
Autobrewery Syndrome and Endogenous Ethanol Production in Patients with MASLD: A Perspective from Chronic Liver Disease

 , ,
, ,  and
and
Abstract
1. Introduction

{kind=link}
{kind=link}
| Author | Age | Sex | Comorbidities | Symptoms | Microorganism | Blood Ethanol Level | Treatment | Outcome |
|---|---|---|---|---|---|---|---|---|
| Kaji et al. [19] | 1. 24 2. 35 | 1. F 2. M | 1. None 2. None | 1. Nausea, faintness 2. Slurred speech, blurred vision, balance problems | 1. C. albicans and C. krusei 2. C. albicans | 1. 254 mg/dL 2. NA | 1.Cabimicina, laxatives, carbohydrates restriction 2.Nystatinum | 1. Symptoms resolution 2. Symptoms resolution |
| Spinucci et al. [13] | 44 | M | Chronic intestinal pseudo-obstruction, on long-term home parenteral nutrition | Abdominal pain, bloating, disorientation, slurred speech after recent antibiotic therapy and simple-sugar based diet | C. albicans (small bowel cultures and stool) S. cerevisiae (small bowel cultures and stool) | 24.9 mmol/L | Sugar-free diet + fluconazole | Symptoms resolution, normalization of ethanolemia |
| Welch et al. [11] | 71 | M | Crohn’s disease, small bowel resection | Slurred speech, dizziness after receiving antibiotic therapy and increasing sugar intake | C. glabrata (small bowel cultures) | 234 mg/dL | Low carbohydrate diet, avoiding antibiotics | Symptoms resolution, normalization of ethanol levels |
| Vandekerckhove et al. [20] | 47 | M | Roux-en-J bypass | Dizziness after receiving antibiotic therapy | Candida glabrata (stool) | 34.7 mmol/L | Low carbohydrate diet + fluconazole nystatin; amphotericin; FMT | Diet and antifungal unsuccessful; FMT symptoms resolution, normalization of ethanol levels and liver enzymes |
| Dahshan and Donovan [21] | 13 | F | Short bowel syndrome in jejunal atresia, 5–6 UA per week | Disorientation, somnolence, bizarre behaviour, fruity odor of breath, especially after meals | C. glabrata (small bowel cultures) S. cerevisiae (small bowel cultures) | 250–350 mg/dL | Antifungal (fluconazole) | Symptoms resolution, normalization of ethanol levels |
| Saverimuttu et al. [22] | 45 | M | Diabetes mellitus type 2, hypertension, hyperlipidemia | Seizures, slurred speech, poor coordination related to meal intake, after receiving antibiotic therapy | S. cerevisiae (stool) C. intermedia (small bowel cultures) K. pneumoniae (small bowel cultures) E. faecalis (small bowel cultures) | 410 mg/dL | Low carbohydrate diet, antifungal (micafungin), probiotics | Symptoms resolution, normalization of ethanol levels |
| Malik et al. [23] | 46 | M | None | Memory loss, mood changes, depression after receiving antibiotic therapy | S. Cerevisiae (stool) S. boulardii (stool) C. Albicans (small bowel cultures) C. parapsilosis (small bowel cultures) | 200 mg/dL at admission, up to 400 mg/dL | Carbohydrate-free diet, multiple antifungal therapy, probiotics | Symptoms resolution |
| Ser et al. [24] | 58 | F | Hemicolectomy, herpetic encephalitis | Recurrent encephalopathy episodes, chronic cognitive disturbances, carbohydrates craving, fruity odor of breath | C. krusei (stool) C. parapsilosis (stool) | 315 mg/dL | Low carbohydrate diet, antifungal (nystatin) | Symptoms resolution |
| Jansson-Nettelbladt et al. [25] | 3 | F | Small bowel malformation | Balance problems | C. kefyr (small bowel cultures and stool) S. cerevisiae (small bowel cultures) | 15 mmol/L | Low carbohydrate diet, antifungal (fluconazole) | Symptoms resolution |
| Kruckenberg et al. [1] | 61 | F | Poorly controlled diabetes mellitus, cirrhosis | None | C. glabrata (urinary) S. cerevisiae (urinary) | Urine ethanol level 32 mg/dL | Antifungal regimen | None improvement of urine ethanol levels |
| Gruszecki et al. [26] | 19 | F | Diabetes mellitus type 1 | Severe diabetic ketoacidosis | C. glabrata (urinary) | Urine ethanol level 0.32 g/dL | NA | Death |
| Cordell et al. [27] | 1. 60 2. 42 3. 32 | 1. M 2. F 3. M | 1. Alcohol abuse; hepatitis C, hypertension, pre-diabetes 2. None 3. None | 1. drunkenness 2. drunkenness, loss of coordination 3. Abdominal pain, reflux, diarrhea, nausea | 1. C. albicans, C. krusei (stool) 2. S. cerevesiae (stool), S. bulardii (stool) 3. None | 1. 170 mg/dL 2. 0.40% on breathalyzer = 400 mg/dL 3. NA | Low-carbohydrate diet Diet NA | Symptoms relief Symptoms relief NA |
| Yuan et al. [17] | NA | NA | NA | NA | K. pneumoniae high-alcohol-producing (HiAlc Kpn) (stool) | ∼400 mg/dL | NA | Oral gavage of HiAlc Kpn and FMT with HiAlc Kpn induced MASLD |
