
Natural Flavonoids for the Prevention of Sarcopenia: Therapeutic Potential and Mechanisms



Abstract
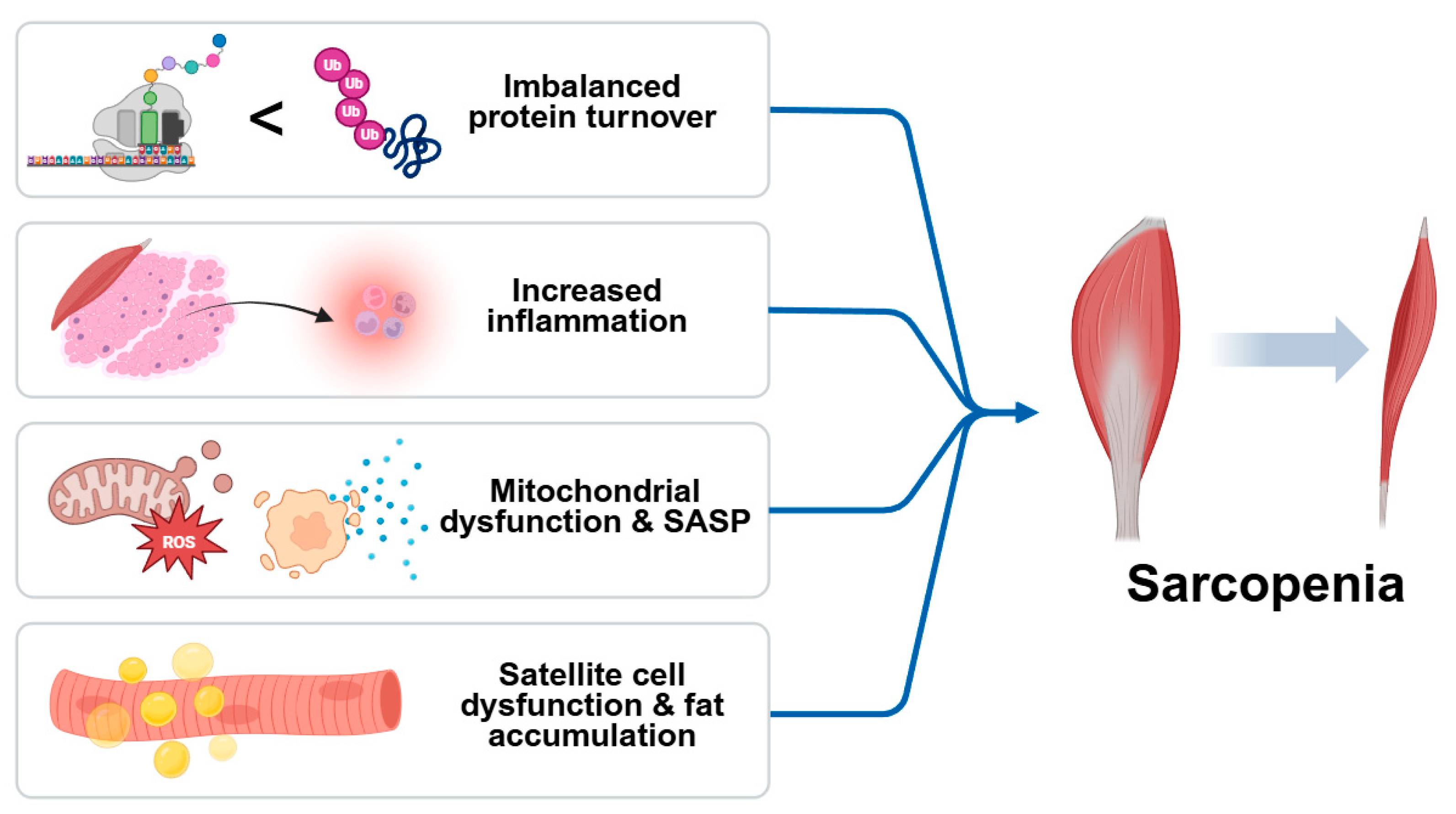
1. Introduction
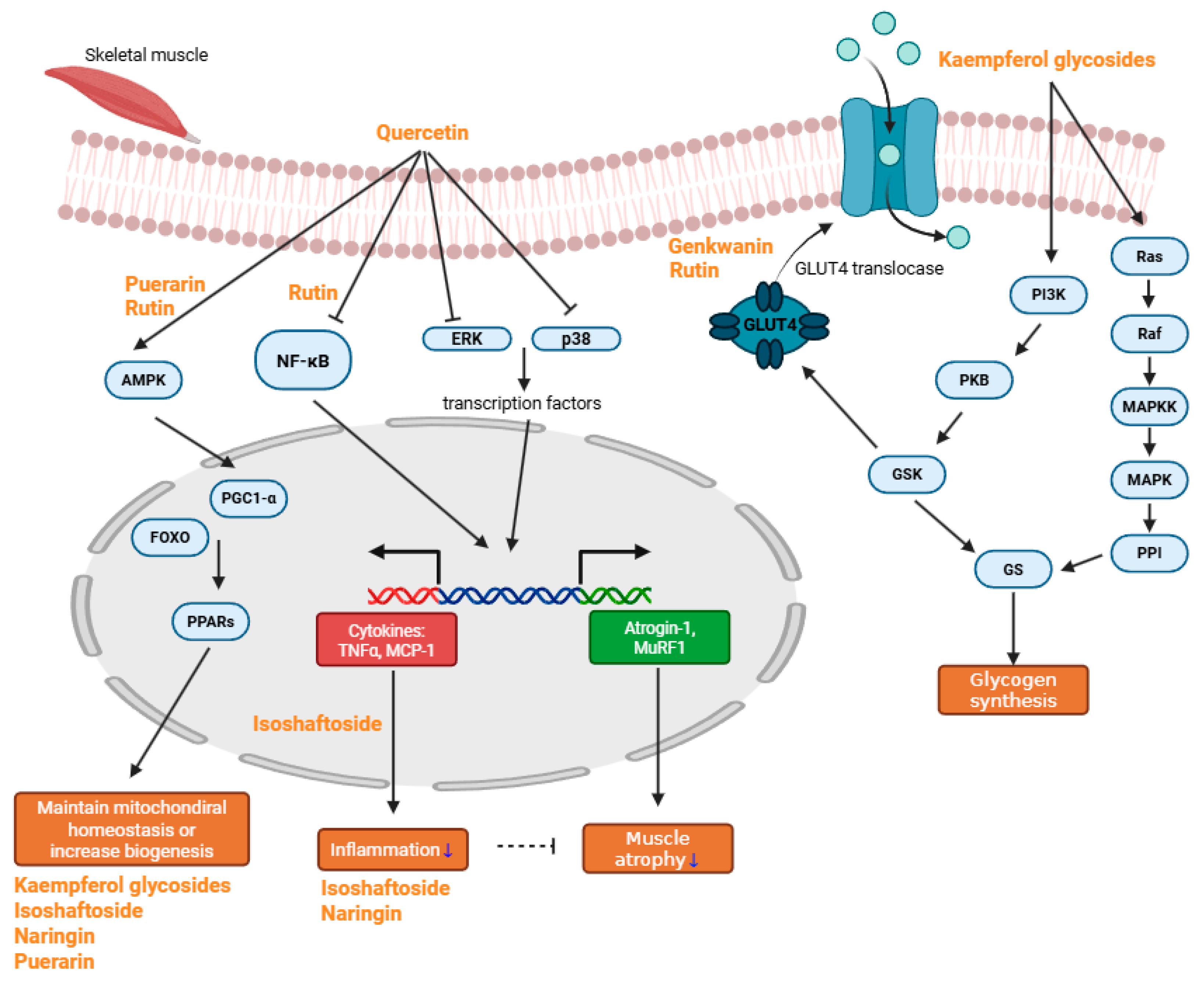
2. Anti-Sarcopenic Effects of Natural Compounds
2.1. Quercetin
2.2. Rutin
2.3. Kaempferol Glycosides
2.4. Baicalin
2.5. Genkwanin
2.6. Isoschaftoside
2.7. Naringin
2.8. Eriocitrin
2.9. Puerarin
3. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-NBDG | 2-(N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino)-2-deoxyglucose |
| ACC | Acetyl-CoA Carboxylase |
| ADIPOQ | Adiponectin |
| AKT | Protein Kinase B |
| AMPK | AMP-activated Protein Kinase |
| ATP/ADP translocase | Adenine Nucleotide Translocator |
| Atrogin-1 | Muscle Atrophy F-box Protein |
| BUN | Blood Urea Nitrogen |
| Bax | Bcl-2-associated X Protein |
| Bcl2 | B-cell lymphoma 2 |
| CAMKII | Calcium/Calmodulin-dependent Protein Kinase II |
| CAMKKβ | Calcium/Calmodulin-dependent Protein Kinase Kinase Beta |
| CAT | Catalase |
| CEBPA | CCAAT/Enhancer-binding Protein Alpha |
| CPT1 | Carnitine Palmitoyltransferase 1 |
| EF-1α | Elongation factor 1-alpha |
| eIF2α | eukaryotic translation initiation factor 2 subunit alpha |
| ERK | Extracellular Signal-Regulated Kinase |
| Elongation factor 1-alpha | Translation Elongation Factor 1 Alpha |
| FABP4 | Fatty Acid Binding Protein 4 |
| FOXO3 | Forkhead Box O3 |
| Foxo1 | Forkhead Box O1 |
| Foxo6 | Forkhead Box O6 |
| GLUT4 | Glucose Transporter Type 4 |
| GOT | Glutamate Oxaloacetate Transaminase |
| GPT | Glutamate Pyruvate Transaminase |
| GSH-Px | Glutathione Peroxidase |
| GSK-3 | Glycogen Synthase Kinase-3 |
| HO1 | Heme Oxygenase 1 |
| IKK | Inhibitor of κB Kinase |
| IL-1α | Interleukin-1 Alpha |
| IL-1β | Interleukin-1 Beta |
| IL-6 | Interleukin-6 |
| IκBα | Inhibitor of Nuclear Factor Kappa-B Alpha |
| LC3-II | Microtubule-associated Proteins 1A/1B Light Chain 3B |
| MAFbx | Muscle Atrophy F-box Protein |
| MAPK | Mitogen-Activated Protein Kinase |
| MDA | Malondialdehyde |
| MHC | Myosin Heavy Chain |
| MuRF1 | Muscle RING-finger Protein-1 |
| MyHC-II | Myosin Heavy Chain II |
| Myf5 | Myogenic Factor 5 |
| Myf6 | Myogenic Factor 6 |
| Myh4 | Myosin Heavy Chain 4 |
| MyoD | Myogenic Differentiation 1 |
| MyoG | Myogenin |
| Myogenin | Myogenic Regulatory Factor |
| NF-κB | Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells |
| NRF1 | Nuclear Respiratory Factor 1 |
| Nox4 | NADPH Oxidase 4 |
| Nrf1 | Nuclear Respiratory Factor 1 |
| Nrf2 | Nuclear Factor Erythroid 2-related Factor 2 |
| PGC-1α | Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha |
| PI3K | Phosphoinositide 3-Kinase |
| PINK1/Parkin | PTEN-induced Kinase 1/Parkin RBR E3 Ubiquitin Protein Ligase |
| PP1 | Protein Phosphatase 1 |
| PPAR-γ | Peroxisome Proliferator-Activated Receptor Gamma |
| PPAR-δ | Peroxisome Proliferator-Activated Receptor Delta |
| Pax | Paired Box Protein |
| ROS | Reactive Oxygen Species |
| SIRT1 | Sirtuin 1 |
| SOD1 | Superoxide Dismutase 1 |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| TBARS | Thiobarbituric Acid Reactive Substances |
| TNFα | Tumor Necrosis Factor Alpha |
| Tfam | Transcription Factor A, Mitochondrial |
| iNOS | Inducible Nitric Oxide Synthase |
| nNOS | Neuronal Nitric Oxide Synthase |
References
- Ligthart-Melis, G.C.; Luiking, Y.C.; Kakourou, A.; Cederholm, T.; Maier, A.B.; de van der Schueren, M.A.E. Frailty, Sarcopenia, and Malnutrition Frequently (Co-)occur in Hospitalized Older Adults: A Systematic Review and Meta-analysis. J. Am. Med. Dir. Assoc. 2020, 21, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Petermann-Rocha, F.; Balntzi, V.; Gray, S.R.; Lara, J.; Ho, F.K.; Pell, J.P.; Celis-Morales, C. Global prevalence of sarcopenia and severe sarcopenia: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2022, 13, 86–99. [Google Scholar] [CrossRef]
- Carvalho do Nascimento, P.R.; Bilodeau, M.; Poitras, S. How do we define and measure sarcopenia? A meta-analysis of observational studies. Age Ageing 2021, 50, 1906–1913. [Google Scholar] [CrossRef]
- Zheng, Y.; Feng, J.; Yu, Y.; Ling, M.; Wang, X. Advances in sarcopenia: Mechanisms, therapeutic targets, and intervention strategies. Arch. Pharm. Res. 2024, 47, 301–324. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyere, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.K.; Woo, J.; Assantachai, P.; Auyeung, T.W.; Chou, M.Y.; Iijima, K.; Jang, H.C.; Kang, L.; Kim, M.; Kim, S.; et al. Asian Working Group for Sarcopenia: 2019 Consensus Update on Sarcopenia Diagnosis and Treatment. J. Am. Med. Dir. Assoc. 2020, 21, 300–307.e2. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wong, P.Y.; Chung, Y.L.; Chow, S.K.; Cheung, W.H.; Law, S.W.; Chan, J.C.N.; Wong, R.M.Y. Deciphering the “obesity paradox” in the elderly: A systematic review and meta-analysis of sarcopenic obesity. Obes. Rev. 2023, 24, e13534. [Google Scholar] [CrossRef]
- Gao, Q.; Hu, K.; Yan, C.; Zhao, B.; Mei, F.; Chen, F.; Zhao, L.; Shang, Y.; Ma, Y.; Ma, B. Associated Factors of Sarcopenia in Community-Dwelling Older Adults: A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 4291. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Shi, W.; Zou, M.; Zeng, Q.S.; Feng, Y.; Luo, Z.Y.; Gan, H.T. Diagnosis, prevalence, and outcomes of sarcopenia in kidney transplantation recipients: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2023, 14, 17–29. [Google Scholar] [CrossRef]
