
DNA-Programmable Oligonucleotide Insecticide Eriola-11 Targets Mitochondrial 16S rRNA and Exhibits Strong Insecticidal Activity Against Woolly Apple Aphid (Eriosoma lanigerum) Hausmann

 ,
, 
 and
and 
Abstract
1. Introduction
2. Results
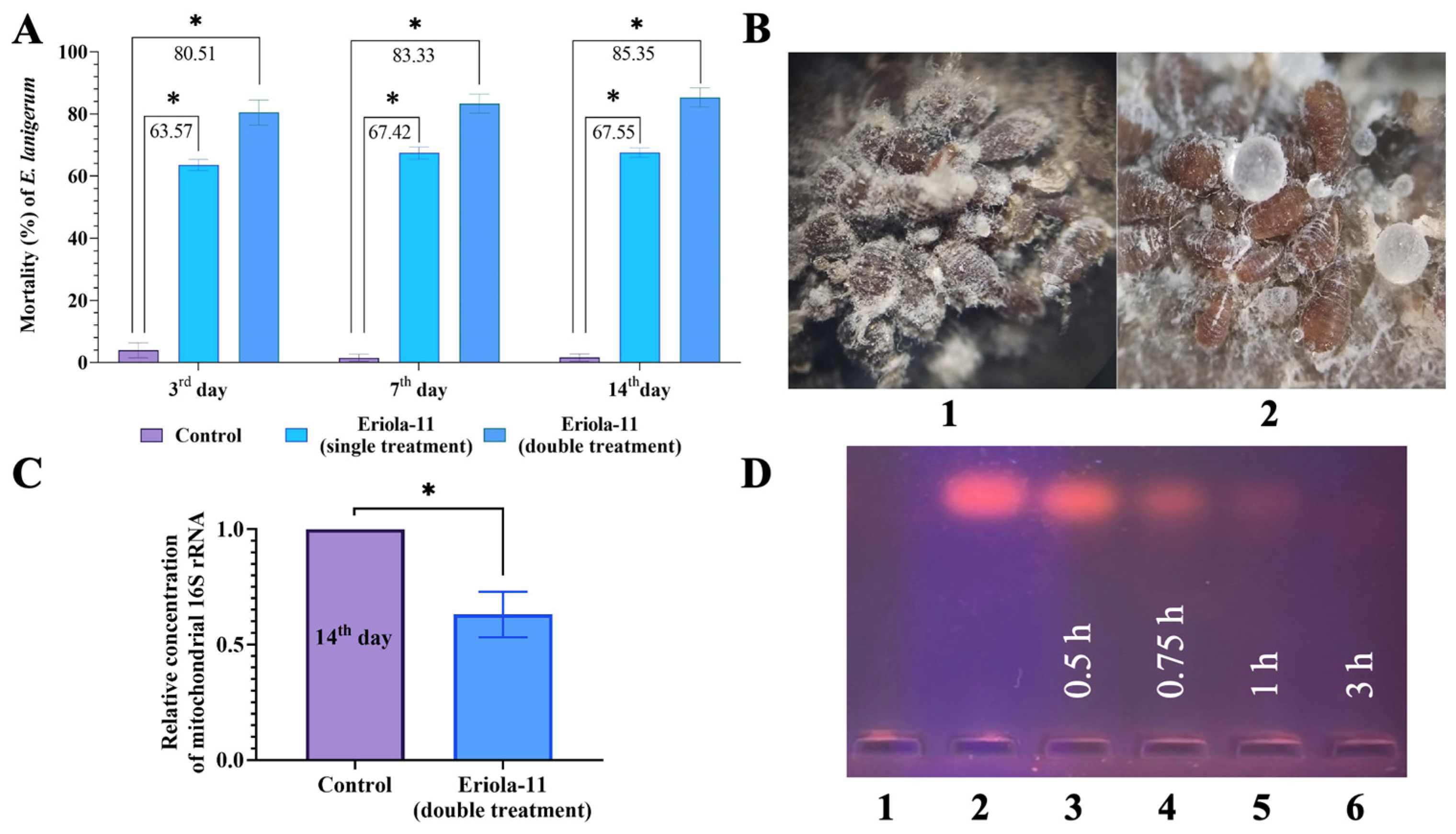
2.1. The Olinscide Eriola-11 Shows Pronounced Insecticidal Effects and Changes Morphology of E. lanigerum After Contact Application
2.2. Olinscide Eriola-11 Significantly Decreases the Concentration of the Mitochondrial 16S rRNA
2.3. Fast Biodegradability of Olinscide Eriola-11 via Tissue Homogenate Deoxyribonucleases
3. Discussion
4. Materials and Methods
4.1. Insect
4.2. DNA Synthesis and Mass Spectrometry Analysis of Oligonucleotides
4.3. Application of Eriola-11 as a Contact Insecticide
4.4. Reverse Transcription and Real-Time PCR (RT-PCR) for 16S rRNA Quantification
4.5. Analysis of DNase Activity
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damavandian, M.R.; Pringle, K.L. The field biology of subterranean populations of the woolly apple aphid, Eriosoma lanigerum (Hausmann) (Hemiptera: Aphididae), in South African apple orchards. Afr. Entomol. 2007, 15, 287–294. [Google Scholar] [CrossRef]
- Singh, J.; Chandel, R.S.; Sharma, P.L. Spatial distribution and interaction of Eriosoma lanigerum and Aphelinus mali on apple under dry- temperate conditions of India. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 88, 383–390. [Google Scholar] [CrossRef]
- Tan, X.M.; Yang, Z.S.; Zhou, H.; Yang, Q.M.; Zhou, H.X. Resistance performance of four principal apple cultivars to woolly apple aphid, Eriosoma lanigerum (Hemiptera: Pemphigidae), by simulated seasonal temperature in northern China. Arthropod-Plant Interact. 2021, 15, 59–69. [Google Scholar] [CrossRef]
- Sayimov, N.; Anorbayev, A.; Bababekov, K. Biological properties and influencing factors of woolly apple aphid (Eriosoma lanigerum Hausm) in the condition of orchard agrobiocenosis. E3S Web Conf. 2021, 244, 02009. [Google Scholar] [CrossRef]
- Khan, A.A.; Kundoo, A.A.; Nissar, M.; Mushtaq, M. Sucking Pests of Temperate Fruits. In Sucking Pests of Crops; Omkar, Ed.; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Stokwe, N.F.; Malan, A.P. Laboratory bioassays to determine susceptibility of woolly apple aphid, Eriosoma lanigerum (Hausmann) (Hemiptera: Aphididae), to entomopathogenic nematodes. Afr. Entomol. 2017, 25, 123–136. [Google Scholar] [CrossRef]
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchard IPM program. BioControl 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Stokwe, N.F.; Malan, A.P. Woolly apple aphid, Eriosoma lanigerum (Hausmann), in South Africa: Biology and management practices, with focus on the potential use of entomopathogenic nematodes and fungi. Afr. Entomol. 2016, 24, 267–278. [Google Scholar] [CrossRef]
