Abstract
Daphnis nerii cypovirus-23 (DnCPV-23) is a new type of cypovirus that has a lethal effect on many species of Sphingidae pests. DnCPV-23 can replicate in Spodoptera frugiperda Sf9 cells, but the replication characteristics of the virus in this cell line are still unclear. To determine the replication characteristics of DnCPV-23 in Sf9 cells, uninfected Sf9 cells and Sf9 cells at 24 and 72 h after DnCPV-23 infection were collected for transcriptome analysis. Compared to uninfected Sf9 cells, a total of 188 and 595 differentially expressed genes (DEGs) were identified in Sf9 cells collected at 24 hpi and 72 h, respectively. KEGG analyses revealed that 139 common DEGs in two treatment groups were related to nutrition and energy metabolism-related processes, cell membrane integrity and function-related pathways, detoxification-related pathways, growth and development-related pathways, and so on. We speculated that these cellular processes might be manipulated by viruses to promote replication. This study provides an important basis for further in-depth research on the mechanism of interaction between viruses and hosts. It provides additional basic information for the future exploitation of DnCPV-23 as a biological insecticide.
1. Introduction
Daphnis nerii cypovirus-23 (DnCPV-23) was initially isolated from naturally diseased larvae of Daphnis nerii (D. nerii), which belongs to the order Lepidoptera and family Sphingidae, and is a worldwide pest [1] that mainly damages the leaves of the ornamental plant oleander and the medicinal plants Rauvolfia vomitoria Afzel and Catharanthus [1,2]. Compared with other cypoviruses, this new type of cypovirus has different electrophoretic migration patterns and conserved terminal sequences [1,3,4]. In addition to D. nerii, DnCPV-23 can infect and induce death in various harmful insects of the Sphingidae family. However, as a potential biopesticide, the molecular mechanism of the interaction between DnCPV-23 and its hosts remains unclear. The study of the interaction mechanism between DnCPV-23 and hosts is essential for understanding virus virulence and virus genome function, screening host molecules that promote virus replication, and screening complex drugs that improve virus insecticidal efficacy.
We previously performed a transcriptome analysis of DnCPV-23-infected and uninfected D. nerii midguts [1]. DnCPV-23 can infect wild insects such as D. nerii, but the lifecycle of these insects is strongly affected by season and cannot be artificially reproduced at present. Therefore, it is essential to study the effects of DnCPV-23 on cell lines. In addition, DnCPV-23 infection of the D. nerii midgut occurs in the form of occlusion-derived viruses (ODVs) in vivo, and the midgut cell composition is complex. However, the Sf9 cell line can be infected in vitro with free virus particles of DnCPV-23. Based on different infection forms and cell types, we speculate that the transcriptome differs between cultured cell lines and D. nerii midguts. Our previous results revealed that DnCPV-23 can be effectively replicated and passaged in Sf9 cells [5]. However, the molecular mechanism underlying DnCPV-23 infection in Sf9 cells has not been fully elucidated. Moreover, there are few reports on the replication of insect RNA viruses in heterologous cell lines. Perina nuda virus (PnV) can establish persistent infection in a heterologous Lymantria xylina cell line, NTU-LY. Various methods were used to investigate virus replication in NTU-LY cells [6]. Moreover, the honeybee virus deformed wing virus (DWV) can infect the heterologous Lepidopteran haemocytic cell line P1 cells [7]. This study reports the replication of another RNA virus, DnCPV-23, in the heterologous cell line Sf9. Our work provides an important foundation for in-depth research on the replication of RNA viruses in heterologous cells and for establishing a sustained infection cell model for insect RNA viruses. To investigate how Sf9 cells react to DnCPV-23 infection, we used a high-throughput sequencing approach to assess the impact of DnCPV-23 infection on global gene expression in Sf9 cells and analysed the host factors that may affect virus replication. In this study, uninfected Sf9 cells and DnCPV-23-infected Sf9 cells were collected at 24 h and 72 h after infection for transcriptome analysis to investigate changes in the gene expression profiles of Sf9 cells after virus infection and identify the host signalling pathways that may affect virus replication.
We speculate that upregulating some processes (including nutrition and energy metabolism, cell membrane integrity and functions, detoxification, and growth and development) after viral infection is beneficial for DnCPV-23 replication. For example, juvenile hormone acid O-methyltransferase-like (JHAMT) can promote virus proliferation by prolonging insect pupation time; virus replication can be promoted by inhibiting host apoptosis through upregulating the expression of cytochrome P450 9e2 (CYP9E2); and Sequestosome 1 (p62/SQSTM1) affects the lifecycle of viruses by regulating autophagy. Through this study, we have identified some host genes that may affect the replication of DnCPV-23. In future work, we will further verify the interaction between these genes and DnCPV-23, which provides an important theoretical basis for understanding the interaction between viruses and hosts, virus replication in heterologous cell lines, and viral biological insecticide development.
2. Results
2.1. Viral Infection of Cells
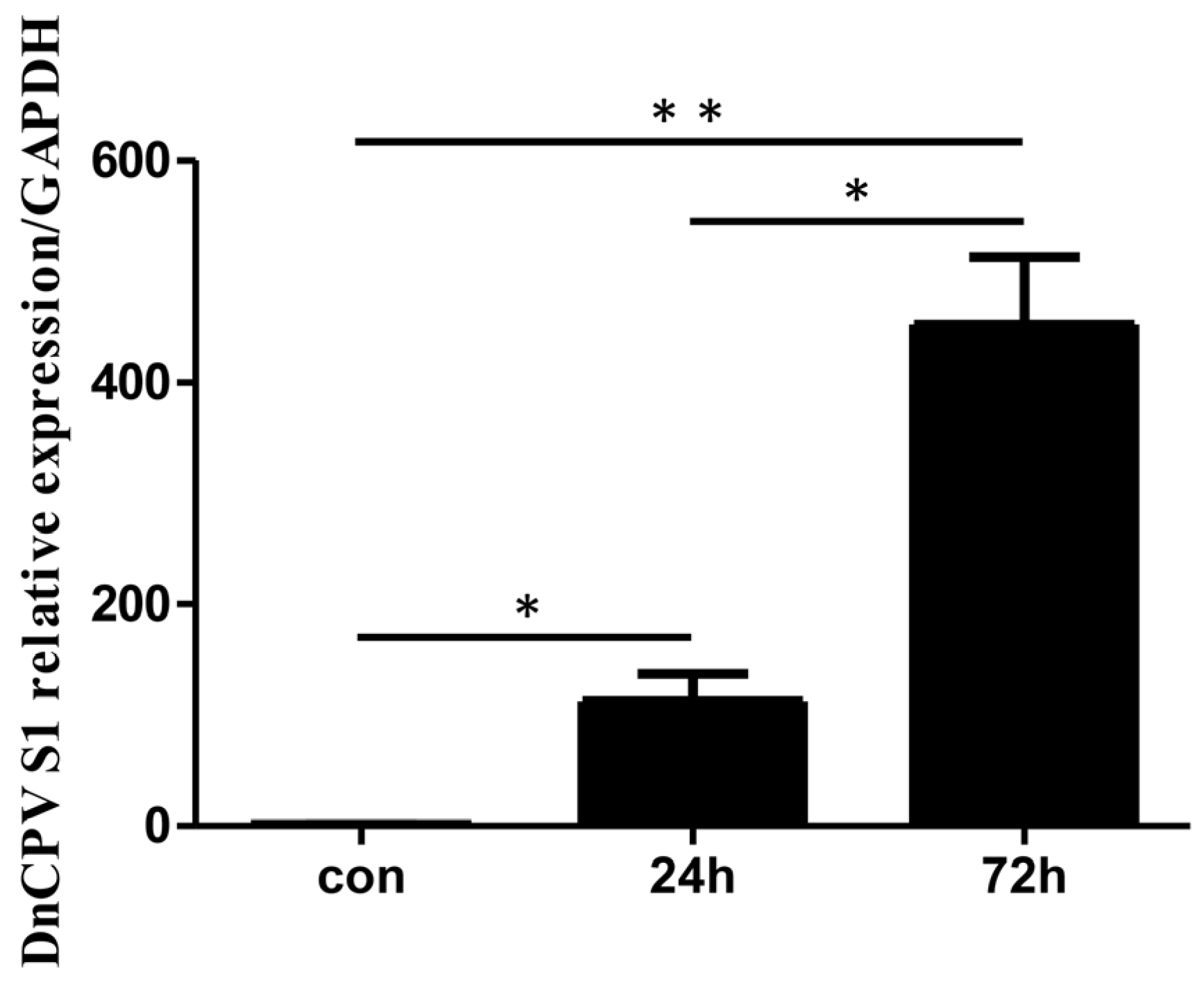
The virus-to-cell ratio is 0.2 polyhedra per cell. Prior to transcriptome analysis, qRT–PCR was used to assess the mRNA levels of the DnCPV-23 S1 gene in infected cells (samples collected at 24 h and 72 h post-infection (hpi)) and in uninfected cells. The results revealed higher relative expression of viral gene mRNA in infected cells than in uninfected cells (Figure 1).
