
Preliminary Study on the Geochemical Characterization of Viticis Fructus Cuticular Waxes: From Latitudinal Variation to Origin Authentication
 , , ,
, , ,  and
and 
Abstract

1. Introduction
2. Results and Discussion
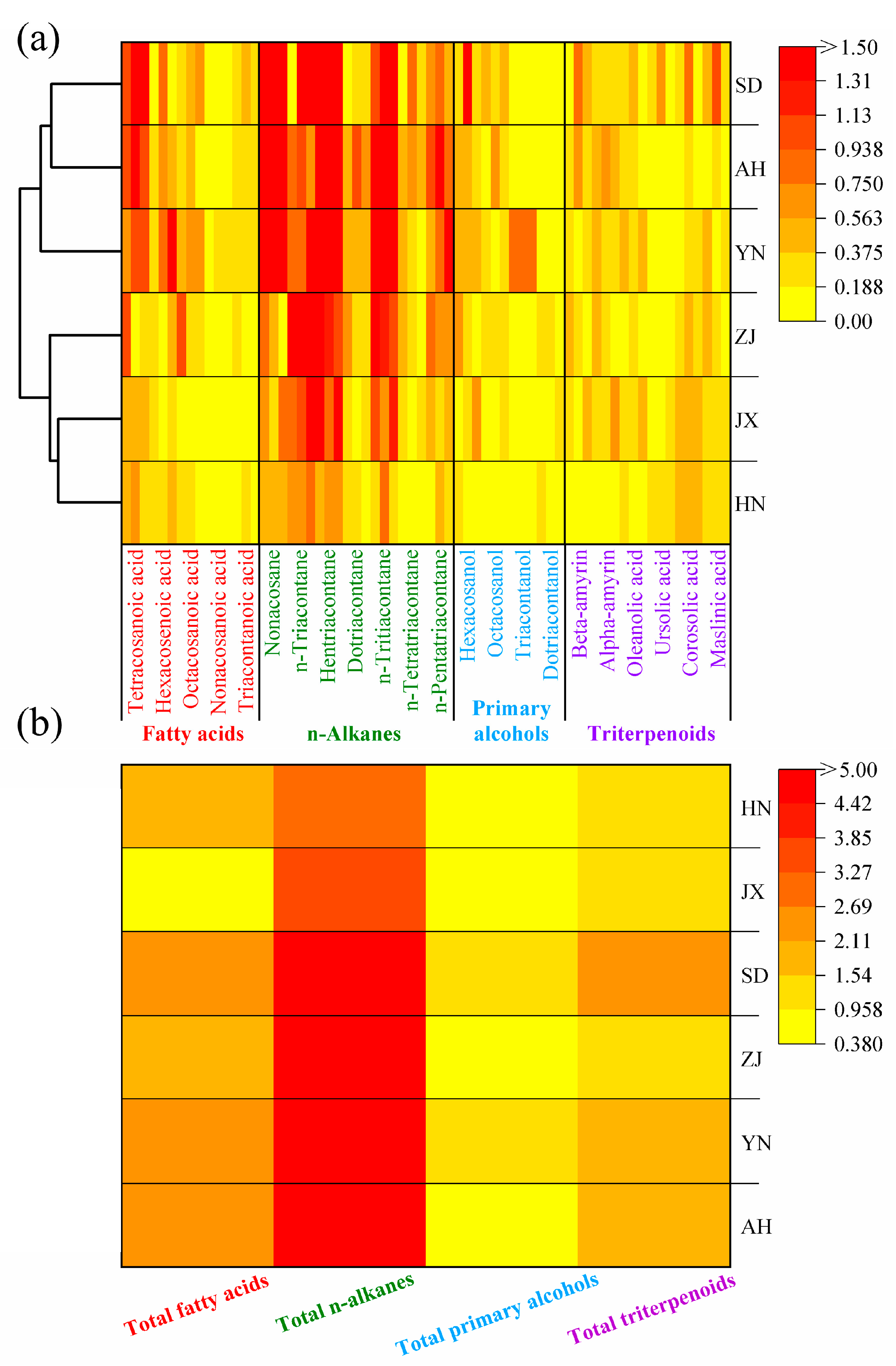
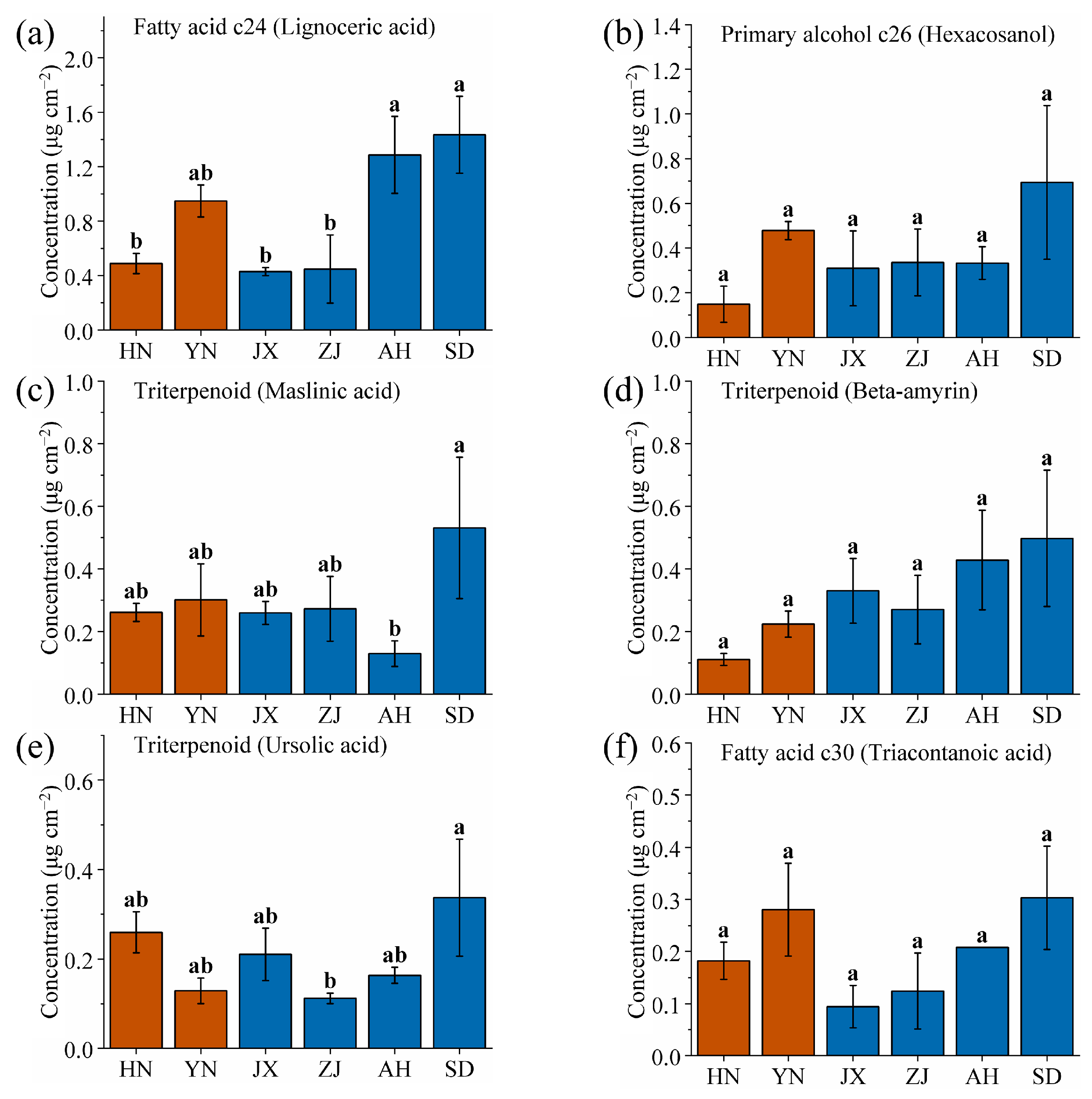
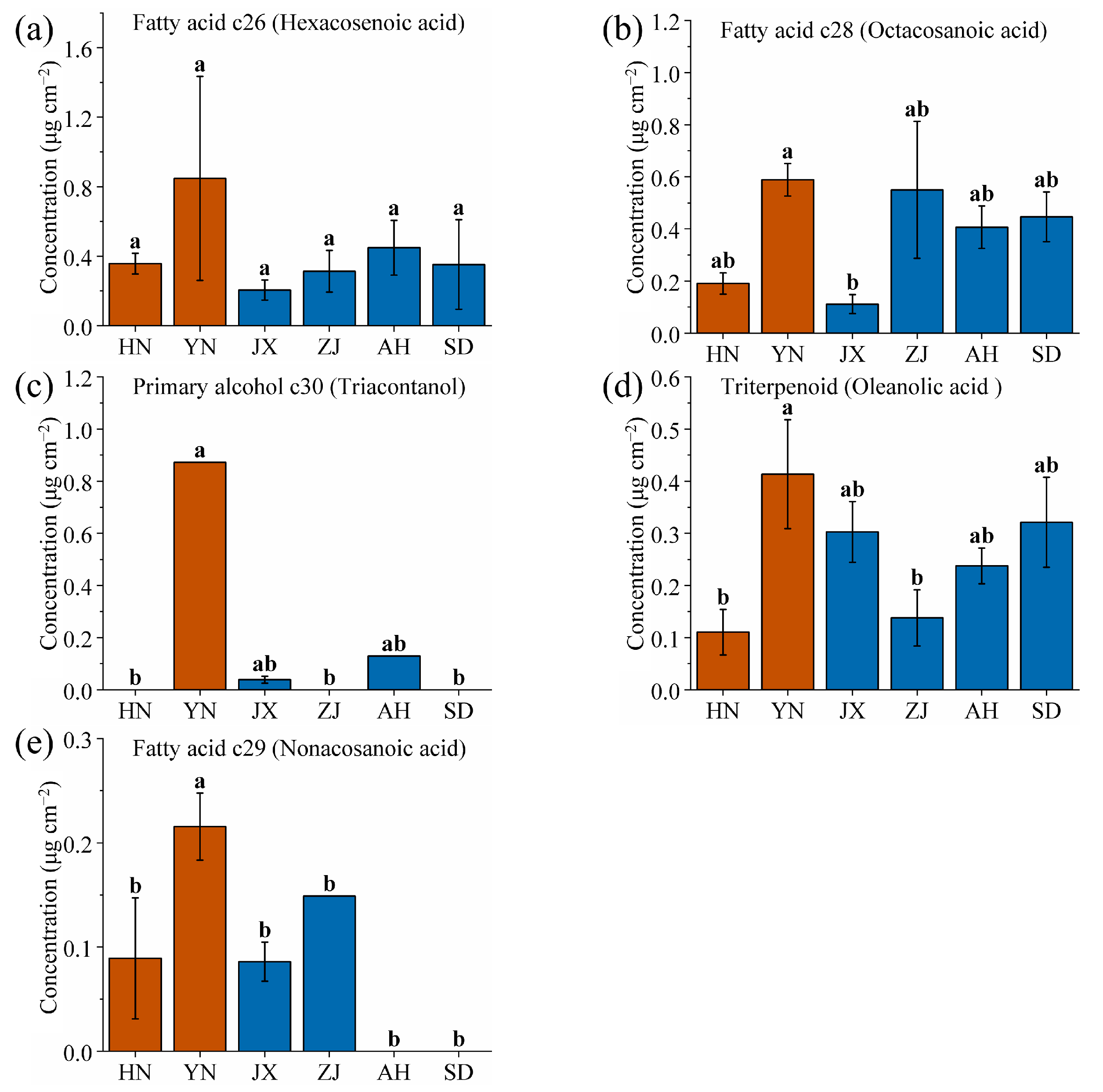
2.1. Chemical Composition and Content Analysis of VF Cuticular Waxes
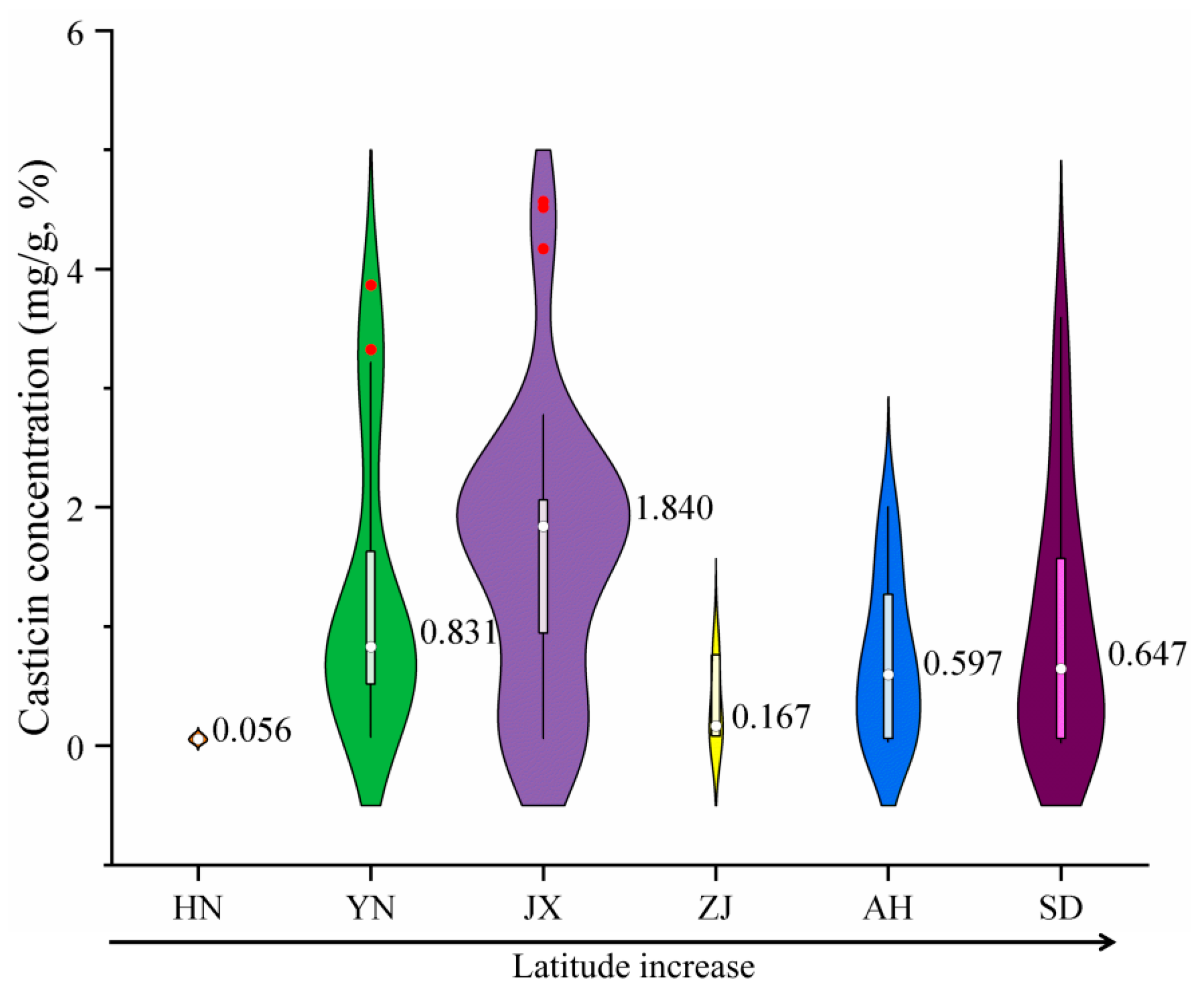
2.2. Special Production Area Priority Identification and Screening of VF
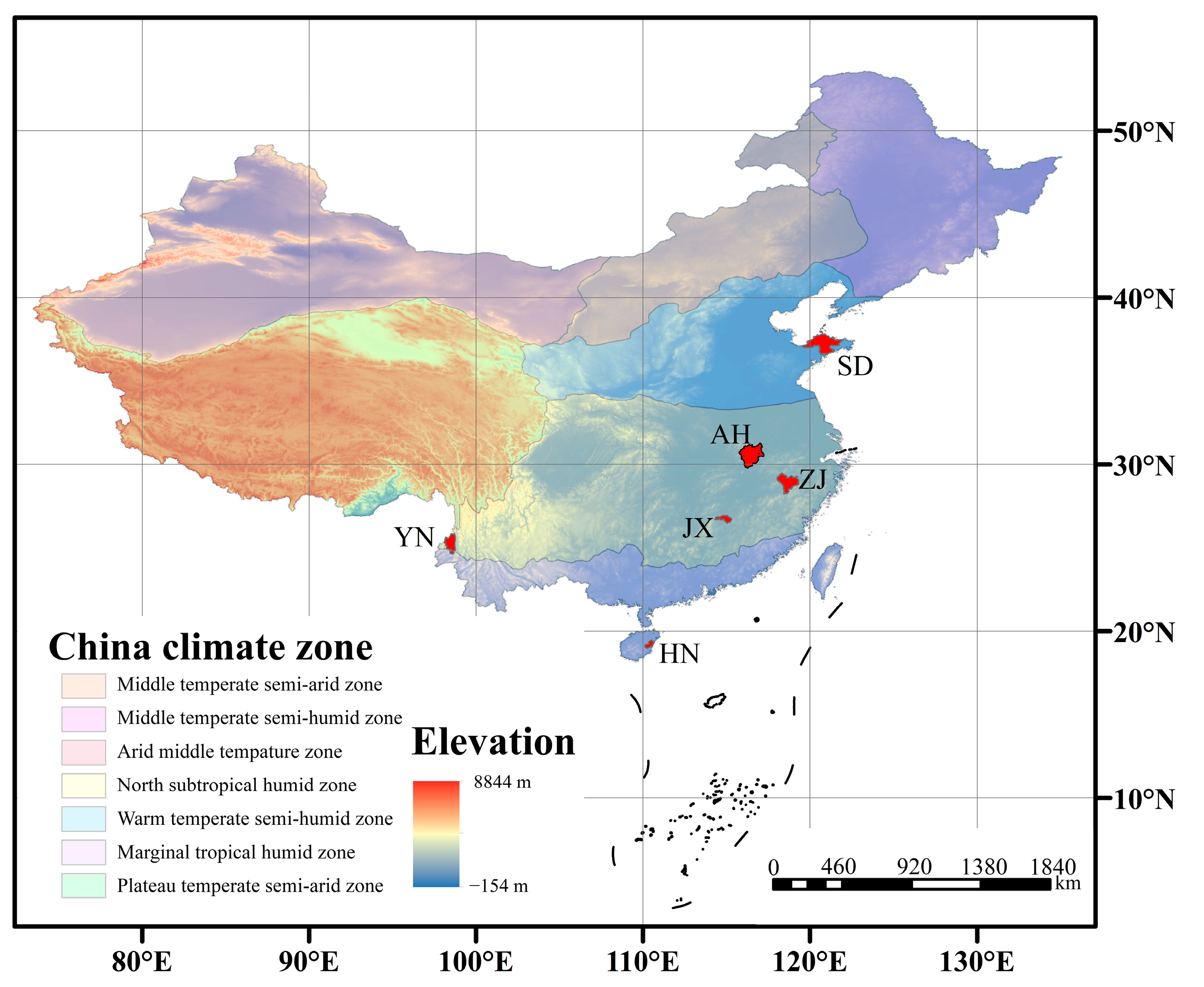
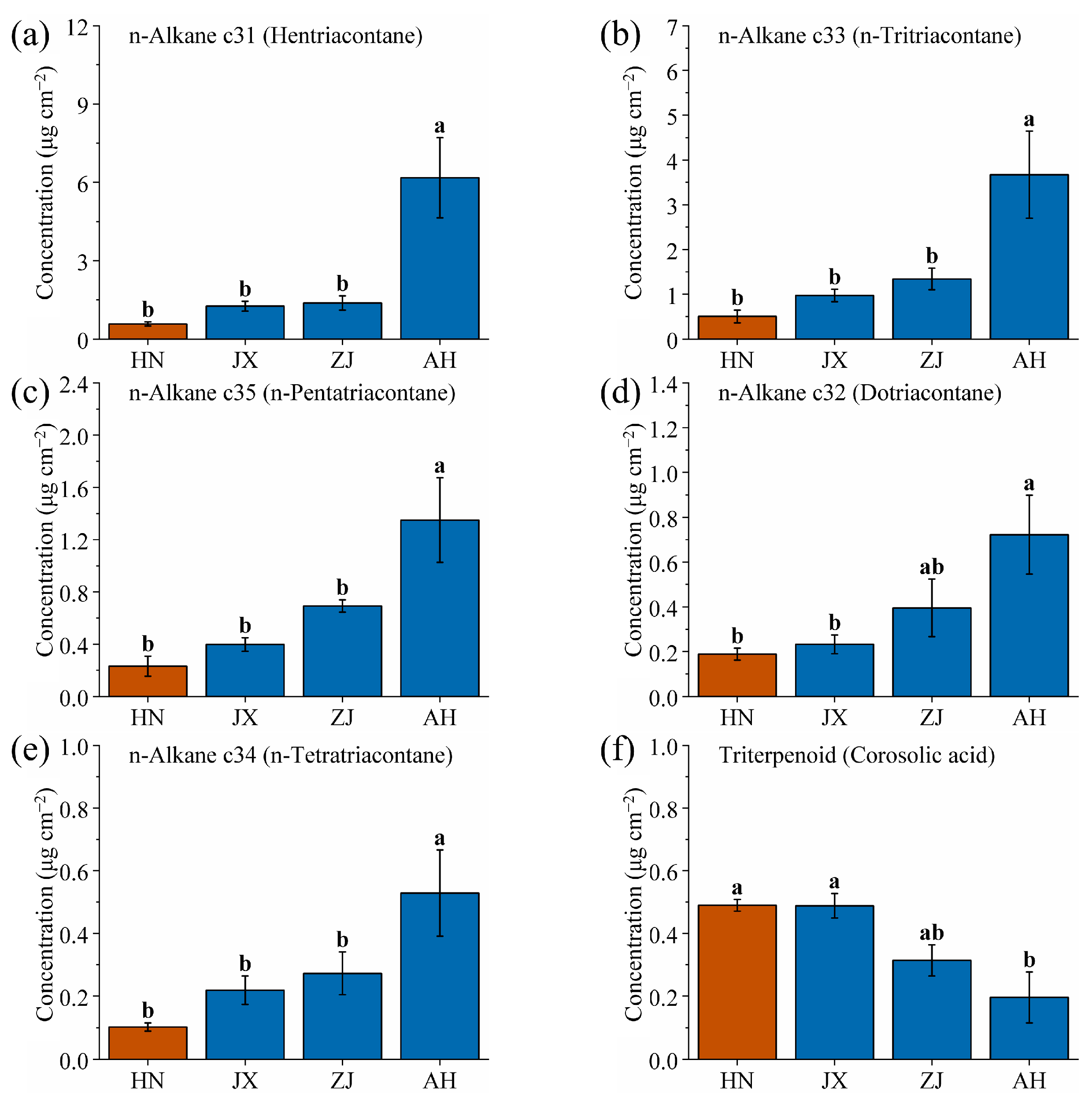
2.3. Search for the Relationship Between VF Production Area and Latitude
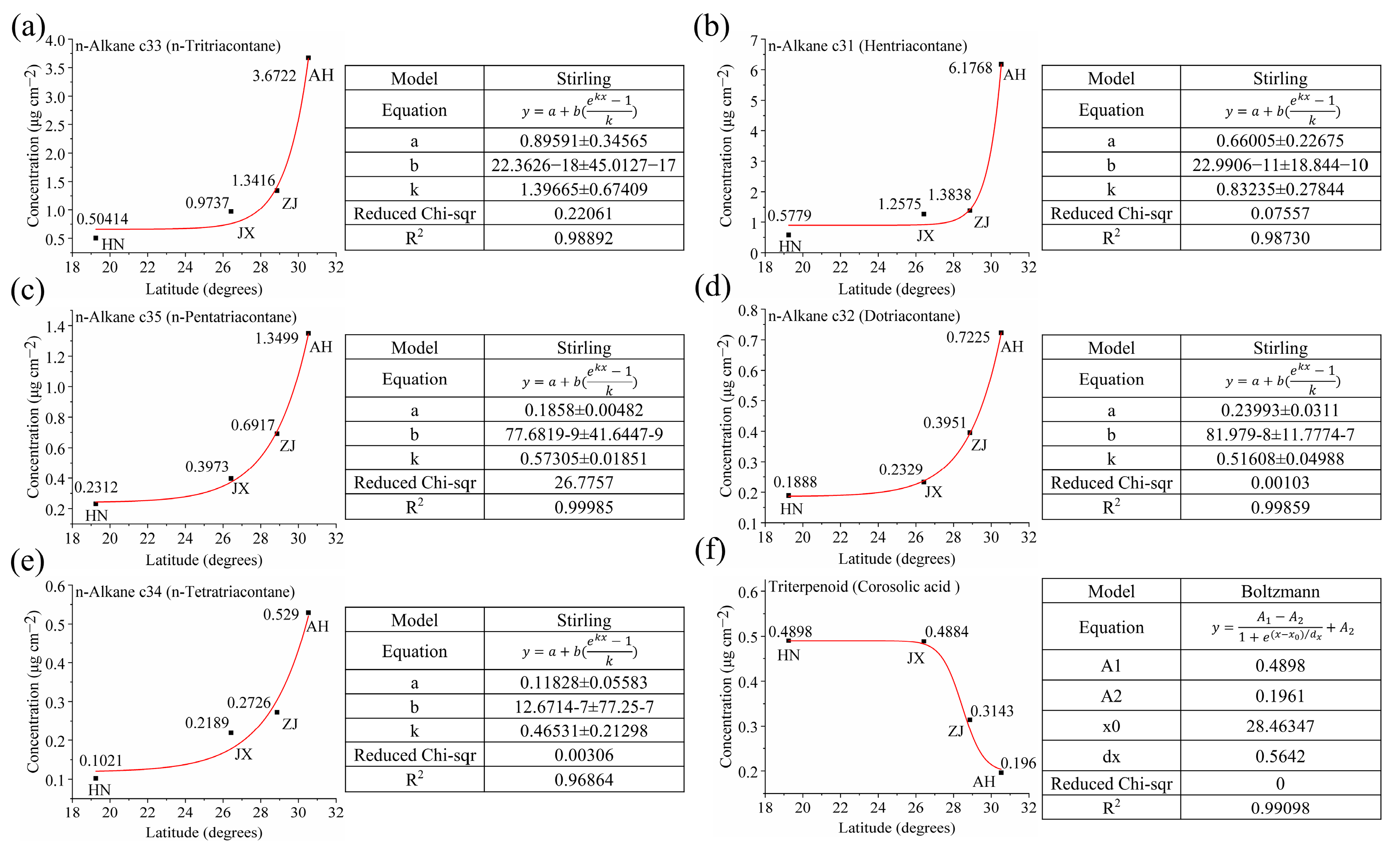
2.4. Establishment of a Model of the Relationship Between Single-Component Waxes and Latitude That Can Be Used to Identify the VF Production Area
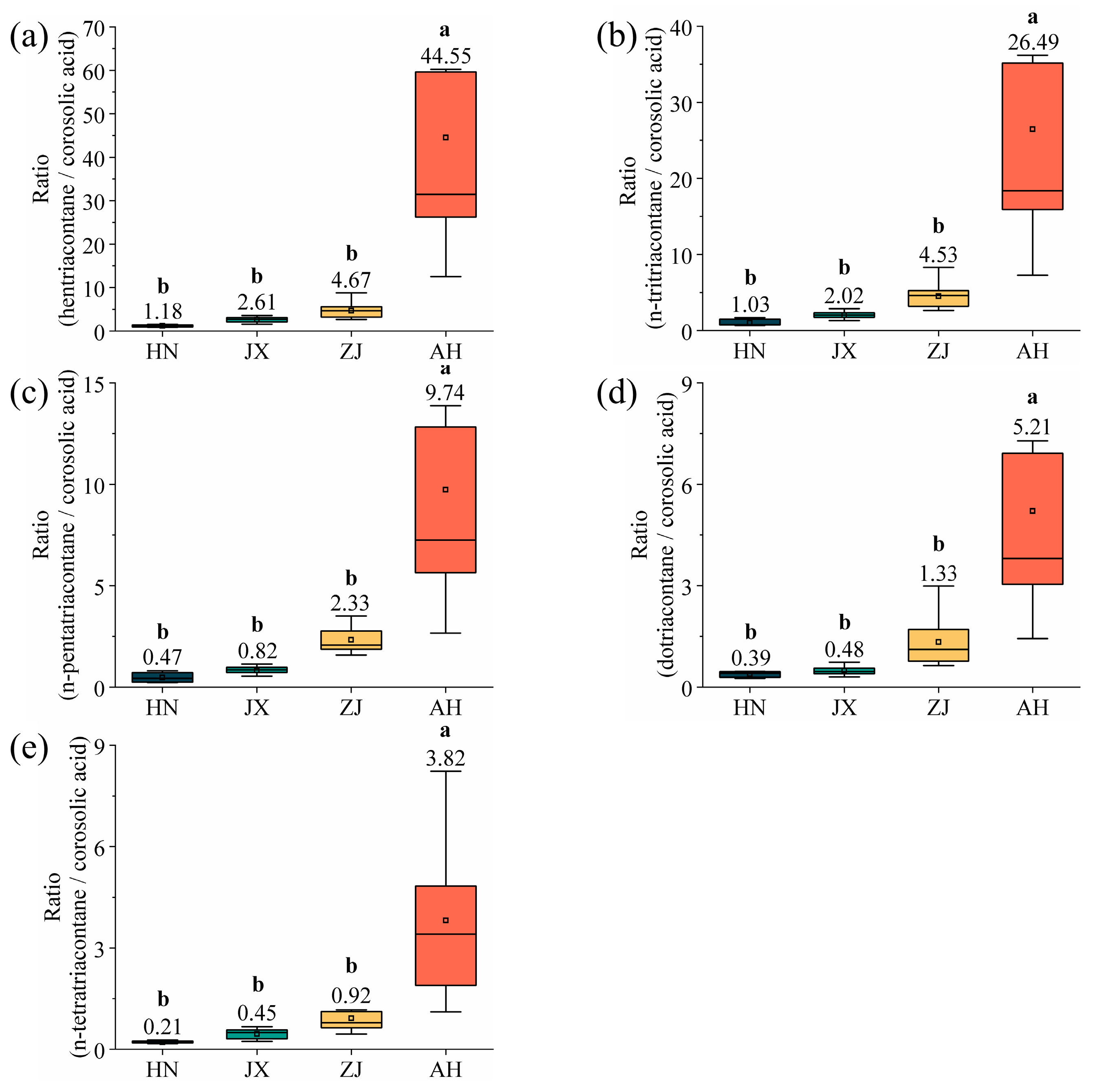
2.5. Establishment of a Model of the Relationship Between Multi-Component Waxes and Latitude That Can Be Used to Identify the VF Production Area
2.6. Identification of the Production Area of VF Using the Multi-Component Wax Ratio
2.7. Investigate the Relationship Between Cuticular Waxes and VF Quality by Internal Metabolites
3. Materials and Methods
3.1. Reagents
3.2. Selection of VF Sample for Cuticular Wax Extraction
3.3. GC-MS Detection of Wax Extracts
3.4. Internal Metabolomics LC-MS Detection of VF
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le, D.D.; Han, S.; Ahn, J.; Yu, J.; Kim, C.-K.; Lee, M. Analysis of Antioxidant Phytochemicals and Anti-Inflammatory Effect from Vitex rotundifolia L. f. Antioxidants 2022, 11, 454. [Google Scholar] [CrossRef]
- Ong, H.G.; Kim, Y.D. Quantitative ethnobotanical study of the medicinal plants used by the Ati Negrito indigenous group in Guimaras island, Philippines. J. Ethnopharmacol. 2014, 157, 228–242. [Google Scholar] [CrossRef]
- Xavier, T.F.; Kannan, M.; Lija, L.; Auxillia, A.; Rose, A.K.F.; Sen, S. Ethnobotanical study of Kani tribes in Thoduhills of Kerala, South India. J. Ethnopharmacol. 2014, 152, 78–90. [Google Scholar] [CrossRef]
- Feng, J.; Li, M.; Wei, Q.; Li, S.; Song, S.; Hua, Z. Unconjugated bilirubin induces pyroptosis in cultured rat cortical astrocytes. J. Neuroinflammation 2018, 15, 23. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Guo, L.; Cao, L.-J.; Zhang, J.; Yin, Z.-Q. A new iridoid glycoside from the fruits of Vitex rotundifolia. J. Asian Nat. Prod. Res. 2017, 19, 2491–2496. [Google Scholar] [CrossRef]
- Joshi, A.R.; Joshi, K. Indigenous knowledge and uses of medicinal plants by local communities of the Kali Gandaki Watershed Area, Nepal. J. Ethnopharmacol. 2000, 73, 175–183. [Google Scholar] [CrossRef]
- Rani, A.; Sharma, A. The Genus Vitex: A Review. Pharmacogn. Rev. 2013, 7, 188–198. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Kamali, M.; Doustimotlagh, A.H.; Nowroozzadeh, M.H.; Rasekh, F.; Hashempur, M.H.; Iraji, A. A comprehensive review of ethnomedicinal approaches, phytochemical analysis, and pharmacological potential of Vitex trifolia L. Front. Pharmacol. 2024, 15, 1322083. [Google Scholar] [CrossRef]
- Gentallan, R.P., Jr.; Madayag, R.E.; Bartolome, M.C.B.; Alvaran, B.B.S.; Magtoltol, J.B.; Quiñones, K.J.O.; Coronado, N.B.; Altoveros, N.C.; Borromeo, T.H.; Endonela, L.E.; et al. Towards the establishment of “lagundi” (Vitex trifolia s. l.) reference germplasm from the Philippines: An agro-morphological and phytochemical evaluation of native genotypes. Ind. Crops Prod. 2014, 208, 117758. [Google Scholar] [CrossRef]
- Li, B.; Cantino, P.D.; Olmstead, R.G.; Bramley, G.L.; Xiang, C.-L.; Ma, Z.-H.; Tan, Y.-H.; Zhang, D.-X. A large-scale chloroplast phylogeny of the Lamiaceae sheds new light on its subfamilial classification. Sci. Rep. 2016, 6, 34343. [Google Scholar] [CrossRef]
- Yan, C.X.; Wei, Y.W.; Li, H.; Xu, K.; Zhai, R.X.; Meng, D.C.; Fu, X.J.; Ren, X. Vitex rotundifolia L. f. and Vitex trifolia L.: A review on their traditional medicine, phytochemistry, pharmacology. J. Ethnopharmacol. 2023, 308, 116273. [Google Scholar] [CrossRef]