- Gao, Q.; Mei, F.; Shang, Y.; Hu, K.; Chen, F.; Zhao, L.; Ma, B. Global prevalence of sarcopenic obesity in older adults: A systematic review and meta-analysis. Clin. Nutr. 2021, 40, 4633–4641. [Google Scholar] [CrossRef]
- Negm, A.M.; Lee, J.; Hamidian, R.; Jones, C.A.; Khadaroo, R.G. Management of Sarcopenia: A Network Meta-Analysis of Randomized Controlled Trials. J. Am. Med. Dir. Assoc. 2022, 23, 707–714. [Google Scholar] [CrossRef]
- Beaudart, C.; Dawson, A.; Shaw, S.C.; Harvey, N.C.; Kanis, J.A.; Binkley, N.; Reginster, J.Y.; Chapurlat, R.; Chan, D.C.; Bruyere, O.; et al. Nutrition and physical activity in the prevention and treatment of sarcopenia: Systematic review. Osteoporos. Int. 2017, 28, 1817–1833. [Google Scholar] [CrossRef]
- Webster, J.M.; Kempen, L.; Hardy, R.S.; Langen, R.C.J. Inflammation and Skeletal Muscle Wasting During Cachexia. Front. Physiol. 2020, 11, 597675. [Google Scholar] [CrossRef]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Petersen, A.M.; Pedersen, B.K. The role of IL-6 in mediating the anti-inflammatory effects of exercise. J. Physiol. Pharmacol. 2006, 57 (Suppl. S10), 43–51. [Google Scholar]
- Carey, A.L.; Steinberg, G.R.; Macaulay, S.L.; Thomas, W.G.; Holmes, A.G.; Ramm, G.; Prelovsek, O.; Hohnen-Behrens, C.; Watt, M.J.; James, D.E.; et al. Interleukin-6 increases insulin-stimulated glucose disposal in humans and glucose uptake and fatty acid oxidation in vitro via AMP-activated protein kinase. Diabetes 2006, 55, 2688–2697. [Google Scholar] [CrossRef]
- Yang, W.; Hu, P. Skeletal muscle regeneration is modulated by inflammation. J. Orthop. Transl. 2018, 13, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, B.; Hassounah, F.; Price, S.R.; Klein, J.; Mohamed, T.M.A.; Wang, Y.; Park, J.; Cai, H.; Zhang, X.; et al. The impact of senescence on muscle wasting in chronic kidney disease. J. Cachexia Sarcopenia Muscle 2023, 14, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Freire, M.; de Cabo, R.; Studenski, S.A.; Ferrucci, L. The Neuromuscular Junction: Aging at the Crossroad between Nerves and Muscle. Front. Aging Neurosci. 2014, 6, 208. [Google Scholar] [CrossRef] [PubMed]
- Uchitomi, R.; Oyabu, M.; Kamei, Y. Vitamin D and Sarcopenia: Potential of Vitamin D Supplementation in Sarcopenia Prevention and Treatment. Nutrients 2020, 12, 3189. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.H.; Tsai, M.C.; Tsai, H.W.; Chang, C.C.; Hou, W.H. Effects of branched-chain amino acid-rich supplementation on EWGSOP2 criteria for sarcopenia in older adults: A systematic review and meta-analysis. Eur. J. Nutr. 2022, 61, 637–651. [Google Scholar] [CrossRef]
- Le, N.H.; Kim, C.-S.; Park, T.; Park, J.H.Y.; Sung, M.-K.; Lee, D.G.; Hong, S.-M.; Choe, S.-Y.; Goto, T.; Kawada, T. Quercetin protects against obesity-induced skeletal muscle inflammation and atrophy. Mediat. Inflamm. 2014, 2014, 834294. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, C.-S.; Joe, Y.; Chung, H.T.; Ha, T.Y.; Yu, R. Quercetin reduces tumor necrosis factor alpha-induced muscle atrophy by upregulation of heme oxygenase-1. J. Med. Food 2018, 21, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, Y.; Egawa, K.; Kanzaki, N.; Izumo, T.; Rogi, T.; Shibata, H. Quercetin glycosides prevent dexamethasone-induced muscle atrophy in mice. Biochem. Biophys. Rep. 2019, 18, 100618. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, T.; Kanzaki, N.; Otsuka, Y.; Izumo, T.; Shibata, H.; Machida, S. Quercetin inhibits adipogenesis of muscle progenitor cells in vitro. Biochem. Biophys. Rep. 2018, 13, 39–44. [Google Scholar] [CrossRef]
- Ohmae, S.; Akazawa, S.; Takahashi, T.; Izumo, T.; Rogi, T.; Nakai, M. Quercetin attenuates adipogenesis and fibrosis in human skeletal muscle. Biochem. Biophys. Res. Commun. 2022, 615, 24–30. [Google Scholar] [CrossRef]
- Chen, X.; Liang, D.; Huang, Z.; Jia, G.; Zhao, H.; Liu, G. Anti-fatigue effect of quercetin on enhancing muscle function and antioxidant capacity. J. Food Biochem. 2021, 45, e13968. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, Z.; Sun, L.; Qiu, M.; Deng, Q.; Fang, Z.; Wang, Y. Protective mechanisms of three antioxidants against T-2 toxin-induced muscle protein deterioration in shrimp. J. Sci. Food Agric. 2022, 102, 4883–4891. [Google Scholar] [CrossRef]
- Jiang, H.; Horiuchi, Y.; Hironao, K.-Y.; Kitakaze, T.; Yamashita, Y.; Ashida, H. Prevention effect of quercetin and its glycosides on obesity and hyperglycemia through activating AMPKα in high-fat diet-fed ICR mice. J. Clin. Biochem. Nutr. 2020, 67, 75–83. [Google Scholar] [CrossRef]
- Yamashita, Y.; Jiang, H.; Okada, F.; Kitakaze, T.; Yoshioka, Y.; Ashida, H. Single oral administration of quercetin glycosides prevented acute hyperglycemia by promoting GLUT4 translocation in skeletal muscles through the activation of AMPK in mice. J. Clin. Biochem. Nutr. 2024, 74, 37. [Google Scholar] [CrossRef]
- Kappel, V.D.; Zanatta, L.; Postal, B.G.; Silva, F.R.M.B. Rutin potentiates calcium uptake via voltage-dependent calcium channel associated with stimulation of glucose uptake in skeletal muscle. Arch. Biochem. Biophys. 2013, 532, 55–60. [Google Scholar] [CrossRef]