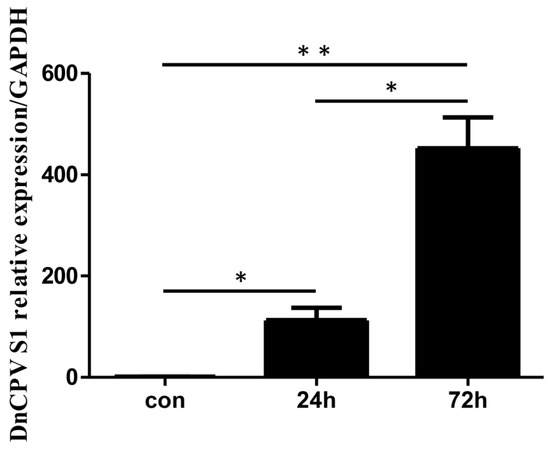
Figure 1.
Analysis of viral RNA in infected cells. Sf9 cells were collected at 24 and 72 h post-infection (hpi). The mRNA levels of DnCPV-23 S1 in control cells and infected cells were assessed via qRT–PCR. Statistically significant differences are indicated as * p < 0.05, ** p < 0.01. Data with p values < 0.05 were considered statistically significant.
2.2. Overview of Transcriptome Sequencing
All of the samples were sequenced independently. Transcriptome sequencing analysis of 15 samples resulted in a total of 81.39 Gb of clean data. The clean data of each sample ranged from 6.41 to 7.04 G. The Q30 of each sample ranged from 94.12% to 94.42%, and the average GC content was 45.96%. The genome alignment of each sample was obtained by mapping reads to the reference genome, with a mapping rate of 81.42% to 82.46%. A total of 595 DEGs were identified, with 506 being upregulated and 89 being downregulated between the CPV_72h samples and the control samples.
2.3. Effects of Viral Infection on the Transcriptome Expression of Sf9 Cells
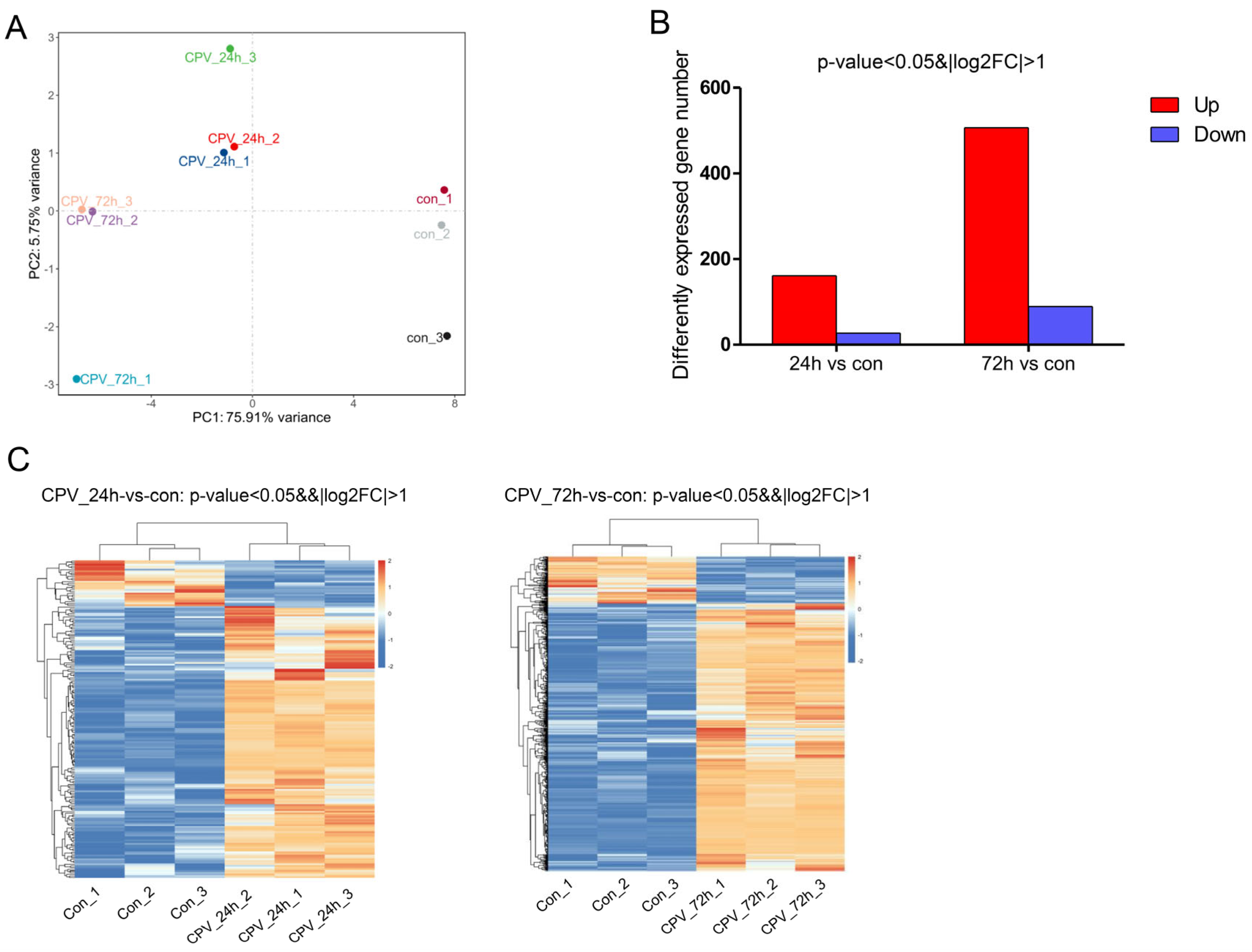
As shown in Figure 2, PCA1 accounted for 75.91%, and PCA2 accounted for 5.75%. Therefore, the percentage of the total of the two was 81.66%, thus accounting for a high proportion and representing the overall population to a large extent. The principal component analysis revealed a clear separation of the samples among the three groups (Figure 2A), indicating that the samples had good repeatability. Compared with uninfected cells, in infected cells, the number of upregulated genes at 24 hpi and 72 hpi was 161 and 506, respectively, and the number of downregulated genes at 24 hpi and 72 hpi was 27 and 89, respectively (Figure 2B). In addition, a heatmap of the gene expression data is presented in Figure 2C. The results suggested that these DEGs could be used to distinguish the samples. The results revealed that viral infection could influence Sf9 gene expression at different time points.
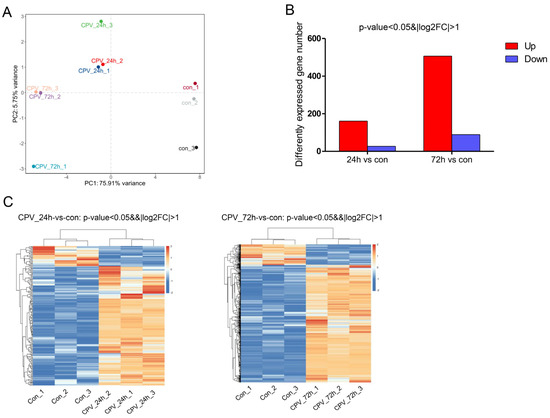
Figure 2.
