
A Host Cell Vector Model for Analyzing Viral Protective Antigens and Host Immunity
 , , , , , and
, , , , , and
Abstract

1. Introduction
2. Results
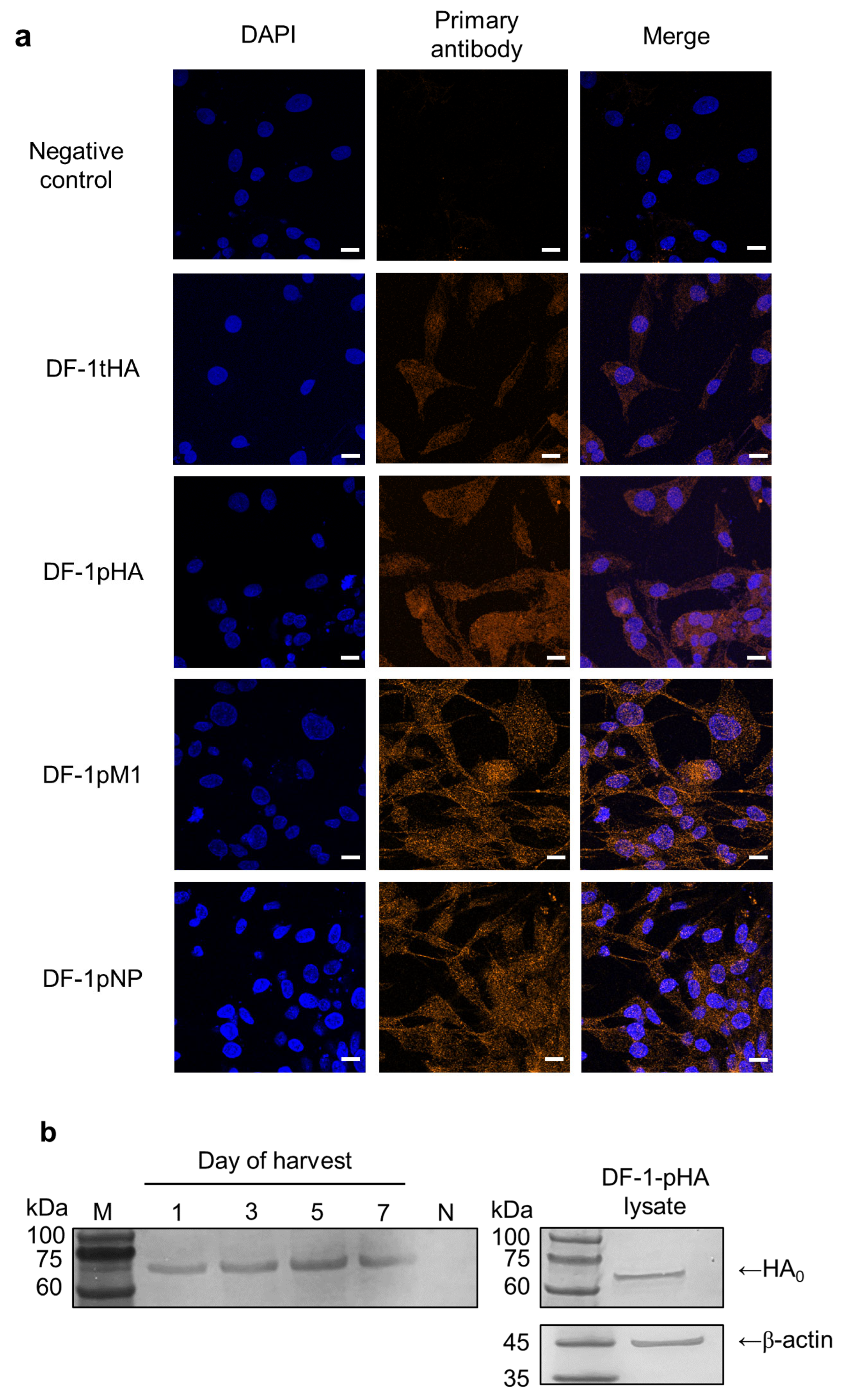
2.1. DF-1 Cell Lines Are Adequate for Efficient Expression of Transient and Permanent IAV Genes
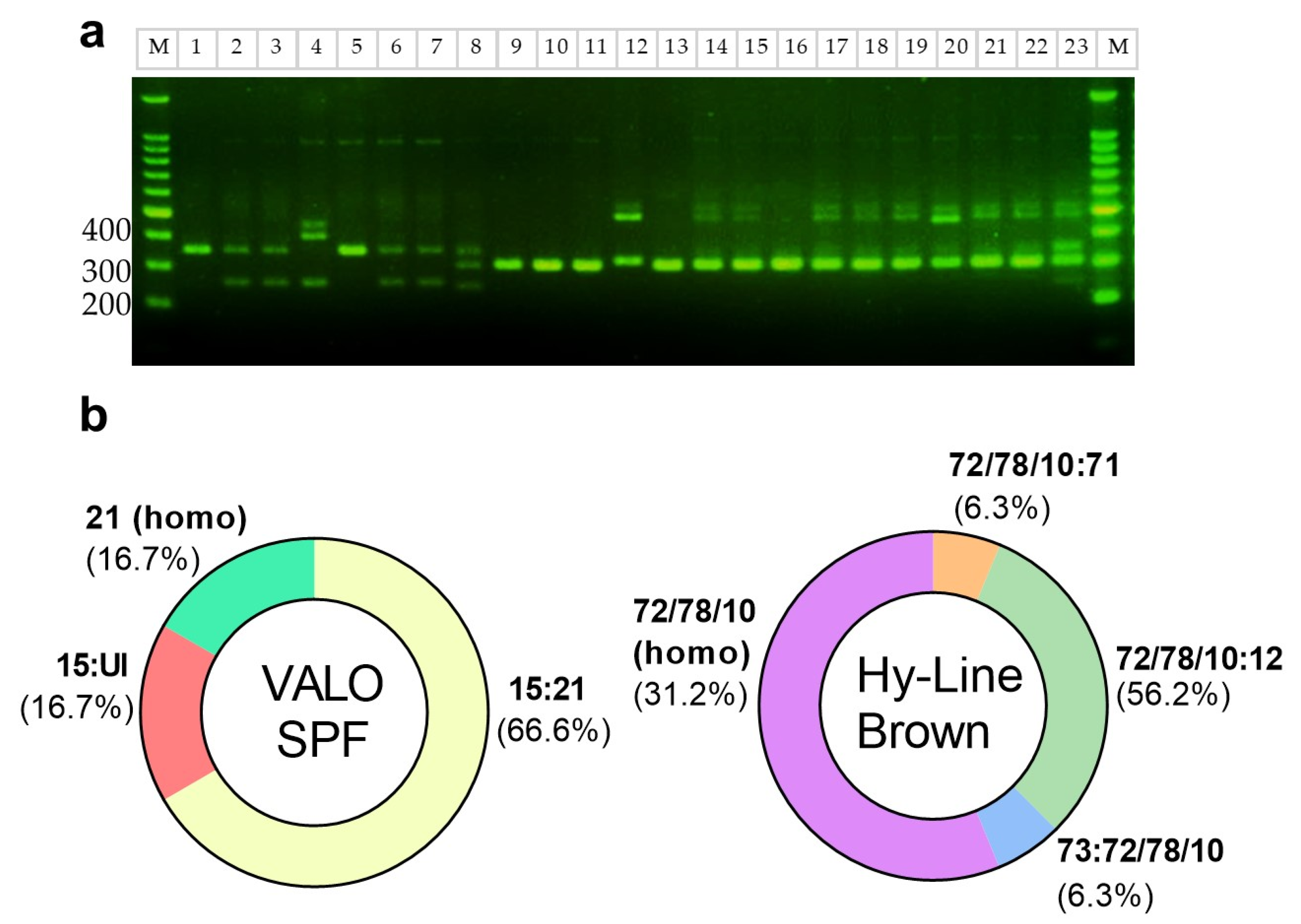
2.2. MHC Class 1 Molecule Haplotype of DF-1 Cells Matches with Most VALO SPF Chickens
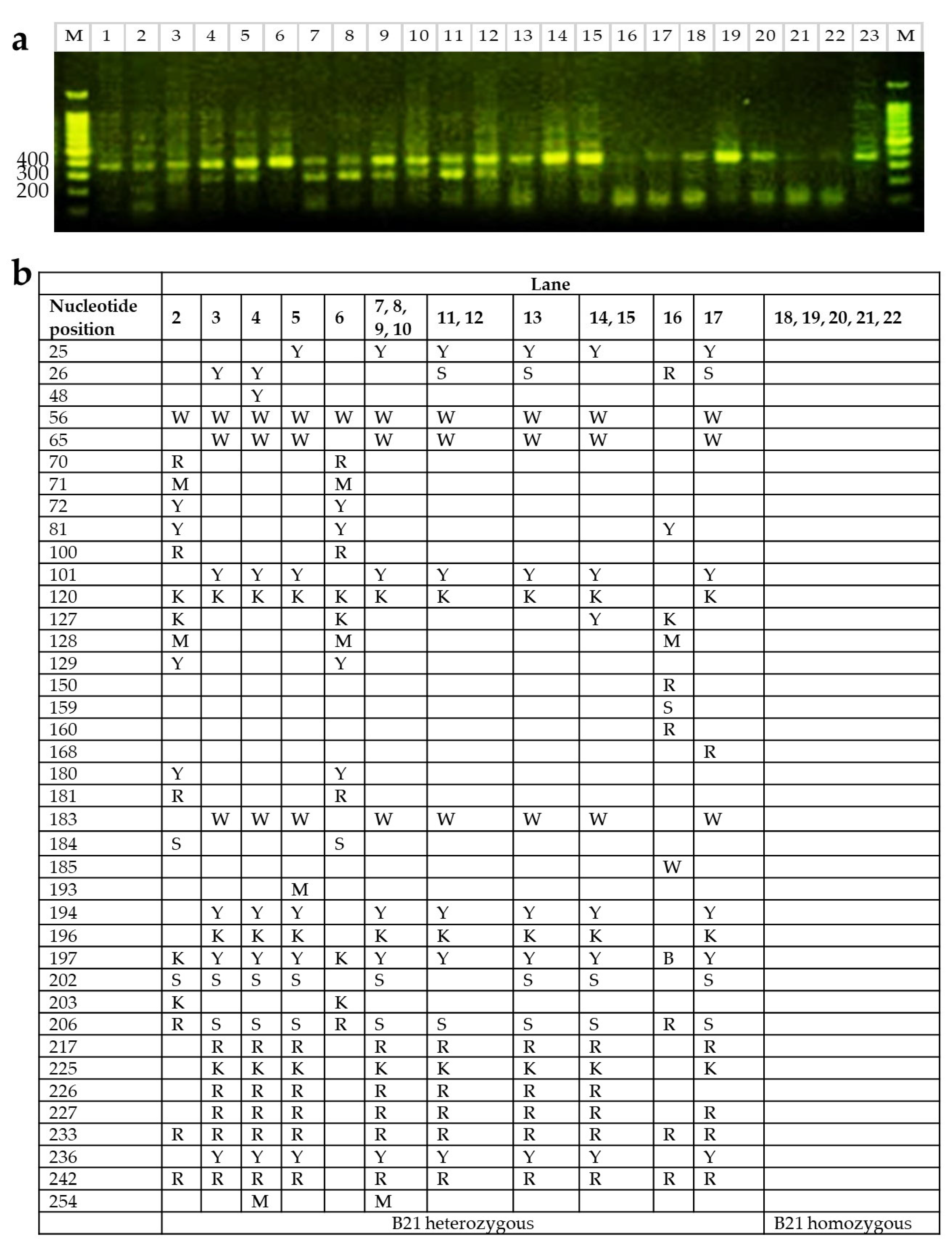