- Seo, S.; Lee, M.-S.; Chang, E.; Shin, Y.; Oh, S.; Kim, I.-H.; Kim, Y. Rutin increases muscle mitochondrial biogenesis with AMPK activation in high-fat diet-induced obese rats. Nutrients 2015, 7, 8152–8169. [Google Scholar] [CrossRef]
- Lee, H.; Kim, Y.I.; Kim, M.J.; Hahm, J.-H.; Seo, H.D.; Ha, T.Y.; Jung, C.H.; Ahn, J. Castor oil plant (Ricinus communis L.) leaves improve dexamethasone-induced muscle atrophy via Nrf2 activation. Front. Pharmacol. 2022, 13, 891762. [Google Scholar] [CrossRef]
- Liu, S.; Adewole, D.; Yu, L.; Sid, V.; Wang, B.; Karmin, O.; Yang, C. Rutin attenuates inflammatory responses induced by lipopolysaccharide in an in vitro mouse muscle cell (C2C12) model. Poult. Sci. 2019, 98, 2756–2764. [Google Scholar] [CrossRef] [PubMed]
- Hah, Y.-S.; Lee, W.K.; Lee, S.-J.; Lee, S.Y.; Seo, J.-H.; Kim, E.J.; Choe, Y.-I.; Kim, S.G.; Yoo, J.-I. Rutin Prevents Dexamethasone-Induced Muscle Loss in C2C12 Myotube and Mouse Model by Controlling FOXO3-Dependent Signaling. Antioxidants 2023, 12, 639. [Google Scholar] [CrossRef]
- Li, Y.; Qin, R.; Yan, H.; Wang, F.; Huang, S.; Zhang, Y.; Zhong, M.; Zhang, W.; Wang, Z. Inhibition of vascular smooth muscle cells premature senescence with rutin attenuates and stabilizes diabetic atherosclerosis. J. Nutr. Biochem. 2018, 51, 91–98. [Google Scholar] [CrossRef]
- Li, Y.; Mei, H.; Liu, Y.; Li, Z.; Qamar, H.; Yu, M.; Ma, X. Dietary supplementation with rutin alters meat quality, fatty acid profile, antioxidant capacity, and expression levels of genes associated with lipid metabolism in breast muscle of qingyuan partridge chickens. Foods 2023, 12, 2302. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, H.; Zhang, J.; Zhou, B.; He, X.; Wang, T.; Wang, C. Dietary rutin improves breast meat quality in heat-stressed broilers and protects mitochondria from oxidative attack via the AMPK/PINK1–Parkin pathway. J. Sci. Food Agric. 2023, 103, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhao, Y. Effect of clove on improving running ability in aging mice. J. Food Biochem. 2022, 46, e14339. [Google Scholar] [CrossRef] [PubMed]
- de Santana Aquino, D.F.; Monteiro, T.A.; Cardoso, C.A.L.; Vieira, S.C.H.; do Carmo Vieira, M.; de Picoli Souza, K.; Amaya-Farfan, J.; Carvalho, G.C.B.C.; Moura, C.S.; Morato, P.N. Investigation of the antioxidant and hypoglycemiant properties of Alibertia edulis (LC Rich.) AC Rich. leaves. J. Ethnopharmacol. 2020, 253, 112648. [Google Scholar] [CrossRef]
- Cazarolli, L.H.; Folador, P.; Pizzolatti, M.G.; Mena Barreto Silva, F.R. Signaling pathways of kaempferol-3-neohesperidoside in glycogen synthesis in rat soleus muscle. Biochimie 2009, 91, 843–849. [Google Scholar] [CrossRef]
- Kashyap, B.; Saikia, K.; Samanta, S.K.; Thakur, D.; Banerjee, S.K.; Borah, J.C.; Talukdar, N.C. Kaempferol 3-O-rutinoside from Antidesma acidum Retz. Stimulates glucose uptake through SIRT1 induction followed by GLUT4 translocation in skeletal muscle L6 cells. J. Ethnopharmacol. 2023, 301, 115788. [Google Scholar] [CrossRef] [PubMed]
- Ramon, P.; Bergmann, D.; Abdulla, H.; Sparks, J.; Omoruyi, F. Bioactive ingredients in K. pinnata extract and synergistic effects of combined K. pinnata and metformin preparations on antioxidant activities in diabetic and non-diabetic skeletal muscle cells. Int. J. Mol. Sci. 2023, 24, 6211. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.-B.; Fang, J.; Lu, X.-Y.; Sun, Z.-L. Kaempferol improves carcase characteristics in broiler chickens by regulating ANGPTL3 gene expression. Br. Poult. Sci. 2012, 53, 836–842. [Google Scholar] [CrossRef]
- Visco, D.B.; Manhães de Castro, R.; da Silva, M.M.; Costa de Santana, B.J.R.; Bezerra Gouveia, H.J.C.; de Moura Ferraz, M.L.R.; de Albuquerque, G.L.; Lacerda, D.C.; de Vasconcelos, D.A.A.; Guzman Quevedo, O. Neonatal kaempferol exposure attenuates gait and strength deficits and prevents altered muscle phenotype in a rat model of cerebral palsy. Int. J. Dev. Neurosci. 2023, 83, 80–97. [Google Scholar] [CrossRef]
- Li, B.; Wan, L.; Li, Y.; Yu, Q.; Chen, P.; Gan, R.; Yang, Q.; Han, Y.; Guo, C. Baicalin, a component of Scutellaria baicalensis, alleviates anorexia and inhibits skeletal muscle atrophy in experimental cancer cachexia. Tumor Biol. 2014, 35, 12415–12425. [Google Scholar] [CrossRef]
- Chae, H.-S.; Dale, O.; Mir, T.M.; Avula, B.; Zhao, J.; Khan, I.A.; Khan, S.I. A multitarget approach to evaluate the efficacy of Aquilaria sinensis flower extract against metabolic syndrome. Molecules 2022, 27, 629. [Google Scholar] [CrossRef]
- Salvadori, L.; Paiella, M.; Castiglioni, B.; Belladonna, M.L.; Manenti, T.; Ercolani, C.; Cornioli, L.; Clemente, N.; Scircoli, A.; Sardella, R. Equisetum arvense standardized dried extract hinders age-related osteosarcopenia. Biomed. Pharmacother. 2024, 174, 116517. [Google Scholar] [CrossRef]
- Han, J.-W.; Shin, S.-K.; Bae, H.R.; Lee, H.; Moon, S.Y.; Seo, W.D.; Kwon, E.-Y. Wheat seedlings extract ameliorates sarcopenia in aged mice by regulating protein synthesis and degradation with anti-inflammatory and mitochondrial biogenesis effects. Phytomedicine 2024, 130, 155747. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xue, Y.; Jia, G.; Zhao, H.; Liu, G.; Huang, Z. Antifatigue effect of naringin on improving antioxidant capacity and mitochondrial function and preventing muscle damage. Exp. Biol. Med. 2022, 247, 1776–1784. [Google Scholar] [CrossRef]
