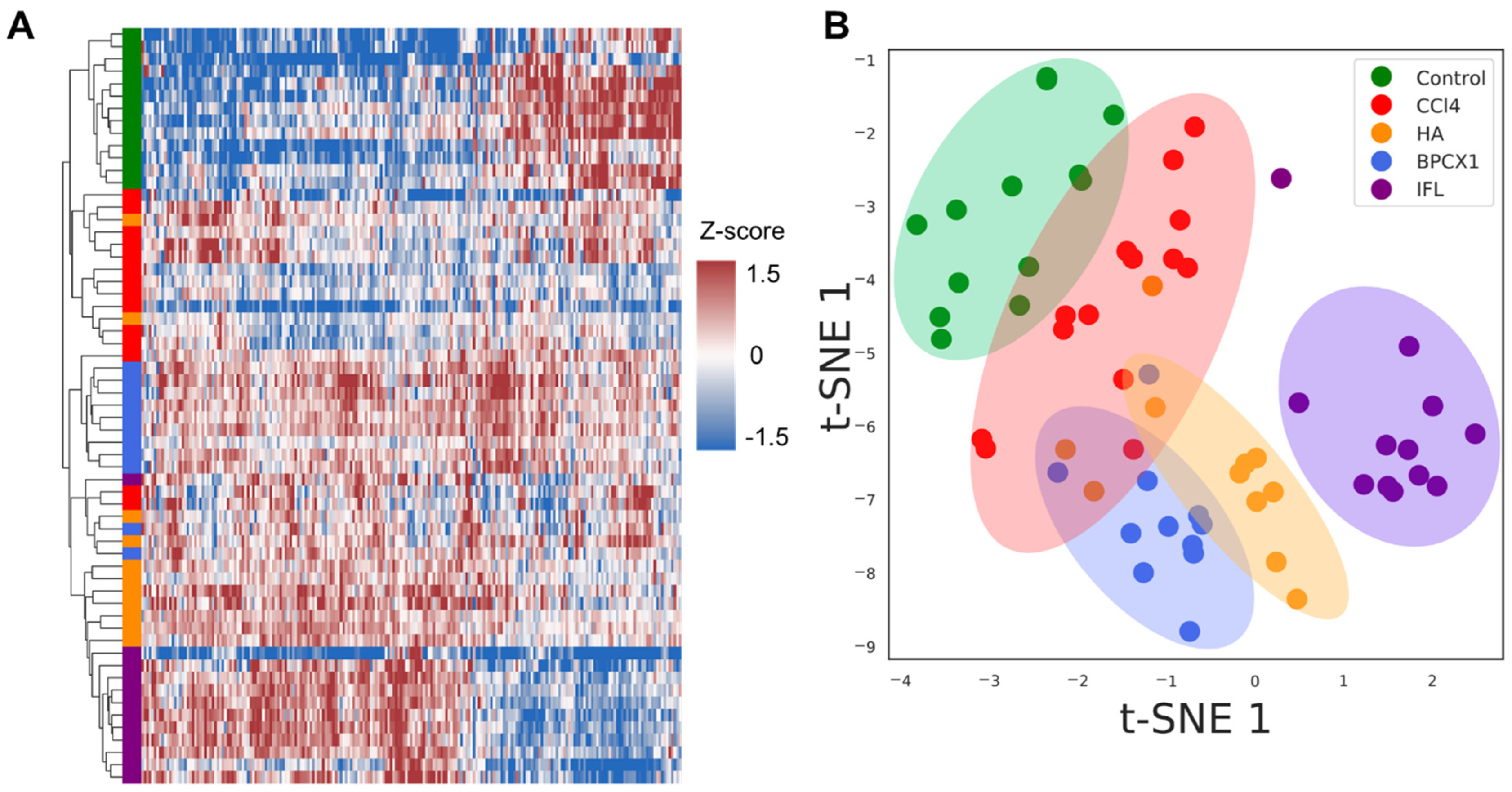
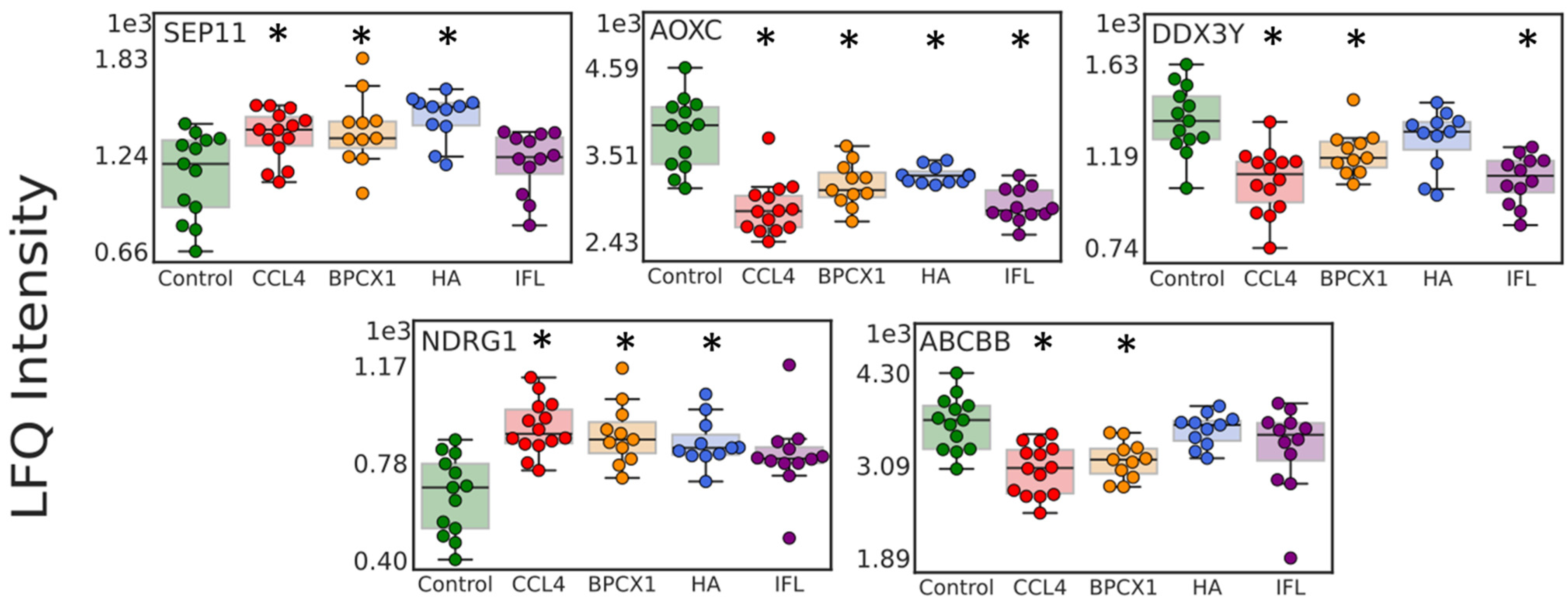
2.4.2. Liver Proteome Fraction Analysis
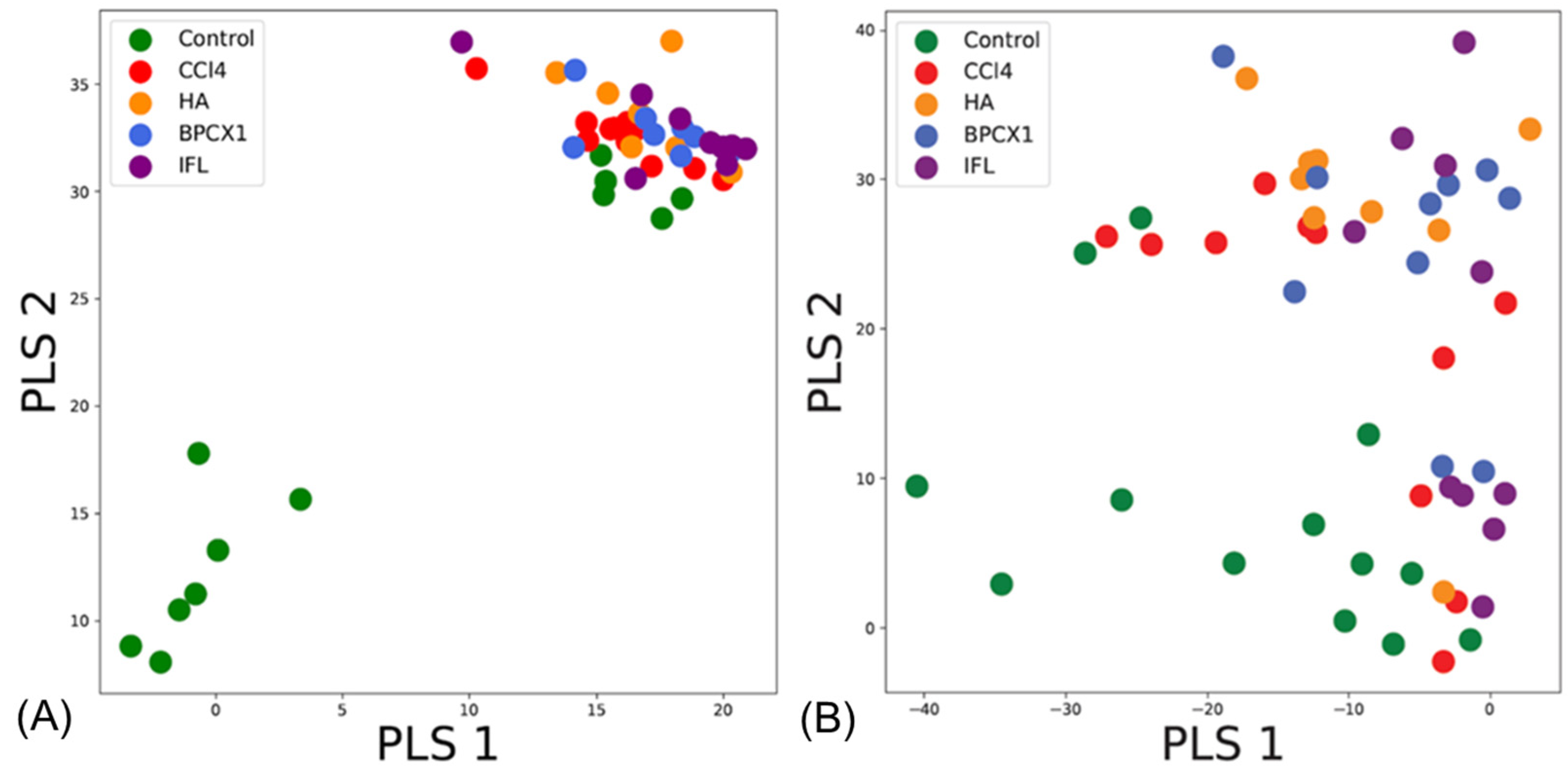
2.5. Results of Molecular Fingerprinting by FT-ICR MS
3. Discussion
4. Materials and Methods
4.1. Chemicals
- Fraction of humic acids (HA) from low-mineralized silt sulfide mud (peloids) of lake Molochka, Sergievsky district, Samara region, Russia (MRC “Sergievskie Mineral Waters” FMBA of Russia).
- Fulvic acid (FA) of low-mineralized silt sulfide mud (peloids) of lake Molochka, Sergievsky district, Samara region, Russia (MRC “Sergievskie Mineral Waters” FMBA of Russia).
- Castoreum extract (CE). Light orange powder [41]; 100 g of raw material contains 46.6 g of beaver musk extract (FSUE NPC “Pharmzashchita” FMBA of Russia). For in vitro study, powder of beaver musk substance was diluted in distilled water until a homogeneous suspension was formed. Sample weight 0.499 g. An alcohol extract was prepared by adding ethyl alcohol in a weight ratio of 1:2; the solution was treated with ultrasound and centrifuged at 13,000 rpm to remove the sediment. The solution was evaporated and re-dissolved in a nutrient medium.
- Isoflavones from kudzu Pueraria lobata roots (IFL)—kudzu root extract (Shaanxi Sheng, Xi’an, China) [42]. Sample weight 0.82 g. An alcohol extract was prepared by adding ethyl alcohol in a weight ratio of 1:2; the solution was treated with ultrasound and centrifuged at 13,000 rpm to remove the sediment. The solution was evaporated and redissolved.