- Musa, N.; Banerjee, S.; Maspalma, G.A.; Usman, L.U.; Hussaini, B. Assessment of the phytochemical, antioxidant and larvicidal activity of essential oil extracted from Simpleleaf Chastetree [Vitex trifolia] leaves obtained from Ganye Local Government, Adamawa State-Nigeria. Mater. Today 2021, 49, 3435–3438. [Google Scholar] [CrossRef]
- Mustarichie, R.; Sumiwi, S.A.; Hanifah, A.T. ED50 from anti-inflammatory properties of Vitex trifolia L. ethanol extract. Natl. J. Physiol. Pharm. Pharmacol. 2018, 8, 719–725. [Google Scholar] [CrossRef]
- Nwodo, N.; Okoye, F.; Lai, D.; Debbab, A.; Kaiser, M.; Brun, R.; Proksch, P. Evaluation of the in vitro trypanocidal activity of methylated flavonoid constituents of Vitex simplicifolia leaves. BMC Complement. Altern. Med. 2015, 15, 82. [Google Scholar] [CrossRef]
- Sathiamoorthy, B.; Gupta, P.; Kumar, M.; Chaturvedi, A.K.; Shukla, P.K.; Maurya, R. New antifungal flavonoid glycoside from Vitex negundo. Bioorg. Med. Chem. Lett. 2007, 17, 239–242. [Google Scholar] [CrossRef]
- Garbi, M.I.; Osman, E.E.; Abdallah, E.T.; Kabbashi, A.S.; Salam, H.A.A. Cytotoxicity of Vitex trifolia leaf extracts on MCF-7 and Vero cell lines. J. Sci. Innov. Res. 2015, 4, 89–93. [Google Scholar] [CrossRef]
- Rana, G. Inhibition efficiency of a newly isolated flavonoid compound from Vitex negundo L. leaves against cattle-endosymbiont Setaria cervi: Phytomedicine for lymphatic filariasis. Parasite Epidemiol. Control 2018, 3, 88–95. [Google Scholar] [CrossRef]
- Matsui, M.; Kumar-Roine, S.; Darius, H.T.; Chinain, M.; Laurent, D.; Pauillac, S. Characterisation of the anti-inflammatory potential of Vitex trifolia L. (Labiatae), a multipurpose plant of the Pacific traditional medicine. J. Ethnopharmacol. 2009, 126, 427–433. [Google Scholar] [CrossRef]
- Chokpaisarn, J.; Paduka, W.; Sotthibandhu, D.S.; Phoopha, S.; Limsuwan, S.; Maneenoon, K.; Mahaprom, K.; Chaiwong, K. Evaluation of biological properties and phytochemical profile of Vitex trifolia L. extract and its infused oil as a value-added product for health promotion. South Afr. J. Bot. 2024, 169, 543–552. [Google Scholar] [CrossRef]
- Zahid, H.; Rizwani, G.H.; Ishaqe, S. Phytopharmacological Review on Vitex agnus-castus: A Potential Medicinal Plant. Chin. Herb. Med. 2016, 8, 24–29. [Google Scholar] [CrossRef]
- Eha, B.; Jannavi, R.; Sukumaran, P. Phyto-pharmacological and Biological Aspects of Vitex negundo Medicinal Plant—A Review. J. Pharm. Res. Int. 2021, 33, 17–32. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Wong, S.K.; Chan, H.T. Casticin from Vitex species: A short review on its anticancer and anti-inflammatory properties. J. Integr. Med. 2018, 16, 147–152. [Google Scholar] [CrossRef]
- Wee, H.N.; Neo, S.Y.; Singh, D.; Yew, H.-C.; Qiu, Z.-Y.; Tsai, X.-R.C.; How, S.-Y.; Yip, K.-Y.C.; Tan, C.-H.; Koh, H.-L. Effects of Vitex trifolia L. leaf extracts and phytoconstituents on cytokine production in human U937 macrophages. BMC Complement. Med. Ther. 2020, 20, 91. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Huang, H.; Yin, M.; Jenks, M.A.; Kosma, D.K.; Yang, P.; Yang, X.; Zhao, H.; Lü, S. Deciphering the core shunt mechanism in Arabidopsis cuticular wax biosynthesis and its role in plant environmental adaptation. Nat. Plants 2025, 11, 165–175. [Google Scholar] [CrossRef]
- Kosma, D.K.; Bourdenx, B.; Bernard, A.; Parsons, E.P.; Lu, S.; Joubès, J.; Jenks, M.A. The impact of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol. 2009, 151, 1918–1929. [Google Scholar] [CrossRef]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef]
- Lee, S.B.; Suh, M.C. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef]
- Kim, H.; Yu, S.-I.; Jung, S.H.; Lee, B.-H.; Suh, M.C. The F-Box Protein SAGL1 and ECERIFERUM3 Regulate Cuticular Wax Biosynthesis in Response to Changes in Humidity in Arabidopsis. Plant Cell 2019, 31, 2223–2240. [Google Scholar] [CrossRef]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–712. [Google Scholar] [CrossRef]