2.3. B21 Haplotype Confirmation Through Sequence-Based Typing (SBT)
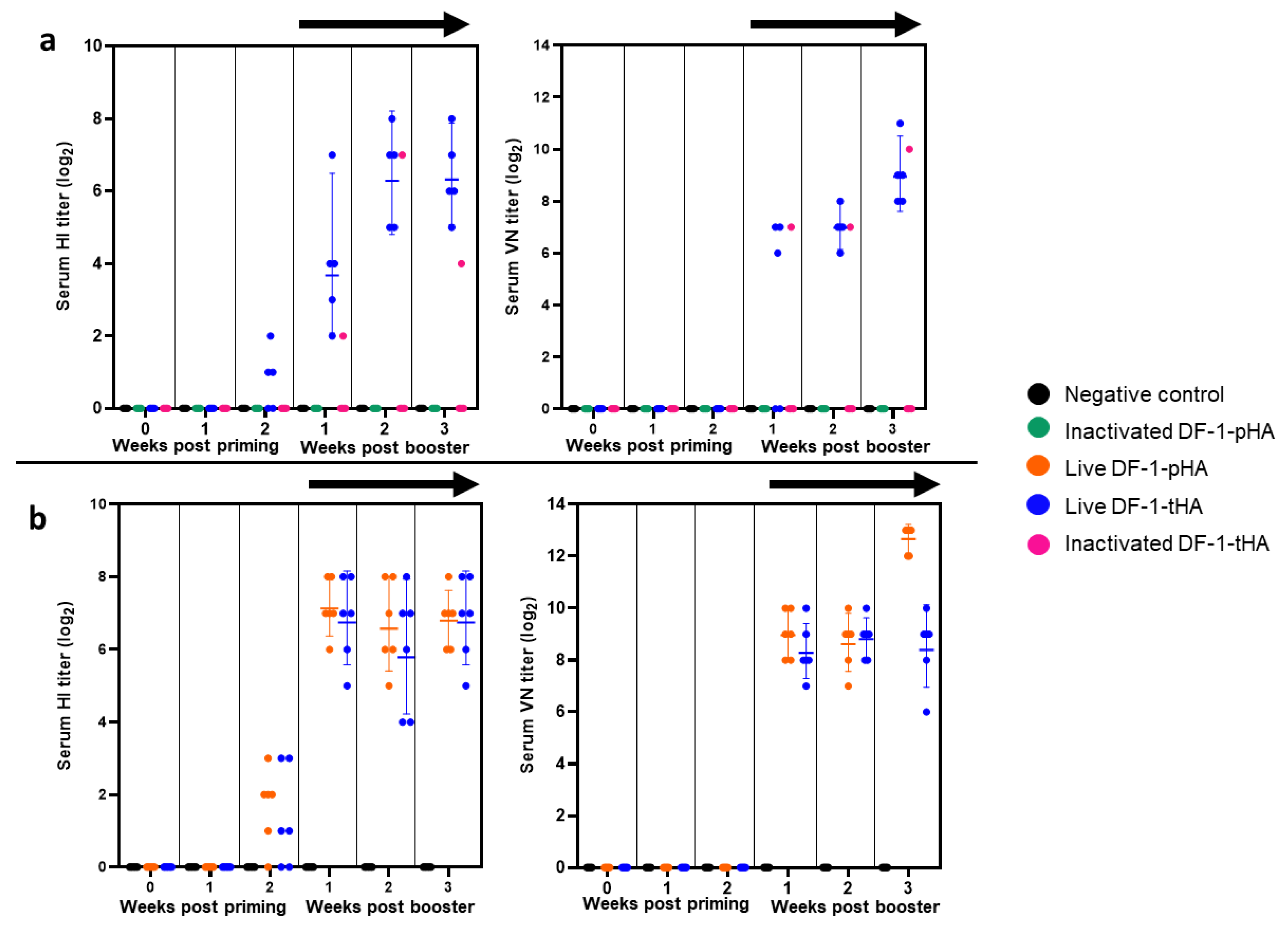
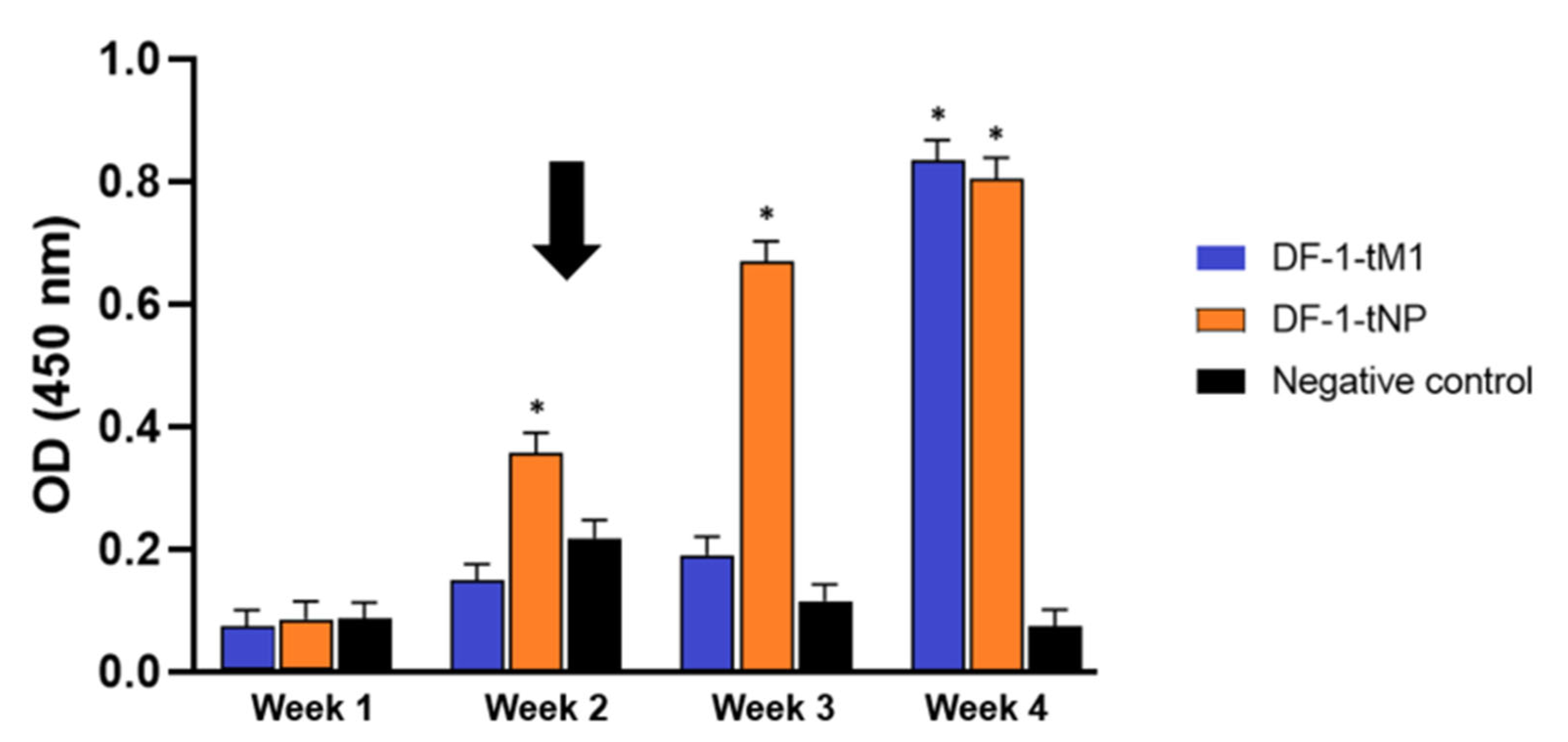
2.4. Live DF-1 Cell Expressing HA Gene Induces Strong Humoral Immunity in VALO SPF Chickens
2.5. Intramuscular Immunization with DF-1 Cells Transiently Expressing M1 or NP Was Sufficient to Induce Virus-Specific Cellular Immune Responses