- Water-soluble lignin derivative BP-Cx-1 [3] (Nobel Ltd., Saint Petersburg, Russia)—a sterile 0.42% ammonia solution (batch X0621D33). In the in vitro studies, BP-Cx-1 was tested at a concentration of 0.0042% (V/V), and for the in vivo studies, at a concentration of 0.42% (V/V).
4.2. Animal Studies
4.3. Experimental Design
- Control—negative control—intraperitoneal injection of sunflower oil (0.2 mL/mouse) (n = 13).
- Control CCl4—positive control—intraperitoneal injection of CCl4 (1 mL/kg) diluted ten times in sunflower oil 2 times per week (6 injections in total) (n = 14).
- CCl4 + Fraction of humic acids (HA)—intraperitoneal injection of CCl4 (1 mL/kg) diluted ten times in sunflower oil 2 times per week (6 injections in total) + intragastric administration of HA (60 mg/kg) 3 times a week, for 4 weeks (n = 12).
- CCl4 + BP-Cx-1—intraperitoneal injection of CCl4 (1 mL/kg) diluted ten times in sunflower oil 2 times per week (6 injections in total) + intragastric administration of BP-Cx-1 (60 mg/kg) 3 times a week, for 4 weeks (n = 11).
- CCl4 + isoflavones isolated from the root of Pueralia lobata (IFL)—intraperitoneal injection of CCl4 (1 mL/kg) diluted ten times in sunflower oil 2 times per week (6 injections in total) + intragastric administration of IFL (60 mg/kg) 3 times a week, for 4 weeks (n = 12).
4.4. MTT Colorimetric Assay
4.5. Proteomic Analysis
4.5.1. Urine Sample Preparation
4.5.2. Liver Sample Preparation
4.5.3. LC–MS/MS Analysis
4.5.4. Data Analysis
4.6. Molecular Fingerprinting by FT-ICR MS
4.6.1. Extraction of Metabolites
4.6.2. FT-ICR MS Analysis
4.6.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Slamenová, D.; Kosíková, B.; Lábaj, J.; Ruzeková, L. Oxidative/Antioxidative Effects of Different Lignin Preparations on DNA in Hamster V79 Cells. Neoplasma 2000, 47, 349–353. [Google Scholar]
- Yoshikawa, T.; Yagi, T.; Shinohara, S.; Fukunaga, T.; Nakasaka, Y.; Tago, T.; Masuda, T. Production of Phenols from Lignin via Depolymerization and Catalytic Cracking. Fuel Process. Technol. 2013, 108, 69–75. [Google Scholar] [CrossRef]
- Fedoros, E.I.; Orlov, A.A.; Zherebker, A.; Gubareva, E.A.; Maydin, M.A.; Konstantinov, A.I.; Krasnov, K.A.; Karapetian, R.N.; Izotova, E.I.; Pigarev, S.E.; et al. Novel Water-Soluble Lignin Derivative BP-Cx-1: Identification of Components and Screening of Potential Targets in Silico and in Vitro. Oncotarget 2018, 9, 18578–18593. [Google Scholar] [CrossRef]
- Coccia, A.; Mosca, L.; Puca, R.; Mangino, G.; Rossi, A.; Lendaro, E. Extra-Virgin Olive Oil Phenols Block Cell Cycle Progression and Modulate Chemotherapeutic Toxicity in Bladder Cancer Cells. Oncol. Rep. 2016, 36, 3095–3104. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, J.; Lin, J. A Simple High-Performance Liquid Chromatography Method for the Determination of Throat-Burning Oleocanthal with Probated Antiinflammatory Activity in Extra Virgin Olive Oils. J. Agric. Food Chem. 2006, 54, 3204–3208. [Google Scholar] [CrossRef]
- Espinoza-Acosta, J.L.; Torres-Chávez, P.I.; Ramírez-Wong, B.; López-Saiz, C.M.; Montaño-Leyva, B. Antioxidant, Antimicrobial, and Antimutagenic Properties of Technical Lignins and Their Applications. BioResources 2016, 11, 5452–5481. [Google Scholar] [CrossRef]
