
The Impact of Nanoparticles and Molecular Forms of TiO2 on the Rhizosphere of Plants in the Example of Common Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions


Abstract
1. Introduction
2. Results
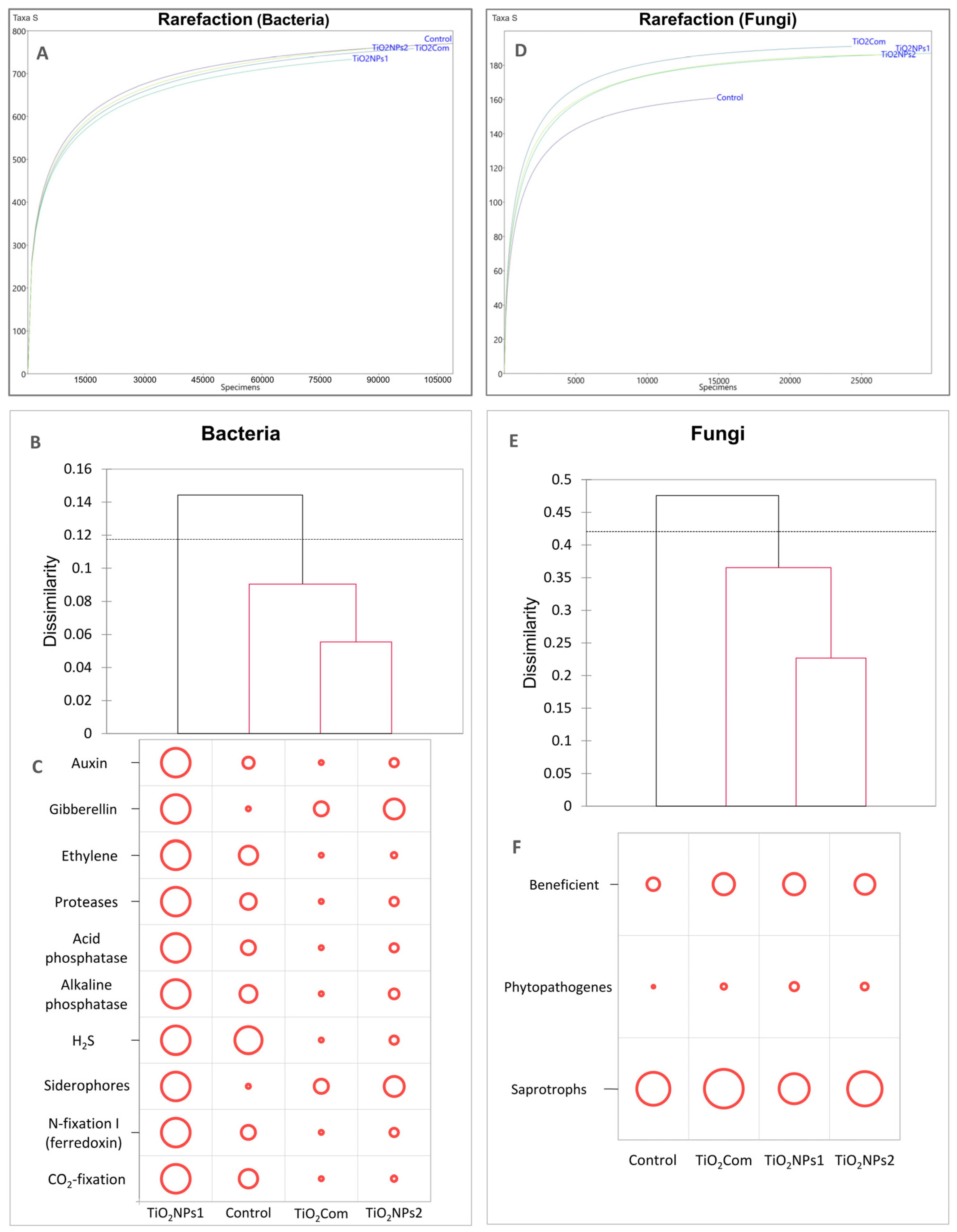
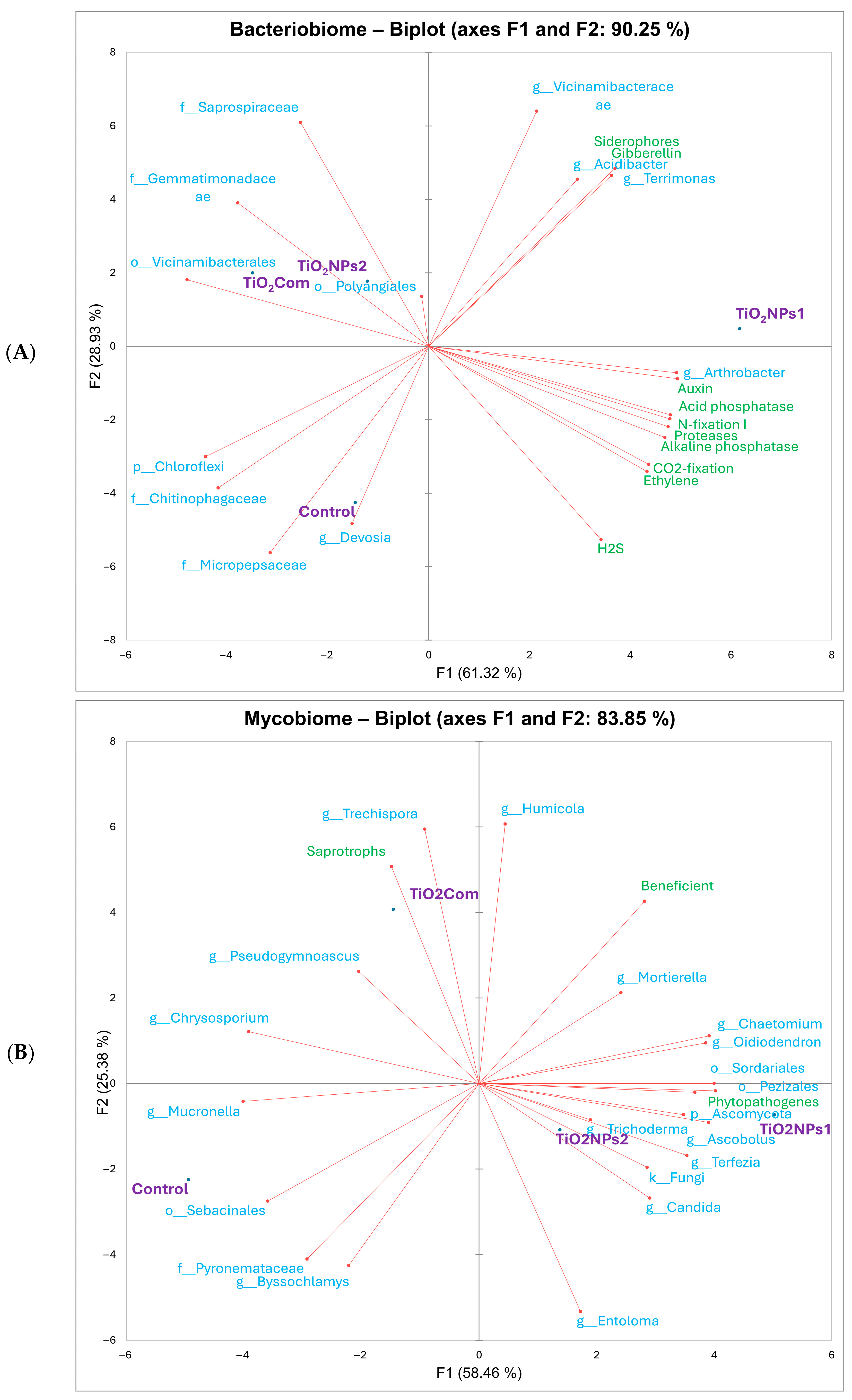
2.1. Bacteriobiome Characteristics
2.2. Mycobiome Characteristics
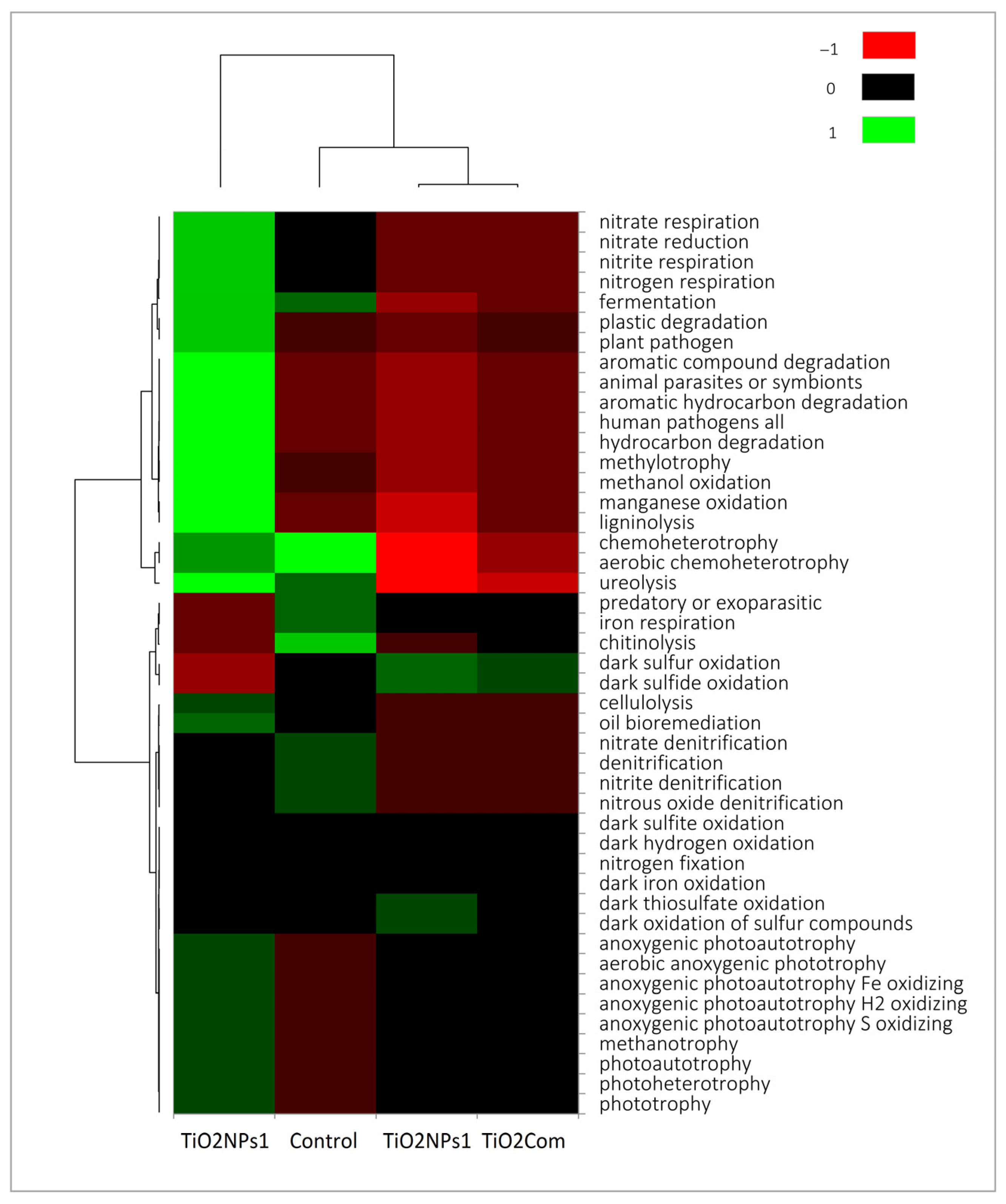
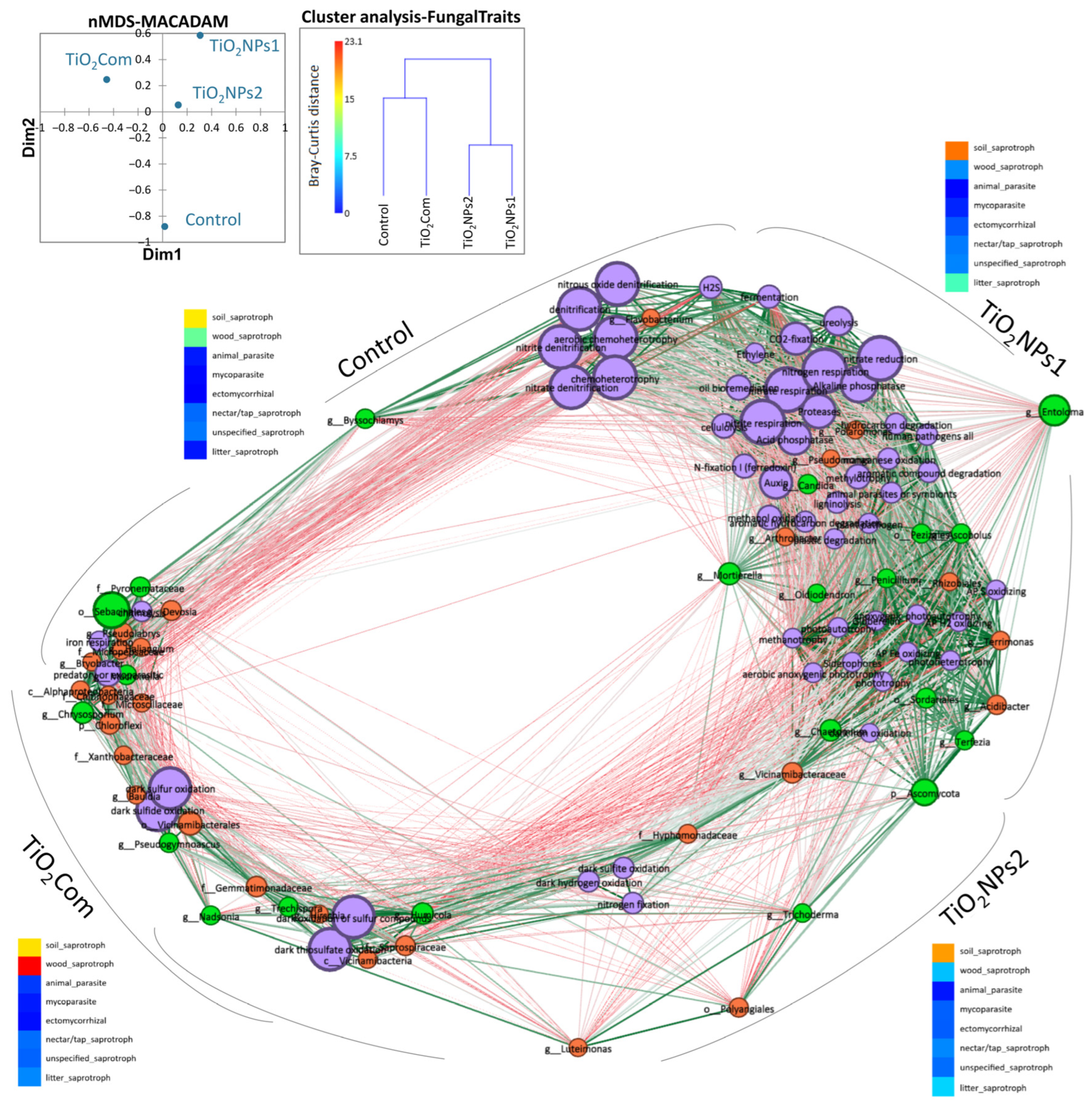
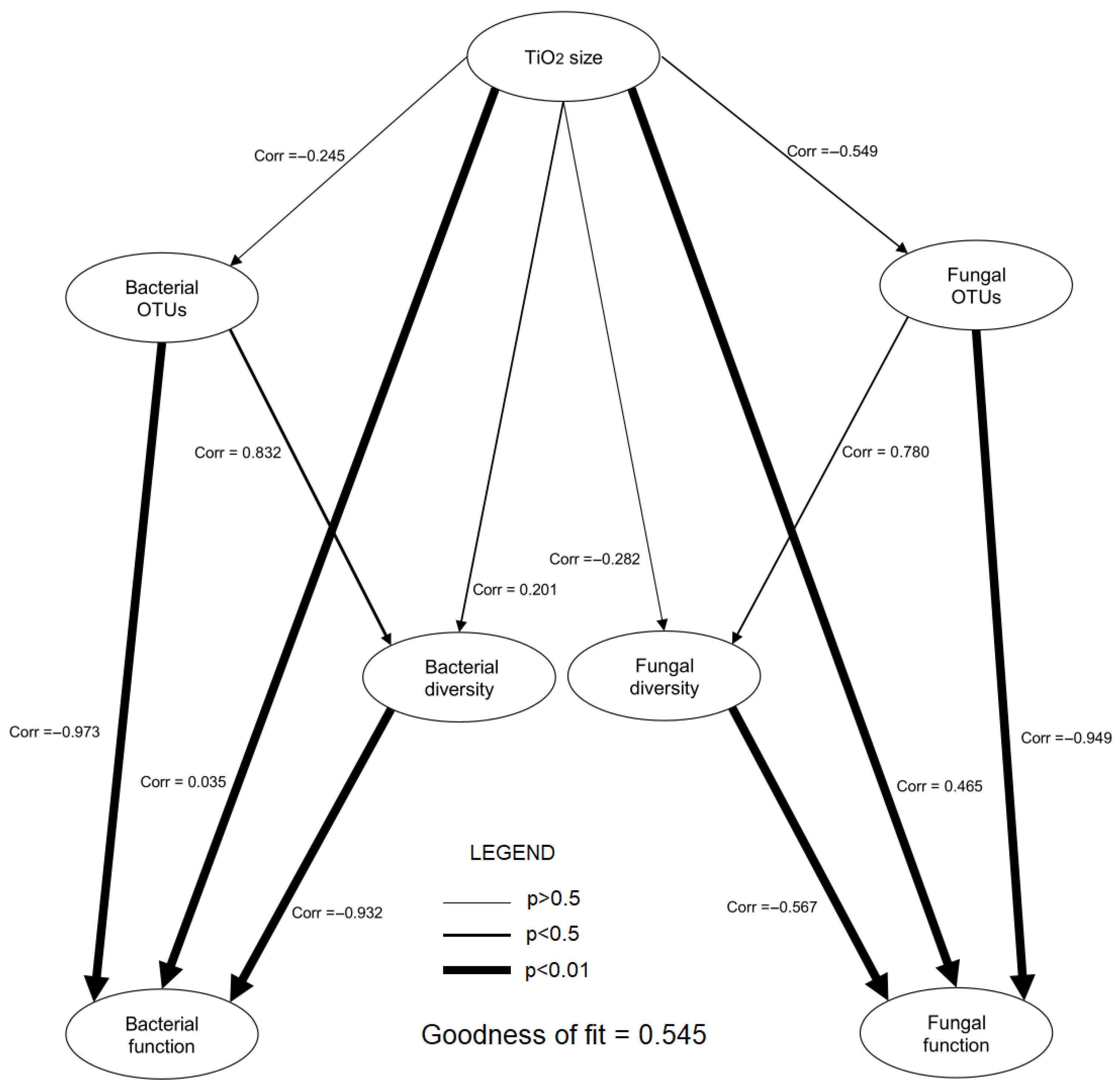
2.3. Predicting the Function of Microbiome, Network and PLS-PM Analysis
3. Discussion
3.1. Bacteriobiome
3.2. Mycobiome
3.3. Relationship Between Changes in Microbiomes and Rhizobiome Metabolism
4. Materials and Methods
4.1. Experimental Setup
4.2. Sequencing
4.3. Statistical Calculation and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, W.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of Titanium Dioxide Nanoparticles with Soil Components and Plants: Current Knowledge and Future Research Needs—A Critical Review. Environ. Sci. Nano 2018, 5, 257–278. [Google Scholar] [CrossRef]
- He, X.; Deng, H.; Hwang, H. The Current Application of Nanotechnology in Food and Agriculture. J. Food Drug Anal. 2019, 27, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Ruraż, K.; Kosewska, O.; Oćwieja, M.; Gorczyca, A. The Impact of Various Forms of Silver Nanoparticles on the Rhizosphere of Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions. Sci. Total Environ. 