
NRF2 Dysregulation and Therapeutic Insights Across Chronic Kidney Diseases


Abstract
1. Introduction
2. NRF2 Regulation
2.1. Transcriptional, Epigenetic, and Post-Transcriptional Regulation
2.2. KEAP1-Dependent Regulation
2.3. KEAP1-Independent Regulation
3. Mechanisms of NRF2 Dysregulation in Chronic Kidney Diseases
3.1. Redox Balance and Antioxidant Defense
3.2. Inflammation
3.3. Fibrosis
3.4. Metabolic Signaling
3.5. NRF2 Dysregulation in CKD Stages
4. Therapeutic Insights Across CKD Etiologies
4.1. Diabetic Kidney Disease (DKD)

{kind=link}
| NRF2 Modulators | Chemical Class | Study Design | Dosage and Duration of Treatment | Pathway/ Mechanism | Reference |
|---|---|---|---|---|---|
| Lixisenatide | Polypeptide | In vivo (STZ-induced diabetic rats) | 10 µg/kg for 4 weeks | Modulate NRF2/HO-1 signaling pathway | [97] |
| Telmisartan | Benzimidazole | In vivo (STZ-induced diabetic rats) | 5 or 10 mg/kg for 8 weeks | [98] | |
| Digitoflavone | Flavonoid | In vitro (SV40-MES13 cells) In vivo (STZ-induced diabetic mice) | 25 or 50 mg/kg | [99] | |
| Notoginsenoside R1 | Saponin | In vitro (HK-2 cells) In vivo (db/db mice) | 30 mg/kg/day for 20 weeks | [100] | |
| Sinapic acid | Cinnamate | In vivo (STZ-induced diabetic rats) | 20 or 40 mg/kg for 8 weeks | [101] | |
| Compound centella | Herbal and Dietary Supplements | In vivo (STZ-induced diabetic rats) | 0.8 g Centella asiatica, 0.8 g Astragalus membranaceus, and 0.4 g Tripterygium wildorfii per rat for 112 days | [102] | |
| Phosphocreatine | Amidine | In vitro (NRK-52E cells) In vivo (STZ-induced diabetic rats) | In vitro: 10, 20, or 40 mm for 4 h In vivo: 25 or 50 mg/kg for 72 h | [103] | |
| Artemisinin | Sesquiterpene | In vivo (STZ-induced diabetic rats) | 25, 50 or 75 mg/kg for 8 weeks | [104] | |
| Coptisine | Alkaloid | In vivo (STZ-induced diabetic rats) | 25 or 50 mg/kg/day for 8 weeks | [105] | |
| L-ergothioneine and metformin | Sulfur Compound Amidine | In vivo (STZ-induced diabetic rats) | 35 mg/kg of L-egt, 500 mg/kg of metformin or both for 7 weeks | [95] | |
| Ethyl ferulate | Cinnamate | In vivo (STZ-induced diabetic rats) | 50, 75, or 100 mg/kg for 4 weeks | [106] | |
| Isoeucommin A | Polyphenol | In vitro (HRMC and RTECs) In vivo (STZ-induced diabetic rat) | In vitro: 31.25, 61.3, or 125 µm for 2 h In vivo: 2.5, 5, or 10 mg/kg/day for 8 weeks | [107] | |
| Kaempferol | Flavonoid | In vivo (STZ-induced diabetic rats) | 200 mg/kg for 8 weeks | [108] | |
| Beta-cryptoxanthin | Tetraterpenoid | In vitro (human podocyte cells) In vivo (db/db mice) | In vitro: 10 µm for 2 h In vivo: 10 mg/kg for 6 weeks | [109] | |
| Simvastatin | Naphthalene | In vivo (STZ-induced diabetic rats) | 10 mg/kg for 8 weeks | [110] | |
| Saxagliptin | Peptide | In vivo (STZ-induced diabetic rats) | 10 mg/kg 45 min before ischemia | [111] | |
| Maackiain | Isoflavonoid | In vivo (STZ-induced diabetic rats) | 10 or 20 mg/kg for7 weeks | [112] | |
| Trametenolic acid | Triterpene | In vivo (db/db mice) | 10 mg/kg/day for 4 weeks | Modulate NRF2-mediated downregulation of Nf-kB | [113] |
| Akebia saponin D | Saponin | In vitro (HK-2 cells) In vivo (STZ-induced diabetic rats) | In vitro: 15, 30 or 60 µg/mL In vivo: 50, 100, or 150 mg/kg for 8 weeks | [114] | |
| Salvianolic acid | Cinnamate | In vivo (STZ-induced diabetic rats) | 3 mg/kg for 18 weeks | [96] | |
| Carnosic acid | Diterpene | In vitro (SV40 MES 13 cells) In vivo (STZ-induced diabetic mice) | In vitro: 2.5, 5, 10, or 15 µm for 24 h In vivo: 15 or 30 mg/kg/day for 20 weeks | [115] | |
| Phloretamide | Phenolic amide | In vivo (STZ-induced diabetic rats) | 200 mg/kg, twice a week for 12 weeks | [116] | |
| Gentisic acid | Hydroxybenzoate | In vivo (STZ-induced diabetic rats) | 100 mg/kg for 2 weeks | [117] | |
| Eriodictyol | Flavonoid | In vivo (STZ-induced diabetic rats) | 20 mg/kg for 12 weeks | [118] | |
| Resveratol | Stilbenoid | In vivo (STZ-induced diabetic rat) | 5 mg/kg/day for 30 days | Modulate KEAP1 | [119] |
| AB-38b | Aromatic diester | In vitro (mouse GMCs) In vivo (STZ-induced diabetic mice) | In vitro: 2.5, 5, or 10 μm In vivo: 10, 20, or 40 mg/kg/day for 8 weeks | [120] | |
| Epigallocatechin-3-gallate | Flavonoid | In vitro (NRK-52E cells) In vivo (STZ-induced diabetic rats) In vivo (STZ-induced diabetic mice) | In vitro: 5 μm for 24 h In vivo: 100 mg/kg for 30 days In vivo: 100 mg/kg for 24 weeks | [90,91] | |
| Shenkang Injection | Herbal extract | In vivo (STZ-induced diabetic rats) | 5 mL/kg for 16 weeks | [121] | |
| Icariin | Flavonoid | In vitro (human glomerular mesangial cells) In vivo (STZ-induced diabetic rats) | In vitro: 1, 3, or 10 μm for 48 h In vivo: 20, 40, or 80 mg/kg for 9 weeks | [122] | |
| Mexacalcitol | Steroid derivative | In vivo (spontaneously diabetic torii rat model) | 0.2 μg/kg 3 times/week for 10 weeks | [89] | |
| Chrysophanol | Quinone | In vivo (STZ-induced diabetic mice) | 2.5, 5, or 10 mg/kg/day for 8 weeks | [123] | |
| Tilianin | Glycoside | In vivo (STZ-induced diabetic rats) | 10 or 20 mg/kg for 28 days | [124] | |
| Caffeoylisocitric acid | Cinnamic acid | In vitro (human GMCs) | 10 µm for 24 h | [125] | |
| Deferiprone | Pyridine | In vivo (STZ-induced diabetic rats) | 50 mg/kg for 16 weeks | Promote nuclear accumulation/translocation of NRF2 | [126] |
| Liraglutide (GLP-1 receptor agonist) | Peptide | In vitro (human glomerular mesangial cells) In vivo (STZ-induced diabetic rats) | In vitro: 100 or 1000 nm for 48 h In vivo: 200 μg/kg/day, duration unknown | [127] | |
| Tert-butylhydroquinone (tBHQ) | Phenol | In vitro (HK2 cells) In vivo (STZ-induced diabetic rats) In vivo (STZ-induced diabetic rats) | In vitro: 10, 20, 30, or 40 μm for 2 h In vivo: 50 mg/kg every other day for 10 weeks 1% tBHQ for 12 weeks | [128,129] | |
| Biphenyl Diester Derivative-39 | Aromatic ester | In vivo (STZ-induced diabetic rats) | 15 or 45 mg/kg/day for 8 weeks | [130] | |
| MitoQ | Quinone derivative | In vitro (HK-2 cells) In vivo (db/db mice) | In vitro: concentration unknown, treated for 2 h In vivo: 12 mg/kg twice weekly for 12 weeks | [131] | |
| Chlorogenic acid | Carboxylic Acid | In vitro (HBZY-1 cells) In vivo (STZ-induced diabetic rats) | In vitro: 5, 10, 25, 50, 100, or 200 μm for 24 h In vivo: 10 mg/kg/day for 2 weeks | [132] | |
| SP600125 | Anthrapyrazolone | In vitro (mouse GMCs) In vivo (STZ-induced diabetic rats) | In vitro: 10 μm for 48 h In vivo: 5 mg/kg every other day for 24 weeks | [133] | |
| Astaxanthin | Carotenoid | In vivo (STZ-induced diabetic rats) | 25 mg/kg/day for 12 weeks | [134] | |
| Myricetin | Flavonoid | In vivo (STZ-induced diabetic mice) | 100 mg/kg/day for 6 months | [135] | |
| Obacunone | Triterpene | In vitro (NRK-52E cells) | 40 μm for 48 h | [136] | |
| Triptolide | Diterpene | In vitro (MPC-5 cells) In vivo (STZ-induced diabetic mice) | In vitro: 10 μm for 48 h In vivo: 100 μg/kg/day for 12 weeks | Inhibit NLRP3 inflammasome | [137] |
| Solasonine | Steroidal glycoside | In vitro (MPC-5 cells) | 5, 10, or 20 μm for 48 h | [138] | |
| Minocycline | Naphthalene | In vitro (human and mouse podocytes) In vivo (db/db mice and STZ-induced diabetic mice) | In vitro: 10 μm for 24 h In vivo: 5 mg/kg/day for 10 weeks | [139] | |
| AB-38b | Aromatic diester | In vitro (mouse GMCs) In vivo (STZ-induced diabetic mice) | Not specified | [120,140] | |
| WJ-39 | Synthetic small molecule | In vitro (rat mesangial cells) In vivo (STZ-induced diabetic mice) | In vitro: 1, 10, or 100 μm for 24, 48, and 72 h. In vivo: 10, 20, or 40 mg/kg for 12 weeks | [141] | |
| Berberine | Alkaloids | In vivo (STZ-induced diabetic hamsters) | 100 or 200 mg/kg for 8 weeks | [142] | |
| Icariin | Flavonoid | In vitro (MPC-5 cells) In vivo (STZ-induced diabetic rats) | In vitro: 1, 3, or 10 μm for 48 h In vivo: 20, 40, or 80 mg/kg for 8 weeks | [143] | |
| Quercetin | Flavonoid | In vitro (HK-2 cells) In vivo (STZ-induced diabetic rats) In vitro (HK-2 cells) In vivo (STZ-induced diabetic rats | In vitro: 25 μm for 48 h In vivo: 100 mg/kg for 12 weeks In vitro: 6.25, 12.5, 25, 50, or 100 μm for 48 h In vivo: 25 or 100 mg/kg for 12 weeks | Inhibit ferroptosis | [144,145] |
| Triptolide | Diterpene | In vitro (human podocyte cells) In vivo (db/db mice) | In vitro: 5 nm for 48 h In vivo: 50 μg/kg/day for 8 weeks | [146] | |
| Paricalcitol | Steroids and derivative | In vitro (HK-2 cells) In vivo (db/db mice) | In vitro: 0.1 μm for 48 h In vivo: 0.1 µg/kg 5 consecutive days/week for 10 weeks | [147] | |
| Carnosine | Peptide | In vitro (HK-2 cells) In vivo (STZ-induced diabetic mice) | Not specified | [148] | |
| Empagliflozin | Glucoside | In vitro (HK-2 cells) In vivo (STZ-induced diabetic mice) | In vitro: 500 nm for 24 h In vivo: 10 mg/kg/day for 8 weeks | [149] | |
| Emodin | Quinone | In vitro (HK-2 cells) In vivo (STZ-induced diabetic rats) | In vitro: 40 μm for 48 h In vivo: 12 weeks | [150] | |
| Crocin | Carotenoid | In vivo (db/db mice) | 40 mg/kg for 8 weeks | Modulate PI3K/AKT/NRF2 | [151] |
| Carnosine | Peptide | In vitro (MPC-5 cells) | 5, 10, 20, or 30 mm for 48 h | [152] | |
| Beta-sitosterol | Steroids and derivative | In vivo (high fat- and sucrose-induced diabetic rats) | 20 or 50 mg/kg/day for 30 days | Modulate TGF-β1/Nrf2/SIRT1/p53 pathway | [153] |
| Gastrodin | Glucoside | In vivo (STZ-induced diabetic mice) | 5, 10 or 20 mg/kg for 6 weeks | Modulate AMPK/NRF2 pathway | [154] |
| Neferine | Alkaloid | In vivo (STZ-induced diabetic mice) | 60, 120 or 240 mg/kg for 12 weeks | Reduce expression of miR-17-5p | [155] |
| Theaflavin-3,3′-digallate | Flavonoid | In vitro (HepG2 and HK-2 cells) In vivo (STZ-induced diabetic rats) | In vitro: not specified In vivo: 10 or 20 mg/kg/day for 6 weeks | Activate Circ-ITCH and NRF2 | [156] |
| Astragaloside IV | Saponin | In vitro (mouse podocyte cells) In vivo (STZ-induced diabetic mice) | In vitro: 0, 0.3, 1.0, 3.0, 10, 20, 40, 80, or 100 μmol/L for 24 h In vivo: 6 mg/kg/day for 10 weeks | Modulate NRF2-ARE/TFAM pathway | [157] |
| Asiatic acid | Triterpene | In vitro (HK-2 cells) In vivo (STZ-induced diabetic rats) | In vitro: 10 or 20 μm for 24 h In vivo: 10 or 30 mg/kg/day for 10 weeks | Modulate mitochondrial dynamics via NRF2 | [158] |
| Thymoquinone | Quinone | In vivo (STZ-induced diabetic rats) | 10 mg/kg/day for 8 weeks | Modulate NRF2/NOX2 pathway | [159] |
| 1, 2, 3, 4, 6-penta-O-galloyl-beta-D-glucose (PGG) | Tannin | In vitro (MPC-5 cells) In vivo (SFZ-induced diabetic rats) | In vitro: 20, 40, or 80 μm for 24 h In vivo: 5 or 20 mg/kg/day for 8 weeks | Suppress MAPK/NF-KB and ERK/NRF2/HO-1 | [160] |
| Fraxin | Coumarin | In vitro (rat GMCs) In vitro (db/db mice) | In vitro: 0, 20, 40, 60, 80, 160, or 320 μm for 24 h In vivo: 25, 50, or 100 6 days/week for 8 weeks | Modulate Cx43-AKT-NRF2/ARE pathway | [161] |