- Lábaj, J.; Slamenová, D.; Kosikova, B. Reduction of Genotoxic Effects of the Carcinogen N-Methyl-N’-Nitro-N-Nitrosoguanidine by Dietary Lignin in Mammalian Cells Cultured in Vitro. Nutr. Cancer 2003, 47, 95–103. [Google Scholar] [CrossRef]
- Barapatre, A.; Meena, A.S.; Mekala, S.; Das, A.; Jha, H. In Vitro Evaluation of Antioxidant and Cytotoxic Activities of Lignin Fractions Extracted from Acacia Nilotica. Int. J. Biol. Macromol. 2016, 86, 443–453. [Google Scholar] [CrossRef]
- Sato, S.; Mukai, Y.; Tokuoka, Y.; Mikame, K.; Funaoka, M.; Fujita, S. Effect of Lignin-Derived Lignophenols on Hepatic Lipid Metabolism in Rats Fed a High-Fat Diet. Environ. Toxicol. Pharmacol. 2012, 34, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Andrei, G.; Lisco, A.; Vanpouille, C.; Introini, A.; Balestra, E.; van den Oord, J.; Cihlar, T.; Perno, C.-F.; Snoeck, R.; Margolis, L.; et al. Topical Tenofovir, a Microbicide Effective against HIV, Inhibits Herpes Simplex Virus-2 Replication. Cell Host Microbe 2011, 10, 379–389. [Google Scholar] [CrossRef]
- Saluja, B.; Thakkar, J.N.; Li, H.; Desai, U.R.; Sakagami, M. Novel Low Molecular Weight Lignins as Potential Anti-Emphysema Agents: In Vitro Triple Inhibitory Activity against Elastase, Oxidation and Inflammation. Pulm. Pharmacol. Ther. 2013, 26, 296–304. [Google Scholar] [CrossRef]
- Vinardell, M.P.; Mitjans, M. Lignins and Their Derivatives with Beneficial Effects on Human Health. Int. J. Mol. Sci. 2017, 18, 1219. [Google Scholar] [CrossRef] [PubMed]
- Zherebker, A.Y.; Rukhovich, G.D.; Kharybin, O.N.; Fedoros, E.I.; Perminova, I.V.; Nikolaev, E.N. Fourier Transform Ion Cyclotron Resonance Mass Spectrometry for the Analysis of Molecular Composition and Batch-to-Batch Consistency of Plant-Derived Polyphenolic Ligands Developed for Biomedical Application. Rapid Commun. Mass Spectrom. 2020, 34, e8850. [Google Scholar] [CrossRef]
- Björklund, L.; Larsson, S.; Jönsson, L.J.; Reimann, E.; Nilvebrant, N.-O. Treatment with Lignin Residue: A Novel Method for Detoxification of Lignocellulose Hydrolysates. Appl. Biochem. Biotechnol. 2002, 98–100, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Vengerovskiĭ, A.I.; Golovina, E.L.; Burkova, V.N.; Saratikov, A.S. Enteric sorbents potentiate hepatoprotective effect of eplir in experimental toxic hepatitis. Eksp. Klin. Farmakol. 2001, 64, 46–48. [Google Scholar] [PubMed]
- Nebbioso, A.; Piccolo, A. Basis of a Humeomics Science: Chemical Fractionation and Molecular Characterization of Humic Biosuprastructures. Biomacromolecules 2011, 12, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Zhernov, Y.V.; Konstantinov, A.I.; Zherebker, A.; Nikolaev, E.; Orlov, A.; Savinykh, M.I.; Kornilaeva, G.V.; Karamov, E.V.; Perminova, I.V. Antiviral Activity of Natural Humic Substances and Shilajit Materials against HIV-1: Relation to Structure. Environ. Res. 2021, 193, 110312. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant Flavonoids: Classification, Distribution, Biosynthesis, and Antioxidant Activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.E.; Peltier-Heap, R.E.; Leveridge, M.; Annan, R.S.; Büttner, F.H.; Trost, M. Advances in High-Throughput Mass Spectrometry in Drug Discovery. EMBO Mol. Med. 2023, 15, e14850. [Google Scholar] [CrossRef] [PubMed]