- Jovičić, I. Eriosoma lanigerum (Woolly Aphid); CABI Compendium: Oxford, UK, 2024. [Google Scholar] [CrossRef]
- Van Emden, H.F.; Harrington, R. Aphids as Crop Pests; CABI: Wallingford, UK, 2017. [Google Scholar] [CrossRef]
- Ali, J. The Peach Potato Aphid (Myzus persicae): Ecology and Management, 1st ed.; CRC Press: Boca Raton, FL, USA, 2023; p. 132. [Google Scholar] [CrossRef]
- Tun, K.M.; Clavijo McCormick, A.; Jones, T.; Garbuz, S.; Minor, M. Honeydew deposition by the giant willow aphid (Tuberolachnus salignus) affects soil biota and soil biochemical properties. Insects 2020, 11, 460. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, P. Observation on the Influence of Apple Woolly Aphid (Eriosoma lanigerum Hausm.) on Vegetative Parameters of Grafted Plants in Container Production. Proc. Bulg. Acad. Sci. 2024, 77, 1251–1259. [Google Scholar] [CrossRef]
- Yang, L.; Ali, J.; Ahmad, B.; Yang, S.; Huang, J.; Zhao, J.; Alam, A.; Khan, K.; Ghramh, H.; Rahman, N.; et al. Garlic as a companion plant for suppressing Myzus persicae infestation in Brassica rapa. Crop Prot. 2025, 187, 106970. [Google Scholar] [CrossRef]
- Alins, G.; Lordan, J.; Rodríguez-Gasol, N.; Arnó, J.; Peñalver-Cruz, A. Earwig Releases Provide Accumulative Biological Control of the Woolly Apple Aphid over the Years. Insects 2023, 14, 890. [Google Scholar] [CrossRef]
- Jayasinghe, W.H.; Akhter, M.S.; Nakahara, K.; Maruthi, M.N. Effect of aphid biology and morphology on plant virus transmission. Pest Manag. Sci. 2022, 78, 416–427. [Google Scholar] [CrossRef]
- Shaw, B.; Nagy, C.; Fountain, M.T. Organic Control Strategies for Use in IPM of Invertebrate Pests in Apple and Pear Orchards. Insects 2021, 12, 1106. [Google Scholar] [CrossRef]
- Šarac, V.; Narandžić, T.; Rodić, V.; Popović, B.M.; Uka, D.; Tomaš Simin, M.; Ljubojević, M. Harnessing Koelreuteria paniculata Seed Extracts and Oil for Sustainable Woolly Apple Aphid Control. Horticulturae 2024, 10, 826. [Google Scholar] [CrossRef]
- Lordan, J.; Alegre, S.; Gatius, F.; Sarasúa, M.J.; Alins, G. Woolly apple aphid Eriosoma lanigerum Hausmann ecology and its relationship with climatic variables and natural enemies in Mediterranean areas. Bull. Entomol. Res. 2015, 105, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bangels, E.; Alhmedi, A.; Akkermans, W.; Bylemans, D.; Belien, T. Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali. Insects 2021, 12, 479. [Google Scholar] [CrossRef] [PubMed]
- Stehle, S.; Schulz, R. Agricultural insecticides threaten surface waters at the global scale. Proc. Natl. Acad. Sci. USA 2015, 112, 5750–5755. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a reduced reliance on conventional pesticides in European agriculture. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef]