Influence of DnCPV-23 infection on the Sf9 cell transcriptome. (A). Plot of the 1st and 2nd principal components of sample variation, as determined using principal component analysis. (B). Compared with uninfected cells, in infected cells, 161 and 506 genes were upregulated (red bars), and 27 and 89 genes were downregulated (blue bars) at 24 hpi and 72 hpi, respectively. (C). Heatmap of 188 and 595 differentially expressed genes (DEGs) between uninfected cells and cells collected at 24 hpi and 72 hpi.
2.4. Analysis of DEGs
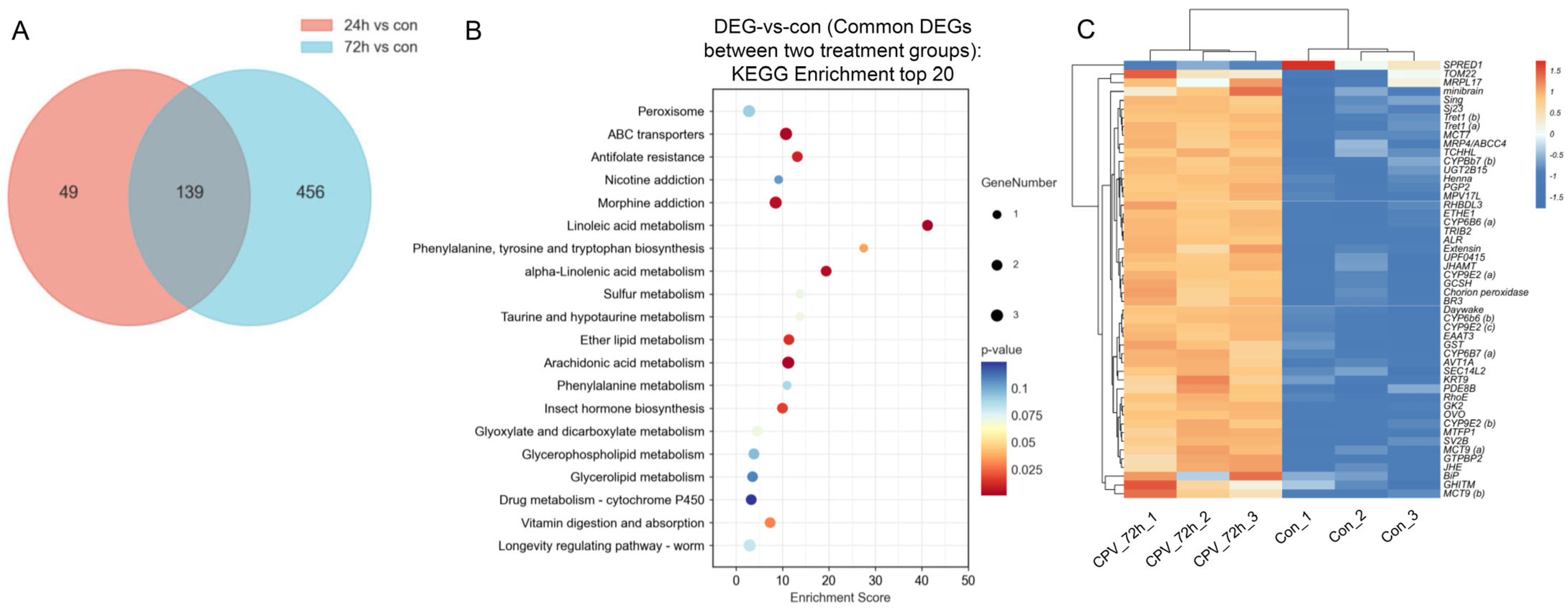
We analysed 139 DEGs shared by the two treatment groups for further analysis (Figure 3A). KEGG functional enrichment analysis was performed using the common DEGs between the two treatment groups to identify the relevant biological pathways associated with these DEGs. Based on the enrichment score obtained from the KEGG analysis, we identified the 20 most enriched pathways. These pathways play important roles in nutrition and energy metabolism-related processes (for example, “linoleic acid metabolism” and “vitamin digestion and absorption”), cell membrane integrity and function-related pathways (for example, “glycerophospholipid metabolism” and “ether lipid metabolism”), detoxification-related pathways (for example, “ABC transporters”), growth and development-related pathways (for example, “insect hormone biosynthesis”), and insect immune responses (for example, “arachidonic acid metabolism”) (Figure 3B). We selected 50 DEGs with the highest expression levels from these 139 DEGs for heatmap analysis. A heatmap of the gene expression data is presented in Figure 3C. The results suggested that the identified DEGs could distinguish the samples and that DnCPV-23 infection influenced gene expression in Sf9 cells.
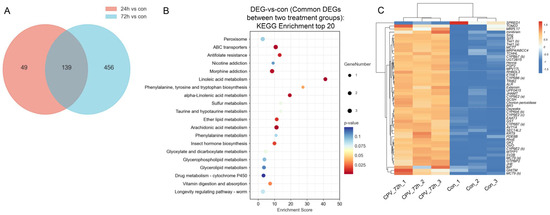
Figure 3.
Signalling pathways and genes involved in the interaction between DnCPV-23 and Sf9 cells. (A) Venn diagram of DEGs obtained from the two treatment groups; there were 139 DEGs that exhibited differential expression between samples infected with DnCPV-23 for 24 h and 72 h compared to the control. (B) KEGG classifications of 139 common DEGs between the two treatment groups (top 20). The larger the bubble is, the greater the number of DEGs. The bubble colour changes (purple–blue–green–red) indicate that the smaller the enrichment p value is, the greater the significance. (C) Heatmap analysis for 50 DEGs with the highest expression level selected from the overlapping 139 DEGs.
Figure 3C shows the top 50 genes with the highest expression levels among the overlapping 139 DEGs, and the relevant information of these genes is presented in Table 1.

Table 1.
Information of 50 DEGs with the highest expression levels selected from the 139 common DEGs in two treatment groups (P62/SQSTM1 only differentially expressed at 72 hpi).