2024, 914, 169824. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Borsuk-Stanulewicz, M.; Purwin, C.; Kosewska, O.; Oćwieja, M. The Effect of Different Forms of Titanium Dioxide on the Yield, Chemical and Microbiological Parameters of Perennial Ryegrass (Lolium perenne L.) Herbage and Silage. Agriculture 2023, 13, 1588. [Google Scholar] [CrossRef]
- Cox, A.; Venkatachalam, P.; Sahi, S.; Sharma, N. Silver and Titanium Dioxide Nanoparticle Toxicity in Plants: A Review of Current Research. Plant Physiol. Biochem. 2016, 107, 147–163. [Google Scholar] [CrossRef]
- Gojznikar, J.; Zdravković, B.; Vidak, M.; Leskošek, B.; Ferk, P. TiO2 Nanoparticles and Their Effects on Eukaryotic Cells: A Double-Edged Sword. Int. J. Mol. Sci. 2022, 23, 12353. [Google Scholar] [CrossRef] [PubMed]
- Ajiboye, T.T.; Ajiboye, T.O.; Babalola, O.O. Impacts of Binary Oxide Nanoparticles on the Soybean Plant and Its Rhizosphere, Associated Phytohormones, and Enzymes. Molecules 2023, 28, 1326. [Google Scholar] [CrossRef]
- Moll, J.; Klingenfuss, F.; Widmer, F.; Gogos, A.; Bucheli, T.D.; Hartmann, M.; van der Heijden, M.G.A. Effects of Titanium Dioxide Nanoparticles on Soil Microbial Communities and Wheat Biomass. Soil Biol. Biochem. 2017, 111, 85–93. [Google Scholar] [CrossRef]
- Gorczyca, A.; Przemieniecki, S.W.; Kurowski, T.; Oćwieja, M. Early Plant Growth and Bacterial Community in Rhizoplane of Wheat and Flax Exposed to Silver and Titanium Dioxide Nanoparticles. Environ. Sci. Pollut. Res. 2018, 25, 33820–33826. [Google Scholar] [CrossRef] [PubMed]