| Akbaba et al. [2] | 38 | M | Recently stopped alcohol abuse, sleep disorders, hypertension | Screening after a car accident | Pseudomonas (small bowel cultures) | 322 mg/dL | NA | NA |
| Yates and Saito [28] | 52 | M | Recent SARS-CoV2 infection | Dizziness, slurred speech, behavior changes | None | 212 mg/dL | Low-carbohydrate diet, probiotics, fluconazole | Symptoms relief |
| Akhavan et al. [29] | 25 | M | Recently undertaken ketogenic diet | Slurred speech, stumbling, dizziness, nausea | None, but resolved after empiric fluconazole therapy | 0.3 g/dL | Empiric fluconazole | Symptoms resolution |
1.1. General Approach to the Autobrewery Syndrome
1.1.1. Endogenous Ethanol Production
1.1.2. ABS Clinical Presentation
2. The Intricacy of the Composition of the Microbiota and Its Metabolic Role
Alterations in the Composition of the Gut Microbiota in Autobrewery Syndrome
3. Alcohol-Mediated Liver Damage and Endogenous Ethanol in Chronic Liver Disease
3.1. Metabolism of Alcohol and Alcohol Mediated Liver Damage
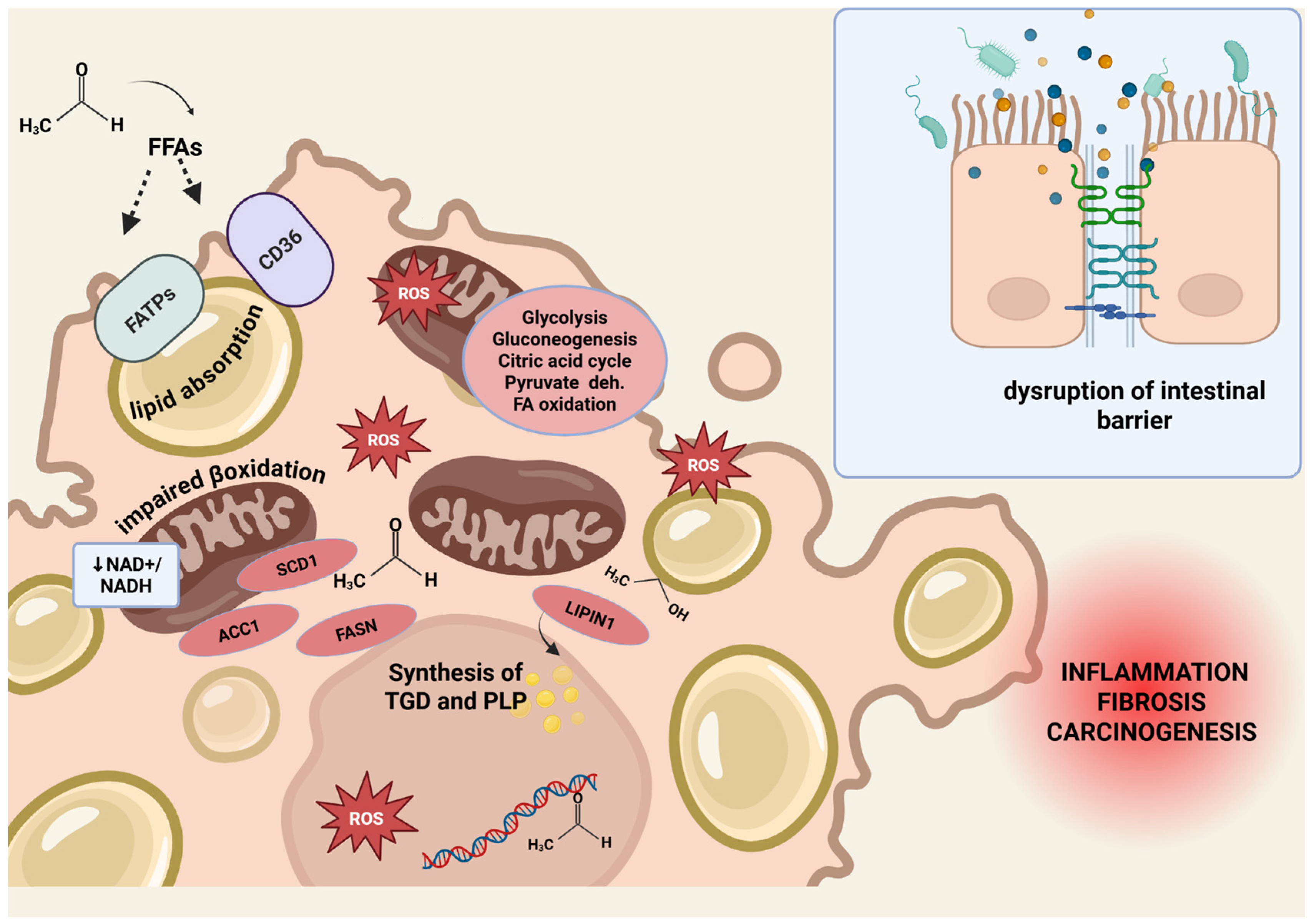
3.1.1. Lipid Accumulation, Mitochondrial Dysfunction and Oxidative Stress
3.1.2. Intestinal Barrier Impairment
3.1.3. Hepatic Stellate Cell Activation and Liver Fibrogenesis
4. Autobrewery Syndrome, MASLD, and Chronic Liver Disease
5. Treatment and Future Perspectives of Autobrewery Syndrome
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABS | autobrewery syndrome |
| ACC1 | acetyl-CoA carboxylase 1 |
| ADH | alcohol dehydrogenase |
| AJs | adherens junctions |
| ALD | alcoholic liver disease |
| ALDH | aldehyde dehydrogenase |
| AMPK | adenosine monophosphate kinase |
| CD36/FAT | fatty acid translocase |
| COL1A2 | collagen type I alpha 2 chain |
| CYP2E1 | cytochrome P450 2E1 |
| CYP2E1 | cytochrome P450 family 2 subfamily E member 1 |
| ECM | extracellular matrix |
| EnEth | endogenous ethanol |
| ERK1/2 | extracellular signal-regulated kinase ½ |
| EtG | ethyl glucuronide |
| EtS | ethyl sulfate |
| FAEEs | fatty acid ethyl esters |
| FASN | fatty acid synthase |
| FATPs | fatty acid transporters |
| FFAs | free fatty acids |
| GMT | fecal microbiota transplantation |
| HSCs | hepatic stellate cells |
| IL-1β | interleukin-1β |
| LPIN | lipin1 protein |
| LPS | lipopolysaccharide |
| MAPK | mitogen-activated protein kinase |
| MASH | metabolic associated steatohepatitis |
| MASLD | metabolic-associated steatotic liver disease |
| MCJ | methylation-controlled J protein |
| MEOS | microsomal ethanol oxidizing system |
| MLCK | myosin light chain kinase |
| MMP-1, MMP-2 | matrix metalloproteinase-1, 2 |
| n-3 PUFA | omega-3 polyunsaturated fatty acid |
| NA | non applicable |
| NAD | nicotinamide adenine dinucleotide |
| NADH | nicotinamide adenine dinucleotide (NAD) + hydrogen (H) |
| NAFLD | non-alcoholic fatty liver disease |
| NF-κB | nuclear factor-kappa B |
| NO | nitric oxide |
| NRF2 | nuclear factor erythroid 2-related factor 2 |
| p70S6K | P70-S6 kinase 1 |
| Peth | phosphatidylethanol |
| PI3K | phosphatidylinositol 3-kinase |
| PPARα | peroxisome proliferator-activated receptor alpha |
| PTPase | protein tyrosine phosphatase |
| Reg3 | regenerating islet-derived protein 3 gamma |
| ROS | reactive oxygen species |
| SCD1 | stearoyl-CoA desaturase 1 |
| SIRT1 | sirtuin 1 |
| SMAD3 | mothers against decapentaplegic homolog 3 |
| SREBP-1c | sterol regulatory element-binding protein 1c |
| TBARS | thiobarbituric acid reactive substance |
| TGF | tumor growth factor |
| TJs | tight junctions |
| TLR/ | toll-like receptor 7 |
| TLR4 | toll-lik receptor4 |
| TNF-α | tumor necrosis factor alpha |
| ZO-1 | zonula occludens 1 |
References
- Kruckenberg, K.M.; DiMartini, A.F.; Rymer, J.A.; Pasculle, A.W.; Tamama, K. Urinary Auto-brewery Syndrome: A Case Report. Ann. Intern. Med. 2020, 172, 702–704. [Google Scholar] [CrossRef]
- Akbaba, M. A medicolegal approach to the very rare Auto-Brewery (endogenous alcohol fermentation) syndrome. Traffic Inj. Prev. 2020, 21, 295–297. [Google Scholar] [CrossRef]
- Bayoumy, A.B.; Mulder, C.J.J.; Mol, J.J.; Tushuizen, M.E. Gut fermentation syndrome: A systematic review of case reports. United Eur. Gastroenterol. J. 2021, 9, 332–342. [Google Scholar] [CrossRef]
- Logan, B.K.; Jones, A.W. Endogenous ethanol “auto-brewery syndrome” as a drunk-driving defence challenge. Med. Sci. Law 2000, 40, 206–215. [Google Scholar] [CrossRef]
- Ladkin, R.G.; Davies, J.N.P. Rupture of the stomach in an African child. Br. Med. J. 1948, 1, 644. [Google Scholar] [CrossRef]