- Termkwancharoen, C.; Malakul, W.; Phetrungnapha, A.; Tunsophon, S. Naringin ameliorates skeletal muscle atrophy and improves insulin resistance in high-fat-diet-induced insulin resistance in obese rats. Nutrients 2022, 14, 4120. [Google Scholar] [CrossRef]
- Takase, T.; Ikeuchi, S.; Inoue, T.; Mukai, R. Eriocitrin contained in lemon peel ameliorates disuse muscle atrophy by suppressing the expression of atrogin-1 and murf-1 in denervated mice. J. Nat. Prod. 2021, 84, 2048–2052. [Google Scholar] [CrossRef]
- Yin, L.; Chen, X.; Li, N.; Jia, W.; Wang, N.; Hou, B.; Yang, H.; Zhang, L.; Qiang, G.; Yang, X. Puerarin ameliorates skeletal muscle wasting and fiber type transformation in STZ-induced type 1 diabetic rats. Biomed. Pharmacother. 2021, 133, 110977. [Google Scholar] [CrossRef]
- Chen, X.; Yi, L.; Song, S.; Wang, L.; Liang, Q.; Wang, Y.; Wu, Y.; Gao, Q. Puerarin attenuates palmitate-induced mitochondrial dysfunction, impaired mitophagy and inflammation in L6 myotubes. Life Sci. 2018, 206, 84–92. [Google Scholar] [CrossRef]
- Arai, Y.; Watanabe, S.; Kimira, M.; Shimoi, K.; Mochizuki, R.; Kinae, N. Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 2000, 130, 2243–2250. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, D.; Costelli, P.; Sampaolesi, M.; Penna, F. Role of inflammation in muscle homeostasis and myogenesis. Mediat. Inflamm. 2015, 2015, 805172. [Google Scholar] [CrossRef]
- Durham, W.J.; Dillon, E.L.; Sheffield-Moore, M. Inflammatory burden and amino acid metabolism in cancer cachexia. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-T.; Yin, Y.; Yang, Y.-J.; Lv, P.-J.; Shi, Y.; Lu, L.; Wei, L.-B. Resveratrol prevents TNF-α-induced muscle atrophy via regulation of Akt/mTOR/FoxO1 signaling in C2C12 myotubes. Int. Immunopharmacol. 2014, 19, 206–213. [Google Scholar] [CrossRef]
- Kumar, S.; Boehm, J.; Lee, J.C. p38 MAP kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Xu, T.; Wang, Y.; Wu, C.; Wang, L.; Yang, X.; Sun, H. The role of inflammatory factors in skeletal muscle injury. Biotarget 2018, 2, 2–7. [Google Scholar] [CrossRef]
- He, W.A.; Berardi, E.; Cardillo, V.M.; Acharyya, S.; Aulino, P.; Thomas-Ahner, J.; Wang, J.; Bloomston, M.; Muscarella, P.; Nau, P. NF-κB–mediated Pax7 dysregulation in the muscle microenvironment promotes cancer cachexia. J. Clin. Investig. 2013, 123, 4821–4835. [Google Scholar] [CrossRef]
- Reid, W.D.; Rurak, J.; Harris, R.L. Skeletal muscle response to inflammation—Lessons for chronic obstructive pulmonary disease. Crit. Care Med. 2009, 37, S372–S383. [Google Scholar] [CrossRef]
- Fanzani, A.; Zanola, A.; Rovetta, F.; Rossi, S.; Aleo, M.F. Cisplatin triggers atrophy of skeletal C2C12 myotubes via impairment of Akt signalling pathway and subsequent increment activity of proteasome and autophagy systems. Toxicol. Appl. Pharmacol. 2011, 250, 312–321. [Google Scholar] [CrossRef]
- Moylan, J.S.; Smith, J.D.; Chambers, M.A.; McLoughlin, T.J.; Reid, M.B. TNF induction of atrogin-1/MAFbx mRNA depends on Foxo4 expression but not AKT-Foxo1/3 signaling. Am. J. Physiol.-Cell Physiol. 2008, 295, C986–C993. [Google Scholar] [CrossRef]
- Penna, F.; Costamagna, D.; Fanzani, A.; Bonelli, G.; Baccino, F.M.; Costelli, P. Muscle wasting and impaired myogenesis in tumor bearing mice are prevented by ERK inhibition. PLoS ONE 2010, 5, e13604. [Google Scholar] [CrossRef]
- Zhang, G.; Li, Y.-P. p38β MAPK upregulates atrogin1/MAFbx by specific phosphorylation of C/EBPβ. Skelet. Muscle 2012, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morand, C.; Manach, C.; Crespy, V.; Remesy, C. Respective bioavailability of quercetin aglycone and its glycosides in a rat model. Biofactors 2000, 12, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and bioavailability of quercetin glycosides in humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Morand, C.; Demigne, C.; Texier, O.; Regerat, F.; Remesy, C. Bioavailability of rutin and quercetin in rats. FEBS Lett. 1997, 409, 12–16. [Google Scholar] [CrossRef]
- Collins, C.A.; Olsen, I.; Zammit, P.S.; Heslop, L.; Petrie, A.; Partridge, T.A.; Morgan, J.E. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell 2005, 122, 289–301. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Carlson, C.L.; Visser, M.; Kelley, D.E.; Scherzinger, A.; Harris, T.B.; Stamm, E.; Newman, A.B. Attenuation of skeletal muscle and strength in the elderly: The Health ABC Study. J. Appl. Physiol. 2001, 90, 2157–2165. [Google Scholar] [CrossRef]
- Azzabou, N.; Hogrel, J.-Y.; Carlier, P.G. NMR based biomarkers to study age-related changes in the human quadriceps. Exp. Gerontol. 2015, 70, 54–60. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Margioris, A.N. Sarcopenic obesity. Hormones 2018, 17, 321–331. [Google Scholar] [CrossRef]
- Zamboni, M.; Mazzali, G.; Fantin, F.; Rossi, A.; Di Francesco, V. Sarcopenic obesity: A new category of obesity in the elderly. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 388–395. [Google Scholar] [CrossRef]
- Baumgartner, R.N.; Koehler, K.M.; Gallagher, D.; Romero, L.; Heymsfield, S.B.; Ross, R.R.; Garry, P.J.; Lindeman, R.D. Epidemiology of sarcopenia among the elderly in New Mexico. Am. J. Epidemiol. 1998, 147, 755–763. [Google Scholar] [CrossRef]