- Orlov, A.; Semenov, S.; Rukhovich, G.; Sarycheva, A.; Kovaleva, O.; Semenov, A.; Ermakova, E.; Gubareva, E.; Bugrova, A.E.; Kononikhin, A.; et al. Hepatoprotective Activity of Lignin-Derived Polyphenols Dereplicated Using High-Resolution Mass Spectrometry, In Vivo Experiments, and Deep Learning. Int. J. Mol. Sci. 2022, 23, 16025. [Google Scholar] [CrossRef]
- Joseph, M.; Cross, T.J.S. Amyloidosis and Subacute Liver Failure. Gastroenterol. Hepatol. 2012, 8, 208–211. [Google Scholar]
- O’Grady, J.G. Acute Liver Failure. Postgrad. Med. J. 2005, 81, 148–154. [Google Scholar] [CrossRef]
- Shrestha, S.M.; Shrestha, S. Subacute Hepatic Failure: Its Possible Pathogenesis. Euroasian J. Hepato-Gastroenterol. 2012, 2, 41–46. [Google Scholar] [CrossRef]
- Scholten, D.; Trebicka, J.; Liedtke, C.; Weiskirchen, R. The Carbon Tetrachloride Model in Mice. Lab. Anim. 2015, 49, 4–11. [Google Scholar] [CrossRef]
- von Mering, C.; Huynen, M.; Jaeggi, D.; Schmidt, S.; Bork, P.; Snel, B. STRING: A Database of Predicted Functional Associations between Proteins. Nucleic Acids Res. 2003, 31, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Sarycheva, A.; Perminova, I.V.; Nikolaev, E.N.; Zherebker, A. Formulae Differences Commence a Database for Interlaboratory Studies of Natural Organic Matter. Environ. Sci. Technol. 2023, 57, 6238–6247. [Google Scholar] [CrossRef]
- Belousov, M.V.; Akhmedzhanov, R.R.; Zykova, M.V.; Gur’ev, A.M.; Yusubov, M.S. Hepatoprotective Properties of Native Humic Acids Isolated from Lowland Peat of Tomsk Region. Pharm. Chem. J. 2014, 48, 249–252. [Google Scholar] [CrossRef]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (Osteopontin): An Early Component of Type-1 (Cell-Mediated) Immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Zacchigna, S.; Oh, H.; Wilsch-Bräuninger, M.; Missol-Kolka, E.; Jászai, J.; Jansen, S.; Tanimoto, N.; Tonagel, F.; Seeliger, M.; Huttner, W.B.; et al. Loss of the Cholesterol-Binding Protein Prominin-1/CD133 Causes Disk Dysmorphogenesis and Photoreceptor Degeneration. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 2297–2308. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, N.; Schäffer, A.A.; Liu, X.; Zhao, Z.; Elliott, G.; Garrett, L.; Choi, N.T.; Wang, Y.; Wang, Y.; et al. Mutations in COMP Cause Familial Carpal Tunnel Syndrome. Nat. Commun. 2020, 11, 3642. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, A.M.; Heinrich, G.; Dai, T.; Bowman, T.A.; Patel, P.R.; Lee, S.J.; Hong, E.-G.; Jung, D.Y.; Assmann, A.; Kulkarni, R.N.; et al. Carcinoembryonic Antigen-Related Cell Adhesion Molecule 1: A Link between Insulin and Lipid Metabolism. Diabetes 2008, 57, 2296–2303. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, S.; Chen, Z.; Baker, K.; Chen, L.; Huang, Y.-H.; Olszak, T.; Zeissig, S.; Wang, J.H.; Mandelboim, O.; Beauchemin, N.; et al. CEACAM1 on Activated NK Cells Inhibits NKG2D-Mediated Cytolytic Function and Signaling. Eur. J. Immunol. 2013, 43, 2473–2483. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Pan, H.; Shively, J.E. CEACAM1 Negatively Regulates IL-1β Production in LPS Activated Neutrophils by Recruiting SHP-1 to a SYK-TLR4-CEACAM1 Complex. PLoS Pathog. 2012, 8, e1002597. [Google Scholar] [CrossRef]