| Fucoxanthin | Carotenoid | In vitro (GMCs) In vivo (STZ-induced diabetic rats) | In vitro: 2 μm In vivo: 200 mg/kg/day for 12 weeks | ModulateSIRT1/NRF2/HO-1 pathway | [162] |
| Isoliquiritigenin | Flavonoid | In vitro (NRK-52E cells) In vivo (STZ-induced diabetic mice) | In vitro: 10 or 20 μm In vivo: 10 or 20 mg/kg every other day for 12 weeks | Activate NRF2 in a SIRT1-dependent manner | [163] |
| Baicalin | Flavonoid | In vivo (db/db mice) | 400 mg/kg/day for 8 weeks | Activate NRF2, inhibit MAPK-mediated inflammatory signaling pathway | [164] |
| Proanthocyanidins | Flavonoid | In vivo (high fat and sugar induced diabetic mice) | 5 mg/kg/day for 4 weeks | Activate p38 MAPK, KEAP1/NRF2 | [165] |
| Oligo-fucoidan | Oligosaccharides | In vitro (NRK-52E cells) In vivo (STZ-induced diabetic mice) | In vitro: 100 μg/mL for 24 h In vivo: 300 mg/kg/day for 8 weeks | Modulate SIRT-1/GLP-1R/NRF2-HO-1 pathway | [166] |
| Sulforaphane | Sulfur compound | In vivo (STZ-induced diabetic mice) In vitro (mouse podocyte cells) In vivo (STZ-induced diabetic podocyte specific Nrf2 KO mice) In vivo (STZ-induced diabetic mice) In vitro (RMCs) In vivo (STZ-induced diabetic rats) | In vivo: 3 months In vitro: 10 μm for 72 h In vivo: 6.5 or 12.5 mg/kg 3 times/week for 12 weeks 0.5 mg/kg 5 days/week for 4 months In vitro: 5 μm for 24 h In vivo: 5 mg/kg/day for 12 weeks | Activate AMPK-mediated lipid metabolic pathways, activate NRF2 via AMPK/AKT/GSK-3β/Fyn, modulate NRF2/PINK1 pathway | [45,167,168,169] |
| 4-O-methylhonokiol | Lignan | In vivo (STZ-induced diabetic mice) | In vivo: 1 mg/kg 5 days/week for 3 months | Activate AMPK/PGC-1a/CPT1B fatty acid oxidation, NRF2/SOF-2-mediated anti-oxidative stress | [170] |
| Moringa Isothiocyanate | Isothiocyanate | In vitro (HepG2-C8 and HK-2 cells) | 1.25, 2.5, or 5 μm for 24 h | Activate NRF2-ARE signaling, suppress inflammation and TGF-β1 signaling | [171] |
| Hesperetin | Glycoside | In vivo (STZ-induced diabetic rats) | 50 or 150 mg/kg/day for 68 days | Modulate NRF2/ARE/Glyoxalase 1 pathway | [172] |
| Paeonol | Ketone | In vitro (rat GMCs) In vivo (STZ-induced diabetic mice) | In vitro: 5, 10, or 20 μg/mL for 2 h In vivo: 150 mg/kg 6 times/week for 8 weeks | Modulate SIRT1/NRF2/ARE pathway | [173] |
| MG132 | Peptide | In vivo (STZ-induced diabetic mice) In vitro (HK11 cells) In vivo (OVE26 diabetic mice model) | 10 μg/kg/day for 4 months In vitro: 2 μm for 9 h In vivo: 10 μg/kg/day for 3 months | Inhibit proteasomal activity resulting in upregulation of NRF2 | [174,175] |
| Polydatin | Glucoside | In vitro (rat GMCs) In vivo (STZ-induced diabetic mice) | In vitro: 10, 20, or 40 μm for 2 h In vivo: 100 or 200 mg/kg/day for 8 weeks | Modulate CKIP-1/NRF2/ARE pathway | [176] |
| Sodium butyrate | Fatty acid | In vivo (STZ-induced diabetic mice) | 5 g/kg/day for 20 weeks | Inhibit histone deacetylase (HDAC) activity to elevate expression of NRF2, independent of KEAP1 and nuclear translocation | [177] |
| Sitagliptin | Pyrazine | In vitro (HK-2 cells) In vivo (Goto–Kakizaki rats) | In vitro: 0.05 or 0.1 μm for 1 h In vivo: 10 mg/kg/day for 20 weeks | Downregulate miR-200a/KEAP1/NRF2 pathway | [178] |
| AICAR | Purine nucleotide | In vitro (MCT cells) In vivo (db/db mice) | In vitro: 2 mm for 48 h In vivo: 2 mg/kg 5 days/week for 4 weeks | Activate AMPK, OGG1 and NRF2 expression | [179] |
| Gynostemma pentaphyllum saponins | Saponin | In vivo (STZ-induced diabetic rats) | 200 or 400 mg/kg/day for 40 days | Activate NRF2 pathway, increased SOD and GSH activity | [180] |
| Fenofibrate | Aryl carboxylic acid derivative | In vivo (STZ-induced diabetic mice) | 100 mg/kg every other day for 3 months | Upregulate FGF21, stimulating P13K/AKT/GSK-3β/Fyn-mediated NRF2 activation | [181] |
4.2. Alport Syndrome (AS)
4.3. Autosomal Dominant Polycystic Kidney Disease (ADPKD)
4.4. Lupus Nephritis (LN)
4.5. Adverse Aspects of NRF2 Activation
5. Targeting Fibrosis—The Key Mechanism Underlying CKD Progression
| Compound | Chemical Class | Study Design | Dosage and Duration of Treatment | Pathway/Mechanism | Reference |
|---|---|---|---|---|---|
| Geniposidic acid | Glucoside | In vitro (NRK-52E cells) In vivo (tubulointerstitial nephropathy rat model) | In vitro: 0, 1, 10, 20, 40, 80, 100, or 200 μm for 24 h In vivo: 20 mg/kg/day for 3 weeks | Repressing aryl hydrocarbon receptor (AHR), inhibit NF-kB, activate NRF2 | [221] |
| Betaine | Amine | In vivo (Doxorubicin-induced nephrotoxicity rat model) | 200 or 400 mg/kg for 28 days | Downregulate inflammatory and fibrotic gene expression | [222] |
| Catalpol | Glucoside | In vitro (NRK-52E cells) In vivo (aristolochic acid-induced kidney injury rat model) | In vitro: 5 or 10 μm for 24 h In vivo: 10 or 100 mg/kg/day for 29 days | Activate NRF2, inhibit NF-kB | [223] |
| Icariin | Flavonoid | In vitro (HK-2 cells) In vivo (UUO mice model) | In vitro: 50 μm for 24 h In vivo: 50 mg/kg/day for 14 days | Activate NRF2/HO-1 pathway, attenuate mitochondrial dysfunction, decrease profibrotic gene expression | [224] |
| Fasudil | Sulfur compound | In vitro (HK-2 cells) In vivo (hyperuricemic mice model) | In vitro: 50 μm for 48 h In vivo: 5 or 9 mg/kg/day for 5 weeks | Activate NRF2 via NEH2 domain, suppress EMT | [225] |
| Melatonin and Zileuton | Amine Amide | In vitro (HKC-8 and HK-2 cells) In vivo (UUO mice model) | In vitro: 1 mm melatonin and 5 μm zileuton In vivo: 20 mg/kg/day melatonin, 20 mg/kg/day zileuton or both for 1 week | Upregulate AKT/mTOR/NRF2 pathway | [226] |
| Spermidine | Amine | In vitro (HK-2 cells) In vivo (UUO mice model) | In vitro: 20 μm for 24 h In vivo: 10 mg/kg/day for 2 weeks | Activate NRF2 and suppress fibrotic signals | [227] |
| Bixin | Carotenoid | In vivo (carbon tetrachloride-induced renal injury mice model) In vitro (HK-2 cells) In vivo (UUO mice model) | 100 or 200 mg/kg/day for 4 weeks In vitro: 0, 20, or 40 μm for 4 or 24 h In vivo: 100 mg/kg once every 3 days for 7 days | Activate NRF2/TLR4/MyD88 pathway, suppress PPAR-gamma/TGF-β1/SMAD3 pathway Suppressing NRF2 ubiquitination, suppress EMT | [228,229] |
| Saponins from Panax japonicus | Saponin | In vivo (naturally aging rats) | 10 or 60 mg/kg/day for 3 or 6 months | Activate NRF2/ARE pathway, suppress NF-kB and TGF-β1/SMAD pathway | [230] |
| Dihydroquercetin | Flavonoid | In vitro (NRK-49F) In vivo (UUO mice model) | In vitro: 80 μm for 1 h In vivo: single dose of 50, 100, or 200 mg/kg | Activate NRF2 pathway, suppress TGF-β1/SMAD pathway | [231] |
| Gastrodin | Glucoside | In vivo (carbon tetrachloride-induced renal injury mice model) | 200 or 400 mg/kg/day for 4 weeks | Activate AMPK/NRF2/HMGB1 pathway, inhibit NF-kB and TGF-β1 pathways | [232] |
| Roxadustat | Isoquinoline | In vivo (folic acid-induced kidney injury mice model) | Single dose of 10 mg/kg | Activate AKT/GSK-3β/NRF2 pathway, decreasing ferroptosis | [233] |
| Bardoxolone methyl | Triterpene | In vitro (mouse GMCs) In vivo (aristolochic acid-induced injury mice model) | In vitro: 0.025, 0.05, or 0.1 μm for 24 h In vivo: 5 or 10 mg/kg/day for 16 days | Activate NRF2/SMAD7 pathway, downregulate TGF-β/SMAD/ECM expression | [63] |
| Epigallocatechin-3-gallate | Flavonoid | In vitro (MDCK cells) In vitro (NRK-52E cells) | 25 μm for 1 h 0, 1, 2, or 5 μm for 48 h | Activate NRF2/HO-1 pathway, protect against EMT and inflammation | [217,218,219] |
| Carnosic acid | Diterpene | In vitro (NKE cells) In vitro (cadmium-induced nephrotoxic mice model) | In vitro: 5 μm for 24 h In vivo: 10 mg/kg/day for 2 weeks | Activate NRF2/HO-1, Downregulate TGF-β1/SMAD/collagen IV signaling | [234] |
| Testosterone propionate | Steroids and steroid derivative | In vivo (naturally aging rats) | 2 mg/kg/day for 12 weeks | Activate NRF2/ARE signaling, suppress TGF-β1/SMAD signaling | [235] |
| Oltipraz | Sulfur compound | In vivo (UUO mice model) | 30 mg/kg/day for 14 days | Decrease expression of TGF-β1, E-cadherin | [236] |
| Sinomenine | Alkaloid | In vitro (HEK293 and RAW264.7 cells) In vivo (UUO mice model) | In vitro: 0, 25, 50, or 100 μm for 4 or 24 h In vivo: 100 mg/kg/day for 7 days | Activate NRF2 pathway, inhibit TGF-β/SMAD and WNT/β-catenin pathways | [65] |
| RTA dh404 | Triterpenoid | In vivo (5/6 nephrectomy mice model) | 2 mg/kg/day for 12 weeks | Activate NRF2 pathway, suppress NF-kB and TGF-β pathways | [237] |
| Dimethylfumarate | Carboxylic acid | In vitro (NRK-49F and RMCs) In vivo (UUO mice model) | In vitro: 0, 20, 40, or 80 µmol/l for 1 h In vivo: 25 mg/kg/day for 7 days | Activate NRF2 pathway, inhibit TGF-β/SMAD signaling | [62] |
6. Current Strategies, Challenges, and Future Perspectives
6.1. Current Therapeutic Strategies
6.2. Challenges
6.3. Future Perspectives
7. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ADPKD | Autosomal dominant polycystic kidney disease |
| a-SMA | Alpha-smooth muscle actin |
| ACACA | Acetyl-CoA carboxylase alpha |
| ACSL1 | Acyl-CoA synthetase long-chain family member 1 |
| AGT | Angiotensionogen |
| AMPK | AMP-activated protein kinase |
| AREs | Antioxidant response elements |
| AS | Alport syndrome |
| b-TrCP | Beta-transcducin repeat containing E3 ubiquitin protein ligase |
| CBP/p300 | CREB-binding protein |
| CKD | Chronic kidney disease |
| COX-2 | Cyclooxygenase-2 |
| CRIF1 | CR6-interacting factor 1 |
| Cul3-Rbx1 | Cullin3-ring box 1 |
| CuZn-SOD | Copper zinc superoxide dismutase |
| DKD | Diabetic kidney disease |
| EGCG | Epigallocatechin-3-gallate |
| eGFR | Estimated glomerular filtration rate |
| EMT | Epithelial–mesenchymal transition |
| FASN | Fatty acid synthase |
| GCS | Glutamylcysteine synthetase |
| GFR | Glomerular filtration rate |
| GSH | Gluthathion |
| GSK-3β | Glycogen synthase kinase-3β |
| GST | Glutathione S-transferase |
| HA | Hippuric acid |
| HO-1 | Heme oxygenase-1 |
| IL-1β | Interleukin 1 beta |
| IL-6 | Interleukin 6 |
| KEAP1 | Kelch-like ECH-associated protein 1 |
| LN | Lupus nephritis |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MRPL12 | Mitochondrial ribosomal protein L12 |
| NQO1 | NADPH-dehydrogenase quinone 1 |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| PBUT | Protein-bound uremic toxin |
| PPP | Pentose phosphate pathway |
| ROS | Reactive oxygen species |
| SCD1 | Stearoyl-CoA desturase 1 |
| SGLT2 | Sodium-glucose cotransporter 2 |
| SIAH2 | Seven in absentia homolog 2 |
| SLE | Systematic lupus erythematosus |
| sMAF | Small musculo-aponeutoric fibrosarcoma |
| TGF-β1 | Transforming growth factor beta 1 |
| TNF | Tumor necrosis factor |
| UUO | Unilateral ureteral obstruction |
| XRE | Xenobiotic response element |
References
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef]