- Ryalls, J.M.W.; Garratt, M.P.D.; Spadaro, D.; Mauchline, A.L. The benefits of integrated pest management for apple depend on pest type and production metrics. Front. Sustain. Food Syst. 2024, 8, 1321067. [Google Scholar] [CrossRef]
- Ansari, I.; El-Kady, M.M.; Arora, C.; Sundararajan, M.; Maiti, D.; Khan, A. A review on the fatal impact of pesticide toxicity on environment and human health. In Global Climate Change; Singh, S., Singh, P., Rangabhashiyam, S., Srivastava, K.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 361–391. [Google Scholar] [CrossRef]
- Punniyakotti, P.; Vinayagam, S.; Rajamohan, R.; Priya, S.D.; Moovendhan, M.; Sundaram, T. Environmental Fate and Ecotoxicological Behaviour of Pesticides and Insecticides in Non-Target Environments: Nanotechnology-Based Mitigation Strategies. J. Environ. Chem. Eng. 2024, 12, 113349. [Google Scholar] [CrossRef]
- Maffia, A.; Marra, F.; Canino, F.; Battaglia, S.; Mallamaci, C.; Oliva, M.; Muscolo, A. Humic Substances from Waste-Based Fertilizers for Improved Soil Fertility. Agronomy 2024, 14, 2657. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Ahmad, F.A.; Alsayegh, A.A.; Zeyaullah, M.; AlShahrani, A.M.; Muzammil, K.; Saati, A.A.; Wahab, S.; Elbendary, E.Y.; Kambal, N.; et al. Pesticides impacts on human health and the environment with their mechanisms of action and possible countermeasures. Heliyon 2024, 10, e29128. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Iqbal, K.; Sohail, M.; Hussain Rind, K.; Habib, S.S. Agrochemical contamination and fish health: Eco-toxicological impacts and mitigation strategies. Chem. Ecol. 2025, 41, 959–993. [Google Scholar] [CrossRef]
- Khursheed, A.; Rather, M.A.; Jain, V.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant based natural products as potential ecofriendly and safer biopesticides: A comprehensive overview of their advantages over conventional pesticides, limitations and regulatory aspects. Microb. Pathog. 2022, 173 Pt A, 105854. [Google Scholar] [CrossRef]
- Lazarević-Pašti, T.; Milanković, V.; Tasić, T.; Petrović, S.; Leskovac, A. With or Without You?—A Critical Review on Pesticides in Food. Foods 2025, 14, 1128. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Novikov, I.A.; Yatskova, E.V.; Bilyk, A.I.; Sharmagiy, A.K.; Gal’chinsky, N.V. Potent and selective ‘genetic zipper’ method for DNA-programmable plant protection: Innovative oligonucleotide insecticides against Trioza alacris Flor. Chem. Biol. Technol. Agric. 2024, 11, 144. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Puzanova, Y.V.; Gal’chinsky, N.V. The ‘genetic zipper’ method offers a cost-effective solution for aphid control. Front. Insect Sci. 2024, 4, 1467221. [Google Scholar] [CrossRef]