- Friedrichs, B.; Tepel, C.; Reinheckel, T.; Deussing, J.; von Figura, K.; Herzog, V.; Peters, C.; Saftig, P.; Brix, K. Thyroid Functions of Mouse Cathepsins B, K, and L. J. Clin. Investig. 2003, 111, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.A.; Ribeiro, A.L.C.; Costa, B.R.C.; Vago, J.P.; Lima, K.M.; Carneiro, F.S.; Ortiz, M.M.O.; Lima, G.L.N.; Carmo, A.A.F.; Rocha, R.M.; et al. Plasmin and Plasminogen Induce Macrophage Reprogramming and Regulate Key Steps of Inflammation Resolution via Annexin A1. Blood 2017, 129, 2896–2907. [Google Scholar] [CrossRef] [PubMed]
- Croessmann, S.; Wong, H.Y.; Zabransky, D.J.; Chu, D.; Mendonca, J.; Sharma, A.; Mohseni, M.; Rosen, D.M.; Scharpf, R.B.; Cidado, J.; et al. NDRG1 Links P53 with Proliferation-Mediated Centrosome Homeostasis and Genome Stability. Proc. Natl. Acad. Sci. USA 2015, 112, 11583–11588. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xu, L.; Chu, H.; Peng, J.; Sacharidou, A.; Hsieh, H.-H.; Weinstock, A.; Khan, S.; Ma, L.; Durán, J.G.B.; et al. Macrophage-to-Endothelial Cell Crosstalk by the Cholesterol Metabolite 27HC Promotes Atherosclerosis in Male Mice. Nat. Commun. 2023, 14, 4101. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, V.G.; Rubtsova, M.Y.; Andreeva, I.P.; Presnova, G.V.; Mikhnevich, T.A.; Yiming, S.; Kravtsova, D.S.; Konstantinov, A.I.; Arutyunyan, D.A.; Perminova, I.V. Screening of Inhibitory Activity of Narrow Fractions of Humic Substances towards Different Types of Class A β-Lactamases. Appl. Biochem. Microbiol. 2024, 60, 1196–1208. [Google Scholar] [CrossRef]
- Semenov, A.L.; Tyndyk, M.L.; Von, J.D.; Ermakova, E.D.; Dorofeeva, A.A.; Tumanyan, I.A.; Radetskaya, E.A.; Yurova, M.N.; Zherebker, A.; Gorbunov, A.Y.; et al. Effects of Isoflavone-Rich NADES Extract of Pueraria Lobata Roots and Astaxanthin-Rich Phaffia Rhodozyma Extract on Prostate Carcinogenesis in Rats. Plants 2023, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Badun, G.A.; Chernysheva, M.G.; Zhernov, Y.V.; Poroshina, A.S.; Smirnov, V.V.; Pigarev, S.E.; Mikhnevich, T.A.; Volkov, D.S.; Perminova, I.V.; Fedoros, E.I. A Use of Tritium-Labeled Peat Fulvic Acids and Polyphenolic Derivatives for Designing Pharmacokinetic Experiments on Mice. Biomedicines 2021, 9, 1787. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, J.; Shen, F.; Qin, Z.; Jiang, M.; Zhu, J.; Wang, Z.; Zhou, J.; Fu, Y.; Chen, X.; et al. Systems Pharmacology Analysis of Synergy of TCM: An Example Using Saffron Formula. Sci. Rep. 2018, 8, 380. [Google Scholar] [CrossRef]
- Semenov, A.L.; Gubareva, E.A.; Ermakova, E.D.; Dorofeeva, A.A.; Tumanyan, I.A.; Radetskaya, E.A.; Yurova, M.N.; Aboushanab, S.A.; Kanwugu, O.N.; Fedoros, E.I.; et al. Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance. Plants 2021, 10, 2735. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, Y.; Yang, L.; Wang, K.; Zhang, X.; Zong, Y.; Ding, Y.; Wang, C.; Zhang, L.; Ji, G. Resveratrol Alleviates FFA and CCl4 Induced Apoptosis in HepG2 Cells via Restoring Endoplasmic Reticulum Stress. Oncotarget 2017, 8, 43799–43809. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Waskom, M. Seaborn: Statistical Data Visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- McKinney, W. Data Structures for Statistical Computing in Python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; Volume 445, pp. 51–56. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar] [CrossRef]