- Ling, X.C.; Kuo, K.-L. Oxidative stress in chronic kidney disease. Ren. Replace. Ther. 2018, 4, 53. [Google Scholar] [CrossRef]
- Ho, H.J.; Shirakawa, H. Oxidative Stress and Mitochondrial Dysfunction in Chronic Kidney Disease. Cells 2022, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jose, P.A. Coordinated Contribution of NADPH Oxidase- and Mitochondria-Derived Reactive Oxygen Species in Metabolic Syndrome and Its Implication in Renal Dysfunction. Front. Pharmacol. 2021, 12, 670076. [Google Scholar] [CrossRef]
- Rispoli, R.M.; Popolo, A.; De Fabrizio, V.; d’Emmanuele di Villa Bianca, R.; Autore, G.; Dalli, J.; Marzocco, S. Targeting Inflammatory Imbalance in Chronic Kidney Disease: Focus on Anti-Inflammatory and Resolution Mediators. Int. J. Mol. Sci. 2025, 26, 3072. [Google Scholar] [CrossRef]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Pedraza-Chaverri, J.; Scholze, A. Nrf2 Activation in Chronic Kidney Disease: Promises and Pitfalls. Antioxidants 2022, 11, 1112. [Google Scholar] [CrossRef]
- Suzuki, T.; Seki, S.; Hiramoto, K.; Naganuma, E.; Kobayashi, E.H.; Yamaoka, A.; Baird, L.; Takahashi, N.; Sato, H.; Yamamoto, M. Hyperactivation of Nrf2 in early tubular development induces nephrogenic diabetes insipidus. Nat. Commun. 2017, 8, 14577. [Google Scholar] [CrossRef]
- Zhao, S.; Lo, C.S.; Miyata, K.N.; Ghosh, A.; Zhao, X.P.; Chenier, I.; Cailhier, J.F.; Ethier, J.; Lattouf, J.B.; Filep, J.G.; et al. Overexpression of Nrf2 in Renal Proximal Tubular Cells Stimulates Sodium-Glucose Cotransporter 2 Expression and Exacerbates Dysglycemia and Kidney Injury in Diabetic Mice. Diabetes 2021, 70, 1388–1403. [Google Scholar] [CrossRef]
- Su, K.; Zhao, S.L.; Yang, W.X.; Lo, C.S.; Chenier, I.; Liao, M.C.; Pang, Y.C.; Peng, J.Z.; Miyata, K.N.; Cailhier, J.F.; et al. NRF2 Deficiency Attenuates Diabetic Kidney Disease in Db/Db Mice via Down-Regulation of Angiotensinogen, SGLT2, CD36, and FABP4 Expression and Lipid Accumulation in Renal Proximal Tubular Cells. Antioxidants 2023, 12, 1715. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Hu, L.; Scrivens, P.J.; Batist, G. Transcriptional regulation of NF-E2 p45-related factor (NRF2) expression by the aryl hydrocarbon receptor-xenobiotic response element signaling pathway: Direct cross-talk between phase I and II drug-metabolizing enzymes. J. Biol. Chem. 2005, 280, 20340–20348. [Google Scholar] [CrossRef]
- Rushworth, S.A.; Zaitseva, L.; Murray, M.Y.; Shah, N.M.; Bowles, K.M.; MacEwan, D.J. The high Nrf2 expression in human acute myeloid leukemia is driven by NF-κB and underlies its chemo-resistance. Blood 2012, 120, 5188–5198. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.K.; Itoh, K.; Yamamoto, M.; Kensler, T.W. Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: Role of antioxidant response element-like sequences in the nrf2 promoter. Mol. Cell Biol. 2002, 22, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Wu, R.; Guo, Y.; Kong, A.N. Regulation of Keap1-Nrf2 signaling: The role of epigenetics. Curr. Opin. Toxicol. 2016, 1, 134–138. [Google Scholar] [CrossRef]
- Yu, S.; Khor, T.O.; Cheung, K.-L.; Li, W.; Wu, T.-Y.; Huang, Y.; Foster, B.A.; Kan, Y.W.; Kong, A.-N. Nrf2 Expression Is Regulated by Epigenetic Mechanisms in Prostate Cancer of TRAMP Mice. PLoS ONE 2010, 5, e8579. [Google Scholar] [CrossRef]
- Yang, P.L.; Lai, T.S.; Chou, Y.H.; Lai, L.C.; Lin, S.L.; Chen, Y.M. DNA methylation in peripheral blood is associated with renal aging and renal function decline: A national community study. Clin. Epigenet. 2024, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Palsamy, P.; Ayaki, M.; Elanchezhian, R.; Shinohara, T. Promoter demethylation of Keap1 gene in human diabetic cataractous lenses. Biochem. Biophys. Res. Commun. 2012, 423, 542–548. [Google Scholar] [CrossRef]
- Liu, Z.Z.; Zhao, X.Z.; Zhang, X.S.; Zhang, M. Promoter DNA demethylation of Keap1 gene in diabetic cardiomyopathy. Int. J. Clin. Exp. Pathol. 2014, 7, 8756–8762. [Google Scholar]
- Sun, Z.; Chin, Y.E.; Zhang, D.D. Acetylation of Nrf2 by p300/CBP augments promoter-specific DNA binding of Nrf2 during the antioxidant response. Mol. Cell Biol. 2009, 29, 2658–2672. [Google Scholar] [CrossRef]
- Harithpriya, K.; Juttada, U.; Jayasuriya, R.; Kumpatla, S.; Viswanathan, V.; Ramkumar, K.M. Comprehensive gene expression analysis of histone deacetylases and the transcription factor Nrf2 in the progression of diabetic nephropathy. Int. J. Diabetes Dev. Ctries. 2025, 45, 462–471. [Google Scholar] [CrossRef]
- He, R.; Liu, B.; Geng, B.; Li, N.; Geng, Q. The role of HDAC3 and its inhibitors in regulation of oxidative stress and chronic diseases. Cell Death Discov. 2023, 9, 131. [Google Scholar] [CrossRef]
- Kim, S.K.; Bae, G.S.; Bae, T.; Ku, S.K.; Choi, B.H.; Kwak, M.K. Renal microRNA-144-3p is associated with transforming growth factor-β1-induced oxidative stress and fibrosis by suppressing the NRF2 pathway in hypertensive diabetic kidney disease. Free Radic. Biol. Med. 2024, 225, 546–559. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, W.; Li, W.; Yu, W.; Li, S.; Rao, T.; Ruan, Y.; Zhou, X.; Liu, C.; Qi, Y.; et al. LncRNA TUG1 attenuates ischaemia-reperfusion-induced apoptosis of renal tubular epithelial cells by sponging miR-144-3p via targeting Nrf2. J. Cell Mol. Med. 2021, 25, 9767–9783. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: Oxidative stress sensing by a Cul3-Keap1 ligase. Mol. Cell Biol. 2004, 24, 8477–8486. [Google Scholar] [CrossRef]
- Lo, S.C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Jaiswal, A.K. An autoregulatory loop between Nrf2 and Cul3-Rbx1 controls their cellular abundance. J. Biol. Chem. 2010, 285, 21349–21358. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Song, Y.; Qu, Y.; Mao, C.; Zhang, R.; Jiang, D.; Sun, X. Post-translational modifications of Keap1: The state of the art. Front. Cell Dev. Biol. 2024, 11, 1332049. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef]
- Li, W.; Yu, S.; Liu, T.; Kim, J.H.; Blank, V.; Li, H.; Kong, A.N. Heterodimerization with small Maf proteins enhances nuclear retention of Nrf2 via masking the NESzip motif. Biochim. Biophys. Acta 2008, 1783, 1847–1856. [Google Scholar] [CrossRef]
- Otsuki, A.; Yamamoto, M. Cis-element architecture of Nrf2–sMaf heterodimer binding sites and its relation to diseases. Arch. Pharmacal Res. 2020, 43, 275–285. [Google Scholar] [CrossRef]
- Wasserman, W.W.; Fahl, W.E. Functional antioxidant responsive elements. Proc. Natl. Acad. Sci. USA 1997, 94, 5361–5366. [Google Scholar] [CrossRef]
- Shelton, L.M.; Lister, A.; Walsh, J.; Jenkins, R.E.; Wong, M.H.; Rowe, C.; Ricci, E.; Ressel, L.; Fang, Y.; Demougin, P.; et al. Integrated transcriptomic and proteomic analyses uncover regulatory roles of Nrf2 in the kidney. Kidney Int. 2015, 88, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Rojo, A.I.; Chowdhry, S.; McMahon, M.; Hayes, J.D.; Cuadrado, A. SCF/β-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner. Mol. Cell Biol. 2011, 31, 1121–1133. [Google Scholar] [CrossRef]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct β-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef]
- Mo, C.; Wang, L.; Zhang, J.; Numazawa, S.; Tang, H.; Tang, X.; Han, X.; Li, J.; Yang, M.; Wang, Z.; et al. The crosstalk between Nrf2 and AMPK signal pathways is important for the anti-inflammatory effect of berberine in LPS-stimulated macrophages and endotoxin-shocked mice. Antioxid. Redox Signal 2014, 20, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhao, T.; Xiao, H. The Implication of Oxidative Stress and AMPK-Nrf2 Antioxidative Signaling in Pneumonia Pathogenesis. Front. Endocrinol. 2020, 11, 400. [Google Scholar] [CrossRef]
- Wang, X.; Tan, X.; Zhang, J.; Wu, J.; Shi, H. The emerging roles of MAPK-AMPK in ferroptosis regulatory network. Cell Commun. Signal. 2023, 21, 200. [Google Scholar] [CrossRef]
- Kang, H.J.; Hong, Y.B.; Kim, H.J.; Bae, I. CR6-interacting factor 1 (CRIF1) regulates NF-E2-related factor 2 (NRF2) protein stability by proteasome-mediated degradation. J. Biol. Chem. 2010, 285, 21258–21268. [Google Scholar] [CrossRef]
- Baba, K.; Morimoto, H.; Imaoka, S. Seven in absentia homolog 2 (Siah2) protein is a regulator of NF-E2-related factor 2 (Nrf2). J. Biol. Chem. 2013, 288, 18393–18405. [Google Scholar] [CrossRef]
- Liu, Y.; Uruno, A.; Saito, R.; Matsukawa, N.; Hishinuma, E.; Saigusa, D.; Liu, H.; Yamamoto, M. Nrf2 deficiency deteriorates diabetic kidney disease in Akita model mice. Redox Biol. 2022, 58, 102525. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ming, H.; Qin, S.; Nice, E.C.; Dong, J.; Du, Z.; Huang, C. Redox regulation: Mechanisms, biology and therapeutic targets in diseases. Signal Transduct. Target. Ther. 2025, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Fu, J.; Liu, N.; Jiao, C.; Guo, G.; Luan, J.; Wang, H.; Yao, L.; Wang, L.; Yamamoto, M.; et al. Nrf2 deficiency promotes the progression from acute tubular damage to chronic renal fibrosis following unilateral ureteral obstruction. Nephrol. Dial. Transplant. 2018, 33, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, H.; Li, J.; Ma, T.; Zhou, S.; Zhang, Z.; Miao, L.; Cai, L. Sulforaphane prevents type 2 diabetes-induced nephropathy via AMPK-mediated activation of lipid metabolic pathways and Nrf2 antioxidative function. Clin. Sci. 2020, 134, 2469–2487. [Google Scholar] [CrossRef]
- Ribeiro, M.; Alvarenga, L.; Coutinho-Wolino, K.S.; Nakao, L.S.; Cardozo, L.F.M.F.; Mafra, D. Sulforaphane upregulates the mRNA expression of NRF2 and NQO1 in non-dialysis patients with chronic kidney disease. Free Radic. Biol. Med. 2024, 221, 181–187. [Google Scholar] [CrossRef]
- Rasmussen, M.; Hansen, K.H.; Scholze, A. Nrf2 Protein Serum Concentration in Human CKD Shows a Biphasic Behavior. Antioxidants 2023, 12, 932. [Google Scholar] [CrossRef]
- Zager, R.A.; Johnson, A.C.M.; Guillem, A.; Keyser, J.; Singh, B. A Pharmacologic “Stress Test” for Assessing Select Antioxidant Defenses in Patients with CKD. Clin. J. Am. Soc. Nephrol. 2020, 15, 633–642. [Google Scholar] [CrossRef]
- Declèves, A.-E.; Sharma, K. Novel targets of antifibrotic and anti-inflammatory treatment in CKD. Nat. Rev. Nephrol. 2014, 10, 257–267. [Google Scholar] [CrossRef]
- Kok, H.M.; Falke, L.L.; Goldschmeding, R.; Nguyen, T.Q. Targeting CTGF, EGF and PDGF pathways to prevent progression of kidney disease. Nat. Rev. Nephrol. 2014, 10, 700–711. [Google Scholar] [CrossRef]
- Ziady, A.G.; Sokolow, A.; Shank, S.; Corey, D.; Myers, R.; Plafker, S.; Kelley, T.J. Interaction with CREB binding protein modulates the activities of Nrf2 and NF-κB in cystic fibrosis airway epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, L1221–L1231. [Google Scholar] [CrossRef]