- Deng, Z.L.; Münch, P.C.; Mreches, R.; McHardy, A.C. Rapid and accurate identification of ribosomal RNA sequences via deep learning. Nucleic Acids Res. 2022, 50, e60. [Google Scholar] [CrossRef] [PubMed]
- Szaflarski, W.; Leśniczak-Staszak, M.; Sowiński, M.; Ojha, S.; Aulas, A.; Dave, D.; Malla, S.; Anderson, P.; Ivanov, P.; Lyons, S.M. Early rRNA processing is a stress-dependent regulatory event whose inhibition maintains nucleolar integrity. Nucleic Acids Res. 2022, 50, 1033–1051. [Google Scholar] [CrossRef]
- Kumar, H.; Gal’chinsky, N.; Sweta, V.; Negi, N.; Filatov, R.; Chandel, A.; Ali, J.; Oberemok, V.; Laikova, K. Perspectives of RNAi, CUADb and CRISPR/Cas as Innovative Antisense Technologies for Insect Pest Control: From Discovery to Practice. Insects 2025, 16, 746. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Laikova, K.V.; Gal’chinsky, N.V. Contact unmodified antisense DNA (CUAD) biotechnology: List of pest species successfully targeted by oligonucleotide insecticides. Front. Agron. 2024, 6, 1415314. [Google Scholar] [CrossRef]
- Gal’chinsky, N.V.; Yatskova, E.V.; Novikov, I.A.; Sharmagiy, A.K.; Plugatar, Y.V.; Oberemok, V.V. Mixed insect pest populations of Diaspididae species under control of oligonucleotide insecticides: 3′-end nucleotide matters. Pestic. Biochem. Physiol. 2024, 200, 105838. [Google Scholar] [CrossRef] [PubMed]
- Puzanova, Y.V.; Novikov, I.A.; Bilyk, A.I.; Sharmagiy, A.K.; Plugatar, Y.V.; Oberemok, V.V. Perfect complementarity mechanism for aphid control: Oligonucleotide insecticide macsan-11 selectively causes high mortality rate for Macrosiphoniella sanborni Gillette. Int. J. Mol. Sci. 2023, 24, 11690. [Google Scholar] [CrossRef] [PubMed]
- Gavrilova, D.; Grizanova, E.; Novikov, I.; Laikova, E.; Zenkova, A.; Oberemok, V.; Dubosvkiy, I. Antisense DNA acaricide targeting pre-rRNA of two-spotted spider mite Tetranychus urticae as efficacy-enhancing agent of fungus Metarhizium robertsii. J. Invertebr. Pathol. 2025, 211, 108297. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M. Mitochondria. In The Cell: A Molecular Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9896/ (accessed on 18 June 2025).
- Oberemok, V.V.; Gal’chinsky, N.V.; Novikov, I.A.; Sharmagiy, A.K.; Yatskova, E.V.; Laikova, E.V.; Plugatar, Y. rRNA-specific antisense DNA and dsDNA trigger rRNA biogenesis and cause potent insecticidal effect on insect pest C. hesperidum L. bioRxiv 2024, 10, 618468. [Google Scholar]
- Dias, N.; Stein, C.A. Antisense oligonucleotides: Basic concepts and mechanisms. Mol. Cancer Ther. 2002, 1, 347–355. [Google Scholar]
- Britannica. Available online: https://www.britannica.com/science/lipid/Waxes (accessed on 10 April 2025).