- Ostadi, A.; Javanmard, A.; Amani Machiani, M.; Sadeghpour, A.; Maggi, F.; Nouraein, M.; Morshedloo, M.R.; Hano, C.; Lorenzo, J.M. Co-Application of TiO2 Nanoparticles and Arbuscular Mycorrhizal Fungi Improves Essential Oil Quantity and Quality of Sage (Salvia officinalis L.) in Drought Stress Conditions. Plants 2022, 11, 1659. [Google Scholar] [CrossRef]
- Asadishad, B.; Chahal, S.; Akbari, A.; Cianciarelli, V.; Azodi, M.; Ghoshal, S.; Tufenkji, N. Amendment of Agricultural Soil with Metal Nanoparticles: Effects on Soil Enzyme Activity and Microbial Community Composition. Environ. Sci. Technol. 2018, 52, 1908–1918. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.; Pietrasiak, N.; Situ, S.; Abenojar, E.; Porche, M.; Kraj, P.; Lakliang, Y.; Samia, A. Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes. Int. J. Mol. Sci. 2015, 16, 23630–23650. [Google Scholar] [CrossRef] [PubMed]
- Simonin, M.; Richaume, A.; Guyonnet, J.P.; Dubost, A.; Martins, J.M.F.; Pommier, T. Titanium Dioxide Nanoparticles Strongly Impact Soil Microbial Function by Affecting Archaeal Nitrifiers. Sci. Rep. 2016, 6, 33643. [Google Scholar] [CrossRef]