2.6. CD8+ T Cell Epitopes of HA, M1, and NP Are Present on the Class 1 MHC Molecules of DF-1 Cells
2.7. Multiple CD8+ T Cell Epitope-Binding Motifs in Various Chicken Haplotypes Emphasize the Significance of the NP T Cell Epitope-Binding Region
3. Discussion
4. Materials and Methods
4.1. Cells, Reagents, and Plasmids
4.2. MHC Genotyping
4.3. Vaccine Preparation
4.4. Confocal Microscopy
4.5. SDS-PAGE and Western Blotting
4.6. Vaccination and Sampling
4.7. Hemagglutination Inhibition (HI) and Virus Neutralization (VN) Assays
4.8. ELISA for IFN-γ Detection
4.9. Multi-Omics Mass Spectrometry for CD8+ T Cell Epitope Mapping
4.10. Sequence Variation and MHC-I Binding Predictions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IAV | Influenza A virus |
| SPF | Specific pathogen-free |
| MHC | Major histocompatibility complex |
| HA | Hemagglutinin |
| M1 | Matrix 1 |
| NP | Nucleocapsid protein |
| PBMC | Peripheral blood mononuclear cells |
| HI | Hemagglutination inhibition |
| VN | Virus neutralization |
| DF-1-tHA | Live DF-1 cells transiently expressing HA |
| DF-1-pHA | DF-1 cells permanently expressing HA |
| DF-1-pM1 | DF-1 cells permanently expressing M1 |
| DF-1-pNP | DF-1 cells permanently expressing NP |
References
- Food and Agriculture Organization of the United Nations. FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 22 February 2025).