- Moss, R.; Jackson, R.R.; Pollard, S.D. Mask of wax: Secretions of wax conceal aphids from detection by spider’s eyes. N. Z. J. Zool. 2006, 33, 215–220. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Laikova, K.V.; Andreeva, O.A.; Gal’chinsky, N.V. Biodegradation of insecticides: Oligonucleotide insecticides and double-stranded RNA biocontrols paving the way for eco-innovation. Front. Environ. Sci. 2024, 12, 1430170. [Google Scholar] [CrossRef]
- Patil, V.; Jangra, S.; Ghosh, A. Advances in Antisense Oligo Technology for Sustainable Crop Protection. Crit. Rev. Plant Sci. 2024, 43, 405–427. [Google Scholar] [CrossRef]
- Paul, N.M.; Yoder, R.J.; Callam, C.S. Incorporating Chemical Structure Drawing Software throughout the Organic Laboratory Curriculum. J. Chem. Educ. 2019, 96, 2638–2642. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Jiang, Q.; Rao, M.; Sheng, Z.; Zhang, Y.; Du, W.; Hao, H.; Zhao, X.; Xu, Z.; et al. Identification of Valid Housekeeping Genes for Real-Time Quantitative PCR Analysis of Collapsed Lung Tissues of Neonatal Somatic Cell Nuclear Transfer-Derived Cattle. Cell Reprogram. 2015, 17, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Rebouças, E.D.L.; Costa, J.J.D.N.; Passos, M.J.; Passos, J.R.D.S.; Hurk, R.V.D.; Silva, J.R.V. Real time PCR and importance of housekeepings genes for normalization and quantification of mRNA expression in different tissues. Braz. Arch. Biol. Technol. 2013, 56, 143–154. [Google Scholar] [CrossRef]
- Paule, M.R.; White, R.J. Transcription by RNA polymerases I and III. Nucleic Acids Res. 2000, 28, 1283–1298. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M., Jr.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb Protoc. 2010, 2010, pdb.prot5439. [Google Scholar] [CrossRef] [PubMed]
- Arezi, B.; Hogrefe, H. Novel mutations in Moloney Murine Leukemia Virus reverse transcriptase increase thermostability through tighter binding to template-primer. Nucleic Acids Res. 2009, 37, 473–481. [Google Scholar] [CrossRef]
- Khehra, N.; Padda, I.S.; Swift, C.J. Polymerase Chain Reaction (PCR) [Updated 6 March 2023]. In StatPearls; StatPearls: Treasure Island, FL, USA, 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK589663/ (accessed on 12 June 2025).
- Smith, D.R. Agarose gel electrophoresis. Methods Mol. Biol. 1993, 18, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Appling, D.R. Prism 4 GraphPad Software, Inc. 5755 Oberlin Drive, #110, San Diego, CA, 92121. www.graphpad.com. See web site for pricing information. J. Am. Chem. Soc. 2003, 125, 10482. [Google Scholar]


{kind=link}
{kind=link}
| Olinscides | Theoretical m/z Ratio | Received m/z Ratio |
|---|---|---|
| Eriola-11 | 3323.61 | 3329.56 |
| Primers | Theoretical m/z Ratio | Received m/z Ratio |
| Eriola-F | 5511.96 | 5505.47 |
| Eriola-R | 5373.92 | 5365.84 |
| Gene | Primer | Primer Sequence (5′-3′) | Tm | PCR Product | GenBank (Sequence ID) |
|---|---|---|---|---|---|
| 16S rRNA | Eriola-F | TATAGGATCTGCTCAATG | 50 °C | 100 bp | AF275225.1 |
| Eriola-R | TCTTTCATTCAAGTCTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberemok, V.; Laikova, K.; Andreeva, O.; Dmitrienko, A.; Rybareva, T.; Ali, J.; Gal’chinsky, N. DNA-Programmable Oligonucleotide Insecticide Eriola-11 Targets Mitochondrial 16S rRNA and Exhibits Strong Insecticidal Activity Against Woolly Apple Aphid (Eriosoma lanigerum) Hausmann. Int. J. Mol. Sci. 2025, 26, 7486. https://doi.org/10.3390/ijms26157486
Oberemok V, Laikova K, Andreeva O, Dmitrienko A, Rybareva T, Ali J, Gal’chinsky N. DNA-Programmable Oligonucleotide Insecticide Eriola-11 Targets Mitochondrial 16S rRNA and Exhibits Strong Insecticidal Activity Against Woolly Apple Aphid (Eriosoma lanigerum) Hausmann. International Journal of Molecular Sciences. 2025; 26(15):7486. https://doi.org/10.3390/ijms26157486
Chicago/Turabian StyleOberemok, Vol, Kate Laikova, Oksana Andreeva, Anastasia Dmitrienko, Tatiana Rybareva, Jamin Ali, and Nikita Gal’chinsky. 2025. "DNA-Programmable Oligonucleotide Insecticide Eriola-11 Targets Mitochondrial 16S rRNA and Exhibits Strong Insecticidal Activity Against Woolly Apple Aphid (Eriosoma lanigerum) Hausmann" International Journal of Molecular Sciences 26, no. 15: 7486. https://doi.org/10.3390/ijms26157486
APA StyleOberemok, V., Laikova, K., Andreeva, O., Dmitrienko, A., Rybareva, T., Ali, J., & Gal’chinsky, N. (2025). DNA-Programmable Oligonucleotide Insecticide Eriola-11 Targets Mitochondrial 16S rRNA and Exhibits Strong Insecticidal Activity Against Woolly Apple Aphid (Eriosoma lanigerum) Hausmann. International Journal of Molecular Sciences, 26(15), 7486. https://doi.org/10.3390/ijms26157486