- Buschhaus, C.; Jetter, R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal cuticulars? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Wu, X.; Wang, L.; Li, J.; Wan, M.; Jia, B.; Ye, Z.; Liu, L.; Tang, X.; et al. Characterization of the PbrLACS2 in pear and role in wax biosynthesis in Arabidopsis. Plant Biotechnol. J. 2025, 21, 3. [Google Scholar] [CrossRef]
- Suresh, K.; Zeisler-Diehl, V.; Schreiber, L. Comparing anatomy, chemical composition, and water permeability of suberized organs in five plant species: Wax makes the difference. Planta 2022, 256, 60. [Google Scholar] [CrossRef]
- Wang, Y.; Su, S.; Chen, G.; Mao, H.; Jiang, Y. Relationship Between Cuticular Waxes and Storage Quality Parameters of Korla Pear Under Different Storage Methods. J. Plant Growth Regul. 2021, 40, 1152–1165. [Google Scholar] [CrossRef]
- Dimopoulos, N.; Tindjau, R.; Wong, D.C.J.; Matzat, T.; Haslam, T.; Song, C.; Gambetta, G.A.; Kunst, L.; Castellarin, S.D. Drought stress modulates cuticular wax composition of the grape berry. J. Exp. Bot. 2020, 71, 3126–3141. [Google Scholar] [CrossRef]
- Zuo, H.; Si, X.; Li, P.; Li, J.; Chen, Z.; Li, P.; Chen, C.; Liu, Z.; Zhao, J. Dynamic change of tea (Camellia sinensis) leaf cuticular wax in white tea processing for contribution to tea flavor formation. Food Res. Int. 2023, 163, 112182. [Google Scholar] [CrossRef]
- Chen, M. The Tea Plant Leaf Cuticle: From Plant Protection to Tea Quality. Front. Plant Sci. 2021, 12, 751547. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Zhang, Y.; Du, Z.; Chen, X.; Kong, X.; Sun, W.; Chen, C. Drought stress modify cuticle of tender tea leaf and mature leaf for transpiration barrier enhancement through common and distinct modes. Sci. Rep. 2020, 10, 6696. [Google Scholar] [CrossRef]
- He, Y.; Gao, J.; Guo, N.; Guo, Y. Variations of leaf cuticular waxes among C3 and C4 Gramineae herbs. Chem. Biodivers. 2016, 13, 1460–1468. [Google Scholar] [CrossRef]
- Zhu, Y.; Guan, M.; Jia, Q.; Wang, G.; Pan, L.; Li, Y. Degradation mechanism of cuticular wax composition and surface properties of bamboo culm during storage. Ind. Crops Prod. 2024, 214, 118558. [Google Scholar] [CrossRef]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z.-H. Molecular and evolutionary mechanisms of cuticular wax for plant drought tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Li, J.; Xiao, H.; Xu, Y.; Fan, S.; Xie, Z.; Guo, M.; Yang, J.; Jing, X.; et al. Chemical characterization and discovery of novel quality markers in Citrus aurantium L. fruit from traditional cultivation areas in China using GC–MS-based cuticular waxes analysis. Food Chem. 2023, 20, 100890. [Google Scholar] [CrossRef]
- Schuster, A.C.; Burghardt, M.; Alfarhan, A.; Bueno, A.; Hedrich, R.; Leide, J.; Thomas, J.; Riederer, M. Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants 2016, 8, plw027. [Google Scholar] [CrossRef]
- Bueno, A.; Alfarhan, A.; Arand, K.; Burghardt, M.; Deininger, A.-C.; Hedrich, R.; Leide, J.; Seufert, P.; Staiger, S.; Riederer, M. Effects of temperature on the cuticular transpiration barrier of two desert plants with water-spender and water-saver strategies. J. Exp. Bot. 2019, 70, 1613–1625. [Google Scholar] [CrossRef]
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef]
- Rao, I.M.; Zeigler, R.S.; Vera, R.; Sarkarung, S. Selection and breeding for acid-soil tolerance in crops. BioScience 1993, 43, 454–465. [Google Scholar] [CrossRef]
- Schultz, J.C.; Appel, H.M.; Ferrieri, A.P.; Arnold, T.M. Flexible resource allocation during plant defense responses. Front. Plant Sci. 2013, 4, 324. [Google Scholar] [CrossRef]
- Kidd, P.S.; Proctor, J. Why plants grow poorly on very acid soils: Are ecologists missing the obvious? J. Exp. Bot. 2001, 52, 791–799. [Google Scholar] [CrossRef]
- Shi-zhen, L. Characteristics of some types of paddy soil in Northeast China. In Proceedings of Symposium on Paddy Soils; Springer: Berlin/Heidelberg, Germany, 1981; pp. 680–685. [Google Scholar] [CrossRef]
- Bi, X.; Chu, H.; Fu, M.; Xu, D.; Zhao, W.; Zhong, Y.; Wang, M.; Li, K.; Zhang, Y. Distribution characteristics of organic carbon (nitrogen) content, cation exchange capacity, and specific surface area in different soil particle sizes. Sci. Rep. 2023, 13, 12242. [Google Scholar] [CrossRef]