- Dinis-Oliveira, R.J. The Auto-Brewery Syndrome: A Perfect Metabolic “Storm” with Clinical and Forensic Implications. J. Clin. Med. 2021, 10, 4637. [Google Scholar] [CrossRef]
- Al-Awadhi, A.; Wasfi, I.A.; Al Reyami, F.; Al-Hatali, Z. Autobrewing revisited: Endogenous concentrations of blood ethanol in residents of the United Arab Emirates. Sci. Justice 2004, 44, 149–152. [Google Scholar] [CrossRef]
- Ragab, A.R. Endogenous Ethanol Production Levels in Saudi Arabia Residents. J. Alcohol. Drug Depend. 2015, 3, 3. [Google Scholar] [CrossRef]
- Malik, F.; Wickremesinghe, P.; Saleem, A. Auto-Brewery Syndrome: A Schematic for Diagnosis and Appropriate Treatment. Pract. Gastroenterol. 2021, 45, 10–20. [Google Scholar]
- Hafez, E.M.; Hamad, M.A.; Fouad, M.; Abdel-Lateff, A. Auto-brewery syndrome: Ethanol pseudo-toxicity in diabetic and hepatic patients. Hum. Exp. Toxicol. 2017, 36, 445–450. [Google Scholar] [CrossRef]
- Welch, B.T.; Coelho Prabhu, N.; Walkoff, L.; Trenkner, S.W. Auto-brewery Syndrome in the Setting of Long-standing Crohn’s Disease: A Case Report and Review of the Literature. J. Crohns Colitis 2016, 10, 1448–1450. [Google Scholar] [CrossRef]
- Dibaise, J.K.; Young, R.J.; Vanderhoof, J.A. Enteric microbial flora, bacterial overgrowth, and short-bowel syndrome. Clin. Gastroenterol. Hepatol. 2006, 4, 11–20. [Google Scholar] [CrossRef]
- Spinucci, G.; Guidetti, M.; Lanzoni, E.; Pironi, L. Endogenous ethanol production in a patient with chronic intestinal pseudo-obstruction and small intestinal bacterial overgrowth. Eur. J. Gastroenterol. Hepatol. 2006, 18, 799. [Google Scholar] [CrossRef]
- Steinert, R.E.; Rehman, A.; Souto Lima, E.J.; Agamennone, V.; Schuren, F.H.J.; Gero, D.; Schreiner, P.; Vonlanthen, R.; Ismaeil, A.; Tzafos, S.; et al. Roux-en-Y gastric bypass surgery changes fungal and bacterial microbiota in morbidly obese patients—A pilot study. PLoS ONE 2020, 15, e0236936. [Google Scholar] [CrossRef]
- Takahashi, G.; Hoshikawa, K.; Kan, S.; Akimaru, R.; Kodama, Y.; Sato, T.; Kakisaka, K.; Yamada, Y. Auto-brewery syndrome caused by oral fungi and periodontal disease bacteria. Acute Med. Surg. 2021, 8, e652. [Google Scholar] [CrossRef]
- Bivin, W.S.; Heinen, B.N. Production of ethanol from infant food formulas by common yeasts. J. Appl. Bacteriol. 1985, 58, 355–357. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, C.; Cui, J.; Lu, J.; Yan, C.; Wei, X.; Zhao, X.; Li, N.; Li, S.; Xue, G.; et al. Fatty Liver Disease Caused by High-Alcohol-Producing Klebsiella pneumoniae. Cell Metab. 2019, 30, 675–688.e7. [Google Scholar] [CrossRef]
- Dawes, E.A.; Foster, S.M. The formation of ethanol in Escherichia coli. Biochim. Biophys. Acta 1956, 22, 253–265. [Google Scholar] [CrossRef]
- Kaji, H.; Asanuma, Y.; Yahara, O.; Shibue, H.; Hisamura, M.; Saito, N.; Kawakami, Y.; Murao, M. Intragastrointestinal alcohol fermentation syndrome: Report of two cases and review of the literature. J. Forensic. Sci. Soc. 1984, 24, 461–471. [Google Scholar] [CrossRef]
- Vandekerckhove, E.; Janssens, F.; Tate, D.; De Looze, D. Treatment of Gut Fermentation Syndrome with Fecal Microbiota Transplantation. Ann. Intern. Med. 2020, 173, 855. [Google Scholar] [CrossRef]
- Dahshan, A.; Donovan, K. Auto-brewery syndrome in a child with short gut syndrome: Case report and review of the literature. J. Pediatr. Gastroenterol. Nutr. 2001, 33, 214–215. [Google Scholar] [CrossRef]
- Saverimuttu, J.; Malik, F.; Arulthasan, M.; Wickremesinghe, P. A Case of Auto-brewery Syndrome Treated with Micafungin. Cureus 2019, 11, e5904. [Google Scholar] [CrossRef]
- Malik, F.; Wickremesinghe, P.; Saverimuttu, J. Case report and literature review of auto-brewery syndrome: Probably an underdiagnosed medical condition. BMJ Open Gastroenterol. 2019, 6, e000325. [Google Scholar] [CrossRef]
- Ser, M.H.; Çalıkuşu, F.Z.; Erener, N.; Destanoğlu, O.; Kıykım, E.; Siva, A. Auto brewery syndrome from the perspective of the neurologist. J. Forensic Leg. Med. 2023, 96, 102514. [Google Scholar] [CrossRef]
- Jansson-Nettelbladt, E.; Meurling, S.; Petrini, B.; Sjölin, J. Endogenous ethanol fermentation in a child with short bowel syndrome. Acta Paediatr. 2006, 95, 502–504. [Google Scholar] [CrossRef]
- Gruszecki, A.C.; Robinson, C.A.; Kloda, S.; Brissie, R.M. High urine ethanol and negative blood and vitreous ethanol in a diabetic woman: A case report, retrospective case survey, and review of the literature. Am. J. Forensic Med. Pathol. 2005, 26, 96–98. [Google Scholar] [CrossRef]