- Bellezza, I.; Tucci, A.; Galli, F.; Grottelli, S.; Mierla, A.L.; Pilolli, F.; Minelli, A. Inhibition of NF-κB nuclear translocation via HO-1 activation underlies α-tocopheryl succinate toxicity. J. Nutr. Biochem. 2012, 23, 1583–1591. [Google Scholar] [CrossRef]
- Nallasamy, P.; Si, H.; Babu, P.V.; Pan, D.; Fu, Y.; Brooke, E.A.; Shah, H.; Zhen, W.; Zhu, H.; Liu, D.; et al. Sulforaphane reduces vascular inflammation in mice and prevents TNF-α-induced monocyte adhesion to primary endothelial cells through interfering with the NF-κB pathway. J. Nutr. Biochem. 2014, 25, 824–833. [Google Scholar] [CrossRef]
- Kim, H.J.; Vaziri, N.D. Contribution of impaired Nrf2-Keap1 pathway to oxidative stress and inflammation in chronic renal failure. Am. J. Physiol.-Ren. Physiol. 2010, 298, F662–F671. [Google Scholar] [CrossRef]
- Lau, W.L.; Liu, S.M.; Pahlevan, S.; Yuan, J.; Khazaeli, M.; Ni, Z.; Chan, J.Y.; Vaziri, N.D. Role of Nrf2 dysfunction in uremia-associated intestinal inflammation and epithelial barrier disruption. Dig. Dis. Sci. 2015, 60, 1215–1222. [Google Scholar] [CrossRef]
- Tan, R.J.; Chartoumpekis, D.V.; Rush, B.M.; Zhou, D.; Fu, H.; Kensler, T.W.; Liu, Y. Keap1 hypomorphism protects against ischemic and obstructive kidney disease. Sci. Rep. 2016, 6, 36185. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Fu, P.; Ma, L. Kidney fibrosis: From mechanisms to therapeutic medicines. Signal Transduct. Target. Ther. 2023, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.P.; Williams, C.E.; Lemmon, C.A. Cell-Matrix Interactions in Renal Fibrosis. Kidney Dial. 2022, 2, 607–624. [Google Scholar] [CrossRef]
- Lv, W.; Booz, G.W.; Fan, F.; Wang, Y.; Roman, R.J. Oxidative Stress and Renal Fibrosis: Recent Insights for the Development of Novel Therapeutic Strategies. Front. Physiol. 2018, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, H.; Huang, L.; Zhu, X.; Sha, J.; Li, G.; Ma, G.; Zhang, W.; Gu, M.; Guo, Y. Nrf2 signaling attenuates epithelial-to-mesenchymal transition and renal interstitial fibrosis via PI3K/Akt signaling pathways. Exp. Mol. Pathol. 2019, 111, 104296. [Google Scholar] [CrossRef]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: From mechanisms to medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.J.; Kim, J.Y.; Choi, Y.K.; Kim, H.J.; Jeong, J.Y.; Bae, K.H.; Park, K.G.; Lee, I.K. Dimethylfumarate attenuates renal fibrosis via NF-E2-related factor 2-mediated inhibition of transforming growth factor-beta/Smad signaling. PLoS ONE 2012, 7, e45870. [Google Scholar] [CrossRef] [PubMed]
- Song, M.K.; Lee, J.H.; Ryoo, I.G.; Lee, S.H.; Ku, S.K.; Kwak, M.K. Bardoxolone ameliorates TGF-beta1-associated renal fibrosis through Nrf2/Smad7 elevation. Free Radic. Biol. Med. 2019, 138, 33–42. [Google Scholar] [CrossRef]
- Nezu, M.; Souma, T.; Yu, L.; Suzuki, T.; Saigusa, D.; Ito, S.; Suzuki, N.; Yamamoto, M. Transcription factor Nrf2 hyperactivation in early-phase renal ischemia-reperfusion injury prevents tubular damage progression. Kidney Int. 2017, 91, 387–401. [Google Scholar] [CrossRef]
- Qin, T.; Yin, S.; Yang, J.; Zhang, Q.; Liu, Y.; Huang, F.; Cao, W. Sinomenine attenuates renal fibrosis through Nrf2-mediated inhibition of oxidative stress and TGFbeta signaling. Toxicol. Appl. Pharmacol. 2016, 304, 1–8. [Google Scholar] [CrossRef]
- Sun, B.; Wang, X.; Liu, X.; Wang, L.; Ren, F.; Wang, X.; Leng, X. Hippuric Acid Promotes Renal Fibrosis by Disrupting Redox Homeostasis via Facilitation of NRF2-KEAP1-CUL3 Interactions in Chronic Kidney Disease. Antioxidants 2020, 9, 783. [Google Scholar] [CrossRef]
- Fan, X.; Yang, M.; Lang, Y.; Lu, S.; Kong, Z.; Gao, Y.; Shen, N.; Zhang, D.; Lv, Z. Mitochondrial metabolic reprogramming in diabetic kidney disease. Cell Death Dis. 2024, 15, 442. [Google Scholar] [CrossRef]
- Rinaldi, A.; Lazareth, H.; Poindessous, V.; Nemazanyy, I.; Sampaio, J.L.; Malpetti, D.; Bignon, Y.; Naesens, M.; Rabant, M.; Anglicheau, D.; et al. Impaired fatty acid metabolism perpetuates lipotoxicity along the transition to chronic kidney injury. JCI Insight 2022, 7, e161783. [Google Scholar] [CrossRef]
- Yates, M.S.; Tran, Q.T.; Dolan, P.M.; Osburn, W.O.; Shin, S.; McCulloch, C.C.; Silkworth, J.B.; Taguchi, K.; Yamamoto, M.; Williams, C.R.; et al. Genetic versus chemoprotective activation of Nrf2 signaling: Overlapping yet distinct gene expression profiles between Keap1 knockout and triterpenoid-treated mice. Carcinogenesis 2009, 30, 1024–1031. [Google Scholar] [CrossRef]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Ludtmann, M.H.; Angelova, P.R.; Zhang, Y.; Abramov, A.Y.; Dinkova-Kostova, A.T. Nrf2 affects the efficiency of mitochondrial fatty acid oxidation. Biochem. J. 2014, 457, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; He, L.; Yang, Y.; Chen, Y.; Song, Y.; Lu, X.; Liang, Y. The inhibition of Nrf2 accelerates renal lipid deposition through suppressing the ACSL1 expression in obesity-related nephropathy. Ren. Fail. 2019, 41, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H.; Schachner, D.; Zimmermann, K.; Dirsch, V.M. Glucose availability is a decisive factor for Nrf2-mediated gene expression. Redox Biol. 2013, 1, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Zoccarato, A.; Smyrnias, I.; Reumiller, C.M.; Hafstad, A.D.; Chong, M.; Richards, D.A.; Santos, C.X.C.; Visnagri, A.; Verma, S.; Bromage, D.I.; et al. NRF2 activation in the heart induces glucose metabolic reprogramming and reduces cardiac dysfunction via upregulation of the pentose phosphate pathway. Cardiovasc. Res. 2024, 121, 339–352. [Google Scholar] [CrossRef]
- Tan, V.W.T.; Salmi, T.M.; Karamalakis, A.P.; Gillespie, A.; Ong, A.J.S.; Balic, J.J.; Chan, Y.C.; Bladen, C.E.; Brown, K.K.; Dawson, M.A.; et al. SLAM-ITseq identifies that Nrf2 induces liver regeneration through the pentose phosphate pathway. Dev. Cell 2024, 59, 898–910.e896. [Google Scholar] [CrossRef]
- Spoto, B.; Politi, C.; Postorino, M.; Parlongo, R.M.; Testa, A.; Tripepi, G.L.; Mallamaci, F.; Zoccali, C. Role of kidney function on Nrf2 mRNA levels in type 2 diabetes. BMJ Open Diabetes Res. Care 2024, 12, e003929. [Google Scholar] [CrossRef]
- Jiang, X.S.; Liu, T.; Xia, Y.F.; Gan, H.; Ren, W.; Du, X.G. Activation of the Nrf2/ARE signaling pathway ameliorates hyperlipidemia-induced renal tubular epithelial cell injury by inhibiting mtROS-mediated NLRP3 inflammasome activation. Front. Immunol. 2024, 15, 1342350. [Google Scholar] [CrossRef]
- Gu, X.; Liu, Y.; Wang, N.; Zhen, J.; Zhang, B.; Hou, S.; Cui, Z.; Wan, Q.; Feng, H. Transcription of MRPL12 regulated by Nrf2 contributes to the mitochondrial dysfunction in diabetic kidney disease. Free Radic. Biol. Med. 2021, 164, 329–340. [Google Scholar] [CrossRef]
- Podrini, C.; Cassina, L.; Boletta, A. Metabolic reprogramming and the role of mitochondria in polycystic kidney disease. Cell Signal 2020, 67, 109495. [Google Scholar] [CrossRef]
- Song, X.; Pickel, L.; Sung, H.K.; Scholey, J.; Pei, Y. Reprogramming of Energy Metabolism in Human PKD1 Polycystic Kidney Disease: A Systems Biology Analysis. Int. J. Mol. Sci. 2024, 25, 7173. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Chen, H.; Chen, D.Q.; Vaziri, N.D.; Su, W.; Ma, S.X.; Shang, Y.Q.; Mao, J.R.; Yu, X.Y.; Zhang, L.; et al. Activated NF-kappaB/Nrf2 and Wnt/beta-catenin pathways are associated with lipid metabolism in CKD patients with microalbuminuria and macroalbuminuria. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2317–2332. [Google Scholar] [CrossRef] [PubMed]
- Juul-Nielsen, C.; Shen, J.; Stenvinkel, P.; Scholze, A. Systematic review of the nuclear factor erythroid 2-related factor 2 (NRF2) system in human chronic kidney disease: Alterations, interventions and relation to morbidity. Nephrol. Dial. Transplant. 2022, 37, 904–916. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, Y.; Liu, Z.; Lu, Y.; Zhu, X.; Lan, B.; Mi, Z.; Dang, L.; Li, N.; Zhan, W.; et al. Activation of NRF2 ameliorates oxidative stress and cystogenesis in autosomal dominant polycystic kidney disease. Sci. Transl. Med. 2020, 12, eaba3613. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, C. Oxidative Stress: A Culprit in the Progression of Diabetic Kidney Disease. Antioxidants 2024, 13, 455. [Google Scholar] [CrossRef]
- Jiang, T.; Tian, F.; Zheng, H.; Whitman, S.A.; Lin, Y.; Zhang, Z.; Zhang, N.; Zhang, D.D. Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-κB-mediated inflammatory response. Kidney Int. 2014, 85, 333–343. [Google Scholar] [CrossRef]
- Rush, B.M.; Bondi, C.D.; Stocker, S.D.; Barry, K.M.; Small, S.A.; Ong, J.; Jobbagy, S.; Stolz, D.B.; Bastacky, S.I.; Chartoumpekis, D.V.; et al. Genetic or pharmacologic Nrf2 activation increases proteinuria in chronic kidney disease in mice. Kidney Int. 2021, 99, 102–116. [Google Scholar] [CrossRef]
- Ng, D.; Hartman, H.L.; Yeh, H.; Gilbert, J.; Stewart, J.A.; Bondi, C.D.; Miner, J.H.; Tan, R.J. Genetic NRF2 Activation Worsens Kidney Injury in an Alport Syndrome Mouse Model. J. Am. Soc. Nephrol. 2024, 35, 686. [Google Scholar] [CrossRef]
- Nie, P.; Lou, Y.; Bai, X.; Zhu, Y.; Guo, Q.; Luo, P.; Zhang, W.; Li, B. Influence of zinc levels and Nrf2 expression in the clinical and pathological changes in patients with diabetic nephropathy. Nutr. Diabetes 2022, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Nakai, K.; Fujii, H.; Kono, K.; Goto, S.; Kitazawa, R.; Kitazawa, S.; Hirata, M.; Shinohara, M.; Fukagawa, M.; Nishi, S. Vitamin D Activates the Nrf2-Keap1 Antioxidant Pathway and Ameliorates Nephropathy in Diabetic Rats. Am. J. Hypertens. 2013, 27, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Mohan, T.; Narasimhan, K.K.S.; Ravi, D.B.; Velusamy, P.; Chandrasekar, N.; Chakrapani, L.N.; Srinivasan, A.; Karthikeyan, P.; Kannan, P.; Tamilarasan, B.; et al. Role of Nrf2 dysfunction in the pathogenesis of diabetic nephropathy: Therapeutic prospect of epigallocatechin-3-gallate. Free Radic. Biol. Med. 2020, 160, 227–238. [Google Scholar] [CrossRef]
- Sun, W.; Liu, X.; Zhang, H.; Song, Y.; Li, T.; Liu, X.; Liu, Y.; Guo, L.; Wang, F.; Yang, T.; et al. Epigallocatechin gallate upregulates NRF2 to prevent diabetic nephropathy via disabling KEAP1. Free Radic. Biol. Med. 2017, 108, 840–857. [Google Scholar] [CrossRef]
- Li, S.; Zheng, L.; Zhang, J.; Liu, X.; Wu, Z. Inhibition of ferroptosis by up-regulating Nrf2 delayed the progression of diabetic nephropathy. Free Radic. Biol. Med. 2021, 162, 435–449. [Google Scholar] [CrossRef]