- Steinmüller, D.; Tevini, M. Action of ultraviolet radiation (UV-B) upon cuticular waxes in some crop plants. Planta 1985, 164, 557–564. [Google Scholar] [CrossRef]
- Tevini, M.; Steinmüller, D. Influence of light, UV-B radiation, and herbicides on wax biosynthesis of cucumber seedlings. J. Plant Physiol. 1987, 131, 111–121. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Wen, X.; Wang, X.-B.; Shen, D.-P.; Ding, L.-W.; He, J. UV-B induced expression of wax synthesis gene LbCYP96A15 in leaves of goji plants (Lycium barbarum) to improve powdery mildew resistance. Chem. Biol. Technol. Agric. 2025, 12, 43. [Google Scholar] [CrossRef]
- Acharya, K.P.; De Frenne, P.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; Diekmann, M.; Hermy, M.; Kolb, A.; Lemke, I.; Plue, J.; et al. Latitudinal variation of life-history traits of an exotic and a native impatiens species in Europe. Acta Oecologica 2017, 81, 40–47. [Google Scholar] [CrossRef]
- Malizia, A.; Blundo, C.; Carilla, J.; Acosta, O.O.; Cuesta, F.; Duque, A.; Aguirre, N.; Aguirre, Z.; Ataroff, M.; Baez, S.; et al. Elevation and latitude drives structure and tree species composition in Andean forests: Results from a large-scale plot network. PLoS ONE 2020, 15, e0231553. [Google Scholar] [CrossRef]
- Shen, P.; Jiang, X.; Zhang, J.; Wang, J.; Raj, R.; Li, G.; Ge, H.; Wang, W.; Yu, B.; Zhang, J. Isolation and microbial transformation of tea sapogenin from seed pomace of Camellia oleifera with anti-inflammatory effects. Chin. J. Nat. Med. 2024, 22, 280–288. [Google Scholar] [CrossRef]
- Davidyants, E.S. Triterpene glycosides as plant growth regulators: Application potential and prospects (A review). Russ. J. Bioorganic Chem. 2024, 50, 2730–2752. [Google Scholar] [CrossRef]
- Saleem, S.; Mushtaq, N.U.; Shah, W.H.; Rasool, A.; Hakeem, K.R.; Rehman, R.U. Beneficial role of phytochemicals in oxidative stress mitigation in plants. In Antioxidant Defense in Plants; Aftab, T., Hakeem, K.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 435–451. [Google Scholar] [CrossRef]
- Grassmann, J. Terpenoids as plant antioxidants. Vitam. Horm. 2005, 72, 505–535. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, C.; Zheng, T.; Zuo, Z. Thermal protection function of camphor on Cinnamomum camphora cell membrane by acting as a signaling molecule. Plant Physiol. Biochem. 2023, 198, 107672. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, L.; Rusalepp, L.; Kaurilind, E.; Sulaiman, H.Y.; Püssa, T.; Niinemets, Ü. Heat priming improved heat tolerance of photosynthesis, enhanced terpenoid and benzenoid emission and phenolics accumulation in Achillea millefolium. Plant Cell Environ. 2021, 44, 2365–2385. [Google Scholar] [CrossRef]
- Long, L.M.; Patel, H.P.; Cory, W.C.; Stapleton, A.E. The maize epicuticular wax layer provides UV protection. Funct. Plant Biol. 2003, 30, 75–81. [Google Scholar] [CrossRef]
- Wu, H.; Liu, L.; Chen, Y.; Liu, T.; Jiang, Q.; Wei, Z.; Li, C.; Wang, Z. Tomato SlCER1-1 catalyzes the synthesis of wax alkanes which increases the drought tolerance and fruit storability. Hortic. Res. 2022, 9, uhac004. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Zhi, P.; Wang, X.; Xu, B.; Gong, Z.; Chang, C. Origins and Evolution of Cuticle Biosynthetic Machinery in Land Plants. Plant Physiol. 2020, 184, 1998–2010. [Google Scholar] [CrossRef]
- Pathak, N.; Negi, A.S. Plant based steroidal and triterpenoid sapogenins: Chemistry on diosgenin and biological aspects. Eur. J. Med. Chem. 2024, 279, 116915. [Google Scholar] [CrossRef]
- Rodler, C.; Schinnerl, J.; Schalamun, M.; Hinterdobler, W.; Brecker, L. Diosgenin and solasodine-based saponins and sapogenines—Effects of structural modifications on Panagrellus redivivus, Spodoptera littoralis and Trichoderma reesei. Results Chem. 2025, 13, 101994. [Google Scholar] [CrossRef]
- Mortici, C. Best estimates of the generalized Stirling formula. Appl. Math. Comput. 2010, 215, 4044–4048. [Google Scholar] [CrossRef]