- Pulit-Penaloza, J.A.; Brock, N.; Belser, J.A.; Sun, X.; Pappas, C.; Kieran, T.J.; Basu Thakur, P.; Zeng, H.; Cui, D.; Frederick, J.; et al. Highly pathogenic avian influenza A(H5N1) virus of clade 2.3.4.4b isolated from a human case in Chile causes fatal disease and transmits between co-housed ferrets. Emerg. Microbes Infect. 2024, 13, 2332667. [Google Scholar] [CrossRef]
- Song, J.H.; Son, S.E.; Kim, H.W.; An, S.H.; Lee, C.Y.; Kwon, H.J.; Choi, K.S. A Model H5N2 Vaccine Strain for Dual Protection Against H5N1 and H9N2 Avian Influenza Viruses. Vaccines 2024, 13, 22. [Google Scholar] [CrossRef]
- Valentin, J.; Ingrao, F.; Rauw, F.; Lambrecht, B. Protection conferred by an H5 DNA vaccine against highly pathogenic avian influenza in chickens: The effect of vaccination schedules. Vaccine 2024, 42, 1487–1497. [Google Scholar] [CrossRef]
- Liniger, M.; Summerfield, A.; Ruggli, N. MDA5 Can Be Exploited as Efficacious Genetic Adjuvant for DNA Vaccination against Lethal H5N1 Influenza Virus Infection in Chickens. PLoS ONE 2012, 7, e49952. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Pepini, T.; Pulichino, A.M.; Carsillo, T.; Carlson, A.L.; Sari-Sarraf, F.; Ramsauer, K.; Debasitis, J.C.; Maruggi, G.; Otten, G.R.; Geall, A.J.; et al. Induction of an IFN-Mediated Antiviral Response by a Self-Amplifying RNA Vaccine: Implications for Vaccine Design. J. Immunol. 2017, 198, 4012–4024. [Google Scholar] [CrossRef] [PubMed]
- Tregaskes, C.A.; Kaufman, J. Chickens as a simple system for scientific discovery: The example of the MHC. Mol. Immunol. 2021, 135, 12–20. [Google Scholar] [CrossRef]
- Wang, W.; Wei, Q.; Hao, Q.; Zhang, Y.; Li, Y.; Bi, Y.; Jin, Z.; Liu, H.; Liu, X.; Yang, Z.; et al. Cellular CARD11 Inhibits the Fusogenic Activity of Newcastle Disease Virus via CBM Signalosome-Mediated Furin Reduction in Chicken Fibroblasts. Front. Microbiol. 2021, 12, 607451. [Google Scholar] [CrossRef]
- Shapiro, J.; Sciaky, N.; Lee, J.; Bosshart, H.; Angeletti, R.H.; Bonifacino, J.S. Localization of endogenous furin in cultured cell lines. J. Histochem. Cytochem. 1997, 45, 3–12. [Google Scholar] [CrossRef]
- Chazara, O.; Juul-Madsen, H.R.; Chang, C.S.; Tixier-Boichard, M.; Bed’hom, B. Correlation in chicken between the marker LEI0258 alleles and Major Histocompatibility Complex sequences. BMC Proc. 2011, 5 (Suppl. 4), S29. [Google Scholar] [CrossRef]
- Bertzbach, L.D.; Tregaskes, C.A.; Martin, R.J.; Deumer, U.S.; Huynh, L.; Kheimar, A.M.; Conradie, A.M.; Trimpert, J.; Kaufman, J.; Kaufer, B.B. The Diverse Major Histocompatibility Complex Haplotypes of a Common Commercial Chicken Line and Their Effect on Marek’s Disease Virus Pathogenesis and Tumorigenesis. Front. Immunol. 2022, 13, 908305. [Google Scholar] [CrossRef]
- Fulton, J.E.; Juul-Madsen, H.R.; Ashwell, C.M.; McCarron, A.M.; Arthur, J.A.; O’Sullivan, N.P.; Taylor, R.L., Jr. Molecular genotype identification of the Gallus gallus major histocompatibility complex. Immunogenetics 2006, 58, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.; Milne, S.; Göbel, T.W.; Walker, B.A.; Jacob, J.P.; Auffray, C.; Zoorob, R.; Beck, S. The chicken B locus is a minimal essential major histocompatibility complex. Nature 1999, 401, 923–925. [Google Scholar] [CrossRef] [PubMed]
- Reemers, S.S.; van Haarlem, D.A.; Sijts, A.J.; Vervelde, L.; Jansen, C.A. Identification of novel avian influenza virus derived CD8+ T-cell epitopes. PLoS ONE 2012, 7, e31953. [Google Scholar] [CrossRef] [PubMed]