- Cordell, B.; Kanodia, A. Auto-Brewery as an Emerging Syndrome: Three Representative Case Studies. J. Clin. Med. Case Rep. 2015, 2, 5. [Google Scholar]
- Yates, S.R.; Saito, A. Auto-Brewery Syndrome After COVID-19 Infection. ACG Case Rep. J. 2024, 11, e01248. [Google Scholar] [CrossRef]
- Akhavan, B.J.; Ostrosky-Zeichner, L.; Thomas, E.J. Drunk Without Drinking: A Case of Auto-Brewery Syndrome. ACG Case Rep. J. 2019, 6, e00208. [Google Scholar] [CrossRef]
- Sarkola, T.; Eriksson, C.J. Effect of 4-methylpyrazole on endogenous plasma ethanol and methanol levels in humans. Alcohol. Clin. Exp. Res. 2001, 25, 513–516. [Google Scholar]
- Watanabe-Suzuki, K.; Seno, H.; Ishii, A.; Kumazawa, T.; Suzuki, O. Ultra-sensitive method for determination of ethanol in whole blood by headspace capillary gas chromatography with cryogenic oven trapping. J. Chromatogr. B Biomed. Sci. Appl. 1999, 727, 89–94. [Google Scholar] [CrossRef]
- Xue, G.; Feng, J.; Zhang, R.; Du, B.; Sun, Y.; Liu, S.; Yan, C.; Liu, X.; Du, S.; Feng, Y.; et al. Three Klebsiella species as potential pathobionts generating endogenous ethanol in a clinical cohort of patients with auto-brewery syndrome: A case control study. eBioMedicine 2023, 91, 104560. [Google Scholar] [CrossRef]
- Blomstrand, R. Observations of the formation of ethanol in the intestinal tract in man. Life Sci. 1971, 10, 575–582. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.F.; Bockelmann, W.; Meske, D.; de Vrese, M.; Walte, H.-G.; Schrezenmeir, J.; Heller, K.J. Ethanol Production by Selected Intestinal Microorganisms and Lactic Acid Bacteria Growing under Different Nutritional Conditions. Front. Microbiol. 2016, 7, 47. [Google Scholar] [CrossRef]
- Jokelainen, K.; Matysiak-Budnik, T.; Mäkisalo, H.; Höckerstedt, K.; Salaspuro, M. High intracolonic acetaldehyde values produced by a bacteriocolonic pathway for ethanol oxidation in piglets. Gut 1996, 39, 100–104. [Google Scholar] [CrossRef]
- Nosova, T.; Jokelainen, K.; Kaihovaara, P.; Jousimies-Somer, H.; Siitonen, A.; Heine, R.; Salaspuro, M. Aldehyde Dehydrogenase Activity and Acetate Production by Aerobic Bacteria Representing the Normal Flora of Human Large Intestine. Alcohol Alcohol. 1996, 31, 555–564. [Google Scholar] [CrossRef]
- Logan, B.K.; Jones, A.W. Endogenous Ethanol Production in a Child with Short Gut Syndrome. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 419–420. [Google Scholar] [CrossRef]
- Holford, N.H. Clinical pharmacokinetics of ethanol. Clin. Pharmacokinet. 1987, 13, 273–292. [Google Scholar] [CrossRef]
- Cederbaum, A.I. Alcohol Metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef]
- Madrid, A.M.; Hurtado, C.; Gatica, S.; Chacón, I.; Toyos, A.; Defilippi, C. Endogenous ethanol production in patients with liver cirrhosis, motor alteration and bacterial overgrowth. Revista Medica de Chile 2002, 130, 1329–1334. [Google Scholar]
- Tameez Ud Din, A.; Alam, F.; Tameez-Ud-Din, A.; Chaudhary, F.M.D. Auto-Brewery Syndrome: A Clinical Dilemma. Cureus 2020, 12, e10983. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e16. [Google Scholar] [CrossRef]
- Vilela, S.F.; O Barbosa, J.; Rossoni, R.D.; Santos, J.D.; Prata, M.C.; Anbinder, A.L.; Jorge, A.O.; Junqueira, J.C. Lactobacillus acidophilus ATCC 4356 inhibits biofilm formation by C. albicans and attenuates the experimental candidiasis in Galleria mellonella. Virulence 2015, 6, 29–39. [Google Scholar] [CrossRef]
- Bakker, G.J.; Nieuwdorp, M. Fecal Microbiota Transplantation: Therapeutic Potential for a Multitude of Diseases beyond Clostridium difficile. Microbiol. Spectr. 2017, 5, 4. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Sendid, B.; Hoarau, G.; Colombel, J.F.; Poulain, D.; Ghannoum, M.A. Mycobiota in gastrointestinal diseases. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 77–87. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, 2. [Google Scholar] [CrossRef]
- Pasolli, E.; Asnicar, F.; Manara, S.; Zolfo, M.; Karcher, N.; Armanini, F.; Beghini, F.; Manghi, P.; Tett, A.; Ghensi, P.; et al. Extensive Unexplored Human Microbiome Diversity Revealed by Over 150,000 Genomes from Metagenomes Spanning Age, Geography, and Lifestyle. Cell 2019, 176, 649–662.e20. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Metabolic Syndrome and Altered Gut Microbiota in Mice Lacking Toll-Like Receptor 5|Science. Available online: https://www.science.org/doi/10.1126/science.1179721 (accessed on 2 May 2025).