- Ahmed, A.; He, P.; He, P.; Wu, Y.; He, Y.; Munir, S. Environmental Effect of Agriculture-Related Manufactured Nano-Objects on Soil Microbial Communities. Environ. Int. 2023, 173, 107819. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Zahra, N.; Lang, T.; Zain, M.; Raza, M.; Shakoor, N.; Adeel, M.; Zhou, H. Integrating Nanotechnology with Plant Microbiome for Next-Generation Crop Health. Plant Physiol. Biochem. 2023, 196, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Zehlike, L.; Peters, A.; Ellerbrock, R.H.; Degenkolb, L.; Klitzke, S. Aggregation of TiO2 and Ag Nanoparticles in Soil Solution—Effects of Primary Nanoparticle Size and Dissolved Organic Matter Characteristics. Sci. Total Environ. 2019, 688, 288–298. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere Bacteriome Structure and Functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and Applications of Plant Growth Promoting Rhizobacteria: Current Perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Kurowski, T.P.; Kotlarz, K.; Krawczyk, K.; Damszel, M.; Pszczółkowska, A.; Kacprzak-Siuda, K.; Chareńska, A.; Mastalerz, J. Bacteria Isolated from Treated Wastewater for Biofertilization and Crop Protection Against fusarium spp. Pathogens. J. Soil Sci. Plant Nutr. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Grün, A.-L.; Manz, W.; Kohl, Y.L.; Meier, F.; Straskraba, S.; Jost, C.; Drexel, R.; Emmerling, C. Impact of Silver Nanoparticles (AgNP) on Soil Microbial Community Depending on Functionalization, Concentration, Exposure Time, and Soil Texture. Environ. Sci. Eur. 2019, 31, 15. [Google Scholar] [CrossRef]