- He, T.-X. Generalized Stirling Numbers and Generalized Stirling Functions. arXiv 2011, arXiv:1106.5251. [Google Scholar] [CrossRef]
- Filbet, F.; Russo, G. Accurate numerical methods for the Boltzmann equation. In Modeling and Computational Methods for Kinetic Equations. Modeling and Simulation in Science, Engineering and Technology; Degond, P., Pareschi, L., Russo, G., Eds.; Birkhäuser: Boston, MA, USA, 2004. [Google Scholar] [CrossRef]
- Kunst, L.; Samuels, A.L. Biosynthesis and secretion of plant cuticular wax. Prog. Lipid Res. 2003, 42, 51–80. [Google Scholar] [CrossRef]
- Yang, X.; Huang, H.; Wang, Z.; Haslam, T.M.; Kunst, L.; Wang, P.; Zhao, H.; Lü, S.; Ma, C. Acetyl-CoA carboxylase1 influences ECERIFERUM2 activity to mediate the synthesis of very-long-chain fatty acid past C28. Plant Physiol. 2025, 197, kiae253. [Google Scholar] [CrossRef]
- Cassagne, C.; Lessire, R.; Bessoule, J.-J.; Moreau, P.; Creach, A.; Schneider, F.; Sturbois, B. Biosynthesis of very long chain fatty acids in higher plants. Prog. Lipid Res. 1994, 33, 55–69. [Google Scholar] [CrossRef]
- de Perez Souza, L.; Garbowicz, K.; Brotman, Y.; Tohge, T.; Fernie, A.R. The acetate pathway supports flavonoid and lipid biosynthesis in Arabidopsis. Plant Physiol. 2020, 182, 857–869. [Google Scholar] [CrossRef]
- Wang, Z.H.; Kang, K.A.; Zhang, R.; Piao, M.J.; Jo, S.H.; Kim, J.S.; Kang, S.S.; Lee, J.S.; Park, D.H.; Hyun, J.W. Myricetin suppresses oxidative stress-induced cell damage via both direct and indirect antioxidant action. Environ. Toxicol. Pharmacol. 2010, 29, 12–18. [Google Scholar] [CrossRef]
- Xie, H.-J.; Mou, W.-S.; Lin, F.-R.; Xu, J.-H.; Lei, Q.-F.; Fang, W.-J. Radical scavenging activity of myricetin. Acta Phys. -Chim. Sin. 2013, 29, 1421–1432. [Google Scholar] [CrossRef]
- Rogowska, A.; Szakiel, A. Enhancement of phytosterol and triterpenoid production in plant hairy root cultures—Simultaneous stimulation or competition? Plants 2021, 10, 2028. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Zhang, C.-L.; Wang, G.-L.; Wang, Y.-X.; Qi, C.-H.; Zhao, Q.; You, C.-X.; Li, Y.-Y.; Hao, Y.-J. The R2R3 MYB transcription factor MdMYB30 modulates plant resistance against pathogens by regulating cuticular wax biosynthesis. BMC Plant Biol. 2019, 19, 362. [Google Scholar] [CrossRef]
- Falginella, L.; Andre, C.M.; Legay, S.; Lin-Wang, K.; Dare, A.P.; Deng, C.; Rebstock, R.; Plunkett, B.J.; Guo, L.; Cipriani, G.; et al. Differential regulation of triterpene biosynthesis induced by an early failure in cuticle formation in apple. Hortic. Res. 2021, 8, 75. [Google Scholar] [CrossRef]
- Leide, J.; Hildebrandt, U.; Reussing, K.; Riederer, M.; Vogg, G. The developmental pattern of tomato fruit wax accumulation and its impact on cuticular transpiration barrier properties: Effects of a deficiency in a β-ketoacyl-coenzyme A synthase (LeCER6). Plant Physiol. 2007, 144, 1667–1679. [Google Scholar] [CrossRef]
- Ghafari, A.T.; Jahidin, A.H.; Zakaria, Y.; Hasan, M.H. Phytochemical Screening and High-performance Thin-layer Chromatography Quantification of Vitex trifolia Leaves Hydro-alcoholic Extract: Potential Anti-inflammatory Properties. J. Pharm. Res. Int. 2021, 33, 111–121. [Google Scholar] [CrossRef]
- Huang, H.; Wang, L. Alteration of cuticular morphologies and chemical composition of cuticle in response to chilling injury in papaya (Carica papaya L.) after harvest. Food Chem. 2023, 416, 135751. [Google Scholar] [CrossRef]
- Rafiq, M.; Javaid, A.; Kanwal, A.; Anwar, A.; Khan, I.H.; Kanwal, Q.; Cheng, C. GC-MS analysis and antifungal potential of flower extract of Acacia nilotica subsp. Indica against Macrophomina phaseolina. Microb. Pathog. 2024, 194, 106819. [Google Scholar] [CrossRef]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of Plant Cuticular Waxes. In Plant Cuticles: An Integrated Functional Approach; Riederer, M., Müller, C., Eds.; Garland Science: New York, NY, USA, 2006; Chapter 4. [Google Scholar] [CrossRef]
- Vasilev, N.; Boccard, J.; Lang, G.; Grömping, U.; Fischer, R.; Goepfert, S.; Rudaz, S.; Schillberg, S. Structured plant metabolomics for the simultaneous exploration of multiple factors. Sci. Rep. 2016, 6, 37390. [Google Scholar] [CrossRef]