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandonà, A.; Paci, P.; Capuani, G.; et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017, 65, 451. [Google Scholar] [CrossRef]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Boursier, J.; Diehl, A.M. Nonalcoholic Fatty Liver Disease and the Gut Microbiome. Clin. Liver Dis. 2016, 20, 263–275. [Google Scholar] [CrossRef]
- Jangi, S.; Hsia, K.; Zhao, N.; Kumamoto, C.A.; Friedman, S.; Singh, S.; Michaud, D.S. Dynamics of the Gut Mycobiome in Patients with Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2024, 22, 821–830.e7. [Google Scholar] [CrossRef]
- Joossens, M.; Huys, G.; Cnockaert, M.; de Preter, V.; Verbeke, K.; Rutgeerts, P.; Vandamme, P.; Vermeire, S. Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut 2011, 60, 631–637. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, J.; Feng, J.; Liu, X.; Hu, Q. The gut microbiota-brain connection: Insights into major depressive disorder and bipolar disorder. Front. Psychiatry 2024, 15, 1421490. [Google Scholar] [CrossRef]
- Kanj, A.N.; Skalski, J.H. Gut Mycobiome and Asthma. J. Fungi 2024, 10, 192. [Google Scholar] [CrossRef]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Wang, L.J.; Li, S.C.; Yeh, Y.M.; Lee, S.Y.; Kuo, H.C.; Yang, C.Y. Gut mycobiome dysbiosis and its impact on intestinal permeability in attention-deficit/hyperactivity disorder. J. Child Psychol. Psychiatry 2023, 64, 1280–1291. [Google Scholar] [CrossRef]
- Ma, W.; Sun, H.; Lian, L.; Guo, L.; Wang, Y.; Huang, L. Immunomodulatory effects of Lactiplantibacillus plantarum CCFM8661 + stachyose on cyclophosphamide-induced immunosuppression mice. Front. Immunol. 2025, 16, 1513531. [Google Scholar] [CrossRef]
- Painter, K.; Cordell, B.J.; Sticco, K.L. Auto-Brewery Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. Available online: http://www.ncbi.nlm.nih.gov/books/NBK513346/ (accessed on 2 May 2025).
- Dashko, S.; Zhou, N.; Compagno, C.; Piškur, J. Why, when, and how did yeast evolve alcoholic fermentation? FEMS Yeast Res. 2014, 14, 826–832. [Google Scholar] [CrossRef]
- Pfeiffer, T.; Morley, A. An evolutionary perspective on the Crabtree effect. Front. Mol. Biosci. 2014, 1, 17. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Power, R. Relationship between pH and medium dissolved solids in terms of growth and metabolism of lactobacilli and Saccharomyces cerevisiae during ethanol production. Appl. Environ. Microbiol. 2005, 71, 2239–2243. [Google Scholar] [CrossRef]
- Sherrington, S.L.; Sorsby, E.; Mahtey, N.; Kumwenda, P.; Lenardon, M.D.; Brown, I.; Ballou, E.R.; MacCallum, D.M.; Hall, R.A.; Noverr, M.C. Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition. PLoS Pathog. 2017, 13, e1006403. [Google Scholar] [CrossRef]
- Bode, J.C.; Rust, S.; Bode, C. The Effect of Cimetidine Treatment on Ethanol Formation in the Human Stomach. Scand. J. Gastroenterol. 1984, 19, 853–856. [Google Scholar] [CrossRef]
- Gan, L.; Feng, Y.; Du, B.; Fu, H.; Tian, Z.; Xue, G.; Yan, C.; Cui, X.; Zhang, R.; Cui, J.; et al. Bacteriophage targeting microbiota alleviates non-alcoholic fatty liver disease induced by high alcohol-producing Klebsiella pneumoniae. Nat. Commun. 2023, 14, 3215. [Google Scholar] [CrossRef]
- Li, N.-N.; Li, W.; Feng, J.-X.; Du, B.; Zhang, R.; Du, S.-H.; Liu, S.-Y.; Xue, G.-H.; Yan, C.; Cui, J.-H.; et al. High alcohol-producing Klebsiella pneumoniae causes fatty liver disease through 2,3-butanediol fermentation pathway in vivo. Gut Microbes 2021, 13, 1979883. [Google Scholar] [CrossRef]
- Cordell, B.J. Shifting microbiomes: Pathobionts hiding in our guts. eBioMedicine 2023, 92, 104592. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients with COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Smędra, A.; Trzmielak, M.; Góralska, K.; Dzikowiec, M.; Brzeziańska-Lasota, E.; Berent, J. Oral form of auto-brewery syndrome. J. Forensic Leg. Med. 2022, 87, 102333. [Google Scholar] [CrossRef]
- Smędra, A.; Berent, J. The Influence of the Oral Microbiome on Oral Cancer: A Literature Review and a New Approach. Biomolecules 2023, 13, 815. [Google Scholar] [CrossRef]
- Seitz, H.K.; Stickel, F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nat. Rev. Cancer 2007, 7, 599–612. [Google Scholar] [CrossRef]
- Heier, C.; Xie, H.; Zimmermann, R. Nonoxidative ethanol metabolism in humans-from biomarkers to bioactive lipids. IUBMB Life 2016, 68, 916–923. [Google Scholar] [CrossRef]