- Tonoyan, N.M.; Chagovets, V.V.; Starodubtseva, N.L.; Tokareva, A.O.; Chingin, K.; Kozachenko, I.F.; Adamyan, L.V.; Frankevich, V.E. Alterations in Lipid Profile upon Uterine Fibroids and Its Recurrence. Sci. Rep. 2021, 11, 11447. [Google Scholar] [CrossRef] [PubMed]
- Kessner, D.; Chambers, M.; Burke, R.; Agus, D.; Mallick, P. ProteoWizard: Open Source Software for Rapid Proteomics Tools Development. Bioinformatics 2008, 24, 2534–2536. [Google Scholar] [CrossRef]
- Goloborodko, A.A.; Levitsky, L.I.; Ivanov, M.V.; Gorshkov, M. V Pyteomics-a Python Framework for Exploratory Data Analysis and Rapid Software Prototyping in Proteomics. J. Am. Soc. Mass Spectrom. 2013, 24, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Volikov, A.; Rukhovich, G.; Perminova, I. V NOMspectra: An Open-Source Python Package for Processing High Resolution Mass Spectrometry Data on Natural Organic Matter. J. Am. Soc. Mass Spectrom. 2023, 34, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, A.T.; Kourtchev, I.; Bortolini, C.; Fuller, S.J.; Giorio, C.; Popoola, O.A.M.; Bogialli, S.; Tapparo, A.; Jones, R.L.; Kalberer, M. A New Processing Scheme for Ultra-High Resolution Direct Infusion Mass Spectrometry Data. Atmos. Environ. 2018, 178, 129–139. [Google Scholar] [CrossRef]
- Kozhinov, A.N.; Zhurov, K.O.; Tsybin, Y.O. Iterative Method for Mass Spectra Recalibration via Empirical Estimation of the Mass Calibration Function for Fourier Transform Mass Spectrometry-Based Petroleomics. Anal. Chem. 2013, 85, 6437–6445. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brzhozovskiy, A.G.; Semenov, S.D.; Zherebker, A.Y.; Bugrova, A.E.; Yurova, M.N.; Zhernov, Y.V.; Kovaleva, O.A.; Semenov, A.L.; Abroskin, D.P.; Kruglov, S.S.; et al. Hepatoprotective Activity of Nature-Derived Polyphenols Studied by Mass Spectrometry Based Multi-OMICS Approach. Int. J. Mol. Sci. 2025, 26, 1604. https://doi.org/10.3390/ijms26041604
Brzhozovskiy AG, Semenov SD, Zherebker AY, Bugrova AE, Yurova MN, Zhernov YV, Kovaleva OA, Semenov AL, Abroskin DP, Kruglov SS, et al. Hepatoprotective Activity of Nature-Derived Polyphenols Studied by Mass Spectrometry Based Multi-OMICS Approach. International Journal of Molecular Sciences. 2025; 26(4):1604. https://doi.org/10.3390/ijms26041604
Chicago/Turabian StyleBrzhozovskiy, Alexander G., Savva D. Semenov, Alexander Y. Zherebker, Anna E. Bugrova, Maria N. Yurova, Yury V. Zhernov, Oxana A. Kovaleva, Alexander L. Semenov, Dmitry P. Abroskin, Stepan S. Kruglov, and et al. 2025. "Hepatoprotective Activity of Nature-Derived Polyphenols Studied by Mass Spectrometry Based Multi-OMICS Approach" International Journal of Molecular Sciences 26, no. 4: 1604. https://doi.org/10.3390/ijms26041604
APA StyleBrzhozovskiy, A. G., Semenov, S. D., Zherebker, A. Y., Bugrova, A. E., Yurova, M. N., Zhernov, Y. V., Kovaleva, O. A., Semenov, A. L., Abroskin, D. P., Kruglov, S. S., Fedoros, E. I., Kononikhin, A. S., & Nikolaev, E. N. (2025). Hepatoprotective Activity of Nature-Derived Polyphenols Studied by Mass Spectrometry Based Multi-OMICS Approach. International Journal of Molecular Sciences, 26(4), 1604. https://doi.org/10.3390/ijms26041604