- Khan, S.A.; Ahmad, A. Phase, Size and Shape Transformation by Fungal Biotransformation of Bulk TiO2. Chem. Eng. J. 2013, 230, 367–371. [Google Scholar] [CrossRef]
- Ważny, R.; Jędrzejczyk, R.J.; Domka, A.; Pliszko, A.; Kosowicz, W.; Githae, D.; Rozpądek, P. How Does Metal Soil Pollution Change the Plant Mycobiome? Environ. Microbiol. 2023, 25, 2913–2930. [Google Scholar] [CrossRef] [PubMed]
- Põlme, S.; Abarenkov, K.; Henrik Nilsson, R.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A User-Friendly Traits Database of Fungi and Fungus-like Stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Binkauskiene, E.; Lugauskas, A.; Krunks, M.; Acik, I.O.; Jasulaitiene, V.; Saduikis, G. Interaction of Chrysosporium Merdarium with Titanium Oxide Surface. Synth. Met. 2010, 160, 906–910. [Google Scholar] [CrossRef]
- Chinnaperumal, K.; Govindasamy, B.; Paramasivam, D.; Dilipkumar, A.; Dhayalan, A.; Vadivel, A.; Sengodan, K.; Pachiappan, P. Bio-Pesticidal Effects of Trichoderma viride Formulated Titanium Dioxide Nanoparticle and Their Physiological and Biochemical Changes on Helicoverpa armigera (Hub.). Pestic. Biochem. Physiol. 2018, 149, 26–36. [Google Scholar] [CrossRef]
- Ramírez-Valdespino, C.A.; Orrantia-Borunda, E. Trichoderma and Nanotechnology in Sustainable Agriculture: A Review. Front. Fungal Biol. 2021, 2, 764675. [Google Scholar] [CrossRef]