- Rafiq, M.; Shoaib, A.; Javaid, A.; Parveen, S.; Hassan, M.A.; Nawaz, H.H.; Cheng, C. Application of Asteraceae biomass and biofertilizers to improve potato crop health by controlling black scurf disease. Front. Plant Sci. 2024, 15, 1437702. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Human Serum Metabolome (HUSERMET) Consortium. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Times (min) | Waxes | HN | YN | JX | ZJ | AH | SD | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%) | SD | CV | (%) | SD | CV | (%) | SD | CV | (%) | SD | CV | (%) | SD | CV | (%) | SD | CV | ||
| 14.738 | Fatty acid c24 (Lignoceric acid) | 8.017 ab | 0.005 | 0.067 | 6.190 abc | 0.013 | 0.214 | 4.722 bc | 0.019 | 0.392 | 4.351 c | 0.018 | 0.409 | 6.162 abc | 0.006 | 0.102 | 8.688 a | 0.010 | 0.118 |
| 15.664 | n-Alkane c29 (Nonacosane) | 7.816 c | 0.004 | 0.057 | 11.119 b | 0.017 | 0.154 | 6.754 c | 0.018 | 0.263 | 4.680 c | 0.021 | 0.439 | 14.636 ab | 0.009 | 0.062 | 15.323 a | 0.010 | 0.067 |
| 16.522 | Primary alcohol c26 (Hexacosanol) | 2.439 a | 0.014 | 0.592 | 3.119 a | 0.007 | 0.212 | 3.404 a | 0.016 | 0.473 | 3.260 a | 0.009 | 0.271 | 1.592 a | 0.006 | 0.372 | 4.202 a | 0.013 | 0.316 |
| 17.108 | n-Alkane c30 (n-Triacontane) | 10.905 bc | 0.021 | 0.195 | 8.086 bc | 0.033 | 0.409 | 14.381 ab | 0.022 | 0.153 | 18.009 a | 0.071 | 0.394 | 4.248 c | 0.009 | 0.216 | 8.850 bc | 0.050 | 0.571 |
| 17.805 | Fatty acid c26 (Hexacosenoic acid) | 5.855 a | 0.017 | 0.296 | 5.534 a | 0.035 | 0.628 | 2.258 a | 0.004 | 0.196 | 3.041 a | 0.027 | 0.877 | 2.150 a | 0.007 | 0.303 | 2.133 a | 0.013 | 0.623 |
| 18.680 | n-Alkane c31 (Hentriacontane) | 9.482 d | 0.012 | 0.131 | 22.007 b | 0.009 | 0.041 | 13.852 c | 0.025 | 0.180 | 13.451 c | 0.008 | 0.057 | 29.579 a | 0.013 | 0.043 | 20.125 b | 0.011 | 0.052 |
| 19.623 | Primary alcohol c28 (Octacosanol) | 1.719 a | 0.002 | 0.129 | 1.444 a | 0.002 | 0.120 | 1.791 a | 0.002 | 0.121 | 2.166 a | 0.008 | 0.355 | 1.515 a | 0.010 | 0.635 | 1.711 a | 0.018 | 1.068 |
| 20.243 | n-Alkane c32 (Dotriacontane) | 3.099 a | 0.003 | 0.091 | 3.154 a | 0.003 | 0.102 | 2.565 ab | 0.002 | 0.065 | 3.841 a | 0.004 | 0.111 | 3.460 a | 0.003 | 0.090 | 1.684 b | 0.007 | 0.393 |
| 20.421 | Beta-amyrin | 1.818 b | 0.002 | 0.094 | 1.461 b | 0.003 | 0.218 | 3.635 b | 0.005 | 0.147 | 2.625 ab | 0.009 | 0.341 | 2.052 b | 0.004 | 0.202 | 3.014 ab | 0.009 | 0.288 |
| 20.931 | Alpha-amyrin | 2.172 b | 0.001 | 0.052 | 2.185 b | 0.007 | 0.299 | 4.432 a | 0.012 | 0.273 | 2.593 b | 0.009 | 0.365 | 2.274 b | 0.003 | 0.152 | 1.785 b | 0.009 | 0.531 |
| 21.093 | Fatty acid c28 (Octacosanoic acid) | 3.125 ab | 0.005 | 0.144 | 3.844 a | 0.009 | 0.221 | 1.226 b | 0.002 | 0.136 | 5.344 a | 0.017 | 0.313 | 1.298 b | 0.009 | 0.710 | 2.703 ab | 0.010 | 0.367 |
| 21.959 | n-Alkane c33 (n-Tritiacontane) | 8.272 d | 0.017 | 0.209 | 14.553 b | 0.010 | 0.066 | 10.726 c | 0.013 | 0.122 | 13.041 bc | 0.010 | 0.079 | 17.585 a | 0.010 | 0.060 | 11.216 c | 0.007 | 0.066 |
| 22.630 | Fatty acid c29 (Nonacosanoic acid) | 1.463 a | 0.007 | 0.475 | 1.407 a | 0.006 | 0.436 | 0.946 a | 0.001 | 0.047 | 1.448 a | 0.005 | 0.319 | 0 a | 0 | — | 0 a | 0 | — |
| 22.936 | Primary alcohol c30 (Triacontanol) | 0 a | 0 | — | 5.695 a | 0.027 | 0.466 | 0.430 a | 0.001 | 0.167 | 0 a | 0 | — | 0.620 a | 0.004 | 0.663 | 0 a | 0 | — |
| 23.395 | n-Alkane c34 (n-Tetratriacontane) | 1.676 a | 0.003 | 0.160 | 1.530 a | 0.009 | 0.558 | 2.412 a | 0.004 | 0.183 | 2.651 a | 0.002 | 0.082 | 2.533 a | 0.012 | 0.462 | 2.496 a | 0.008 | 0.318 |
| 23.726 | Fatty acid c30 (Triacontanoic acid) | 2.987 a | 0.001 | 0.032 | 1.831 a | 0.009 | 0.466 | 1.038 a | 0.007 | 0.672 | 1.206 a | 0.005 | 0.453 | 0.996 a | 0.007 | 0.663 | 1.835 a | 0.009 | 0.517 |
| 24.525 | Oleanolic acid | 1.812 ab | 0.008 | 0.434 | 2.700 ab | 0.009 | 0.316 | 3.334 a | 0.005 | 0.141 | 1.340 ab | 0.010 | 0.745 | 1.138 b | 0.004 | 0.390 | 1.945 ab | 0.010 | 0.531 |