- Kurisawa, M.; Chung, J.E.; Uyama, H.; Kobayashi, S. Enzymatic synthesis and antioxidant properties of poly (rutin). Biomacromolecules 2003, 4, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.-C. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, M.; Bagdassarian, V. Rutin content in plant products. J. Univ. Chem. Technol. Metall. 2009, 44, 201–203. [Google Scholar]
- Kreft, I.; Fabjan, N.; Yasumoto, K. Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 2006, 98, 508–512. [Google Scholar] [CrossRef]
- La Casa, C.; Villegas, I.; De La Lastra, C.A.; Motilva, V.; Calero, M.M. Evidence for protective and antioxidant properties of rutin, a natural flavone, against ethanol induced gastric lesions. J. Ethnopharmacol. 2000, 71, 45–53. [Google Scholar] [CrossRef]
- Sheu, J.-R.; Hsiao, G.; Chou, P.-H.; Shen, M.-Y.; Chou, D.-S. Mechanisms involved in the antiplatelet activity of rutin, a glycoside of the flavonol quercetin, in human platelets. J. Agric. Food Chem. 2004, 52, 4414–4418. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a potent antioxidant: Implications for neurodegenerative disorders. Oxidative Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed]
- Eo, S.-H.; Kim, S.-J. Rutin induces cyclooxygenase-2 (COX-2) and matrix metalloproteinase (MMP)-9 expression via the ERK and PI3K/Akt pathways in A549 human lung cancer cells. J. Converg. Inf. Technol. 2013, 8, 414. [Google Scholar]
- Zhao, B.; Xiong, Y.; Zhang, Y.; Jia, L.; Zhang, W.; Xu, X. Rutin promotes osteogenic differentiation of periodontal ligament stem cells through the GPR30-mediated PI3K/AKT/mTOR signaling pathway. Exp. Biol. Med. 2020, 245, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Ochi, A.; Abe, T.; Nakao, R.; Yamamoto, Y.; Kitahata, K.; Takagi, M.; Hirasaka, K.; Ohno, A.; Teshima-Kondo, S.; Taesik, G. N-myristoylated ubiquitin ligase Cbl-b inhibitor prevents on glucocorticoid-induced atrophy in mouse skeletal muscle. Arch. Biochem. Biophys. 2015, 570, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, M.; Shi, L.; Zhou, J.; Ma, R.; Feng, K.; Chen, X.; Xu, X.; Li, X.; Li, T. Panax ginseng total protein facilitates recovery from dexamethasone-induced muscle atrophy through the activation of glucose consumption in C2C12 myotubes. BioMed Res. Int. 2019, 2019, 3719643. [Google Scholar] [CrossRef]
- Choi, K.M. Sarcopenia and sarcopenic obesity. Korean J. Intern. Med. 2016, 31, 1054. [Google Scholar] [CrossRef]
- Singh, P.P.; Chauhan, S. Activity guided isolation of antioxidants from the leaves of Ricinus communis L. Food Chem. 2009, 114, 1069–1072. [Google Scholar] [CrossRef]
- Babu, P.R.; Bhuvaneswar, C.; Sandeep, G.; Ramaiah, C.V.; Rajendra, W. Hepatoprotective role of Ricinus communis leaf extract against d-galactosamine induced acute hepatitis in albino rats. Biomed. Pharmacother. 2017, 88, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Menegati, S.E.L.T.; de Lima, F.F.; Traesel, G.K.; Souza, R.I.C.; Dos Santos, A.C.; de Santana Aquino, D.F.; de Oliveira, V.S.; Vieira, S.C.H.; Cardoso, C.A.L.; do Carmo Vieira, M. Acute and subacute toxicity of the aqueous extract of Alibertia edulis (Rich.) A. Rich. ex DC. in rats. J. Ethnopharmacol. 2016, 194, 1096–1102. [Google Scholar] [CrossRef]
- Aziz, Z.; Tang, W.; Chong, N.; Tho, L. A systematic review of the efficacy and tolerability of hydroxyethylrutosides for improvement of the signs and symptoms of chronic venous insufficiency. J. Clin. Pharm. Ther. 2015, 40, 177–185. [Google Scholar] [CrossRef]
- Malinska, H.; Hüttl, M.; Oliyarnyk, O.; Markova, I.; Poruba, M.; Racova, Z.; Kazdova, L.; Vecera, R. Beneficial effects of troxerutin on metabolic disorders in non-obese model of metabolic syndrome. PLoS ONE 2019, 14, e0220377. [Google Scholar] [CrossRef]
- Pan, R.-Y.; Ma, J.; Kong, X.-X.; Wang, X.-F.; Li, S.-S.; Qi, X.-L.; Yan, Y.-H.; Cheng, J.; Liu, Q.; Jin, W. Sodium rutin ameliorates Alzheimer’s disease–like pathology by enhancing microglial amyloid-β clearance. Sci. Adv. 2019, 5, eaau6328. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Pan, R.; Cheng, J.; Cui, Q.; Chen, J.; Yuan, Z. Sodium rutin extends lifespan and health span in mice including positive impacts on liver health. Br. J. Pharmacol. 2022, 179, 1825–1838. [Google Scholar] [CrossRef]
- Boutry-Regard, C.; Vinyes-Pares, G.; Breuille, D.; Moritani, T. Supplementation with Whey Protein, Omega-3 Fatty Acids and Polyphenols Combined with Electrical Muscle Stimulation Increases Muscle Strength in Elderly Adults with Limited Mobility: A Randomized Controlled Trial. Nutrients 2020, 12, 1866. [Google Scholar] [CrossRef]
- Yang, N.; Qi, X.; Hu, J.; Teng, J.; Wang, Y.; Li, C. Exploring the mechanism of astragalus membranaceus in the treatment of multiple system atrophy based on network pharmacology and molecular docking. Medicine 2023, 102, e32523. [Google Scholar] [CrossRef]
- Kumar, M. A review on phytochemical constituents and pharmacological activities of Ricinus communis L. Plant. Int. J. Pharmacogn. Phytochem. Res. 2017, 9, 466–472. [Google Scholar] [CrossRef]
- Hayes, N.; Fogarty, M.; Sadofsky, L.; Jones, H.S. Physiologically-obtainable polyphenol exposures modulate reactive oxygen and nitrogen species signaling in the C2C12 model of skeletal muscle ageing. Eur. J. Pharmacol. 2023, 941, 175511. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, K.; Gu, Y.; Luo, C.; Mao, W.; Zhou, W.; Zhu, J.; Zhang, H. Kaempferol attenuates spinal cord injury by interfering inflammatory and oxidative stress by targeting the p53 protein: A molecular docking analysis. Mol. Cell. Toxicol. 2021, 17, 257–266. [Google Scholar] [CrossRef]