2.5. qRT–PCR Validation of DEGs
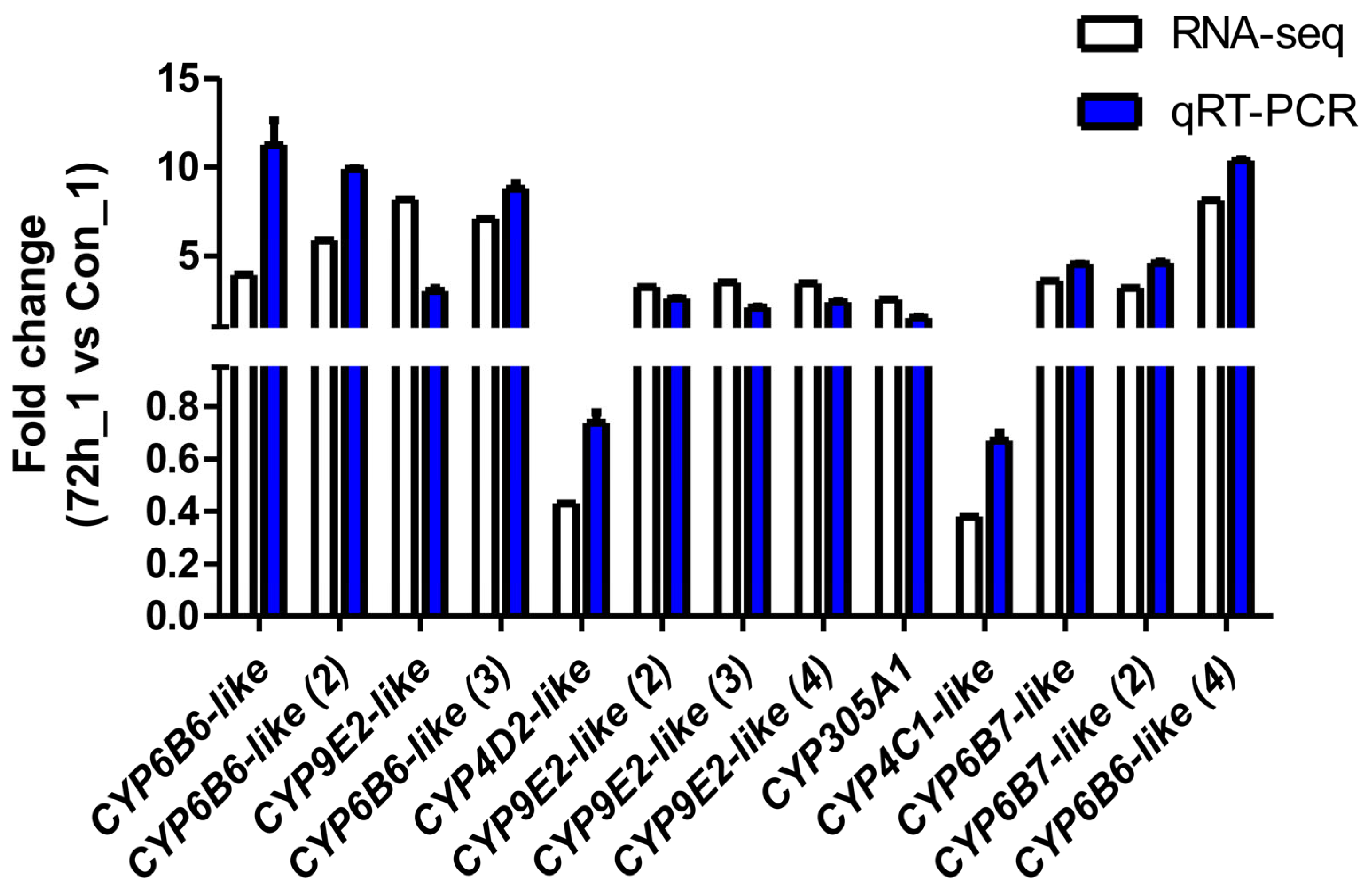
To verify the reliability of the transcriptome data and the DEG results obtained by RNA-seq, thirteen DEGs were selected for qPCR analysis. As shown in Figure 4, the fold-change values for the infected cells (CPV_72_1) vs. the uninfected cells (Con_1) obtained via the qPCR analysis were consistent with the values obtained via RNA-seq for all of the selected genes.
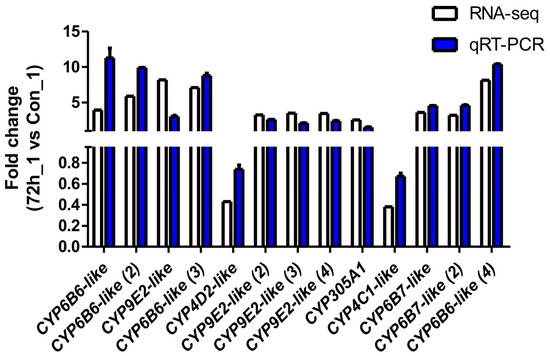
Figure 4.
Validation of RNA-seq profiles by real-time qPCR. To validate the RNA-seq data, the relative mRNA levels of 13 selected DEGs in infected cells (CPV_72_1) were examined by qPCR. The mRNA levels determined via qPCR are presented as the fold change compared with the mRNA levels in the uninfected cells (Con_1) after normalisation against GAPDH. The relative expression levels from the RNA-seq analysis were calculated as RPKM values. The error bars represent means ± SEMs.
3. Discussion
To analyse the important signalling pathways and genes that may be involved in the interaction between DnCPV-23 and Sf9 cells, we selected 139 common genes between 24 h DEG and 72 h DEG for analysis. The KEGG analysis revealed that many processes changed in Sf9 cells after virus infection, including nutrition and energy metabolism-related processes (such as “linoleic acid metabolism” and “vitamin digestion and absorption”), cell membrane integrity and function-related pathways (such as “glycerophospholipid metabolism” and “ether lipid metabolism”), detoxification-related pathways (such as “ABC transporters”), and growth and development-related pathways (such as “insect hormone biosynthesis”). The KEGG analysis indicated that these processes or pathways might have important roles in the interactions between DnCPV-23 and Sf9 cells. Furthermore, these processes or pathways also changed in the midgut of Daphnis nerii after DnCPV-23 infection [1].
DnCPV-23 infection can affect the expression of critical genes involved in energy metabolism, such as glycerol kinase-like (GK; Gene ID: LOC118276660) and trehalose transporter 1 (Tret1; Gene ID: LOC118271571, LOC118279019). Glycerol kinase (GK) is an enzyme that catalyses the formation of glycerol 3-phosphate from ATP and glycerol [8]. GK2 is a glycerol biosynthesis gene that plays a crucial role in survival during the cold period [9]. HCV/HBV infection reduces the expression of miR-451a, which inhibits GK expression, and miR-451a attenuates hepatitis C virus replication by targeting glycerol kinase. Furthermore, the supplementation of miR-451a could impede lipid deposition, reduce steatohepatitis, and inhibit HCV replication in the liver [10]. After virus infection, two Tret1 transcripts were significantly upregulated. Tret1 is a specific and high-capacity facilitated transporter of trehalose, which is the major sugar found in insect haemolymph fluid, providing energy and promoting growth, metamorphosis, stress recovery, chitin synthesis, and virus replication in insects [11,12,13]. In BmN cells, BmNPV infection promotes the expression of trehalose hydrolysis and transport-related genes to facilitate BmNPV proliferation through the trehalose-PI3K-Akt pathway in the midgut [14]. Moreover, trehalose could facilitate virus replication and shedding of dengue virus (DENV) in Aedes aegypti cells. It is possible that trehalose increases DENV2 infection in Aag2 cells by promoting autophagy to prolong cell survival and enhance virus maturation, and enhancing virus cell entry through the modification of cell membranes [12].
In addition to affecting energy metabolism, DnCPV-23 infection also affected insect metamorphosis, day–night rhythm, and oocyte development-related genes. Juvenile hormone esterase-like (JHE, Gene ID: LOC118263046) was upregulated, induced by DnCPV-23 infection. JHE is the primary juvenile hormone (JH)-specific degradation enzyme that plays a crucial role in regulating JH levels. Depletion of JHE resulted in the extension of Bombyx mori larval stages [15] or larval mortality in Telchin licus (Lepidoptera) [16]. JHE activity is downregulated in larval Adoxophyes honmai following dhoNPV and AdorNPV infection, and small interfering RNAs were proposed to play a role in the downregulation of JHE gene expression in baculovirus-infected caterpillars [17]. The Mamestra brassicae Multiple Nucleopolyhedroviruses (MbMNPV) infection of H. armigera larvae inhibits the expression of JHE following the upregulation of JH titre. The upregulation of JH titre prevents the pupation of H. armigera and promotes MbMNPV replication through the JH-Met-Kr-h1 signalling pathway [18]. DnCPV-23 infection promoted the expression of juvenile hormone acid O-methyltransferase-like (JHAMT; Gene ID: LOC118265567). Juvenile hormone (JH) titres in insects are regulated by synthesis with JHAMT and catabolism with JHE [19]. JH could regulate metamorphosis, reproduction, diapause, and polyphenisms [20,21]. The knockdown of JHAMT reduced JH titre, leading to larval death in Aedes aegypti [20] and changing the phenotype of S. frugiperda [22]. During the early stages of infection, Zika virus (ZIKV) regulates the expression of some ribosomal protein genes (RpL23 and RpL27) through the JH-Met-Tai signalling pathway. Moreover, the upregulated expression of RpL23 and RpL27 could facilitate the translation of viral proteins and promotes ZIKV infection in Aedes aegypti [23]. Further research is needed to investigate the detailed effects of these host genes on DnCPV-23 infection.