| 25.213 | Ursolic acid | 4.262 a | 0.004 | 0.091 | 0.842 d | 0.003 | 0.359 | 2.317 b | 0.002 | 0.106 | 1.090 cd | 0.004 | 0.331 | 0.783 d | 0.003 | 0.363 | 2.042 bc | 0.008 | 0.389 |
| 25.493 | n-Alkane c35 (n-Pentatriacontane) | 3.795 b | 0.011 | 0.287 | 5.679 ab | 0.021 | 0.372 | 4.377 ab | 0.006 | 0.146 | 6.724 a | 0.016 | 0.231 | 6.464 ab | 0.005 | 0.081 | 3.731 ab | 0.015 | 0.401 |
| 27.226 | Primary alcohol c32 (Dotriacontanol) | 2.351 a | 0.006 | 0.252 | 0.424 b | 0.002 | 0.406 | 1.859 a | 0.003 | 0.154 | 1.774 a | 0.003 | 0.176 | 0 b | 0 | — | 0 b | 0 | — |
| 28.033 | Maslinic acid | 4.282 a | 0.007 | 0.152 | 1.967 bc | 0.011 | 0.539 | 2.856 ab | 0.005 | 0.166 | 2.648 abc | 0.005 | 0.206 | 0.620 c | 0.002 | 0.256 | 3.218 ab | 0.012 | 0.359 |
| 28.356 | Corosolic acid | 8.036 a | 0.006 | 0.077 | 1.157 d | 0.005 | 0.389 | 5.380 b | 0.010 | 0.190 | 3.055 c | 0.015 | 0.489 | 0.939 d | 0.003 | 0.268 | 2.694 cd | 0.011 | 0.417 |
| Production Areas | Average Latitude | Average Altitude (m) | Soil Types | Soil pH | Annual Average Temperature (°C) | Annual Average Precipitation (mm) | Frost-Free Period (days) |
|---|---|---|---|---|---|---|---|
| Qionghai City Hainan Province | 19.25° N | 10~50 | Laterite soil Red soil | 5.0~6.5 | 24~26 | 2000~2500 | 365 |
| Tengchong City Yunnan Province | 25.02° N | 1600~1900 | Red soil Volcanic ash soil | 5.5~6.8 | 14~16 | 1400~1600 | 270~300 |
| Ji’an City Jiangxi Province | 27.12° N | 50~200 | Red soil Paddy soil | 4.5~6.0 | 17~19 | 1400~1600 | 280~300 |
| Quzhou City Zhejiang Province | 28.97° N | 50~150 | Red soil Yellow soil | 5.0~6.5 | 16~18 | 1500~1800 | 250~280 |
| Anqing City Anhui Province | 30.53° N | 20~100 | Yellow-brown soil Paddy soil | 5.5~7.0 | 15~17 | 1300~1500 | 240~260 |
| Yantai City Shandong Province | 37.52° N | 10~100 | Brown soil Brown soil | 6.5~7.5 | 11~13 | 550~700 | 200~220 |
| HN | YN | JX | ZJ | AH | SD | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Content | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) |
| μg/cm2 | ||||||||||||||||||
| Total fatty acids | 1.59 a | 0.25 | 26.06 | 2.60 a | 1.04 | 16.96 | 1.41 a | 0.64 | 15.49 | 1.75 a | 1.00 | 17.05 | 2.45 a | 1.01 | 11.72 | 2.64 a | 1.16 | 15.96 |
| Total n-alkanes | 2.75 a | 0.50 | 45.04 | 10.13 abc | 2.38 | 66.13 | 5.00 bc | 1.55 | 55.07 | 6.42 bc | 1.75 | 62.40 | 16.22 a | 7.00 | 77.66 | 10.47 ab | 5.56 | 63.43 |
| Total primary alcohols | 0.40 a | 0.18 | 6.51 | 1.01 a | 0.48 | 6.60 | 0.68 a | 0.39 | 7.48 | 0.74 a | 0.29 | 7.20 | 0.59 a | 0.44 | 2.81 | 0.98 a | 0.35 | 5.91 |
| Total triterpenoids | 1.36 a | 0.17 | 22.38 | 1.58 a | 0.42 | 10.31 | 1.99 a | 0.67 | 21.96 | 1.37 a | 0.47 | 13.35 | 1.63 a | 0.64 | 7.81 | 2.43 a | 1.39 | 14.70 |
| Total | 6.09 b | 15.31 ab | 9.08 b | 10.29 b | 20.88 a | 16.51 ab | ||||||||||||
| Classification | Content | HN | YN | JX | ZJ | AH | SD | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| μg/cm2 | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | Mean | SD | (%) | |
| Fatty | Tetracosanoic acid | 0.37 a | 0.20 | 6.01 | 0.16 a | 0.08 | 1.01 | 0.51 a | 0.53 | 5.63 | 0.27 a | 0.13 | 2.60 | 0.37 a | 0.17 | 1.74 | 0.20 a | 0.08 | 1.21 |
| Lignoceric acid | 0.49 b | 0.13 | 7.90 | 0.95 ab | 0.20 | 5.78 | 0.43 b | 0.05 | 4.71 | 0.45 b | 0.43 | 4.31 | 1.29 a | 0.49 | 6.02 | 1.43 a | 0.49 | 8.64 | |
| Hexacosenoic acid | 0.36 a | 0.10 | 5.77 | 0.85 a | 0.83 | 5.17 | 0.21 a | 0.10 | 2.25 | 0.31 a | 0.21 | 3.01 | 0.45 a | 0.27 | 2.10 | 0.35 a | 0.45 | 2.12 | |
| acids | Octacosanoic acid | 0.19 ab | 0.07 | 3.08 | 0.59 a | 0.11 | 3.59 | 0.11 b | 0.06 | 1.22 | 0.55 ab | 0.46 | 5.29 | 0.27 ab | 0.25 | 1.27 | 0.45 ab | 0.17 | 2.69 |
| Nonacosanoic acid | 0.09 b | 0.08 | 1.44 | 0.22 a | 0.05 | 1.32 | 0.09 b | 0.03 | 0.94 | 0.15 b | — | 1.43 | 0 b | — | 0 | 0 b | — | 0 | |
| Triacontanoic acid | 0.18 a | 0.05 | 2.94 | 0.28 a | 0.13 | 1.71 | 0.09 a | 0.06 | 1.03 | 0.12 a | 0.13 | 1.19 | 0.21 a | — | 0.97 | 0.30 a | 0.14 | 1.82 | |
| n-Alkanes | Nonacosane | 0.48 b | 0.08 | 7.70 | 1.70 ab | 0.29 | 10.39 | 0.61 b | 0.24 | 6.73 | 0.48 b | 0.39 | 4.64 | 3.06 a | 1.38 | 14.29 | 2.53 a | 1.21 | 15.23 |