- Pasquoto-Stigliani, T.; Guilger-Casagrande, M.; Campos, E.V.R.; Germano-Costa, T.; Bilesky-José, N.; Migliorini, B.B.; Feitosa, L.O.; Sousa, B.T.; de Oliveira, H.C.; Fraceto, L.F.; et al. Titanium Biogenic Nanoparticles to Help the Growth of Trichoderma Harzianum to Be Used in Biological Control. J. Nanobiotechnol. 2023, 21, 166. [Google Scholar] [CrossRef]
- Ayad, F.; Matallah-Boutiba, A.; Rouane–Hacene, O.; Bouderbala, M.; Boutiba, Z. Tolerance of Trichoderma sp. to Heavy Metals and Its Antifungal Activity in Algerian Marine Environment. J. Pure Appl. Microbiol. 2018, 12, 855–870. [Google Scholar] [CrossRef]
- Maurya, S.; Rashk-E-Eram; Naik, S.K.; Choudhary, J.S.; Kumar, S. Heavy Metals Scavenging Potential of Trichoderma Asperellum and Hypocrea Nigricans Isolated from Acid Soil of Jharkhand. Indian J. Microbiol. 2019, 59, 27–38. [Google Scholar] [CrossRef]
- Srinivasan, R.; Prabhu, G.; Prasad, M.; Mishra, M.; Chaudhary, M.; Srivastava, R. Penicillium. In Beneficial Microbes in Agro-Ecology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 651–667. [Google Scholar]
- Przemieniecki, S.W.; Kosewska, A.; Purwin, C.; Zapałowska, A.; Mastalerz, J.; Kotlarz, K.; Kolaczek, K. Biometric, Chemical, and Microbiological Evaluation of Common Wheat (Triticum aestivum L.) Seedlings Fertilized with Mealworm (Tenebrio molitor L.) Larvae Meal. Appl. Soil Ecol. 2021, 167, 104037. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/bcdecec7-f45f-4dc5-beb1-97022d29fab4/content (accessed on 10 November 2024).