- Laposata, E.A.; Lange, L.G. Presence of nonoxidative ethanol metabolism in human organs commonly damaged by ethanol abuse. Science 1986, 231, 497–499. [Google Scholar] [CrossRef]
- Gordon, E.R. The effect of chronic consumption of ethanol on the redox state of the rat liver. Can. J. Biochem. 1972, 50, 949–957. [Google Scholar] [CrossRef]
- Veech, R.L.; Guynn, R.; Veloso, D. The time-course of the effects of ethanol on the redox and phosphorylation states of rat liver. Biochem. J. 1972, 127, 387–397. [Google Scholar] [CrossRef]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological Aspects of Alcohol Metabolism in the Liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef]
- Doege, H.; Baillie, R.A.; Ortegon, A.M.; Tsang, B.; Wu, Q.; Punreddy, S.; Hirsch, D.; Watson, N.; Gimeno, R.E.; Stahl, A. Targeted deletion of FATP5 reveals multiple functions in liver metabolism: Alterations in hepatic lipid homeostasis. Gastroenterology 2006, 130, 1245–1258. [Google Scholar] [CrossRef]
- Wilson, C.G.; Tran, J.L.; Erion, D.M.; Vera, N.B.; Febbraio, M.; Weiss, E.J. Hepatocyte-Specific Disruption of CD36 Attenuates Fatty Liver and Improves Insulin Sensitivity in HFD-Fed Mice. Endocrinology 2016, 157, 570–585. [Google Scholar] [CrossRef]
- You, M.; Matsumoto, M.; Pacold, C.M.; Cho, W.K.; Crabb, D.W. The role of AMP-activated protein kinase in the action of ethanol in the liver. Gastroenterology 2004, 127, 1798–1808. [Google Scholar] [CrossRef]
- Marmier, S.; Dentin, R.; Daujat-Chavanieu, M.; Guillou, H.; Bertrand-Michel, J.; Gerbal-Chaloin, S.; Girard, J.; Lotersztajn, S.; Postic, C. Novel role for carbohydrate responsive element binding protein in the control of ethanol metabolism and susceptibility to binge drinking. Hepatology 2015, 62, 1086–1100. [Google Scholar] [CrossRef]
- Bi, L.; Jiang, Z.; Zhou, J. The role of lipin-1 in the pathogenesis of alcoholic fatty liver. Alcohol Alcohol. 2015, 50, 146–151. [Google Scholar] [CrossRef]
- Jeon, S.; Carr, R. Alcohol effects on hepatic lipid metabolism. J. Lipid Res. 2020, 61, 470–479. [Google Scholar] [CrossRef]
- Yin, H.; Hu, M.; Zhang, R.; Shen, Z.; Flatow, L.; You, M. MicroRNA-217 promotes ethanol-induced fat accumulation in hepatocytes by down-regulating SIRT1. J. Biol. Chem. 2012, 287, 9817–9826. [Google Scholar] [CrossRef]
- Kersten, S. Integrated physiology and systems biology of PPARα. Mol. Metab. 2014, 3, 354–371. [Google Scholar] [CrossRef]
- Fischer, M.; You, M.; Matsumoto, M.; Crabb, D.W. Peroxisome proliferator-activated receptor alpha (PPARalpha) agonist treatment reverses PPARalpha dysfunction and abnormalities in hepatic lipid metabolism in ethanol-fed mice. J. Biol. Chem. 2003, 278, 27997–28004. [Google Scholar] [CrossRef]
- Malaguarnera, G.; Giordano, M.; Nunnari, G.; Bertino, G.; Malaguarnera, M. Gut microbiota in alcoholic liver disease: Pathogenetic role and therapeutic perspectives. World J. Gastroenterol. 2014, 20, 16639–16648. [Google Scholar] [CrossRef]
- Zhong, Z.; Ramshesh, V.K.; Rehman, H.; Liu, Q.; Theruvath, T.P.; Krishnasamy, Y.; Lemasters, J.J.; Choi, J. Acute ethanol causes hepatic mitochondrial depolarization in mice: Role of ethanol metabolism. PLoS ONE 2014, 9, e91308. [Google Scholar] [CrossRef]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef]
- Purohit, V.; Bode, J.C.; Bode, C.; Brenner, D.A.; Choudhry, M.A.; Hamilton, F.; Kang, Y.J.; Keshavarzian, A.; Rao, R.; Sartor, R.B.; et al. Alcohol, intestinal bacterial growth, intestinal permeability to endotoxin, and medical consequences: Summary of a symposium. Alcohol 2008, 42, 349–361. [Google Scholar] [CrossRef]
- Basuroy, S.; Sheth, P.; Mansbach, C.M.; Rao, R.K. Acetaldehyde disrupts tight junctions and adherens junctions in human colonic mucosa: Protection by EGF and L-glutamine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G367–G375. [Google Scholar] [CrossRef]
- Sheth, P.; Seth, A.; Atkinson, K.J.; Gheyi, T.; Kale, G.; Giorgianni, F.; Desiderio, D.M.; Li, C.; Naren, A.; Rao, R. Acetaldehyde dissociates the PTP1B-E-cadherin-beta-catenin complex in Caco-2 cell monolayers by a phosphorylation-dependent mechanism. Biochem. J. 2007, 402, 291–300. [Google Scholar] [CrossRef]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef]