After viral infection, the expressions of many detoxification-related genes were upregulated, such as cytochrome P450 9e2 (CYP9E2), cytochrome P450 6B7 (CYP6B7), cytochrome P450 6B6 (CYP6B6), glutathione S-transferase (GST), UDP-glucuronosyltransferase (UGT), and multidrug resistance-associated protein 4-like (MRP4/ABCC4). CYP9E2 (Gene ID: LOC118264059, LOC118264054, LOC118262727), CYP6B7 (Gene ID: LOC118279736, LOC118279835) and CYP6B6 (Gene ID: LOC118263048, LOC118262642) are involved in xenobiotic detoxification in insects. In Helicoverpa armigera, CYP6B6 plays crucial roles in the transformation of esfenvalerate and capsaicinoids [24,25]. In H. armigera, CYP6B7 is known as an important detoxification gene in response to fenvalerate [26,27]. In honeybees, thiacloprid can be specifically metabolised by CYP9E2 [28]. In addition, the CYP9E2 gene of Bombyx mori can be regulated by the microRNA bmo-miR-31-5p to inhibit apoptosis and promote BmNPV proliferation [29]. In the future, we will further analyse the role of these CYP450 genes in virus replication. GST (Gene ID: LOC118261931) is a group of enzymes associated with detoxification and plays an important role in detoxifying endogenous and exogenous compounds [30]. The expression of GST could be regulated by viral infection [31,32]. GST may modify lipids to inhibit viral infection, or regulate inflammatory-like mediators to exert antiviral effects [33]. However, GST could promote BmNPV proliferation through regulating glutathione (GSH) levels [34]. DnCPV-23 infection also induced the expression of another detoxification enzyme, UGT2B15-like (Gene ID: LOC118278849). UDP-glucuronosyltransferases (UGTs) could transform various exogenous and endogenous compounds, which play a critical role in detoxification and homeostasis in insects [35]. Moreover, UGTs could regulate viral infection. It is speculated that UGT33D1 reduces oxidative stress to promote BmNPV infection [36]. DnCPV-23 replication could promote the expression of MRP4/ABCC4 (Gene ID: LOC118262225). MRP4 (multidrug resistance-associated protein 4) is a member of the ATP-binding cassette (ABC) transporter C subfamily, which are transporters of various substrates including endogenous compounds, xenobiotic compounds, and nutrients [37,38]. HIV-1 infection induced a significant increase in MRP4 expression in human macrophages. The increase in MRP4 expression may favour the efflux of antiretroviral drugs in macrophages to promote virus replication [39].
DnCPV-23 could upregulate the expression of Sequestosome 1 (p62/SQSTM1, Gene ID: LOC118273096, LOC118273082). BmCPVs can induce mitophagy, which is a form of autophagy, through the interaction of VP4 with host Tom40 to promote self-replication [40]. Additionally, BmCPV replication could be attenuated by vsp21-induced autophagy [41]. Thus, the impact of autophagy on virus replication may be opposite in different situations. Many studies have shown that autophagy is a double-edged sword for virus replication [42,43,44]. We used autophagy inhibitors to analyse the effects of blocking different stages of the autophagy process on virus replication. Compared with the control (DMSO), treatment with Baf A1 significantly restricted virus replication, whereas treatment with 3-MA or CQ did not affect virus replication (Supplementary Figure S1). The results suggested that maintaining the acidic pH and protein degradation ability of lysosomes might be crucial for virus replication [45,46]. In addition, we found that DnCPV-23 infection promoted the expression of Sequestosome 1 (SQSTM1/p62), an adaptor protein of selective autophagy, to promote the degradation of aggregate-prone proteins. SQSTM1 is a well-known autophagy cargo receptor (ACR) that plays a dual role during pathogen infection; it can promote the degradation of viral proteins, aiding in the elimination of the virus, but can also be manipulated by viruses to promote mitochondrial degradation, thereby suppressing immune responses [47]. A previous study reported that Baf A1 treatment can decrease the protein level of SQSTM1 [48]. Above all, we speculate that the DnCPV-23 infection of host cells promotes autophagy and self-replication by upregulating SQSTM1 expression, which can be blocked by Baf A1. However, in this study, we did not explore it thoroughly. To confirm this conclusion, further research is needed to analyse the impact of lysosomal pH on DnCPV-23 replication and elucidate whether Baf A1 affects the replication of DnCPV-23 by downregulating the expression of SQSTM1. In the future, we will (1) use CRISPR/Cas9 to knock out SQSTM1 in Sf9 to determine whether Baf A1 inhibits DnCPV replication through SQSTM1; (2) use immunofluorescence, electron microscopy scanning, Western Blotting, and RT qPCR to determine which stage of the virus Baf A1 specifically affects: (3) use knockdown, overexpression, or inhibitor inhibition assays to analyse the host factors involved in Baf A1 affecting virus replication [49,50,51].
In Sf9 cells, we revealed substantial differences in the transcription of genes related to energy metabolism, growth, development, and detoxification processes induced by DnCPV-23 replication. We speculate that these cellular processes might be regulated by DnCPV-23 to promote its replication. In addition, we found that autophagy-related genes also undergo changes, and we need further research to determine the role of autophagy in the DnCPV-23 replication. In these pathways, we analysed some specific host genes that may be involved in DnCPV-23 replication. This study provides important preliminary work for future validation experiments. This study analysed the differentially expressed genes (DEGs) of Sf9 cells after DnCPV-23 infection for the first time, and identified host genes that may be involved in DnCPV-23 replication. Our work enriched the research on the interaction between cypovirus and hosts. However, there has been no definitive and credible validation of the role of relevant host genes in virus replication, making it difficult for readers to determine which host genes are truly involved in the lifecycle of DnCPV-23. In summary, the results obtained in this study provide an important foundation for further in-depth research on the mechanism of interaction between DnCPV-23 and its hosts.
4. Materials and Methods
4.1. Cell Line, Virus Stock, Inhibitors, and Antibodies
Sf9 cells, an ovary-derived cell line of Spodoptera frugiperda, were cultured at 28 °C in Grace’s insect medium (Gibco, Waltham, MA, USA) supplemented with 10% foetal bovine serum (Gibco, USA). DnCPV-23 was initially isolated from the larvae of D. nerii and propagated in D. nerii larvae [1]. The suspension of DnCPV-23 polyhedra used for infecting Sf9 cells was stored at 4 °C in the dark. 3-Methyladenine (3-MA) (Selleck, Houston, TX, USA) was used at 5 mM, chloroquine (CQ) (MedChemExpress, Monmouth Junction, NJ, USA) was used at 40 µM, and bafilomycin A1 (Baf A1) (MedChemExpress) was used at 20 nM. For immunoblotting, rabbit polyclonal antibodies against the DnCPV-23 protein encoded by the viral S1 gene (ABclonal Biotechnology, Woburn, MA, USA) were used at a 1:500 dilution, and mouse monoclonal antibodies against tubulin (Abbkine, Atlanta, GA, USA) were used at a 1:10,000 dilution.
4.2. Virus Inoculation
To promote virus infection in cells, purified DnCPV-23 polyhedra at a concentration of approximately 2 × 107/mL were treated with 0.2 M Na2CO3–NaHCO3 (pH 10.8), after which the pH was adjusted to 7.4 with 1 M Tris-HCl (pH 6.8) buffer. The free virus particles were stored at −80 °C. Sf9 cells were seeded in 6-well plates, and when they reached a confluency of approximately 75%, they were infected with free DnCPV-23 virus particles (50 μL of the viral suspension containing free DnCPV-23 particles without polyhedral particles). After incubation for 12 h, the cell culture medium was replaced with Grace medium containing 10% (v/v) FBS, and the cells were harvested 24 h and 72 h after infection. Uninfected cells were used as controls.