- Przemieniecki, S.W.; Oćwieja, M.; Ciesielski, S.; Halecki, W.; Matras, E.; Gorczyca, A. Chemical Structure of Stabilizing Layers of Negatively Charged Silver Nanoparticles as an Effector of Shifts in Soil Bacterial Microbiome under Short-Term Exposure. Int. J. Environ. Res. Public Health 2022, 19, 14438. [Google Scholar] [CrossRef] [PubMed]
- Ruraż, K.; Przemieniecki, S.W.; Piwowarczyk, R. Interspecies and Temporal Dynamics of Bacterial and Fungal Microbiomes of Pistil Stigmas in Flowers in Holoparasitic Plants of the Orobanche Series Alsaticae (Orobanchaceae). Sci. Rep. 2023, 13, 6749. [Google Scholar] [CrossRef] [PubMed]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Przemieniecki, S.W.; Damszel, M.; Ciesielski, S.; Kubiak, K.; Mastalerz, J.; Sierota, Z.; Gorczyca, A. Bacterial Microbiome in Armillaria Ostoyae Rhizomorphs Inhabiting the Root Zone during Progressively Dying Scots Pine. Appl. Soil Ecol. 2021, 164, 103929. [Google Scholar] [CrossRef]
- Kurowski, T.; Przemieniecki, S.; Grabowski, K.; Damszel, M.; Kwiatkowska, E. Change in Microbiological Conditions of Lawn Grass Root Zones as a Result of Fertilization with Sewage Sludge. Pol. J. Environ. Stud. 2016, 25, 2015–2026. [Google Scholar] [CrossRef] [PubMed]
- Le Boulch, M.; Déhais, P.; Combes, S.; Pascal, G. The MACADAM Database: A MetAboliC PAthways DAtabase for Microbial Taxonomic Groups for Mining Potential Metabolic Capacities of Archaeal and Bacterial Taxonomic Groups. Database 2019, 2019, baz049. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Billington, R.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Midford, P.E.; Ong, Q.; Ong, W.K.; et al. The MetaCyc Database of Metabolic Pathways and Enzymes. Nucleic Acids Res. 2018, 46, D633–D639. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Lumivero XLSTAT Basic Solutions. Available online: https://www.xlstat.com/en/solutions/basic (accessed on 25 November 2024).
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks Visualization and Exploration of Large Graphs. Available online: https://ojs.aaai.org/index.php/ICWSM/article/view/13937 (accessed on 25 November 2024).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU ID—BACTERIA | Control * | TiO2Com | TiO2NPs1 | TiO2 NPs2 |
|---|---|---|---|---|
| o__Vicinamibacterales | 5.2 | 5.8 | 4.5 | 5.3 |
| f__Gemmatimonadaceae | 2.9 | 3.0 | 2.8 | 2.9 |
| g__Vicinamibacteraceae | 2.1 | 3.0 | 3.2 | 2.9 |
| g__Devosia | 2.5 | 2.2 | 2.2 | 2.4 |
| p__Chloroflexi_KD4-96 | 2.1 | 1.8 | 1.3 | 1.6 |
| f__Saprospiraceae | 2.2 | 2.3 | 2.2 | 2.2 |
| o__Polyangiales_BIrii41 | 1.7 | 1.5 | 1.6 | 2.1 |
| g__Acidibacter | 1.4 | 1.6 | 2.1 | 2.2 |
| f__Xanthobacteraceae | 1.4 | 1.4 | 1.2 | 1.4 |
| g__Haliangium | 1.3 | 1.1 | 1.0 | 1.1 |
| f__Chitinophagaceae | 1.5 | 1.1 | 0.5 | 1.1 |
| f__Hyphomonadaceae | 1.3 | 1.4 | 1.4 | 1.4 |
| f__Micropepsaceae | 1.4 | 1.0 | 0.7 | 1.0 |
| c__Vicinamibacteria | 1.2 | 1.4 | 1.2 | 1.2 |
| c__Alphaproteobacteria | 1.3 | 1.2 | 1.1 | 1.2 |
| g__Bauldia | 1.0 | 1.1 | 0.9 | 1.0 |
| g__Terrimonas | 1.0 | 1.2 | 1.5 | 1.4 |
| g__Bryobacter | 1.1 | 1.0 | 0.8 | 0.9 |
| p__Chloroflexi_JG30 | 1.1 | 1.2 | 0.8 | 1.0 |
| g__Pseudolabrys | 1.1 | 0.7 | 0.5 | 0.8 |