- Ma, T.Y.; Nguyen, D.; Bui, V.; Nguyen, H.; Hoa, N. Ethanol modulation of intestinal epithelial tight junction barrier. Am. J. Physiol. 1999, 276, G965–G974. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Farhadi, A.; Forsyth, C.B.; Rangan, J.; Jakate, S.; Shaikh, M.; Banan, A.; Fields, J.Z. Evidence that chronic alcohol exposure promotes intestinal oxidative stress, intestinal hyperpermeability and endotoxemia prior to development of alcoholic steatohepatitis in rats. J. Hepatol. 2009, 50, 538–547. [Google Scholar] [CrossRef]
- Duddempudi, A.T. Immunology in alcoholic liver disease. Clin. Liver Dis. 2012, 16, 687–698. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol Res. Curr. Rev. 2017, 38, 147–161. [Google Scholar]
- Guo, J.; Friedman, S.L. Toll-like receptor 4 signaling in liver injury and hepatic fibrogenesis. Fibrogenesis Tissue Repair 2010, 3, 21. [Google Scholar] [CrossRef]
- Jia, L.; Chang, X.; Qian, S.; Liu, C.; Lord, C.C.; Ahmed, N.; Lee, C.E.; Lee, S.; Gautron, L.; Mitchell, M.C.; et al. Hepatocyte toll-like receptor 4 deficiency protects against alcohol-induced fatty liver disease. Mol. Metab. 2018, 14, 121–129. [Google Scholar] [CrossRef]
- Lee, J.Y.; Ye, J.; Gao, Z.; Youn, H.S.; Lee, W.H.; Zhao, L.; Sizemore, N.; Hwang, D.H. Reciprocal modulation of Toll-like receptor-4 signaling pathways involving MyD88 and phosphatidylinositol 3-kinase/AKT by saturated and polyunsaturated fatty acids. J. Biol. Chem. 2003, 278, 37041–37051. [Google Scholar] [CrossRef]
- Moshage, H.; Casini, A.; Lieber, C.S. Acetaldehyde selectively stimulates collagen production in cultured rat liver fat-storing cells but not in hepatocytes. Hepatology 1990, 12 Pt 1, 511–518. [Google Scholar] [CrossRef]
- Casini, A.; Cunningham, M.; Rojkind, M.; Lieber, C.S. Acetaldehyde increases procollagen type I and fibronectin gene transcription in cultured rat fat-storing cells through a protein synthesis-dependent mechanism. Hepatology 1991, 13, 758–765. [Google Scholar]
- Svegliati-Baroni, G.; Ridolfi, F.; Di Sario, A.; Saccomanno, S.; Bendia, E.; Benedetti, A.; Greenwel, P. Intracellular signaling pathways involved in acetaldehyde-induced collagen and fibronectin gene expression in human hepatic stellate cells. Hepatology 2001, 33, 1130–1140. [Google Scholar] [CrossRef]
- Casini, A.; Ceni, E.; Salzano, R.; Schuppan, D.; Milani, S.; Pellegrini, G.; Surrenti, C. Regulation of Undulin Synthesis and Gene Expression in Human Fat-Storing Cells by Acetaldehyde and Transforming Growth Factor-β1: Comparison with Fibronectin. Biochem. Biophys. Res. Commun. 1994, 199, 1019–1026. [Google Scholar] [CrossRef]
- Casini, A.; Galli, G.; Salzano, R.; Ceni, E.; Franceschelli, F.; Rotella, C.M.; Surrenti, C. Acetaldehyde induces c-fos and c-jun proto-oncogenes in fat-storing cell cultures through protein kinase C activation. Alcohol Alcohol. 1994, 29, 303–314. [Google Scholar]
- Anania, F.A.; Womack, L.; Potter, J.J.; Mezey, E. Acetaldehyde enhances murine alpha2(I) collagen promoter activity by Ca2+-independent protein kinase C activation in cultured rat hepatic stellate cells. Alcohol Clin. Exp. Res. 1999, 23, 279–284. [Google Scholar]
- Bardag-Gorce, F.; Yuan, Q.X.; Li, J.; French, B.A.; Fang, C.; Ingelman-Sundberg, M.; French, S.W. The effect of ethanol-induced cytochrome p4502E1 on the inhibition of proteasome activity by alcohol. Biochem. Biophys. Res. Commun. 2000, 279, 23–29. [Google Scholar] [CrossRef]
- Butura, A.; Nilsson, K.; Morgan, K.; Morgan, T.R.; French, S.W.; Johansson, I.; Schuppe-Koistinen, I.; Ingelman-Sundberg, M. The impact of CYP2E1 on the development of alcoholic liver disease as studied in a transgenic mouse model. J. Hepatol. 2009, 50, 572–583. [Google Scholar] [CrossRef]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Volynets, V.; Küper, M.A.; Strahl, S.; Maier, I.B.; Spruss, A.; Wagnerberger, S.; Königsrainer, A.; Bischoff, S.C.; Bergheim, I. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD). Dig. Dis. Sci. 2012, 57, 1932–1941. [Google Scholar] [CrossRef]
- Cope, K.; Risby, T.; Diehl, A.M. Increased gastrointestinal ethanol production in obese mice: Implications for fatty liver disease pathogenesis. Gastroenterology 2000, 119, 1340–1347. [Google Scholar] [CrossRef]
- Baker, S.S.; Baker, R.D.; Liu, W.; Nowak, N.J.; Zhu, L. Role of alcohol metabolism in non-alcoholic steatohepatitis. PLoS ONE 2010, 5, e9570. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Davids, M.; Herrema, H.; Aydin, O.; Tremaroli, V.; Rios-Morales, M.; Levels, H.; Bruin, S.; de Brauw, M.; Verheij, J.; et al. Microbiome-derived ethanol in nonalcoholic fatty liver disease. Nat. Med. 2022, 28, 2100–2106. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Z.; Li, H.; Zhao, J.; Wei, X.; Lin, W.; Zhao, X.; Jiang, A.; Yuan, J. Endogenous ethanol produced by intestinal bacteria induces mitochondrial dysfunction in non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2020, 35, 2009–2019. [Google Scholar] [CrossRef]