- Byrd-Leotis, L.; Galloway, S.E.; Agbogu, E.; Steinhauer, D.A. Influenza hemagglutinin (HA) stem region mutations that stabilize or destabilize the structure of multiple HA subtypes. J. Virol. 2015, 89, 4504–4516. [Google Scholar] [CrossRef]
- Ekiert, D.C.; Bhabha, G.; Elsliger, M.A.; Friesen, R.H.; Jongeneelen, M.; Throsby, M.; Goudsmit, J.; Wilson, I.A. Antibody recognition of a highly conserved influenza virus epitope. Science 2009, 324, 246–251. [Google Scholar] [CrossRef]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Warga, E.; Anderson, J.; Tucker, M.; Harris, E.; Elmer, J. Transcriptomic analysis of the innate immune response to in vitro transfection of plasmid DNA. Mol. Ther. Nucleic Acids 2023, 31, 43–56. [Google Scholar] [CrossRef]
- Meunier, M.; Chemaly, M.; Dory, D. DNA vaccination of poultry: The current status in 2015. Vaccine 2016, 34, 202–211. [Google Scholar] [CrossRef]
- Chen, L.; He, Y.; Liu, H.; Shang, Y.; Guo, G. Potential immune evasion of the severe acute respiratory syndrome coronavirus 2 Omicron variants. Front. Immunol. 2024, 15, 1339660. [Google Scholar] [CrossRef]
- Sha, B.; Luo, M. Structure of a bifunctional membrane-RNA binding protein, influenza virus matrix protein M1. Nat. Struct. Biol. 1997, 4, 239–244. [Google Scholar] [CrossRef]
- Li, Z.; Watanabe, T.; Hatta, M.; Watanabe, S.; Nanbo, A.; Ozawa, M.; Kakugawa, S.; Shimojima, M.; Yamada, S.; Neumann, G.; et al. Mutational Analysis of Conserved Amino Acids in the Influenza A Virus Nucleoprotein. J. Virol. 2009, 83, 4153–4162. [Google Scholar] [CrossRef]
- Hou, Y.; Guo, Y.; Wu, C.; Shen, N.; Jiang, Y.; Wang, J. Prediction and identification of T cell epitopes in the H5N1 influenza virus nucleoprotein in chicken. PLoS ONE 2012, 7, e39344. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Lim, J.M.; Yu, B.; Song, S.; Neeli, P.; Sobhani, N.; K, P.; Bonam, S.R.; Kurapati, R.; Zheng, J.; et al. The next-generation DNA vaccine platforms and delivery systems: Advances, challenges and prospects. Front. Immunol. 2024, 15, 1332939. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.; Gurry, C.; Freitas, L.; Schultz, M.B.; Bach, G.; Diallo, A.; Akite, N.; Ho, J.; Lee, R.T.; Yeo, W.; et al. GISAID’s Role in Pandemic Response. China CDC Wkly. 2021, 3, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.; Krauss, S.; Perez, D.; Webby, R.; Webster, R.G. Eight-plasmid system for rapid generation of influenza virus vaccines. Vaccine 2002, 20, 3165–3170. [Google Scholar] [CrossRef]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.; Perez, D. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef]
- Livant, E.J.; Ewald, S.J. High-resolution typing for chicken BF2 (MHC class I) alleles by automated sequencing. Anim. Genet. 2005, 36, 432–434. [Google Scholar] [CrossRef]
- Dong, C.; Yu, B. Mutation surveyor: An in silico tool for sequencing analysis. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 760, pp. 223–237. [Google Scholar] [CrossRef]
- An, S.H.; Hong, S.M.; Song, J.H.; Son, S.E.; Lee, C.Y.; Choi, K.S.; Kwon, H.J. Engineering an Optimal Y280-Lineage H9N2 Vaccine Strain by Tuning PB2 Activity. Int. J. Mol. Sci. 2023, 24, 8840. [Google Scholar] [CrossRef]