4.3. RNA Extraction, Library Preparation, and RNA-Seq
Control Sf9 cells and Sf9 cells infected with DnCPV-23 were collected at 24 h and 72 h postinfection. All RNA sequencing (RNA-seq) procedures were conducted by the Oebiotech Company (Shanghai, China). Total RNA was extracted from Sf9 cells using TRIzol reagent (Invitrogen, Waltham, MA, USA) in accordance with the manufacturer’s protocols. The RNA integrity and concentrations were assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Nine RNA samples (including three uninfected samples and nine infected samples) with confirmed RNA integrity were used to construct the libraries. The cDNA libraries were prepared using a TruSeq RNA Sample Preparation Kit (Illumina, San Diego, CA, USA) in accordance with the manufacturer’s protocols. Thereafter, the obtained cDNA libraries were sequenced on the Illumina HiSeq2500 platform, which generated paired-end raw reads of 150 bp.
4.4. RNA-Seq Data Analysis
To obtain clean reads, raw reads in fastq format were first processed using fastp 1, and the low-quality reads were removed. The clean reads were mapped to the reference genome ZJU_Sfru_1.0 GCF_011064685.1 using HISAT2 (version 2.2.4) [52]. Gene expression levels were normalised as fragments per kilobase of transcript per million mapped reads (FPKM) values. Principal component analysis (PCA) was performed using R (v 3.2.0) to evaluate the biological duplication of samples.
Differentially expressed genes (DEGs) were analysed using DESeq2. A Q value < 0.05 and a fold change > 2 or fold change < 0.5 were set as the thresholds for DEGs. Hierarchical cluster analysis of DEGs was performed using R (v 3.2.0) to investigate the expression patterns of genes in different groups and samples. Furthermore, all DEGs were subjected to GO and KEGG analyses to screen for significantly enriched terms via R (v 3.2.0).
4.5. Quantitative Real-Time PCR
Quantitative real-time PCR (qRT–PCR) was used to analyse the expression level of the DnCPV-23 S1 gene in transcriptome samples and verify the DEGs identified by RNA-seq. Total RNA was isolated from samples for transcriptomic analysis using TRIzol reagent (Life Technologies, Carlsbad, CA, USA) and then treated with DNase I (Fermentas, Glen Burnie, MD, USA). Total RNA (500 ng per sample) was reverse-transcribed into complementary DNA (cDNA) using a PrimeScript RT Reagent Kit (Takara, San Jose, CA, USA). Then, qRT–PCR was performed using Talent qPCR PreMix SYBR Green (Tiangen, Beijing, China) on a QuantStudio™ 7 Flex Real-Time PCR System (Applied Biosystems™, Waltham, MA, USA). One cycle was added for melting curve analysis for all of the reactions to verify the product specificity. Relative expression levels of genes were calculated using the 2−ΔΔCT method, and GAPDH was used as a reference for normalisation [53]. All of the primers for the target genes are listed in Table 2.
Table 2.
Primers used for the qRT–PCR analysis of viral RNA in transcriptome samples and for the validation of RNA-seq data.
4.6. Cell Treatments
Sf9 cells were seeded (1 × 105 cells per well) in a 24-well plate and allowed to attach overnight. The next day, the cells were pretreated with the autophagy inhibitors 3-MA, CQ, and Baf A1 for 2 h prior to DnCPV-23 infection. The final concentrations of 3-MA, CQ, and Baf A1 were 5 mM, 40 µM, and 20 nM, respectively. Two hours later, 10 μL of the viral suspension containing free DnCPV-23 particles without polyhedral particles was added to the cells for 6 h. After the incubation period, the medium was aspirated, and fresh cell culture media containing autophagy inhibitors were added to the cells, which were subsequently cultured for 24 h and 48 h.
4.7. Western Blotting
Cells were collected by centrifugation at 300× g for 5 min at 4 °C, and total protein was extracted using RIPA lysis buffer (Beyotime, Shanghai, China). The cells were placed on ice for 20 min, and the supernatant was obtained by centrifugation at 6000× g for 6 min at 4 °C. Samples were mixed with 5× SDS loading buffer and boiled at 100 °C for 7 min. For Western blot analysis, after SDS-PAGE, proteins were transferred to polyvinylidene fluoride (PVDF) membranes, which were incubated overnight at 4 °C with the appropriate primary antibodies. The membranes were subsequently rinsed three times with TBST (each for 10 min) and incubated with horseradish peroxidase-conjugated goat anti-mouse IgG or goat anti-rabbit IgG as the secondary antibody for 2 h at room temperature. Protein bands were visualised using an enhanced chemiluminescence (ECL) Western blot detection kit and analysed using the ChemiScope 3000 mini system (Clinx, Shanghai, China).
4.8. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 6.0. The values are presented as the mean plus or minus the standard error of the mean (SEM). The unpaired two-tailed t-test was used to compare one factor among different groups. Statistically significant differences are indicated as * p < 0.05, ** p < 0.01, or *** p < 0.001. Data with p values < 0.05 were considered statistically significant.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms26157487/s1.
Author Contributions
W.K.: Writing—review and editing, Writing—original draft, Visualisation, Validation, Project administration, Methodology, Investigation, Data curation, Conceptualisation, Funding. C.Y. (Chenghua Yan): Writing—review and editing, Supervision, Investigation, Formal analysis, Conceptualisation. W.K. and L.J.: Writing—review and editing, Supervision, Resources, Funding. Others (J.Y., J.W., J.C., X.L., C.Y. (Chunhua Yang), Z.Z., L.G., J.L.; T.D., F.Y. and G.M.): Methodology and Suggestions. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of Jiangxi Province (Project no. 20224BAB205004), the Pilot Demonstration Project for Increasing Funding of the Provincial Science and Technology Program of Jiangxi Academy of Sciences (Project no. 2022YSBG21006), the National Natural Science Foundation of China (Project no. 32260705), the Natural Science Foundation of Jiangxi (Project no. 20224BAB216001), the Natural Science Foundation of Jiangxi Province (Project no. 20232BAB215007), the Open Research Fund Program of the State Key Laboratory of Virology of China (Project no. 2022IOV001), the Pilot Demonstration Project for Increasing Funding of the Provincial Science and Technology Program of Jiangxi Academy of Sciences (Project no. 2023YSBG22005), the Natural Science Foundation of Jiangxi Province (Project no. 20224BAB206029), the Overall Rationing System Pilot Project of Jiangxi Academy of Sciences (Project no. Grant number 2022YSBG22013), the Jiangxi Province Science and Technology Matching Support Project (Project no. 20250BFA99002) and Basic Research Program of Jiangxi Academy of Sciences (Grant number 2022YJC2007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. The original data of the transcriptome will be released on 10 July 2025 or upon publication. BioProject accession: PRJNA1279750.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kuang, W.D.; Yan, C.H.; Zhan, Z.G.; Guan, L.M.; Wang, J.C.; Chen, J.H.; Li, J.H.; Ma, G.Q.; Zhou, X.; Jin, L. Transcriptional responses of Daphnis nerii larval midgut to oral infection by Daphnis nerii cypovirus-23. Virol. J. 2021, 18, 250. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, C.; Gao, J.; Abbas, M.N.; Kausar, S.; Qian, C.; Wang, L.; Wei, G.; Zhu, B.J.; Liu, C.L. Comparative mitochondrial genome analysis of Daphnis nerii and other lepidopteran insects reveals conserved mitochondrial genome organization and phylogenetic relationships. PLoS ONE 2017, 12, e0178773. [Google Scholar] [CrossRef]