- De Araújo, E.R.D.; Guerra, G.C.B.; Araújo, D.F.d.S.; De Araújo, A.A.; Fernandes, J.M.; de Araújo Júnior, R.F.; Da Silva, V.C.; De Carvalho, T.G.; Ferreira, L.d.S.; Zucolotto, S.M. Gastroprotective and antioxidant activity of Kalanchoe brasiliensis and Kalanchoe pinnata leaf juices against indomethacin and ethanol-induced gastric lesions in rats. Int. J. Mol. Sci. 2018, 19, 1265. [Google Scholar] [CrossRef] [PubMed]
- Verschuren, O.; Smorenburg, A.R.; Luiking, Y.; Bell, K.; Barber, L.; Peterson, M.D. Determinants of muscle preservation in individuals with cerebral palsy across the lifespan: A narrative review of the literature. J. Cachexia Sarcopenia Muscle 2018, 9, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Imran, M.; Alsagaby, S.A.; Naeem, H.; Al Abdulmonem, W.; Hussain, M.; Abdelgawad, M.A.; El-Ghorab, A.H.; Ghoneim, M.M.; El-Sherbiny, M. Anticancer, antioxidant, ameliorative and therapeutic properties of kaempferol. Int. J. Food Prop. 2023, 26, 1140–1166. [Google Scholar] [CrossRef]
- Yangzom, P.; Amruthanand, S.; Sharma, M.; Mahajan, S.; Lingaraju, M.C.; Parida, S.; Sahoo, M.; Kumar, D.; Singh, T.U. Subacute 28 days oral toxicity study of kaempferol and biochanin-A in the mouse model. J. Biochem. Mol. Toxicol. 2022, 36, e23090. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Akiyama, M.; Mizokami, T.; Ito, H.; Ikeda, Y. A randomized, placebo-controlled trial evaluating the safety of excessive administration of kaempferol aglycone. Food Sci. Nutr. 2023, 11, 5427–5437. [Google Scholar] [CrossRef]
- Srinivas, N.R. Baicalin, an emerging multi-therapeutic agent: Pharmacodynamics, pharmacokinetics, and considerations from drug development perspectives. Xenobiotica 2010, 40, 357–367. [Google Scholar] [CrossRef]
- Lixuan, Z.; Jingcheng, D.; Wenqin, Y.; Jianhua, H.; Baojun, L.; Xiaotao, F. Baicalin attenuates inflammation by inhibiting NF-κB activation in cigarette smoke induced inflammatory models. Pulm. Pharmacol. Ther. 2010, 23, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, J.; Zhang, P.; Li, D.; Zhang, C.; Zhao, H.; Fu, J.; Wang, B.; Liu, J. Baicalin attenuates proinflammatory cytokine production in oxygen–glucose deprived challenged rat microglial cells by inhibiting TLR4 signaling pathway. Int. Immunopharmacol. 2012, 14, 749–757. [Google Scholar] [CrossRef]
- Zhang, N.; Van Crombruggen, K.; Holtappels, G.; Bachert, C. A herbal composition of Scutellaria baicalensis and Eleutherococcus senticosus shows potent anti-inflammatory effects in an ex vivo human mucosal tissue model. Evid.-Based Complement. Altern. Med. 2012, 2012, 673145. [Google Scholar] [CrossRef]
- Xiping, Z.; Guanghua, F.; Jinxian, H.; Weihong, W.; Rujun, X.; Wei, Z.; Jing, Y.; Qijun, Y.; Meijuan, Y.; Qing, W. Baicalin protects thymus of rats with severe acute pancreatitis. Inflammation 2010, 33, 157–165. [Google Scholar] [CrossRef]
- Lim, H.A.; Lee, E.K.; Kim, J.M.; Park, M.H.; Kim, D.H.; Choi, Y.J.; Ha, Y.M.; Yoon, J.-H.; Choi, J.S.; Yu, B.P. PPARγ activation by baicalin suppresses NF-κB-mediated inflammation in aged rat kidney. Biogerontology 2012, 13, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bao, H.; Wu, J.; Duan, X.; Liu, B.; Sun, J.; Gong, W.; Lv, Y.; Zhang, H.; Luo, Q. Baicalin is anti-inflammatory in cigarette smoke-induced inflammatory models in vivo and in vitro: A possible role for HDAC2 activity. Int. Immunopharmacol. 2012, 13, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Frantz, J.D.; Tawa, N.E.; Melendez, P.A.; Oh, B.-C.; Lidov, H.G.; Hasselgren, P.-O.; Frontera, W.R.; Lee, J.; Glass, D.J. IKKβ/NF-κB activation causes severe muscle wasting in mice. Cell 2004, 119, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.R.; Kim, H.J.; Lecker, S.H. Ubiquitin–protein ligases in muscle wasting. Int. J. Biochem. Cell Biol. 2005, 37, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.D.; Lecker, S.H.; Jagoe, R.T.; Navon, A.; Goldberg, A.L. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc. Natl. Acad. Sci. USA 2001, 98, 14440–14445. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.-M.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Bao, Y.; Sun, Y.-W.; Ji, J.; Gan, L.; Zhang, C.-F.; Wang, C.-Z.; Yuan, C.-S. Genkwanin ameliorates adjuvant-induced arthritis in rats through inhibiting JAK/STAT and NF-κB signaling pathways. Phytomedicine 2019, 63, 153036. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Stojanović, Z.; Quirantes-Piné, R.; Arráez-Román, D.; Švarc-Gajić, J.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Rosmarinus officinalis leaves as a natural source of bioactive compounds. Int. J. Mol. Sci. 2014, 15, 20585–20606. [Google Scholar] [CrossRef]
- Santos-Gomes, P.C.; Seabra, R.M.; Andrade, P.B.; Fernandes-Ferreira, M. Phenolic antioxidant compounds produced by in vitro shoots of sage (Salvia officinalis L.). Plant Sci. 2002, 162, 981–987. [Google Scholar] [CrossRef]
- Boukaabache, R.; Boubekri, N.; Boumaza, O.; Mekkiou, R.; Seghiri, R.; Sarri, D.; Zama, D.; Benayache, F.; Benayache, S. Phytochemical study of ethyl acetate extract and antioxidant activity of Genista quadriflora Munby (Fabaceae). Der Pharm. Lett. 2013, 5, 56–59. [Google Scholar]