- Goikoetxea-Usandizaga, N.; Bravo, M.; Egia-Mendikute, L.; Abecia, L.; Serrano-Maciá, M.; Urdinguio, R.G.; Clos-García, M.; Rodríguez-Agudo, R.; Araujo-Legido, R.; López-Bermudo, L.; et al. The outcome of boosting mitochondrial activity in alcohol-associated liver disease is organ-dependent. Hepatology 2023, 78, 878–895. [Google Scholar] [CrossRef]
- Wang, Q.; Kim, S.Y.; Matsushita, H.; Wang, Z.; Pandyarajan, V.; Matsuda, M.; Ohashi, K.; Tsuchiya, T.; Roh, Y.S.; Kiani, C.; et al. Oral administration of PEGylated TLR7 ligand ameliorates alcohol-associated liver disease via the induction of IL-22. Proc. Natl. Acad. Sci. USA 2021, 118, e2020868118. [Google Scholar] [CrossRef]
- Warner, D.R.; Warner, J.B.; Hardesty, J.E.; Song, Y.L.; Chen, C.; Chen, Z.; Kang, J.X.; McClain, C.J.; Kirpich, I.A. Beneficial effects of an endogenous enrichment in n3-PUFAs on Wnt signaling are associated with attenuation of alcohol-mediated liver disease in mice. FASEB J. 2021, 35, e21377. [Google Scholar] [CrossRef]
- Gu, Z.; Li, F.; Liu, Y.; Jiang, M.; Zhang, L.; He, L.; Wilkey, D.W.; Merchant, M.; Zhang, X.; Deng, Z.-B.; et al. Exosome-Like Nanoparticles From Lactobacillus rhamnosusGG Protect Against Alcohol-Associated Liver Disease Through Intestinal Aryl Hydrocarbon Receptor in Mice. Hepatol. Commun. 2021, 5, 846–864. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Ng, S.C.; Schnabl, B. Promises of microbiome-based therapies. J. Hepatol. 2022, 76, 1379–1391. [Google Scholar] [CrossRef]
- Ferrere, G.; Wrzosek, L.; Cailleux, F.; Turpin, W.; Puchois, V.; Spatz, M.; Ciocan, D.; Rainteau, D.; Humbert, L.; Hugot, C.; et al. Fecal microbiota manipulation prevents dysbiosis and alcohol-induced liver injury in mice. J. Hepatol. 2017, 66, 806–815. [Google Scholar] [CrossRef]
- Philips, C.A.; Pande, A.; Shasthry, S.M.; Jamwal, K.D.; Khillan, V.; Chandel, S.S.; Kumar, G.; Sharma, M.K.; Maiwall, R.; Jindal, A.; et al. Healthy Donor Fecal Microbiota Transplantation in Steroid-Ineligible Severe Alcoholic Hepatitis: A Pilot Study. Clin. Gastroenterol. Hepatol. 2017, 15, 600–602. [Google Scholar] [CrossRef]
- Craven, L.; Rahman, A.; Nair Parvathy, S.; Beaton, M.; Silverman, J.; Qumosani, K.; Hramiak, I.; Hegele, R.; Joy, T.; Meddings, J.; et al. Allogenic Fecal Microbiota Transplantation in Patients with Nonalcoholic Fatty Liver Disease Improves Abnormal Small Intestinal Permeability: A Randomized Control Trial. Am. J. Gastroenterol. 2020, 115, 1055–1065. [Google Scholar] [CrossRef]
- Zhou, D.; Pan, Q.; Shen, F.; Cao, H.-X.; Ding, W.-J.; Chen, Y.-W.; Fan, J.-G. Total fecal microbiota transplantation alleviates high-fat diet-induced steatohepatitis in mice via beneficial regulation of gut microbiota. Sci. Rep. 2017, 7, 1529. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andaloro, S.; De Gaetano, V.; Cardone, F.; Ianiro, G.; Cerrito, L.; Pallozzi, M.; Stella, L.; Gasbarrini, A.; Ponziani, F.R. Autobrewery Syndrome and Endogenous Ethanol Production in Patients with MASLD: A Perspective from Chronic Liver Disease. Int. J. Mol. Sci. 2025, 26, 7345. https://doi.org/10.3390/ijms26157345
Andaloro S, De Gaetano V, Cardone F, Ianiro G, Cerrito L, Pallozzi M, Stella L, Gasbarrini A, Ponziani FR. Autobrewery Syndrome and Endogenous Ethanol Production in Patients with MASLD: A Perspective from Chronic Liver Disease. International Journal of Molecular Sciences. 2025; 26(15):7345. https://doi.org/10.3390/ijms26157345
Chicago/Turabian StyleAndaloro, Silvia, Valeria De Gaetano, Ferdinando Cardone, Gianluca Ianiro, Lucia Cerrito, Maria Pallozzi, Leonardo Stella, Antonio Gasbarrini, and Francesca Romana Ponziani. 2025. "Autobrewery Syndrome and Endogenous Ethanol Production in Patients with MASLD: A Perspective from Chronic Liver Disease" International Journal of Molecular Sciences 26, no. 15: 7345. https://doi.org/10.3390/ijms26157345
APA StyleAndaloro, S., De Gaetano, V., Cardone, F., Ianiro, G., Cerrito, L., Pallozzi, M., Stella, L., Gasbarrini, A., & Ponziani, F. R. (2025). Autobrewery Syndrome and Endogenous Ethanol Production in Patients with MASLD: A Perspective from Chronic Liver Disease. International Journal of Molecular Sciences, 26(15), 7345. https://doi.org/10.3390/ijms26157345