- Green, T.B.; White, S.; Rao, S.; Mertens, P.P.; Adler, P.H.; Becnel, J.J. Biological and molecular studies of a cypovirus from the black fly Simulium ubiquitum (Diptera: Simuliidae). J. Invertebr. Pathol. 2007, 95, 26–32. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, T.; Xiao, Y.; Qin, F.; Lei, C.; Sun, X. Genomic and biological characterization of a new cypovirus isolated from Dendrolimus punctatus. PLoS ONE 2014, 9, e0113201. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.D.; Zhan, Z.G.; Guan, L.M.; Wang, J.C.; Yan, C.H.; Chen, J.H.; Li, J.H.; Zhou, X.; Jin, L. Study on proliferation characteristics of Daphnis nerii cypovirus-23 in Sf9 Cells. J. Agric. Biotechnol. 2021, 29, 772–779. (In Chinese) [Google Scholar]
- Nai, Y.S.; Lo, C.M. Persistent PnV (Perina nuda virus) infection in a heterologous Lymantria xylina cell line, NTU-LY. Biocontrol Sci. Technol. 2020, 30, 929–940. [Google Scholar] [CrossRef]
- Erez, T.; Chejanovsky, N. Infection of a lepidopteran cell line with deformed wing virus. Viruses 2020, 12, 739. [Google Scholar] [CrossRef]
- Agosto, J.A.M.; McCabe, E.R.B. Conserved family of glycerol kinase loci in Drosophila melanogaster. Mol. Genet. Metab. 2006, 88, 334–345. [Google Scholar] [CrossRef]
- Vatanparast, M.; Sajjadian, S.M.; Park, Y. Glycerol biosynthesis plays an essential role in mediating cold tolerance the red imported fire ant, Solenopsis invicta. Arch. Insect Biochem. Physiol. 2022, 109, e21861. [Google Scholar] [CrossRef]
- Majumdar, S.; Chowdhury, D.R.; Chakraborty, B.C.; Chowdhury, A.; Datta, S.; Banerjee, S. MiR-451a attenuates hepatic steatosis and hepatitis C virus replication by targeting glycerol kinase. J. Transl. Med. 2025, 23, 322. [Google Scholar] [CrossRef]
- Kikawada, T.; Saito, A.; Kanamori, Y.; Nakahara, Y.; Iwata, K.I.; Tanaka, D.; Watanabe, M.; Okuda, T. Trehalose transporter 1, a facilitated and high-capacity trehalose transporter, allows exogenous trehalose uptake into cells. Proc. Natl. Acad. Sci. USA 2007, 104, 11585–11590. [Google Scholar] [CrossRef]
- Marten, A.D.; Haslitt, D.P.; Martin, C.A.; Karthikeyan, A.; Swanson, D.H.; Kalera, K.; Johnson, U.G.; Swarts, B.M.; Conway, M.J. Trehalose supports the growth of Aedes aegypti cells and modifies gene expression and dengue virus type 2 replication. PLoS Pathog. 2025, 21, e1012795. [Google Scholar] [CrossRef]
- Kaur, A.; Singh, S.; Sharma, S.C. Unlocking Trehalose’s versatility: A comprehensive Journey from biosynthesis to therapeutic applications. Exp. Cell Res. 2024, 442, 114250. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, H.D.; Wang, Y.X.; Guo, Z.X.; Liu, Y.X.; Huang, Z.H.; Zhu, L.B.; Liu, M.H.; Liu, S.H.; Xu, J.P. Trehalose hydrolysis and transport-related genes promote Bombyx mori nucleopolyhedrovirus proliferation through the phosphoinositide 3-kinase-Akt signalling pathway in BmN cell. Dev. Comp. Immunol. 2023, 140, 104625. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Liu, X.J.; Shiotsuki, T.; Wang, Z.S.; Xu, X.; Huang, Y.P.; Li, M.W.; Li, K.; Tan, A.J. Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect Biochem. Mol. Biol. 2017, 81, 72–79. [Google Scholar] [CrossRef]
- Reis, M.A.; Noriega, D.D.; Alves, G.D.; Coelho, R.R.; Grossi-de-Sa, M.F.; Antonino, J.D. Why is oral-induced RNAi inefficient in Diatraea saccharalis? A possible role for DsREase and other nucleases. Pestic. Biochem. Physiol. 2022, 186, 105166. [Google Scholar] [CrossRef]
- Saito, Y.; Kamita, S.G.; Hammock, B.D.; Kunimi, Y.; Inoue, M.N.; Nakai, M. Juvenile hormone (JH) esterase activity but not JH epoxide hydrolase activity is downregulated in larval Adoxophyes honmai following nucleopolyhedroviruses infection. J. Insect Physiol. 2015, 80, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Dai, J.P.; Zhang, G.Z.; Singh, D.; Zhang, X.X.; Liang, Z.P. Mamestra brassicae multiple nucleopolyhedroviruses prevents pupation of Helicoverpa armigera by regulating juvenile hormone titer. Insects 2024, 15, 202. [Google Scholar] [CrossRef]
- Takatsuka, J.; Nakai, M.; Shinoda, T. A virus carries a gene encoding juvenile hormone acid methyltransferase, a key regulatory enzyme in insect metamorphosis. Sci. Rep. 2017, 7, 13522. [Google Scholar] [CrossRef]
- Gu, Y.F.; Yang, X.Y.; Liu, S.H.; Chen, X.W.; Liu, R.; Gao, J.L.; Zhong, Y.H.; Li, X.Y.; Han, W.S. RNAi-mediated knockdown of juvenile hormone acid methyltransferase depresses reproductive performance in female. Pestic. Biochem. Physiol. 2025, 211, 106420. [Google Scholar] [CrossRef]
- Zhu, S.M.; Liu, F.F.; Chen, X.Y.; Xia, S.S.; Wu, Y.T.; Tang, W.; Ren, C.H.; Wang, J.; Li, S. Inter-organelle communication dynamically orchestrates juvenile hormone biosynthesis and female reproduction. Natl. Sci. Rev. 2025, 12, nwaf022. [Google Scholar] [CrossRef]
- Wang, A.Y.; Yang, Y.X.; Zhang, Y.; Xue, C.; Cheng, Y.J.; Zhang, Y.F.; Zhang, W.J.; Zhao, M.; Zhang, J.H. Insecticide-induced sublethal effect in the fall armyworm is mediated by miR-9993/miR-2a-3p-FPPS/JHAMT-JH molecular module. Pestic. Biochem. Physiol. 2025, 210, 106400. [Google Scholar] [CrossRef]