| n-Triacontane | 0.66 a | 0.09 | 10.75 | 1.24 a | 0.81 | 7.56 | 1.31 a | 0.67 | 14.33 | 1.85 a | 0.42 | 17.84 | 0.89 a | 0.18 | 4.15 | 1.46 a | 1.31 | 8.80 | |
| Hentriacontane | 0.58 c | 0.14 | 9.34 | 3.37 b | 0.57 | 20.57 | 1.26 bc | 0.33 | 13.80 | 1.38 bc | 0.47 | 13.32 | 6.18 a | 2.66 | 28.88 | 3.32 b | 1.50 | 20.00 | |
| Dotriacontane | 0.19 b | 0.05 | 3.05 | 0.48 ab | 0.04 | 2.95 | 0.23 b | 0.07 | 2.56 | 0.40 b | 0.22 | 3.80 | 0.72 a | 0.31 | 3.38 | 0.28 b | 0.16 | 1.67 | |
| n-Tritriacontane | 0.50 c | 0.25 | 8.15 | 2.23 ab | 0.45 | 13.60 | 0.97 bc | 0.24 | 10.69 | 1.34 bc | 0.42 | 12.92 | 3.67 a | 1.68 | 17.17 | 1.85 bc | 0.96 | 11.15 | |
| n-Tetratriacontane | 0.10 b | 0.02 | 1.65 | 0.23 ab | 0.16 | 1.43 | 0.22 ab | 0.08 | 2.40 | 0.27 ab | 0.12 | 2.63 | 0.53 a | 0.19 | 2.47 | 0.41 ab | 0.34 | 2.48 | |
| n-Pentatriacontane | 0.23 c | 0.13 | 3.74 | 0.87 ab | 0.50 | 5.31 | 0.40 bc | 0.09 | 4.36 | 0.69 bc | 0.08 | 6.66 | 1.35 a | 0.56 | 6.31 | 0.62 bc | 0.21 | 3.71 | |
| Primary | Hexacosanol | 0.15 a | 0.14 | 2.40 | 0.48 a | 0.07 | 2.92 | 0.31 a | 0.29 | 3.39 | 0.34 a | 0.26 | 3.23 | 0.33 a | 0.13 | 1.55 | 0.69 a | 0.60 | 4.18 |
| Octacosanol | 0.10 a | 0.04 | 1.69 | 0.22 a | 0.04 | 1.35 | 0.16 a | 0.04 | 1.78 | 0.22 a | 0.01 | 2.14 | 0.32 a | 0.45 | 1.48 | 0.28 a | 0.25 | 1.70 | |
| alcohols | Triacontanol | 0 b | — | 0 | 0.87 a | — | 5.32 | 0.04 ab | 0.02 | 0.43 | 0 b | — | 0 | 0.13 ab | — | 0.60 | 0 b | — | 0 |
| Dotriacontanol | 0.14 a | 0.05 | 2.32 | 0.06 b | — | 0.40 | 0.17 a | 0.06 | 1.85 | 0.18 a | 0.04 | 1.76 | 0 b | — | 0 | 0 b | — | 0 | |
| Triterpenoids | Beta-amyrin | 0.11 a | 0.03 | 1.79 | 0.22 a | 0.07 | 1.37 | 0.33 a | 0.18 | 3.62 | 0.27 a | 0.19 | 2.60 | 0.43 a | 0.28 | 2.00 | 0.50 a | 0.38 | 3.00 |
| Alpha-amyrin | 0.13 b | 0.03 | 2.14 | 0.33 ab | 0.12 | 2.04 | 0.40 a | 0.21 | 4.42 | 0.27 ab | 0.16 | 2.57 | 0.47 a | 0.13 | 2.22 | 0.29 ab | 0.03 | 1.77 | |
| Oleanolic acid | 0.11 b | 0.08 | 1.79 | 0.41 a | 0.18 | 2.52 | 0.30 ab | 0.10 | 3.32 | 0.14 b | 0.09 | 1.33 | 0.24 ab | 0.06 | 1.11 | 0.32 ab | 0.15 | 1.93 | |
| Ursolic acid | 0.26 ab | 0.08 | 4.20 | 0.13 ab | 0.05 | 0.79 | 0.21 ab | 0.10 | 2.31 | 0.11 b | 0.02 | 1.08 | 0.16 ab | 0.03 | 0.76 | 0.34 a | 0.23 | 2.03 | |
| Corosolic acid | 0.49 a | 0.03 | 7.92 | 0.18 b | 0.09 | 1.08 | 0.49 a | 0.07 | 5.36 | 0.31 ab | 0.09 | 3.03 | 0.20 b | 0.14 | 0.92 | 0.44 ab | 0.30 | 2.68 | |
| Maslinic acid | 0.26 ab | 0.05 | 4.22 | 0.30 ab | 0.20 | 1.84 | 0.26 ab | 0.06 | 2.85 | 0.27 ab | 0.18 | 2.62 | 0.13 b | 0.07 | 0.60 | 0.53 a | 0.39 | 3.20 | |
| Total | 6.18 b | — | — | 16.39 ab | — | — | 9.11 b | — | — | 10.39 b | — | — | 21.39 a | — | — | 16.61 ab | — | — | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Guo, M.; Hu, L.; Yang, J.; Xu, J.; Rafiq, M.; Wang, Y.; Cheng, C.; Zeng, S. Preliminary Study on the Geochemical Characterization of Viticis Fructus Cuticular Waxes: From Latitudinal Variation to Origin Authentication. Int. J. Mol. Sci. 2025, 26, 7293. https://doi.org/10.3390/ijms26157293
Luo Y, Guo M, Hu L, Yang J, Xu J, Rafiq M, Wang Y, Cheng C, Zeng S. Preliminary Study on the Geochemical Characterization of Viticis Fructus Cuticular Waxes: From Latitudinal Variation to Origin Authentication. International Journal of Molecular Sciences. 2025; 26(15):7293. https://doi.org/10.3390/ijms26157293
Chicago/Turabian StyleLuo, Yiqing, Min Guo, Lei Hu, Jiaxin Yang, Junyu Xu, Muhammad Rafiq, Ying Wang, Chunsong Cheng, and Shaohua Zeng. 2025. "Preliminary Study on the Geochemical Characterization of Viticis Fructus Cuticular Waxes: From Latitudinal Variation to Origin Authentication" International Journal of Molecular Sciences 26, no. 15: 7293. https://doi.org/10.3390/ijms26157293
APA StyleLuo, Y., Guo, M., Hu, L., Yang, J., Xu, J., Rafiq, M., Wang, Y., Cheng, C., & Zeng, S. (2025). Preliminary Study on the Geochemical Characterization of Viticis Fructus Cuticular Waxes: From Latitudinal Variation to Origin Authentication. International Journal of Molecular Sciences, 26(15), 7293. https://doi.org/10.3390/ijms26157293