- Park, S.-H.; Cui, X.; Ahn, D.; Lee, E.B.; Cha, D.S.; Jeon, H.; Zee, O.P.; Kim, Y.-C.; Kim, D.K. Anti-oxidative Activities of 3’-Hydroxygenkwanin from the Flower Buds of Daphne genkwa in Caenorhabditis elegans. Nat. Product. Sci. 2014, 20, 80–85. [Google Scholar]
- Thomas, A.; Rajesh, E.A.; Kumar, D.S. The significance of Tinospora crispa in treatment of diabetes mellitus. Phytother. Res. 2016, 30, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-W.; Bao, Y.; Yu, H.; Chen, Q.-J.; Lu, F.; Zhai, S.; Zhang, C.-F.; Li, F.; Wang, C.-Z.; Yuan, C.-S. Anti-rheumatoid arthritis effects of flavonoids from Daphne genkwa. Int. Immunopharmacol. 2020, 83, 106384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-F.; Zhang, S.-L.; He, X.; Yang, X.-L.; Wu, H.-T.; Lin, B.-Q.; Jiang, C.-P.; Wang, J.; Yu, C.-H.; Yang, Z.-L. Antioxidant effects of Genkwa flos flavonoids on Freund׳s adjuvant-induced rheumatoid arthritis in rats. J. Ethnopharmacol. 2014, 153, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Herbison, G.; Ditunno, J.; Jaweed, M. Muscle atrophy in rheumatoid arthritis. J. Rheumatol. Suppl. 1987, 14, 78–81. [Google Scholar]
- Ortenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef]
- Yao, L.; Liu, W.; Bashir, M.; Nisar, M.F.; Wan, C.C. Eriocitrin: A review of pharmacological effects. Biomed. Pharmacother. 2022, 154, 113563. [Google Scholar] [CrossRef]
- Minato, K.-i.; Miyake, Y.; Fukumoto, S.; Yamamoto, K.; Kato, Y.; Shimomura, Y.; Osawa, T. Lemon flavonoid, eriocitrin, suppresses exercise-induced oxidative damage in rat liver. Life Sci. 2003, 72, 1609–1616. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, B.; Chen, S.; Lin, M.; Chen, Y.; Jin, S.; Chen, W.; Zhang, Y. Applications of network pharmacology in traditional Chinese medicine research. Evid.-Based Complement. Altern. Med. 2020, 2020, 1646905. [Google Scholar] [CrossRef]
- Chen, R.; Xue, J.; Xie, M. Puerarin prevents isoprenaline-induced myocardial fibrosis in mice by reduction of myocardial TGF-β1 expression. J. Nutr. Biochem. 2012, 23, 1080–1085. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Neuroprotective herbs for stroke therapy in traditional eastern medicine. Neurol. Res. 2005, 27, 287–301. [Google Scholar] [CrossRef]
- Chen, X.-F.; Wang, L.; Wu, Y.-Z.; Song, S.-Y.; Min, H.-Y.; Yang, Y.; He, X.; Liang, Q.; Yi, L.; Wang, Y. Effect of puerarin in promoting fatty acid oxidation by increasing mitochondrial oxidative capacity and biogenesis in skeletal muscle in diabetic rats. Nutr. Diabetes 2018, 8, 1. [Google Scholar] [CrossRef]
- Jung, H.W.; Kang, A.N.; Kang, S.Y.; Park, Y.-K.; Song, M.Y. The root extract of Pueraria lobata and its main compound, puerarin, prevent obesity by increasing the energy metabolism in skeletal muscle. Nutrients 2017, 9, 33. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Liu, M.; Song, X.; Wang, D.; Zhang, J. Network pharmacology-based analysis of the effects of puerarin on sarcopenia. Ann. Transl. Med. 2022, 10, 671. [Google Scholar] [CrossRef]
- Yin, L.; Chen, X.; Li, N.; Jia, W.; Wang, N.; Hou, B.; Zhang, L.; Qiang, G.; Yang, X.; Du, G.H. Puerarin ameliorates skeletal muscle atrophy in STZ-induced type 1 diabetic rats by enhancing Akt/mTOR while inhibiting autophagy signaling pathway. Authorea Prepr. 2020. [Google Scholar] [CrossRef]
- Wu, M.; Zhao, G.; Yang, X.; Peng, C.; Zhao, J.; Liu, J.; Li, R.; Gao, Z. Puerarin accelerates neural regeneration after sciatic nerve injury. Neural Regen. Res. 2014, 9, 589–593. [Google Scholar] [CrossRef]
- Speisky, H.; Arias-Sante, M.F.; Fuentes, J. Oxidation of Quercetin and Kaempferol Markedly Amplifies Their Antioxidant, Cytoprotective, and Anti-Inflammatory Properties. Antioxidants 2023, 12, 155. [Google Scholar] [CrossRef]
- Conte, A.; Pellegrini, S.; Tagliazucchi, D. Synergistic protection of PC12 cells from beta-amyloid toxicity by resveratrol and catechin. Brain Res. Bull. 2003, 62, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yue, X.; Sun, Z.; Hambright, W.S.; Feng, Q.; Cui, Y.; Huard, J.; Robbins, P.D.; Wang, Z.; Mu, X. Senolytic elimination of senescent macrophages restores muscle stem cell function in severely dystrophic muscle. Aging 2022, 14, 7650–7661. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, R.; Nakashima, R.; Yano, M.; Iwahara, N.; Asakura, S.; Nojima, I.; Saga, Y.; Kunimoto, R.; Horio, Y.; Kuno, A. Resveratrol, a SIRT1 activator, attenuates aging-associated alterations in skeletal muscle and heart in mice. J. Pharmacol. Sci. 2023, 152, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.H.; Alizargar, J.; Peng, C.Y.; Wu, J.P. Combination of exercise training and resveratrol attenuates obese sarcopenia in skeletal muscle atrophy. Chin. J. Physiol. 2020, 63, 101–112. [Google Scholar] [CrossRef]
- Islam, M.T.; Tuday, E.; Allen, S.; Kim, J.; Trott, D.W.; Holland, W.L.; Donato, A.J.; Lesniewski, L.A. Senolytic drugs, dasatinib and quercetin, attenuate adipose tissue inflammation, and ameliorate metabolic function in old age. Aging Cell 2023, 22, e13767. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Compound | Model | Molecular Targets | Effects | Dose | References |
|---|---|---|---|---|---|
| Quercetin | C2C12 cells and C57BL/6 mice | ERK↓ p38↓ MAPK↓ NF-κB↓ Atrogin-1↓ MuRF1↓ | Synthesis and differentiation of muscle cells and reduction of inflammation | 50 μM 0.05% or 0.1% in diet | [22] |