| g__Luteimonas | 0.9 | 0.9 | 0.9 | 1.0 |
| f__Rhizobiales | 0.7 | 0.9 | 1.1 | 1.0 |
| f__Microscillaceae | 1.0 | 0.9 | 0.8 | 1.0 |
| g__Hirschia | 0.8 | 1.0 | 0.9 | 0.8 |
| g__Arthrobacter | 0.8 | 0.5 | 1.7 | 0.6 |
| g__Flavobacterium | 1.0 | 0.4 | 0.8 | 0.4 |
| g__Polaromonas | 0.6 | 0.4 | 1.2 | 0.5 |
| g__Pseudomonas | 0.3 | 0.1 | 1.2 | 0.2 |
| OTUs | 771 | 759 | 735 | 760 |
| Simpson’s dominance (λ) | 0.010 | 0.011 | 0.010 | 0.011 |
| Shannon diversity (H′) | 5.285 | 5.239 | 5.256 | 5.247 |
| Pielou’s evenness (J′) | 0.795 | 0.790 | 0.796 | 0.791 |
| OTU ID—FUNGI | Control | TiO2Com | TiO2NPs1 | TiO2NPs2 |
|---|---|---|---|---|
| o__Sebacinales | 29.0 | 11.3 * | 5.5 * | 13.6 * |
| g__Entoloma | 10.7 | 8.3 | 11.4 | 11.9 |
| p__Ascomycota | 3.0 | 5.6 | 10.0 * | 11.5 * |
| g__Mortierella | 3.2 | 4.3 | 5.5 | 2.8 |
| g__Chrysosporium | 6.8 | 5.5 | 0.3 * | 1.9 * |
| g__Humicola | 0.6 | 7.7 * | 3.1 | 2.0 |
| g__Chaetomium | 0.4 | 3.1 | 5.4 * | 4.3 |
| o__Sordariales | 0.7 | 1.9 | 3.8 | 3.0 |
| f__Pyronemataceae | 4.3 | 1.1 | 1.0 | 2.6 |
| g__Mucronella | 3.4 | 2.4 | 0.9 | 1.9 |
| g__Byssochlamys | 2.7 | 1.7 | 2.0 | 1.8 |
| g__Candida | 1.9 | 1.6 | 2.7 | 1.8 |
| g__Ascobolus | 0.3 | 1.0 | 4.4 | 2.1 |
| g__Pseudogymnoascus | 1.8 | 2.6 | 0.6 | 2.6 |
| g__Trechispora | 0.0 | 7.5 * | 0.0 | 0.0 |
| o__Pezizales | 0.8 | 1.3 | 3.2 | 1.4 |
| g__Nadsonia | 1.3 | 1.7 | 1.0 | 2.3 |
| g__Terfezia | 0.0 | 0.4 | 2.6 | 2.8 |
| g__Penicillium | 0.9 | 1.3 | 2.0 | 1.5 |
| g__Trichoderma | 0.3 | 0.8 | 1.1 | 2.9 |
| g__Oidiodendron | 0.3 | 1.1 | 2.1 | 1.1 |
| g__Phialemonium | 0.8 | 1.8 | 0.1 | 0.7 |
| k__Fungi | 0.3 | 0.3 | 1.0 | 1.6 |
| OTUs | 161 | 191 | 187 | 186 |
| Simpson’s dominance (λ) | 0.111 | 0.045 | 0.043 | 0.057 |
| Shannon diversity (H′) | 3.224 | 3.750 | 3.743 | 3.618 |
| Pielou’s evenness (J′) | 0.634 | 0.714 | 0.715 | 0.692 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przemieniecki, S.W.; Damszel, M.; Kosewska, O. The Impact of Nanoparticles and Molecular Forms of TiO2 on the Rhizosphere of Plants in the Example of Common Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions. Int. J. Mol. Sci. 2025, 26, 685. https://doi.org/10.3390/ijms26020685
Przemieniecki SW, Damszel M, Kosewska O. The Impact of Nanoparticles and Molecular Forms of TiO2 on the Rhizosphere of Plants in the Example of Common Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions. International Journal of Molecular Sciences. 2025; 26(2):685. https://doi.org/10.3390/ijms26020685
Chicago/Turabian StylePrzemieniecki, Sebastian Wojciech, Marta Damszel, and Olga Kosewska. 2025. "The Impact of Nanoparticles and Molecular Forms of TiO2 on the Rhizosphere of Plants in the Example of Common Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions" International Journal of Molecular Sciences 26, no. 2: 685. https://doi.org/10.3390/ijms26020685
APA StylePrzemieniecki, S. W., Damszel, M., & Kosewska, O. (2025). The Impact of Nanoparticles and Molecular Forms of TiO2 on the Rhizosphere of Plants in the Example of Common Wheat (Triticum aestivum L.)—Shifts in Microbiome Structure and Predicted Microbial Metabolic Functions. International Journal of Molecular Sciences, 26(2), 685. https://doi.org/10.3390/ijms26020685