- Song, J.H.; Son, S.E.; Kim, H.W.; Kim, S.J.; An, S.H.; Lee, C.Y.; Kwon, H.J.; Choi, K.S. Rapid and specific on-site H5Nx avian influenza diagnosis via RPA and PAM-independent CRISPR-Cas12a assay combined with anti-NP antibody-based viral RNA purification. Front. Vet. Sci. 2025, 12, 1520349. [Google Scholar] [CrossRef]
- Westgeest, K.B.; Bestebroer, T.M.; Spronken, M.I.; Gao, J.; Couzens, L.; Osterhaus, A.D.; Eichelberger, M.; Fouchier, R.A.; de Graaf, M. Optimization of an enzyme-linked lectin assay suitable for rapid antigenic characterization of the neuraminidase of human influenza A(H3N2) viruses. J. Virol. Methods 2015, 217, 55–63. [Google Scholar] [CrossRef]
- Storkus, W.J.; Zeh, H.J., 3rd; Salter, R.D.; Lotze, M.T. Identification of T-cell epitopes: Rapid isolation of class I-presented peptides from viable cells by mild acid elution. J. Immunother. Emphas. Tumor Immunol. 1993, 14, 94–103. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Predicted Epitope a | Identified Epitope (Percentage) | Variation (%) |
|---|---|---|---|
| HA | 23YHANNSTEQV32 | 395NKVNSIIDKM404 b,c (97.6) | Others (2.4) |
| 402DKMNTQFEAV411 | |||
| 487HKCDNECMESV497 | |||
| 395NKVNSIIDKM404 | |||
| 465DKVRLQLRDNA475 | |||
| M1 | 133NRMGTVTAEGA143 | 133NRMGTVTAEGA143 b (67.9) | NRMGTVTAEVA (14.3) |
| NRMGTVTTEGA (12.2) | |||
| NRMGTVTAEAA (2.1) | |||
| Other (3.5) | |||
| NP | 435GRTSDMRTEII445 | 212GRRTRVAYERM222 b (98.5) | GRRTRIAYERM (1.5) |
| 99RRDGKWMRELI109 | |||
| 120WRQANKGEDA129 | |||
| 212GRRTRVAYERM222 |
| Haplotypes (Binding Motif) | |||
|---|---|---|---|
| B4 X-D/E-X-X-D/E-X-X-(X)-E/L/I | B12 X-X-X-X-V/I-X-X-(X)-V/L/I | B15 X-R-X-X-X-X-X-(X)-Y | B19 X-R-X-X-X-X-X-Y/P/L/F |
| 10YEQMETGGE18 | 21NATEIRASV29 | 212GRRTRVAY219 * | 64ERMVLSAF71 |
| 111YDKEEIRRI119 | 25IRASVGRMV33 | 76NRYLEEHP83 | |
| 251AEIEDLIFL259 | 29VGRMVSGI36 | 212GRRTRVAY219 * | |
| 371METMDSNTL379 | 53EGRLIQNSI61 | 347IRGTRMVP354 | |
| 59NSITIERMV67 | 451ARPEDVSF458 | ||
| 112DKEEIRRI119 | |||
| 179AGAAVKGI186 | |||
| 182AVKGIGTMV190 | |||
| 186IGTMVMEL193 | |||
| 186IGTMVMELI194 | |||
| 190VMELIRMI197 | |||
| 193LIRMIKRGI201 | |||
| 235QRAMVDQV242 | |||
| 249GNAEIEDL256 | |||
| 249GNAEIEDLI257 | |||
| 276LPACVYGL283 | |||
| 308QNSQVFSL315 | |||
| 308QNSQVFSLI316 | |||
| 339EDLRVSSFI347 | |||
| 410PTFSVQRNL418 | |||
| 459QGRGVFEL466 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, S.-M.; Song, J.-H.; Son, S.-E.; Kim, H.-W.; Kim, G.; Hong, S.-M.; Choi, K.-S.; Kwon, H.-J. A Host Cell Vector Model for Analyzing Viral Protective Antigens and Host Immunity. Int. J. Mol. Sci. 2025, 26, 7492. https://doi.org/10.3390/ijms26157492
Ahn S-M, Song J-H, Son S-E, Kim H-W, Kim G, Hong S-M, Choi K-S, Kwon H-J. A Host Cell Vector Model for Analyzing Viral Protective Antigens and Host Immunity. International Journal of Molecular Sciences. 2025; 26(15):7492. https://doi.org/10.3390/ijms26157492
Chicago/Turabian StyleAhn, Sun-Min, Jin-Ha Song, Seung-Eun Son, Ho-Won Kim, Gun Kim, Seung-Min Hong, Kang-Seuk Choi, and Hyuk-Joon Kwon. 2025. "A Host Cell Vector Model for Analyzing Viral Protective Antigens and Host Immunity" International Journal of Molecular Sciences 26, no. 15: 7492. https://doi.org/10.3390/ijms26157492
APA StyleAhn, S.-M., Song, J.-H., Son, S.-E., Kim, H.-W., Kim, G., Hong, S.-M., Choi, K.-S., & Kwon, H.-J. (2025). A Host Cell Vector Model for Analyzing Viral Protective Antigens and Host Immunity. International Journal of Molecular Sciences, 26(15), 7492. https://doi.org/10.3390/ijms26157492