| C2C12 cells and C57BL/6 mice | Atrogin-1↓ MuRF1↓ HO1↑ Nrf2↑ NF-κB↓ | Inhibition of TNFα-induced inflammatory response and muscle protein degradation | 20–50 μM 0.05% in diet | [23] | |
| C2C12 cells and BALB/c CrSlc mice | Atrogin-1↓ MuRF1↓ Foxo1↓ | Protection of muscle atrophy induced by dexamethasone | 10 μM 0.15% or 0.45% in water | [24] | |
| Primary mouse myoblast | PPAR-γ↓ FABP4↓ MyoD↓ Pax↓ | Inhibition of muscle adipogenesis | 5–50 μM | [25] | |
| Primary human muscle cell | CEBPA↓ ADIPOQ↓ | Inhibition of muscle adipogenesis | 0.3-3 μM | [26] | |
| BALB/c mice | PPAR-δ↑ CPT1↑ HADH↑ UCP3↑ | Promotion of glycogen storage and fatty-acid oxidation | 0.005% in diet | [27] | |
| Rutin | Litopenaeus vannamei (shrimp) | Enolase↑ malate dehydrogenase↑ ATP/ADP translocase↑ EF-1α↑ elF2α↑ aspartate aminotransferase↑ | Protection of protein deterioration induced by T-2 toxin | 2.00–32.00 g/kg in diet | [28] |
| ICR mice | phosphorylation of AMPK↑ CAMKKβ↑ ACC↑ | Promotion of glucose uptake and prevent hyperglycemia in high-fat diet obese mice | 0.1% in diet | [29,30] | |
| Wistar rats | Calcium uptake↑ CAMKII↑ GLUT-4 translocation↑ | Promotion of glucose uptake by insulin-independent calcium pathway | 10−14 M | [31] | |
| Sprague-Dawley rats | AMPK↑ mtDNA↑ Atrogin-1↓ MuRF1↓ PGC-1α↓ Nrf1↓ Bax↓ Bcl2↓ | Promotion of mitochondrial biogenesis in high-fat diet obese rats and inhibits muscle cell apoptosis | 50 mg/kg/day | [32] | |
| Sprague-Dawley rats | PGC-1α↑ NRF1↑ Tfam↑ SIRT1↑ | Promotion of mitochondrial biogenesis in high-fat diet obese rats | 0.1% in diet | [32] | |
| C57BL/6 mice | MyoD↑ MyoG↑ MHC↑ | Protection of muscle atrophy induced by dexamethasone | 0.1% of extract | [33] | |
| C2C12 cells | IL-6↓ TNFα↓ iNOS↓ ROS↓ NF-κB↓ | Inhibition of inflammation induced by LPS | 10–100 μM | [34] | |
| C2C12 cells | MAFbx↓ MuRF1↓ FOXO3↓ | Protection of muscle atrophy induced by dexamethasone | 100 μM | [35] | |
| VSMC of C57BL/6 mice and chickens and broilers | malondialdehyde↓ ROS↓ 3-nitrotyrosine↓ 4-hydroxynonenal↓ p47phox↓ Nox4↓ | Protection from hydrogenperoxide-induced senescence | 40 mg/kg/d | [36,37,38] | |
| Brioiler chickens | LC3-II↓ PINK1/Parkin↓ | Increased antioxidant activity and inhibited mitophagy | 500 mg/kg | [38] | |
| Sqiss mice | MDA↓ BUN↓ lactate↓ GOT↓ GPT↓ nNOS↓ iNOS↓ TNFα↓ syncytin-1↓ phosphorylation of IKK↓ | Anti-oxidant activity in high-fat diet obese mice, increasing running ability | 100–200 mg/kg/day | [39,40] | |
| Kaempferol glycosides | Wistar rats | PI3K↑ GSK-3↑ MAPK↑ PP1↑ | Promotion of glucose uptake and glycogen synthesis in soleus muscle | 1 μM | [41] |
| L6 skeletal muscle from rat | SIRT1↑ p-IRS↑ AKT↑ AMPK↑, GLUT4 translocation | Enhancement of glucose uptake | 100 nM or 1 μM | [42] | |
| HSMM and DHSMM | CAT↑ MDA↓ | Anti-oxidant activity combined with metformin | 400 µg/mL of extract | [43] | |
| Broiler chickens | TBARS↓ MDA↓ | Inhibition of oxidative stress via reduced MDA concentration | 0.3 or 0.6% in diet | [44] | |
| Wistar rats | Oxidative fibers↓ | Attenuation of muscle atrophy in cerebral palsy rats | 1 mg/kg | [45] | |
| Baicalin | BALB/c mice | Atrogin-1↓ MuRF1↓ IκBα↓ p65↓ NF-κB↓ | Inhibition of muscle atrophy in cachexia mice | 50 or 150 mg/kg | [46] |
| Genkwanin | C2C12 cells | 2-NBDG↑ | Enhancement of glucose uptake | 50 µg/mL extract | [47] |
| C57BL/6 mice | MyHC-II↑ Myh4↑ Foxo1↓ Foxo6↓ IL-1β↓ STAT3↓ p65↓ | Anti-inflammation and inhibition of osteosarcopenia in geriatric mice | 500 mg/kg/d extract | [48] | |
| Isoshaftoside | C2C12 cells | Myf5↑ Myf6↑ MyoD↑ Myogenin↑ TNFα↓ IL-1α↓ IL-1β↓ IL-6↓ | Prevention of muscle degradation and promotes muscle regeneration Induction of mitochondrial biogenesis Reduction of inflammation | 50 μM | [49] |
| Naringin | BALB/c mice | SOD↑ CAT↑ GSH-Px↑ ROS↓ | Reduction of fatigue and enhanced endurance Reduced oxidative stress | 400 mg/kg in diet | [50] |
| Obese rats | mTOR↑ PGC1-α↑ Atrogin-1↓ MuRF1↓ SOD1↑ CAT↑ ROS↓ | Reduction of protein degradation and increased protein synthesis Induction of mitochondrial biogenesis | 50 and 100 mg/kg | [51] | |
| Eriocitrin | denervated mice | Atrogin-1↓ MuRF1↓ | Reduction of oxidative stress and inhibits lipid peroxication | 0.5% in diet | [52] |
| Puerarin | Type 1 diabetic rat | Akt/mTOR↑ PGC-1α↑ AMPK↑ | Enhancement of mitochondrial biogenesis Prevents skeletal muscle wasting | 100 mg/kg/d | [53,54] |
| L6 myotubes | 0.3 mM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, Y.E.; Ju, S.H.; Kim, Y.; Lee, S.-J. Natural Flavonoids for the Prevention of Sarcopenia: Therapeutic Potential and Mechanisms. Int. J. Mol. Sci. 2025, 26, 7458. https://doi.org/10.3390/ijms26157458
Yoon YE, Ju SH, Kim Y, Lee S-J. Natural Flavonoids for the Prevention of Sarcopenia: Therapeutic Potential and Mechanisms. International Journal of Molecular Sciences. 2025; 26(15):7458. https://doi.org/10.3390/ijms26157458
Chicago/Turabian StyleYoon, Ye Eun, Seong Hun Ju, Yebean Kim, and Sung-Joon Lee. 2025. "Natural Flavonoids for the Prevention of Sarcopenia: Therapeutic Potential and Mechanisms" International Journal of Molecular Sciences 26, no. 15: 7458. https://doi.org/10.3390/ijms26157458
APA StyleYoon, Y. E., Ju, S. H., Kim, Y., & Lee, S.-J. (2025). Natural Flavonoids for the Prevention of Sarcopenia: Therapeutic Potential and Mechanisms. International Journal of Molecular Sciences, 26(15), 7458. https://doi.org/10.3390/ijms26157458