- Pergola, P.E.; Raskin, P.; Toto, R.D.; Meyer, C.J.; Huff, J.W.; Grossman, E.B.; Krauth, M.; Ruiz, S.; Audhya, P.; Christ-Schmidt, H.; et al. Bardoxolone methyl and kidney function in CKD with type 2 diabetes. N. Engl. J. Med. 2011, 365, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.P.; Wrolstad, D.; Bakris, G.L.; Chertow, G.M.; de Zeeuw, D.; Goldsberry, A.; Linde, P.G.; McCullough, P.A.; McMurray, J.J.; Wittes, J.; et al. Risk factors for heart failure in patients with type 2 diabetes mellitus and stage 4 chronic kidney disease treated with bardoxolone methyl. J. Card. Fail. 2014, 20, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Dare, A.; Channa, M.L.; Nadar, A. L-ergothioneine and its combination with metformin attenuates renal dysfunction in type-2 diabetic rat model by activating Nrf2 antioxidant pathway. Biomed. Pharmacother. 2021, 141, 111921. [Google Scholar] [CrossRef]
- Wu, P.; Yan, Y.; Ma, L.L.; Hou, B.Y.; He, Y.Y.; Zhang, L.; Niu, Z.R.; Song, J.K.; Pang, X.C.; Yang, X.Y.; et al. Effects of the Nrf2 Protein Modulator Salvianolic Acid A Alone or Combined with Metformin on Diabetes-associated Macrovascular and Renal Injury. J. Biol. Chem. 2016, 291, 22288–22301. [Google Scholar] [CrossRef]
- Sleem, M.; Aboubakr, E.M.; Mohamed, W.R.; Khalil, A.M.; Messiha, B.A.S.; Taye, A. Potential roles for eNOS and NrF(2) /HO-1 signaling in the ameliorative effect of lixisenatide on diabetes-induced kidney injury in rats and its amplification by ticagrelor co-administration. Pharmazie 2024, 79, 187–194. [Google Scholar] [CrossRef]
- Antar, S.A.; Abdo, W.; Taha, R.S.; Farage, A.E.; El-Moselhy, L.E.; Amer, M.E.; Abdel Monsef, A.S.; Abdel Hamid, A.M.; Kamel, E.M.; Ahmeda, A.F.; et al. Telmisartan attenuates diabetic nephropathy by mitigating oxidative stress and inflammation, and upregulating Nrf2/HO-1 signaling in diabetic rats. Life Sci. 2022, 291, 120260. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, G.; Cheng, X.; Teng, Z.; Cai, X.; Yang, J.; Sun, X.; Lu, W.; Wang, X.; Yao, Y.; et al. Therapeutic potential of digitoflavone on diabetic nephropathy: Nuclear factor erythroid 2-related factor 2-dependent anti-oxidant and anti-inflammatory effect. Sci. Rep. 2015, 5, 12377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, X.; Zhang, C.; Shen, Q.; Sun, G.; Sun, X. Notoginsenoside R1 Protects db/db Mice against Diabetic Nephropathy via Upregulation of Nrf2-Mediated HO-1 Expression. Molecules 2019, 24, 247. [Google Scholar] [CrossRef]
- Alaofi, A.L. Sinapic Acid Ameliorates the Progression of Streptozotocin (STZ)-Induced Diabetic Nephropathy in Rats via NRF2/HO-1 Mediated Pathways. Front. Pharmacol. 2020, 11, 1119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zeng, J.; Li, J.; Chen, X.; Miao, J.; Jin, Q.; Chen, H. Effects of Compound Centella on Oxidative Stress and Keap1-Nrf2-ARE Pathway Expression in Diabetic Kidney Disease Rats. Evid. Based Complement. Altern. Med. 2020, 2020, 9817932. [Google Scholar] [CrossRef]
- Shopit, A.; Niu, M.; Wang, H.; Tang, Z.; Li, X.; Tesfaldet, T.; Ai, J.; Ahmad, N.; Al-Azab, M.; Tang, Z. Protection of diabetes-induced kidney injury by phosphocreatine via the regulation of ERK/Nrf2/HO-1 signaling pathway. Life Sci. 2020, 242, 117248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qi, S.; Song, Y.; Ling, C. Artemisinin attenuates early renal damage on diabetic nephropathy rats through suppressing TGF-beta1 regulator and activating the Nrf2 signaling pathway. Life Sci. 2020, 256, 117966. [Google Scholar] [CrossRef]
- Zhai, J.; Li, Z.; Zhang, H.; Ma, L.; Ma, Z.; Zhang, Y.; Zou, J.; Li, M.; Ma, L.; Li, X. Coptisine ameliorates renal injury in diabetic rats through the activation of Nrf2 signaling pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 57–65. [Google Scholar] [CrossRef]
- Kaikini, A.A.; Muke, S.; Peshattiwar, V.; Bagle, S.; Dighe, V.; Sathaye, S. Ethyl ferulate, a lipophilic phenylpropanoid, prevents diabetes-associated renal injury in rats by amelioration of hyperglycemia-induced oxidative stress via activation of nuclear factor erythroid 2-related factor 2. J. Food Biochem. 2021, 45, e13607. [Google Scholar] [CrossRef]
- Huang, Q.; Ouyang, D.S.; Liu, Q. Isoeucommin A attenuates kidney injury in diabetic nephropathy through the Nrf2/HO-1 pathway. FEBS Open Bio 2021, 11, 2350–2363. [Google Scholar] [CrossRef]
- Alshehri, A.S. Kaempferol attenuates diabetic nephropathy in streptozotocin-induced diabetic rats by a hypoglycaemic effect and concomitant activation of the Nrf-2/Ho-1/antioxidants axis. Arch. Physiol. Biochem. 2023, 129, 984–997. [Google Scholar] [CrossRef]
- Ke, J.; Zang, H.; Liu, Y.; Teng, Q.; Hua, J.; Peng, D.; Wang, P. beta-cryptoxanthin suppresses oxidative stress via activation of the Nrf2/HO-1 signaling pathway in diabetic kidney disease. Front. Pharmacol. 2024, 15, 1480629. [Google Scholar] [CrossRef]
- Hasan, I.H.; Shaheen, S.Y.; Alhusaini, A.M.; Mahmoud, A.M. Simvastatin mitigates diabetic nephropathy by upregulating farnesoid X receptor and Nrf2/HO-1 signaling and attenuating oxidative stress and inflammation in rats. Life Sci. 2024, 340, 122445. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Leng, Y.F.; Wang, W.; Zhang, J.; Yuan, T.L.; Wang, J. Protective effect of Saxagliptin on diabetic rats with renal ischemia reperfusion injury by targeting oxidative stress and mitochondrial apoptosis pathway through activating Nrf-2/HO-1 signaling. Transpl. Immunol. 2023, 76, 101762. [Google Scholar] [CrossRef]
- Guo, J.; Li, J.; Wei, H.; Liang, Z. Maackiain Protects the Kidneys of Type 2 Diabetic Rats via Modulating the Nrf2/HO-1 and TLR4/NF-kappaB/Caspase-3 Pathways. Drug Des. Devel Ther. 2021, 15, 4339–4358. [Google Scholar] [CrossRef]
- Duan, Q.; Tian, L.; Feng, J.; Ping, X.; Li, L.; Yaigoub, H.; Li, R.; Li, Y. Trametenolic Acid Ameliorates the Progression of Diabetic Nephropathy in db/db Mice via Nrf2/HO-1 and NF-kappaB-Mediated Pathways. J. Immunol. Res. 2022, 2022, 6151847. [Google Scholar] [CrossRef]
- Lu, C.; Fan, G.; Wang, D. Akebia Saponin D ameliorated kidney injury and exerted anti-inflammatory and anti-apoptotic effects in diabetic nephropathy by activation of NRF2/HO-1 and inhibition of NF-KB pathway. Int. Immunopharmacol. 2020, 84, 106467. [Google Scholar] [CrossRef]
- Xie, Z.; Zhong, L.; Wu, Y.; Wan, X.; Yang, H.; Xu, X.; Li, P. Carnosic acid improves diabetic nephropathy by activating Nrf2/ARE and inhibition of NF-kappaB pathway. Phytomedicine 2018, 47, 161–173. [Google Scholar] [CrossRef]
- Al-Hussan, R.; Albadr, N.A.; Alshammari, G.M.; Almasri, S.A.; Alfayez, F.F.; Yahya, M.A. Phloretamide Protects against Diabetic Kidney Damage and Dysfunction in Diabetic Rats by Attenuating Hyperglycemia and Hyperlipidemia, Suppressing NF-kappabeta, and Upregulating Nrf2. Pharmaceutics 2024, 16, 505. [Google Scholar] [CrossRef] [PubMed]
- Noei Razliqi, R.; Ahangarpour, A.; Mard, S.A.; Khorsandi, L. Gentisic acid protects against diabetic nephropathy in Nicotinamide-Streptozotocin administered male mice by attenuating oxidative stress and inflammation: The role of miR-200a/Keap1/Nrf2 pathway, renin-angiotensin system (RAS) and NF-small ka, CyrillicB. Chem. Biol. Interact. 2023, 380, 110507. [Google Scholar] [CrossRef]
- AlTamimi, J.Z.; AlFaris, N.A.; Alshammari, G.M.; Alagal, R.I.; Aljabryn, D.H.; Abdo Yahya, M. Protective effect of eriodictyol against hyperglycemia-induced diabetic nephropathy in rats entails antioxidant and anti-inflammatory effects mediated by activating Nrf2. Saudi Pharm. J. 2023, 31, 101817. [Google Scholar] [CrossRef] [PubMed]
- Palsamy, P.; Subramanian, S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling. Biochim. Biophys. Acta 2011, 1812, 719–731. [Google Scholar] [CrossRef]
- Du, L.; Wang, L.; Wang, B.; Wang, J.; Hao, M.; Chen, Y.B.; Li, X.Z.; Li, Y.; Jiang, Y.F.; Li, C.C.; et al. A novel compound AB38b attenuates oxidative stress and ECM protein accumulation in kidneys of diabetic mice through modulation of Keap1/Nrf2 signaling. Acta Pharmacol. Sin. 2020, 41, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, S.; Jin, G.; Gao, K.; Wang, S.; Zhang, X.; Zhou, K.; Cai, Y.; Zhou, X.; Zhao, Z. Network pharmacology-based study on the mechanism of ShenKang injection in diabetic kidney disease through Keap1/Nrf2/Ho-1 signaling pathway. Phytomedicine 2023, 118, 154915. [Google Scholar] [CrossRef]
- Wang, K.; Zheng, X.; Pan, Z.; Yao, W.; Gao, X.; Wang, X.; Ding, X. Icariin Prevents Extracellular Matrix Accumulation and Ameliorates Experimental Diabetic Kidney Disease by Inhibiting Oxidative Stress via GPER Mediated p62-Dependent Keap1 Degradation and Nrf2 Activation. Front. Cell Dev. Biol. 2020, 8, 559. [Google Scholar] [CrossRef]
- Yuan, X.; Tang, W.; Lin, C.; He, H.; Li, L. Chrysophanol ameliorates oxidative stress and pyroptosis in mice with diabetic nephropathy through the Kelch-like ECH-associated protein 1/nuclear factor erythroid 2-related factor 2 signaling pathway. Acta Biochim. Pol. 2023, 70, 891–897. [Google Scholar] [CrossRef]
- Zhang, R.; Lu, M.; Zhang, S.; Liu, J. Renoprotective effects of Tilianin in diabetic rats through modulation of oxidative stress via Nrf2-Keap1 pathway and inflammation via TLR4/MAPK/NF-kappaB pathways. Int. Immunopharmacol. 2020, 88, 106967. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, W.; Yang, F.; Ai, F.; Du, D.; Li, Y. Discovery of caffeoylisocitric acid as a Keap1-dependent Nrf2 activator and its effects in mesangial cells under high glucose. J. Enzyme Inhib. Med. Chem. 2022, 37, 178–188. [Google Scholar] [CrossRef]
- Pena-Montes, D.J.; Huerta-Cervantes, M.; Riveros-Rosas, H.; Manzo-Avalos, S.; Aguilera-Mendez, A.; Huerta, M.; Trujillo, X.; Cortes-Rojo, C.; Montoya-Perez, R.; Salgado-Garciglia, R.; et al. Iron chelation mitigates mitochondrial dysfunction and oxidative stress by enhancing nrf2-mediated antioxidant responses in the renal cortex of a murine model of type 2 diabetes. Mitochondrion 2024, 78, 101937. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Zhang, Y.; Wei, Q.; Huang, Z. GLP-1 receptor agonist liraglutide alleviates kidney injury by regulating nuclear translocation of NRF2 in diabetic nephropathy. Clin. Exp. Pharmacol. Physiol. 2024, 51, e70003. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, C.Y.; Chen, H.; Liu, J.; Zhong, L.L.; Tang, M.M.; Wang, W.B.; Huang, J.P.; Jiang, X.S. tBHQ attenuates podocyte injury in diabetic nephropathy by inhibiting NADPH oxidase-derived ROS generation via the Nrf2/HO-1 signalling pathway. Heliyon 2022, 8, e10515. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, L.; Wang, F.; Shi, Y.; Ren, Y.; Liu, Q.; Cao, Y.; Duan, H. Attenuation of glomerular injury in diabetic mice with tert-butylhydroquinone through nuclear factor erythroid 2-related factor 2-dependent antioxidant gene activation. Am. J. Nephrol. 2011, 33, 289–297. [Google Scholar] [CrossRef]
- Adelusi, T.; Li, X.; Xu, L.; Du, L.; Hao, M.; Zhou, X.; Chowdhry, A.; Sun, Y.; Gu, X.; Lu, Q.; et al. Novel Chalcone BDD-39 Mitigated Diabetic Nephropathy through the Activation of Nrf2/ARE Signaling. Curr. Mol. Pharmacol. 2022, 15, 658–675. [Google Scholar] [CrossRef]