- Shi, Z.K.; Wen, D.; Chang, M.M.; Sun, X.M.; Wang, Y.H.; Cheng, C.H.; Zhang, L.Q.; Zheng, A.H.; Zou, Z. Juvenile hormone-sensitive ribosomal activity enhances viral replication in Aedes aegypti. Msystems 2021, 6, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Liu, D.; Yuan, Y.Y.; Li, M.; Qiu, X.H. CYP6B6 is involved in esfenvalerate detoxification in the polyphagous lepidopteran pest, Helicoverpa armigera. Pestic. Biochem. Physiol. 2017, 138, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.F.; Yu, M.Y.; Li, M.; Reilly, C.A.; Qiu, X.H. CYP6B6 mediated adaptation to capsaicinoids in the generalist Helicoverpa armigera and specialist H. assulta: Transcriptional response and metabolic detoxification. Int. J. Biol. Macromol. 2025, 286, 138286. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, J.Y.; Wu, P.Z.; Zhang, Y.; Qiu, L.H. Functional study on candidate regulators mediating the expression of CYP6B7 induced by fenvalerate in a susceptible strain of Helicoverpa armigera. Pestic. Biochem. Physiol. 2024, 202, 105918. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zheng, J.Y.; Wu, P.Z.; Zhang, Y.; Qiu, L.H. A comparative study of transcriptional regulation mechanism of Cytochrome P450 CYP6B7 between resistant and susceptible strains of Helicoverpa armigera. J. Agric. Food Chem. 2023, 71, 9314–9323. [Google Scholar] [CrossRef]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the molecular determinants of bee sensitivity to neonicotinoid insecticides. Curr. Biol. 2018, 28, 1137–1143.e5. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Y.; Liu, Z.Y.; Zhu, Y.; Xiao, Y.; Xiao, W.F.; Tang, L.; Dong, Z.Q.; Pan, M.H.; Lu, C.; Chen, P. MicroRNA bmo-miR-31-5p inhibits apoptosis and promotes BmNPV proliferation by targeting the CYP9e2 gene of Bombyx mori. Pest Manag. Sci. 2024, 80, 4564–4574. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, Z.D.; Lv, X.X.; Liu, X.C.; Li, Y.F.; Tian, Z.; Zhang, Y.L.; Liu, J.Y. Molecular mechanism of λ-cyhalothrin detoxification by a delta-class Glutathione S-Transferase (PxGSTD3) from. J. Agric. Food Chem. 2025, 73, 6559–6566. [Google Scholar] [CrossRef]
- Haas, G.; Seiler, M.; Nguyen, J.; Troxler, L.; Pennarun, S.; Lefebvre, E.; Benamrouche, Y.; Loizeau, L.; Reinbolt, C.; Liang, M.; et al. Regulation of detoxifying enzymes expression and restriction of picorna-like virus infection by natural polysaccharide extracts in. Virology 2025, 607, 110513. [Google Scholar] [CrossRef]
- Gui, Z.Z.; Hou, C.X.; Liu, T.; Qin, G.X.; Li, M.W.; Jin, B. Effects of insect viruses and pesticides on Glutathione S-Transferase activity and gene expression in Bombyx mori. J. Econ. Entomol. 2009, 102, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Dalton, T.P. The role of cytochrome P450 enzymes in endogenous signalling pathways and environmental carcinogenesis. Nat. Rev. Cancer 2006, 6, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Kong, W.W.; Ling, B.; Wang, Z.Y.; Zhang, Y.; Guo, Z.X.; Liu, S.H.; Xu, J.P. Bmo-miR-3351 modulates glutathione content and inhibits BmNPV proliferation by targeting BmGSTe6 in Bombyx mori. Insect Sci. 2024, 31, 1378–1396. [Google Scholar] [CrossRef]
- Fan, C.C.; Cui, Z.X.; Yang, T.Y.; Sun, L.L.; Cao, C.W. UDP-glucuronosyltransferase is involved in susceptibility of Chironomus kiiensis Tokunaga, 1936 (Diptera: Chironomidae) to insecticides. Ecotoxicol. Environ. Saf. 2023, 263, 115353. [Google Scholar] [CrossRef]
- Zhu, F.F.; Han, J.Y.; Hong, J.D.; Cai, F.C.; Tang, Q.; Yu, Q.; Ma, S.S.; Liu, X.Y.; Huo, S.H.; Chen, K.P. Characterization of the UDP-glycosyltransferase UGT33D1 in silkworm. Insect Mol. Biol. 2024, 33, 697–707. [Google Scholar] [CrossRef]
- Shang, F.; Ding, B.Y.; Niu, J.Z.; Lu, J.M.; Xie, X.C.; Li, C.Z.; Zhang, W.; Pan, D.; Jiang, R.X.; Wang, J.J. microRNA maintains nutrient homeostasis in the symbiont-host interaction. Proc. Natl. Acad. Sci. USA 2024, 121, e2406925121. [Google Scholar] [CrossRef]
- Huang, H.; Lu-Bo, Y.; Haddad, G.G. A Drosophila ABC transporter regulates lifespan. PLoS Genet. 2014, 10, e1004844. [Google Scholar] [CrossRef]
- Jorajuria, S.; Dereuddre-Bosquet, N.; Naissant-Storck, K.; Dormont, D.; Clayette, P. Differential expression levels of MRP1, MRP4, and MRP5 in response to human immunodeficiency virus infection in human macrophages. Antimicrob. Agents Chemother. 2004, 48, 1889–1891. [Google Scholar] [CrossRef]
- Zhu, M.; Pan, J.; Zhang, M.T.; Tong, X.Y.; Zhang, Y.S.; Zhang, Z.Y.; Liang, Z.; Zhang, X.; Hu, X.L.; Xue, R.Y.; et al. Bombyx mori cypovirus (BmCPV) induces PINK1-Parkin mediated mitophagy via interaction of VP4 with host Tom40. Dev. Comp. Immunol. 2022, 126, 104244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, M.; Pan, J.; Qiu, Q.; Tong, X.; Hu, X.; Gong, C. BmCPV replication is suppressed by the activation of the NF-kappaB/autophagy pathway through the interaction of vsp21 translated by vcircRNA_000048 with ubiquitin carboxyl-terminal hydrolase. Insect Biochem. Mol. Biol. 2023, 156, 103947. [Google Scholar] [CrossRef]
- Choi, Y.; Bowman, J.W.; Jung, J.U. Autophagy during viral infection—A double-edged sword. Nat. Rev. Microbiol. 2018, 16, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tu, S.Y.; Ding, L.; Jin, M.L.; Chen, H.C.; Zhou, H.B. The role of autophagy in viral infections. J. Biomed. Sci. 2023, 30, 5. [Google Scholar] [CrossRef] [PubMed]
- Nardacci, R.; Ciccosanti, F.; Marsella, C.; Ippolito, G.; Piacentini, M.; Fimia, G.M. Role of autophagy in HIV infection and pathogenesis. J. Intern. Med. 2017, 281, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberger, A.J.B.; Sanyal, A.; Saminathan, A.; Bloyet, L.M.; Stumpf, S.; Liu, Z.M.; Ojha, R.; Patjas, M.T.; Geneid, A.; Scanavachi, G.; et al. SARS-CoV-2 requires acidic pH to infect cells. Proc. Natl. Acad. Sci. USA 2022, 119, 422–432. [Google Scholar] [CrossRef]
- Doyle, C.A.; Busey, G.W.; Iobst, W.H.; Kiessling, V.; Renken, C.; Doppalapudi, H.; Stremska, M.E.; Manjegowda, M.C.; Arish, M.; Wang, W.M.; et al. Endosomal fusion of pH-dependent enveloped viruses requires ion channel TRPM7. Nat. Commun. 2024, 15, 8479. [Google Scholar] [CrossRef]
- Li, Y.H.; Li, C.Y.; Zhao, C.C.; Wu, J.Y.; Zhu, Y.; Wang, F.; Zhong, J.P.; Yan, Y.; Jin, Y.L.; Dong, W.R.; et al. Coronavirus M protein promotes mitophagy over virophagy by recruiting PDPK1 to phosphorylate SQSTM1 at T138. Nat. Commun. 2024, 15, 8927. [Google Scholar] [CrossRef]
- Pesti, I.; Barczánfalvi, G.; Dulka, K.; Kata, D.; Farkas, E.; Gulya, K. Bafilomycin 1A affects p62/SQSTM1 autophagy marker protein level and autophagosome puncta formation oppositely under various inflammatory conditions in cultured rat microglial cells. Int. J. Mol. Sci. 2024, 25, 8265. [Google Scholar] [CrossRef]
- Tan, X.Y.; Wang, X.; Liu, Q.S.; Xie, X.Q.; Li, Y.; Li, B.Q.; Li, Z.Q.; Xia, Q.Y.; Zhao, P. Inhibition of silkworm vacuolar-type ATPase activity by its inhibitor Bafilomycin A1 induces caspase-dependent apoptosis in an embryonic cell line of silkworm. Arch. Insect Biochem. Physiol. 2018, 99, e21507. [Google Scholar] [CrossRef]
- Mauvezin, C.; Neufeld, T.P. Bafilomycin A1 disrupts autophagic flux by inhibiting both V-ATPase-dependent acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome fusion. Autophagy 2015, 11, 1437–1438. [Google Scholar] [CrossRef]
- Song, B.Y.W.; Korolkova, O. Bafilomycin A1 inhibits HIV-1 infection by disrupting lysosomal cholesterol transport. Viruses 2024, 16, 1374. [Google Scholar] [CrossRef]
- Jin, M.H.; Shan, Y.X.; Peng, Y.; Wang, W.H.; Zhang, H.H.; Liu, K.Y.; Heckel, D.G.; Wu, K.M.; Tabashnik, B.E.; Xiao, Y.T. Downregulation of a transcription factor associated with resistance to Bt toxin Vip3Aa in the invasive fall armyworm. Proc. Natl. Acad. Sci. USA 2023, 120, e2306932120. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).