- Xiao, L.; Xu, X.; Zhang, F.; Wang, M.; Xu, Y.; Tang, D.; Wang, J.; Qin, Y.; Liu, Y.; Tang, C.; et al. The mitochondria-targeted antioxidant MitoQ ameliorated tubular injury mediated by mitophagy in diabetic kidney disease via Nrf2/PINK1. Redox Biol. 2017, 11, 297–311. [Google Scholar] [CrossRef]
- Bao, L.; Li, J.; Zha, D.; Zhang, L.; Gao, P.; Yao, T.; Wu, X. Chlorogenic acid prevents diabetic nephropathy by inhibiting oxidative stress and inflammation through modulation of the Nrf2/HO-1 and NF-kB pathways. Int. Immunopharmacol. 2018, 54, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, X.; Zhou, S.; Jia, Y.; Li, Y.; Song, Y.; Wang, J.; Wu, H. SP600125 suppresses Keap1 expression and results in NRF2-mediated prevention of diabetic nephropathy. J. Mol. Endocrinol. 2018, 60, 145–157. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y.; Chen, Q.; Yang, H.; Xie, X. Astaxanthin Promotes Nrf2/ARE Signaling to Alleviate Renal Fibronectin and Collagen IV Accumulation in Diabetic Rats. J. Diabetes Res. 2018, 2018, 6730315. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.J.; Wang, H.R.; Wang, Y.I.; Zhai, Z.H.; Wang, L.W.; Li, L.; Zhang, C.; Tang, L. Myricetin Attenuated Diabetes-Associated Kidney Injuries and Dysfunction via Regulating Nuclear Factor (Erythroid Derived 2)-Like 2 and Nuclear Factor-kappaB Signaling. Front. Pharmacol. 2019, 10, 647. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, T.; Wang, H.; Jiang, Y.; Peng, S. Obacunone attenuates high glucose-induced oxidative damage in NRK-52E cells by inhibiting the activity of GSK-3beta. Biochem. Biophys. Res. Commun. 2019, 513, 226–233. [Google Scholar] [CrossRef]
- Lv, C.; Cheng, T.; Zhang, B.; Sun, K.; Lu, K. Triptolide protects against podocyte injury in diabetic nephropathy by activating the Nrf2/HO-1 pathway and inhibiting the NLRP3 inflammasome pathway. Ren. Fail. 2023, 45, 2165103. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, Y.; Hu, J.E.; Zhang, M. Solasonine alleviates high glucose-induced podocyte injury through increasing Nrf2-medicated inhibition of NLRP3 activation. Drug Dev. Res. 2022, 83, 1697–1706. [Google Scholar] [CrossRef]
- Shahzad, K.; Bock, F.; Al-Dabet, M.M.; Gadi, I.; Nazir, S.; Wang, H.; Kohli, S.; Ranjan, S.; Mertens, P.R.; Nawroth, P.P.; et al. Stabilization of endogenous Nrf2 by minocycline protects against Nlrp3-inflammasome induced diabetic nephropathy. Sci. Rep. 2016, 6, 34228. [Google Scholar] [CrossRef]
- Du, L.; Wang, J.; Chen, Y.; Li, X.; Wang, L.; Li, Y.; Jin, X.; Gu, X.; Hao, M.; Zhu, X.; et al. Novel biphenyl diester derivative AB-38b inhibits NLRP3 inflammasome through Nrf2 activation in diabetic nephropathy. Cell Biol. Toxicol. 2020, 36, 243–260. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Z.; Ying, K.; Wang, H.; Liu, P.; Ji, X.; Chi, T.; Zou, L.; Wang, S.; He, Z. WJ-39, an Aldose Reductase Inhibitor, Ameliorates Renal Lesions in Diabetic Nephropathy by Activating Nrf2 Signaling. Oxid. Med. Cell Longev. 2020, 2020, 7950457. [Google Scholar] [CrossRef]
- Ding, B.; Geng, S.; Hou, X.; Ma, X.; Xu, H.; Yang, F.; Liu, K.; Liang, W.; Ma, G. Berberine Reduces Renal Cell Pyroptosis in Golden Hamsters with Diabetic Nephropathy through the Nrf2-NLRP3-Caspase-1-GSDMD Pathway. Evid. Based Complement. Altern. Med. 2021, 2021, 5545193. [Google Scholar] [CrossRef]
- Ding, X.; Zhao, H.; Qiao, C. Icariin protects podocytes from NLRP3 activation by Sesn2-induced mitophagy through the Keap1-Nrf2/HO-1 axis in diabetic nephropathy. Phytomedicine 2022, 99, 154005. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Chang, L.; Ren, Y.; Sui, M.; Fu, Y.; Zhang, L.; Hao, L. Quercetin improves diabetic kidney disease by inhibiting ferroptosis and regulating the Nrf2 in streptozotocin-induced diabetic rats. Ren. Fail. 2024, 46, 2327495. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Yang, Y.; Qiao, Y.; Zheng, Y.; Yu, X.; Liu, F.; Wang, H.; Zheng, B.; Pan, S.; Ren, K.; et al. Quercetin Ameliorates Diabetic Kidney Injury by Inhibiting Ferroptosis via Activating Nrf2/HO-1 Signaling Pathway. Am. J. Chin. Med. 2023, 51, 997–1018. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Wu, H.X.; Shi, W.Q.; Yang, Y.; Lin, M.; Wang, K.; Bian, C.C.; An, X.F.; Wang, T.; Yan, M. Triptolide Attenuates Renal Slit Diagram to Tight Junction Transition in Diabetic Kidney Disease by Regulating Nrf2-Ferroptosis Pathway. Am. J. Chin. Med. 2024, 52, 2161–2185. [Google Scholar] [CrossRef]
- Wang, H.; Yu, X.; Liu, D.; Qiao, Y.; Huo, J.; Pan, S.; Zhou, L.; Wang, R.; Feng, Q.; Liu, Z. VDR Activation Attenuates Renal Tubular Epithelial Cell Ferroptosis by Regulating Nrf2/HO-1 Signaling Pathway in Diabetic Nephropathy. Adv. Sci. 2024, 11, e2305563. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Y.; Liu, X.; Guo, S.; Jiang, L.; Huang, Y.; Wu, Y. Carnosine alleviates kidney tubular epithelial injury by targeting NRF2 mediated ferroptosis in diabetic nephropathy. Amino Acids 2023, 55, 1141–1155. [Google Scholar] [CrossRef]
- Lu, Q.; Yang, L.; Xiao, J.J.; Liu, Q.; Ni, L.; Hu, J.W.; Yu, H.; Wu, X.; Zhang, B.F. Empagliflozin attenuates the renal tubular ferroptosis in diabetic kidney disease through AMPK/NRF2 pathway. Free Radic. Biol. Med. 2023, 195, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Tao, P.; Wang, Q.; Cui, M.; Cao, M.; Xu, Y. Emodin attenuates diabetic kidney disease by inhibiting ferroptosis via upregulating Nrf2 expression. Aging 2023, 15, 7673–7688. [Google Scholar] [CrossRef]
- Wang, G.; Deng, J.; Hua, Z. Crocin protects against endoplasmic reticulum stress-related tubular injury in diabetic nephropathy via the activation of the PI3K/AKT/Nrf2 pathway. Iran. J. Basic. Med. Sci. 2024, 27, 439–446. [Google Scholar] [CrossRef]
- Zhao, K.; Li, Y.; Wang, Z.; Han, N.; Wang, Y. Carnosine Protects Mouse Podocytes from High Glucose Induced Apoptosis through PI3K/AKT and Nrf2 Pathways. Biomed. Res. Int. 2019, 2019, 4348973. [Google Scholar] [CrossRef]
- Jayaraman, S.; Prasad, M.; Natarajan, S.R.; Krishnamoorthy, R.; Alshuniaber, M.A.; Gatasheh, M.K.; Veeraraghavan, V.P.; Rajagopal, P.; Palanisamy, C.P. Molecular mechanisms underlying the effects of beta-sitosterol on TGF-beta1/Nrf2/SIRT1/p53-mediated signaling in the kidney of a high-fat diet and sucrose-induced type-2 diabetic rat. Chem. Biol. Interact. 2025, 411, 111443. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Pan, Q.; Lin, X.; Li, X.; Qu, L. Gastrodin ameliorates diabetic nephropathy by activating the AMPK/Nrf2 pathway. J. Mol. Histol. 2024, 55, 1327–1339. [Google Scholar] [CrossRef]
- Hongmei, H.; Maojun, Y.; Ting, L.I.; Dandan, W.; Ying, L.I.; Xiaochi, T.; Lu, Y.; Shi, G.U.; Yong, X.U. Neferine inhibits the progression of diabetic nephropathy by modulating the miR-17-5p/nuclear factor E2-related factor 2 axis. J. Tradit. Chin. Med. 2024, 44, 44–53. [Google Scholar] [CrossRef]
- Fu, H.; He, J.; Li, C.; Chang, H. Theaflavin-3,3′-Digallate Protects Liver and Kidney Functions in Diabetic Rats by Up-Regulating Circ-ITCH and Nrf2 Signaling Pathway. J. Agric. Food Chem. 2024, 72, 14630–14639. [Google Scholar] [CrossRef]
- Shen, Q.; Fang, J.; Guo, H.; Su, X.; Zhu, B.; Yao, X.; Wang, Y.; Cao, A.; Wang, H.; Wang, L. Astragaloside IV attenuates podocyte apoptosis through ameliorating mitochondrial dysfunction by up-regulated Nrf2-ARE/TFAM signaling in diabetic kidney disease. Free Radic. Biol. Med. 2023, 203, 45–57. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, X.; Chen, J.; Song, S.; Fang, S.; Wang, Z.; Xu, S.; Xu, Y.; Liu, J.; Jiang, C.; et al. Asiatic acid attenuates tubular injury in diabetic kidney disease by regulating mitochondrial dynamics via the Nrf-2 pathway. Phytomedicine 2023, 109, 154552. [Google Scholar] [CrossRef] [PubMed]
- Hofni, A.; Ali, F.E.M.; Ibrahim, A.R.N.; Aboubaker, E.M. Renoprotective Effect of Thymoquinone against Streptozotocin-Induced Diabetic Nephropathy: Role of NOX2 and Nrf2 Signals. Curr. Mol. Pharmacol. 2023, 16, 905–914. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Dai, L.; Wang, Y.; Zhao, C.; Wang, W.; Zhang, Y.; Zhao, Y.; Yu, T. 1,2,3,4,6-penta-O-galloyl-β-D-glucose alleviates inflammation and oxidative stress in diabetic nephropathy rats through MAPK/NF-κB and ERK/Nrf2/HO-1 signaling pathways. Exp. Ther. Med. 2022, 24, 639. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zeng, J.; Li, C.; Xiao, H.; Li, S.; Lin, Z.; Huang, K.; Shen, J.; Huang, H. Fraxin Promotes the Activation of Nrf2/ARE Pathway via Increasing the Expression of Connexin43 to Ameliorate Diabetic Renal Fibrosis. Front. Pharmacol. 2022, 13, 853383. [Google Scholar] [CrossRef]
- Yang, G.; Li, Q.; Peng, J.; Jin, L.; Zhu, X.; Zheng, D.; Zhang, Y.; Wang, R.; Song, Y.; Hu, W.; et al. Fucoxanthin regulates Nrf2 signaling to decrease oxidative stress and improves renal fibrosis depending on Sirt1 in HG-induced GMCs and STZ-induced diabetic rats. Eur. J. Pharmacol. 2021, 913, 174629. [Google Scholar] [CrossRef]
- Huang, X.; Shi, Y.; Chen, H.; Le, R.; Gong, X.; Xu, K.; Zhu, Q.; Shen, F.; Chen, Z.; Gu, X.; et al. Isoliquiritigenin prevents hyperglycemia-induced renal injuries by inhibiting inflammation and oxidative stress via SIRT1-dependent mechanism. Cell Death Dis. 2020, 11, 1040. [Google Scholar] [CrossRef]
- Ma, L.; Wu, F.; Shao, Q.; Chen, G.; Xu, L.; Lu, F. Baicalin Alleviates Oxidative Stress and Inflammation in Diabetic Nephropathy via Nrf2 and MAPK Signaling Pathway. Drug Des. Dev. Ther. 2021, 15, 3207–3221. [Google Scholar] [CrossRef]
- Gong, P.; Wang, P.; Pi, S.; Guo, Y.; Pei, S.; Yang, W.; Chang, X.; Wang, L.; Chen, F. Proanthocyanidins Protect Against Cadmium-Induced Diabetic Nephropathy Through p38 MAPK and Keap1/Nrf2 Signaling Pathways. Front. Pharmacol. 2021, 12, 801048. [Google Scholar] [CrossRef]
- Yu, W.C.; Huang, R.Y.; Chou, T.C. Oligo-Fucoidan Improves Diabetes-Induced Renal Fibrosis via Activation of Sirt-1, GLP-1R, and Nrf2/HO-1: An In Vitro and In Vivo Study. Nutrients 2020, 12, 3068. [Google Scholar] [CrossRef]
- Wu, H.; Kong, L.; Cheng, Y.; Zhang, Z.; Wang, Y.; Luo, M.; Tan, Y.; Chen, X.; Miao, L.; Cai, L. Metallothionein plays a prominent role in the prevention of diabetic nephropathy by sulforaphane via up-regulation of Nrf2. Free Radic. Biol. Med. 2015, 89, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Shang, G.; Tang, X.; Gao, P.; Guo, F.; Liu, H.; Zhao, Z.; Chen, Q.; Jiang, T.; Zhang, N.; Li, H. Sulforaphane attenuation of experimental diabetic nephropathy involves GSK-3 beta/Fyn/Nrf2 signaling pathway. J. Nutr. Biochem. 2015, 26, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Wang, Q.; Guo, F.; Song, Y.; Fan, X.; Shao, M.; Chen, D.; Zhang, W.; Qin, G. Sulforaphane ameliorated podocyte injury according to regulation of the Nrf2/PINK1 pathway for mitophagy in diabetic kidney disease. Eur. J. Pharmacol. 2023, 958, 176042. [Google Scholar] [CrossRef]
- Ma, T.; Zheng, Z.; Guo, H.; Lian, X.; Rane, M.J.; Cai, L.; Kim, K.S.; Kim, K.T.; Zhang, Z.; Bi, L. 4-O-methylhonokiol ameliorates type 2 diabetes-induced nephropathy in mice likely by activation of AMPK-mediated fatty acid oxidation and Nrf2-mediated anti-oxidative stress. Toxicol. Appl. Pharmacol. 2019, 370, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Gao, L.; Su, S.; Sargsyan, D.; Wu, R.; Raskin, I.; Kong, A.N. Moringa Isothiocyanate Activates Nrf2: Potential Role in Diabetic Nephropathy. AAPS J. 2019, 21, 31. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Kong, L.; Tang, Z.Z.; Zhang, Y.M.; Liu, Y.; Wang, T.Y.; Liu, Y.W. Hesperetin ameliorates diabetic nephropathy in rats by activating Nrf2/ARE/glyoxalase 1 pathway. Biomed. Pharmacother. 2019, 111, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, Z.; Gong, W.; Zou, Y.; Xu, F.; Chen, L.; Huang, H. Paeonol Ameliorates Diabetic Renal Fibrosis Through Promoting the Activation of the Nrf2/ARE Pathway via Up-Regulating Sirt1. Front. Pharmacol. 2018, 9, 512. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wang, Y.; Luo, M.; Tan, Y.; Cui, W.; Miao, L. Prevention of Streptozotocin-Induced Diabetic Nephropathy by MG132: Possible Roles of Nrf2 and IkappaB. Oxid. Med. Cell Longev. 2017, 2017, 3671751. [Google Scholar] [CrossRef]
- Cui, W.; Li, B.; Bai, Y.; Miao, X.; Chen, Q.; Sun, W.; Tan, Y.; Luo, P.; Zhang, C.; Zheng, S.; et al. Potential role for Nrf2 activation in the therapeutic effect of MG132 on diabetic nephropathy in OVE26 diabetic mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E87–E99. [Google Scholar] [CrossRef]
- Gong, W.; Li, J.; Chen, Z.; Huang, J.; Chen, Q.; Cai, W.; Liu, P.; Huang, H. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating CKIP-1 to resist HG-induced up-regulation of FN and ICAM-1 in GMCs and diabetic mice kidneys. Free Radic. Biol. Med. 2017, 106, 393–405. [Google Scholar] [CrossRef]
- Dong, W.; Jia, Y.; Liu, X.; Zhang, H.; Li, T.; Huang, W.; Chen, X.; Wang, F.; Sun, W.; Wu, H. Sodium butyrate activates NRF2 to ameliorate diabetic nephropathy possibly via inhibition of HDAC. J. Endocrinol. 2017, 232, 71–83. [Google Scholar] [CrossRef]
- Civantos, E.; Bosch, E.; Ramirez, E.; Zhenyukh, O.; Egido, J.; Lorenzo, O.; Mas, S. Sitagliptin ameliorates oxidative stress in experimental diabetic nephropathy by diminishing the miR-200a/Keap-1/Nrf2 antioxidant pathway. Diabetes Metab. Syndr. Obes. 2017, 10, 207–222. [Google Scholar] [CrossRef]
- Habib, S.L.; Yadav, A.; Kidane, D.; Weiss, R.H.; Liang, S. Novel protective mechanism of reducing renal cell damage in diabetes: Activation AMPK by AICAR increased NRF2/OGG1 proteins and reduced oxidative DNA damage. Cell Cycle 2016, 15, 3048–3059. [Google Scholar] [CrossRef]
- Gao, D.; Zhao, M.; Qi, X.; Liu, Y.; Li, N.; Liu, Z.; Bian, Y. Hypoglycemic effect of Gynostemma pentaphyllum saponins by enhancing the Nrf2 signaling pathway in STZ-inducing diabetic rats. Arch. Pharm. Res. 2016, 39, 221–230. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, J.; Guo, W.; Li, F.; Sun, W.; Chen, J.; Zhang, C.; Lu, X.; Tan, Y.; Feng, W.; et al. Up-regulation of Nrf2 is involved in FGF21-mediated fenofibrate protection against type 1 diabetic nephropathy. Free Radic. Biol. Med. 2016, 93, 94–109. [Google Scholar] [CrossRef]
- Stenvinkel, P.; Chertow, G.M.; Devarajan, P.; Levin, A.; Andreoli, S.P.; Bangalore, S.; Warady, B.A. Chronic Inflammation in Chronic Kidney Disease Progression: Role of Nrf2. Kidney Int. Rep. 2021, 6, 1775–1787. [Google Scholar] [CrossRef]
- Gyarmati, G.; Shroff, U.N.; Izuhara, A.; Hou, X.; Da Sacco, S.; Sedrakyan, S.; Lemley, K.V.; Amann, K.; Perin, L.; Peti-Peterdi, J. Intravital imaging reveals glomerular capillary distension and endothelial and immune cell activation early in Alport syndrome. JCI Insight 2022, 7, e152676. [Google Scholar] [CrossRef]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. Inflammatory processes in renal fibrosis. Nat. Rev. Nephrol. 2014, 10, 493–503. [Google Scholar] [CrossRef]
- Noone, D.; Licht, C. An update on the pathomechanisms and future therapies of Alport syndrome. Pediatr. Nephrol. 2013, 28, 1025–1036. [Google Scholar] [CrossRef]
- Bondi, C.D.; Hartman, H.L.; Tan, R.J. NRF2 in kidney physiology and disease. Physiol. Rep. 2024, 12, e15961. [Google Scholar] [CrossRef]
- Kaseda, S.; Sannomiya, Y.; Horizono, J.; Kuwazuru, J.; Suico, M.A.; Ogi, S.; Sasaki, R.; Sunamoto, H.; Fukiya, H.; Nishiyama, H.; et al. Novel Keap1-Nrf2 Protein-Protein Interaction Inhibitor UBE-1099 Ameliorates Progressive Phenotype in Alport Syndrome Mouse Model. Kidney360 2022, 3, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Chertow, G.M.; Appel, G.B.; Andreoli, S.; Bangalore, S.; Block, G.A.; Chapman, A.B.; Chin, M.P.; Gibson, K.L.; Goldsberry, A.; Iijima, K.; et al. Study Design and Baseline Characteristics of the CARDINAL Trial: A Phase 3 Study of Bardoxolone Methyl in Patients with Alport Syndrome. Am. J. Nephrol. 2021, 52, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Warady, B.A.; Pergola, P.E.; Agarwal, R.; Andreoli, S.; Appel, G.B.; Bangalore, S.; Block, G.A.; Chapman, A.B.; Chin, M.P.; Gibson, K.L.; et al. Effects of Bardoxolone Methyl in Alport Syndrome. Clin. J. Am. Soc. Nephrol. 2022, 17, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Avula, U.M.R.; Harris, L.; Hassanein, M. Bardoxolone for CKD: The Paradox of Confusion and Dogma. Kidney360 2022, 3, 1955–1960. [Google Scholar] [CrossRef]
- Chavez, E.; Rodriguez, J.; Drexler, Y.; Fornoni, A. Novel Therapies for Alport Syndrome. Front. Med. 2022, 9, 848389. [Google Scholar] [CrossRef]
- Miller, G.; Trevino, I.; Mccauley, L.; Hannigan, L.; Probst, B.; Ferguson, D.; Wigley, C. FO006. The NRF2 activator bardoxolone methyl inhibits CYST formation, reduces inflammation, and improves mitochondrial function in cellular models of polycystic kidney disease. Nephrol. Dial. Transplant. 2019, 34 (Suppl. 1), gfz096.FO006. [Google Scholar] [CrossRef]
- Qiu, Z.; He, J.; Shao, G.; Hu, J.; Li, X.; Zhou, H.; Li, M.; Yang, B. Obacunone Retards Renal Cyst Development in Autosomal Dominant Polycystic Kidney Disease by Activating NRF2. Antioxidants 2021, 11, 38. [Google Scholar] [CrossRef]
- Du, X.; Yu, J.; Sun, X.; Qu, S.; Zhang, H.; Hu, M.; Yang, S.; Zhou, P. Impact of epigallocatechin-3-gallate on expression of nuclear factor erythroid 2-related factor 2 and γ-glutamyl cysteine synthetase genes in oxidative stress-induced mouse renal tubular epithelial cells. Mol. Med. Rep. 2018, 17, 7952–7958. [Google Scholar] [CrossRef]
- Kanlaya, R.; Thongboonkerd, V. Protective Effects of Epigallocatechin-3-Gallate from Green Tea in Various Kidney Diseases. Adv. Nutr. 2019, 10, 112–121. [Google Scholar] [CrossRef]
- Kovacic, P.; Jacintho, J.D. Systemic lupus erythematosus and other autoimmune diseases from endogenous and exogenous agents: Unifying theme of oxidative stress. Mini Rev. Med. Chem. 2003, 3, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Yoh, K.; Itoh, K.; Enomoto, A.; Hirayama, A.; Yamaguchi, N.; Kobayashi, M.; Morito, N.; Koyama, A.; Yamamoto, M.; Takahashi, S. Nrf2-deficient female mice develop lupus-like autoimmune nephritis. Kidney Int. 2001, 60, 1343–1353. [Google Scholar] [CrossRef]
- Li, J.; Stein, T.D.; Johnson, J.A. Genetic dissection of systemic autoimmune disease in Nrf2-deficient mice. Physiol. Genom. 2004, 18, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Battelli, L.; Hubbs, A.F. Multiorgan autoimmune inflammation, enhanced lymphoproliferation, and impaired homeostasis of reactive oxygen species in mice lacking the antioxidant-activated transcription factor Nrf2. Am. J. Pathol. 2006, 168, 1960–1974. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, H.; Ding, Q.; Xu, X.; Yu, B.; Huang, Z. Nuclear Factor Erythroid 2-related Factor 2 Deficiency Exacerbates Lupus Nephritis in B6/lpr mice by Regulating Th17 Cell Function. Sci. Rep. 2016, 6, 38619. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guo, Q.; Wei, X.; Zhu, Y.; Luo, M.; Luo, P. Association of serum Nrf2 protein levels with disease activity and renal impairment in lupus nephritis. Front. Immunol. 2024, 15, 1304167. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Ka, S.M.; Chang, J.M.; Chen, H.C.; Shui, H.A.; Li, C.Y.; Hua, K.F.; Chang, W.L.; Huang, J.J.; Yang, S.S.; et al. Epigallocatechin-3-gallate prevents lupus nephritis development in mice via enhancing the Nrf2 antioxidant pathway and inhibiting NLRP3 inflammasome activation. Free Radic. Biol. Med. 2011, 51, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, S.; Tajima, H.; Ono, M. Nuclear factor erythroid 2-related factor 2 is a critical target for the treatment of glucocorticoid-resistant lupus nephritis. Arthritis Res. Ther. 2016, 18, 139. [Google Scholar] [CrossRef]
- Li, D.; Shi, G.; Wang, J.; Zhang, D.; Pan, Y.; Dou, H.; Hou, Y. Baicalein ameliorates pristane-induced lupus nephritis via activating Nrf2/HO-1 in myeloid-derived suppressor cells. Arthritis Res. Ther. 2019, 21, 105. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Soto, M.; Sanchez-Hidalgo, M.; Cardeno, A.; Rosillo, M.A.; Sanchez-Fidalgo, S.; Utrilla, J.; Martin-Lacave, I.; Alarcon-de-la-Lastra, C. Dietary extra virgin olive oil attenuates kidney injury in pristane-induced SLE model via activation of HO-1/Nrf-2 antioxidant pathway and suppression of JAK/STAT, NF-kappaB and MAPK activation. J. Nutr. Biochem. 2016, 27, 278–288. [Google Scholar] [CrossRef]
- Castejon, M.L.; Sanchez-Hidalgo, M.; Aparicio-Soto, M.; Montoya, T.; Martin-LaCave, I.; Fernandez-Bolanos, J.G.; Alarcon-de-la-Lastra, C. Dietary oleuropein and its new acyl-derivate attenuate murine lupus nephritis through HO-1/Nrf2 activation and suppressing JAK/STAT, NF-kappaB, MAPK and NLRP3 inflammasome signaling pathways. J. Nutr. Biochem. 2019, 74, 108229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Wu, H.; Wang, J.; Zhang, S. Esculetin alleviates murine lupus nephritis by inhibiting complement activation and enhancing Nrf2 signaling pathway. J. Ethnopharmacol. 2022, 288, 115004. [Google Scholar] [CrossRef]
- Ka, S.M.; Lin, J.C.; Lin, T.J.; Liu, F.C.; Chao, L.K.; Ho, C.L.; Yeh, L.T.; Sytwu, H.K.; Hua, K.F.; Chen, A. Citral alleviates an accelerated and severe lupus nephritis model by inhibiting the activation signal of NLRP3 inflammasome and enhancing Nrf2 activation. Arthritis Res. Ther. 2015, 17, 331. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Ka, S.M.; Chang, J.M.; Lai, J.H.; Dai, M.S.; Jheng, H.L.; Kuo, M.T.; Chen, P.; Chen, A. Antroquinonol differentially modulates T cell activity and reduces interleukin-18 production, but enhances Nrf2 activation, in murine accelerated severe lupus nephritis. Arthritis Rheum. 2012, 64, 232–242. [Google Scholar] [CrossRef]
- Sogawa, Y.; Nagasu, H.; Iwase, S.; Ihoriya, C.; Itano, S.; Uchida, A.; Kidokoro, K.; Taniguchi, S.; Takahashi, M.; Satoh, M.; et al. Infiltration of M1, but not M2, macrophages is impaired after unilateral ureter obstruction in Nrf2-deficient mice. Sci. Rep. 2017, 7, 8801. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K. Oxidative Stress in SLE T Cells, Is NRF2 Really the Target to Treat? Front. Immunol. 2021, 12, 633845. [Google Scholar] [CrossRef]
- Zhao, S.; Ghosh, A.; Lo, C.S.; Chenier, I.; Scholey, J.W.; Filep, J.G.; Ingelfinger, J.R.; Zhang, S.L.; Chan, J.S.D. Nrf2 Deficiency Upregulates Intrarenal Angiotensin-Converting Enzyme-2 and Angiotensin 1-7 Receptor Expression and Attenuates Hypertension and Nephropathy in Diabetic Mice. Endocrinology 2018, 159, 836–852. [Google Scholar] [CrossRef]
- Köhler, U.A.; Kurinna, S.; Schwitter, D.; Marti, A.; Schäfer, M.; Hellerbrand, C.; Speicher, T.; Werner, S. Activated Nrf2 impairs liver regeneration in mice by activation of genes involved in cell-cycle control and apoptosis. Hepatology 2014, 60, 670–678. [Google Scholar] [CrossRef]
- Gu, L.; Du, Y.; Chen, J.; Hasan, M.N.; Clayton, Y.D.; Matye, D.J.; Friedman, J.E.; Li, T. Cullin 3 RING E3 ligase inactivation causes NRF2-dependent NADH reductive stress, hepatic lipodystrophy, and systemic insulin resistance. Proc. Natl. Acad. Sci. USA 2024, 121, e2320934121. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, F.; Wang, M.; Wang, L.; Ye, C.; Yang, S.; Wang, C. Linking renal hypoxia and oxidative stress in chronic kidney disease: Based on clinical subjects and animal models. Biomol. Biomed. 2024, 24, 1319–1330. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Ibrahim, I.M.; Abd-Alhameed, E.K.; Sharawi, Z.W.; Jaber, F.A.; Althagafy, H.S. Nrf2/HO-1 as a therapeutic target in renal fibrosis. Life Sci. 2023, 334, 122209. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, N.; Su, X.; Zhou, G.; Sun, G.; Du, F.; Bian, X.; Wang, B. Epigallocatechin-3-gallate attenuates transforming growth factor-beta1 induced epithelial-mesenchymal transition via Nrf2 regulation in renal tubular epithelial cells. Biomed. Pharmacother. 2015, 70, 260–267. [Google Scholar] [CrossRef]
- Kanlaya, R.; Thongboonkerd, V. Molecular Mechanisms of Epigallocatechin-3-Gallate for Prevention of Chronic Kidney Disease and Renal Fibrosis: Preclinical Evidence. Curr. Dev. Nutr. 2019, 3, nzz101. [Google Scholar] [CrossRef]
- Kanlaya, R.; Khamchun, S.; Kapincharanon, C.; Thongboonkerd, V. Protective effect of epigallocatechin-3-gallate (EGCG) via Nrf2 pathway against oxalate-induced epithelial mesenchymal transition (EMT) of renal tubular cells. Sci. Rep. 2016, 6, 30233. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Li, X.J.; Wang, W.F.; Zou, L.; Miao, H.; Zhao, Y.Y. Geniposidic Acid Attenuates Chronic Tubulointerstitial Nephropathy Through Regulation of the NF-kB/Nrf2 Pathway via Aryl Hydrocarbon Receptor Signaling. Phytother. Res. 2024, 38, 5441–5457. [Google Scholar] [CrossRef]
- Patel, D.; Yadav, P.; Singh, S.K.; Tanwar, S.S.; Sehrawat, A.; Khurana, A.; Bhatti, J.S.; Navik, U. Betaine alleviates doxorubicin-induced nephrotoxicity by preventing oxidative insults, inflammation, and fibrosis through the modulation of Nrf2/HO-1/NLRP3 and TGF-beta expression. J. Biochem. Mol. Toxicol. 2024, 38, e23559. [Google Scholar] [CrossRef]
- Liu, Z.H.; Xu, Q.Y.; Wang, Y.; Gao, H.X.; Min, Y.H.; Jiang, X.W.; Yu, W.H. Catalpol from Rehmannia glutinosa Targets Nrf2/NF-kappaB Signaling Pathway to Improve Renal Anemia and Fibrosis. Am. J. Chin. Med. 2024, 52, 1451–1485. [Google Scholar] [CrossRef]
- Ding, N.; Sun, S.; Zhou, S.; Lv, Z.; Wang, R. Icariin alleviates renal inflammation and tubulointerstitial fibrosis via Nrf2-mediated attenuation of mitochondrial damage. Cell Biochem. Funct. 2024, 42, e4005. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Y.; Li, W.; Feng, J.; Chen, Y.; Chen, R.; Hu, L.; Wei, J. Fasudil attenuates oxidative stress-induced partial epithelial-mesenchymal transition of tubular epithelial cells in hyperuricemic nephropathy via activating Nrf2. Eur. J. Pharmacol. 2024, 975, 176640. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Kim, S.E.; Go, H.G.; Lee, Y.J.; Park, M.S.; Ko, S.; Han, B.S.; Yoon, Y.C.; Cho, Y.J.; Lee, P.; et al. Synergistic Renoprotective Effect of Melatonin and Zileuton by Inhibition of Ferroptosis via the AKT/mTOR/NRF2 Signaling in Kidney Injury and Fibrosis. Biomol. Ther. 2023, 31, 599–610. [Google Scholar] [CrossRef]
- Aihara, S.; Torisu, K.; Uchida, Y.; Imazu, N.; Nakano, T.; Kitazono, T. Spermidine from arginine metabolism activates Nrf2 and inhibits kidney fibrosis. Commun. Biol. 2023, 6, 676. [Google Scholar] [CrossRef]
- Ma, J.Q.; Zhang, Y.J.; Tian, Z.K.; Liu, C.M. Bixin attenuates carbon tetrachloride induced oxidative stress, inflammation and fibrosis in kidney by regulating the Nrf2/TLR4/MyD88 and PPAR-gamma/TGF-beta1/Smad3 pathway. Int. Immunopharmacol. 2021, 90, 107117. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y.; Wei, S.; Chen, L.; Xue, L.; Tian, H.; Tao, S. Bixin Confers Prevention against Ureteral Obstruction-Caused Renal Interstitial Fibrosis through Activation of the Nuclear Factor Erythroid-2-Related Factor2 Pathway in Mice. J. Agric. Food Chem. 2020, 68, 8321–8329. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yuan, D.; Gai, L.; Wu, X.; Shi, Y.; He, Y.; Liu, C.; Zhang, C.; Zhou, G.; Yuan, C. Saponins from Panax japonicus ameliorate age-related renal fibrosis by inhibition of inflammation mediated by NF-kappaB and TGF-beta1/Smad signaling and suppression of oxidative stress via activation of Nrf2-ARE signaling. J. Ginseng Res. 2021, 45, 408–419. [Google Scholar] [CrossRef]
- Wang, W.; Ma, B.L.; Xu, C.G.; Zhou, X.J. Dihydroquercetin protects against renal fibrosis by activating the Nrf2 pathway. Phytomedicine 2020, 69, 153185. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Sun, Y.Z.; Ming, Q.L.; Tian, Z.K.; Zhang, Y.J.; Liu, C.M. Effects of gastrodin against carbon tetrachloride induced kidney inflammation and fibrosis in mice associated with the AMPK/Nrf2/HMGB1 pathway. Food Funct. 2020, 11, 4615–4624. [Google Scholar] [CrossRef]
- Li, X.; Zou, Y.; Xing, J.; Fu, Y.Y.; Wang, K.Y.; Wan, P.Z.; Zhai, X.Y. Pretreatment with Roxadustat (FG-4592) Attenuates Folic Acid-Induced Kidney Injury through Antiferroptosis via Akt/GSK-3beta/Nrf2 Pathway. Oxid. Med. Cell Longev. 2020, 2020, 6286984. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dewanjee, S.; Dua, T.K.; Joardar, S.; Chakraborty, P.; Bhowmick, S.; Saha, A.; Bhattacharjee, S.; De Feo, V. Carnosic Acid Attenuates Cadmium Induced Nephrotoxicity by Inhibiting Oxidative Stress, Promoting Nrf2/HO-1 Signalling and Impairing TGF-beta1/Smad/Collagen IV Signalling. Molecules 2019, 24, 4176. [Google Scholar] [CrossRef]
- Zhang, G.; Kang, Y.; Zhou, C.; Cui, R.; Jia, M.; Hu, S.; Ji, X.; Yuan, J.; Cui, H.; Shi, G. Amelioratory Effects of Testosterone Propionate on Age-related Renal Fibrosis via Suppression of TGF-beta1/Smad Signaling and Activation of Nrf2-ARE Signaling. Sci. Rep. 2018, 8, 10726. [Google Scholar] [CrossRef]
- Polat, E.C.; Besiroglu, H.; Ozcan, L.; Otunctemur, A.; Eruyar, A.T.; Somay, A.; Ozbay, N.; Cekmen, M.; Eraldemir, C.; Ozbek, E. Beneficial effects of Oltipraz, nuclear factor-erythroid–2-related factor 2 (Nrf2), on renal damage in unilateral ureteral obstruction rat model. Int. Braz. J. Urol. 2018, 44, 1243–1251. [Google Scholar] [CrossRef]
- Aminzadeh, M.A.; Reisman, S.A.; Vaziri, N.D.; Khazaeli, M.; Yuan, J.; Meyer, C.J. The synthetic triterpenoid RTA dh404 (CDDO-dhTFEA) restores Nrf2 activity and attenuates oxidative stress, inflammation, and fibrosis in rats with chronic kidney disease. Xenobiotica 2014, 44, 570–578. [Google Scholar] [CrossRef]
- Berger, A.A.; Sottosanti, E.R.; Winnick, A.; Izygon, J.; Berardino, K.; Cornett, E.M.; Kaye, A.D.; Varrassi, G.; Viswanath, O.; Urits, I. Monomethyl Fumarate (MMF, Bafiertam) for the Treatment of Relapsing Forms of Multiple Sclerosis (MS). Neurol. Int. 2021, 13, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Venci, J.V.; Gandhi, M.A. Dimethyl fumarate (Tecfidera): A new oral agent for multiple sclerosis. Ann. Pharmacother. 2013, 47, 1697–1702. [Google Scholar] [CrossRef]
- Paik, J. Diroximel Fumarate in Relapsing Forms of Multiple Sclerosis: A Profile of Its Use. CNS Drugs 2021, 35, 691–700. [Google Scholar] [CrossRef]
- Abeti, R.; Baccaro, A.; Esteras, N.; Giunti, P. Novel Nrf2-Inducer Prevents Mitochondrial Defects and Oxidative Stress in Friedreich’s Ataxia Models. Front. Cell Neurosci. 2018, 12, 188. [Google Scholar] [CrossRef]
- Hernandez Martinez, A.P.; Cheng, Y.; Gindl-Bracho, A.; Cabrera-Jara, A.; Fernandez Yepez, A.K.; Martĭnez Hernández, M.F.; Sanchez-Lozada, L.G.; Tapia, E.; Castellanos, F.e.R.; Vazquez-Rangel, A.; et al. Effect of Oral Supplementation with Curcumin in Diabetic Subjects with Proteinuric Kidney Disease: A Randomized Controlled Trial: PO1026. J. Am. Soc. Nephrol. 2020, 31, 351. [Google Scholar] [CrossRef]
- Saldanha, J.F.; Leal, V.O.; Rizzetto, F.; Grimmer, G.H.; Ribeiro-Alves, M.; Daleprane, J.B.; Carraro-Eduardo, J.C.; Mafra, D. Effects of Resveratrol Supplementation in Nrf2 and NF-κB Expressions in Nondialyzed Chronic Kidney Disease Patients: A Randomized, Double-Blind, Placebo-Controlled, Crossover Clinical Trial. J. Ren. Nutr. 2016, 26, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Nair, V.; Keller, B.J.; Eichinger, F.; Hawkins, J.J.; Randolph, A.; Böger, C.A.; Gadegbeku, C.A.; Fox, C.S.; Cohen, C.D.; et al. Integrative biology identifies shared transcriptional networks in CKD. J. Am. Soc. Nephrol. 2014, 25, 2559–2572. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yan, Y. Therapeutic potential of sulforaphane in liver diseases: A review. Front. Pharmacol. 2023, 14, 1256029. [Google Scholar] [CrossRef] [PubMed]
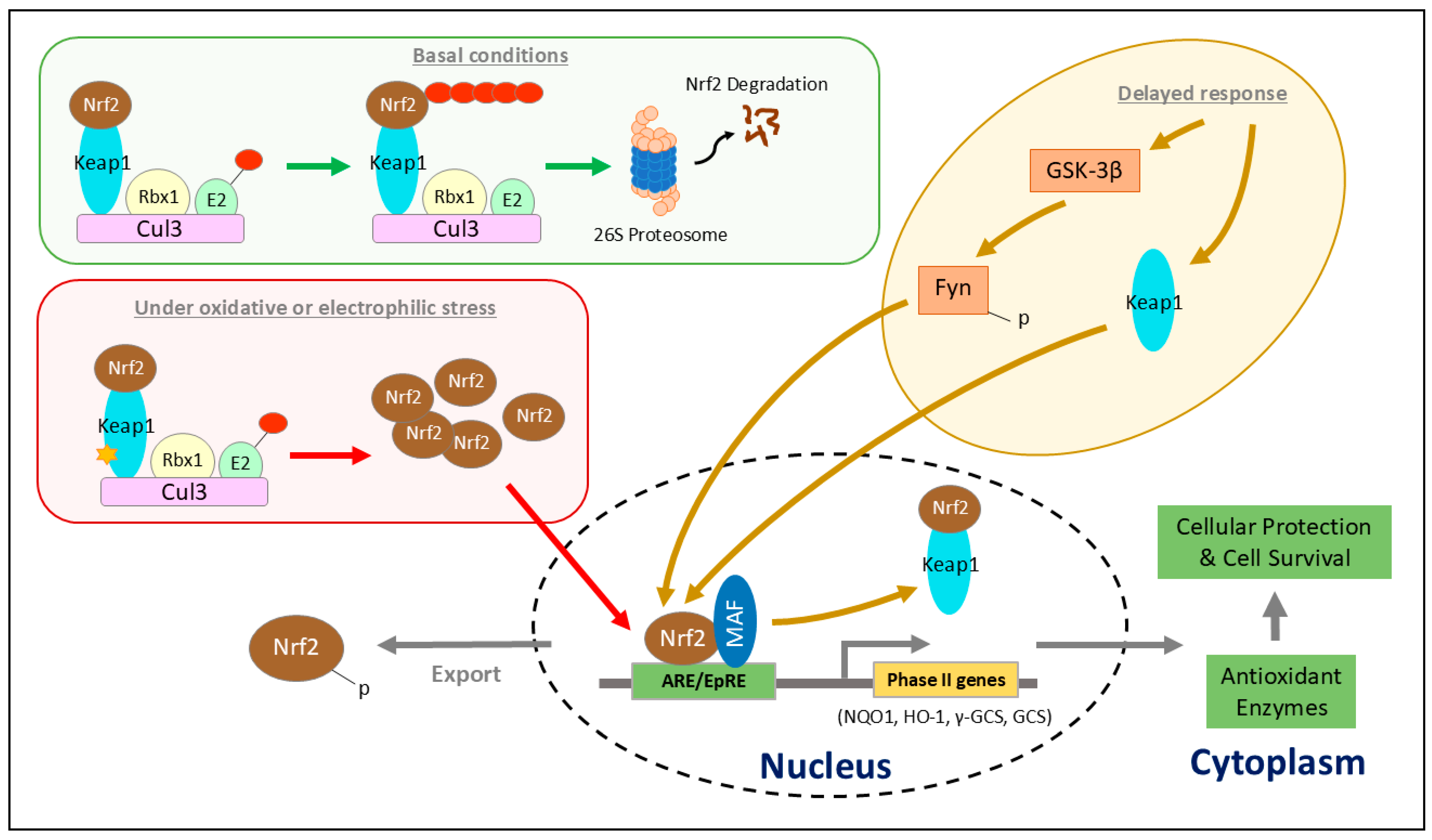
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, T.S.T.; Ng, K.H.; Zhang, Y. NRF2 Dysregulation and Therapeutic Insights Across Chronic Kidney Diseases. Int. J. Mol. Sci. 2025, 26, 7471. https://doi.org/10.3390/ijms26157471
Lim TST, Ng KH, Zhang Y. NRF2 Dysregulation and Therapeutic Insights Across Chronic Kidney Diseases. International Journal of Molecular Sciences. 2025; 26(15):7471. https://doi.org/10.3390/ijms26157471
Chicago/Turabian StyleLim, Tina Si Ting, Kar Hui Ng, and Yaochun Zhang. 2025. "NRF2 Dysregulation and Therapeutic Insights Across Chronic Kidney Diseases" International Journal of Molecular Sciences 26, no. 15: 7471. https://doi.org/10.3390/ijms26157471
APA StyleLim, T. S. T., Ng, K. H., & Zhang, Y. (2025). NRF2 Dysregulation and Therapeutic Insights Across Chronic Kidney Diseases. International Journal of Molecular Sciences, 26(15), 7471. https://doi.org/10.3390/ijms26157471