
Acridine Derivatives as Antifungal and Antivirulence Agents Against Candida albicans


Abstract
1. Introduction
2. Results
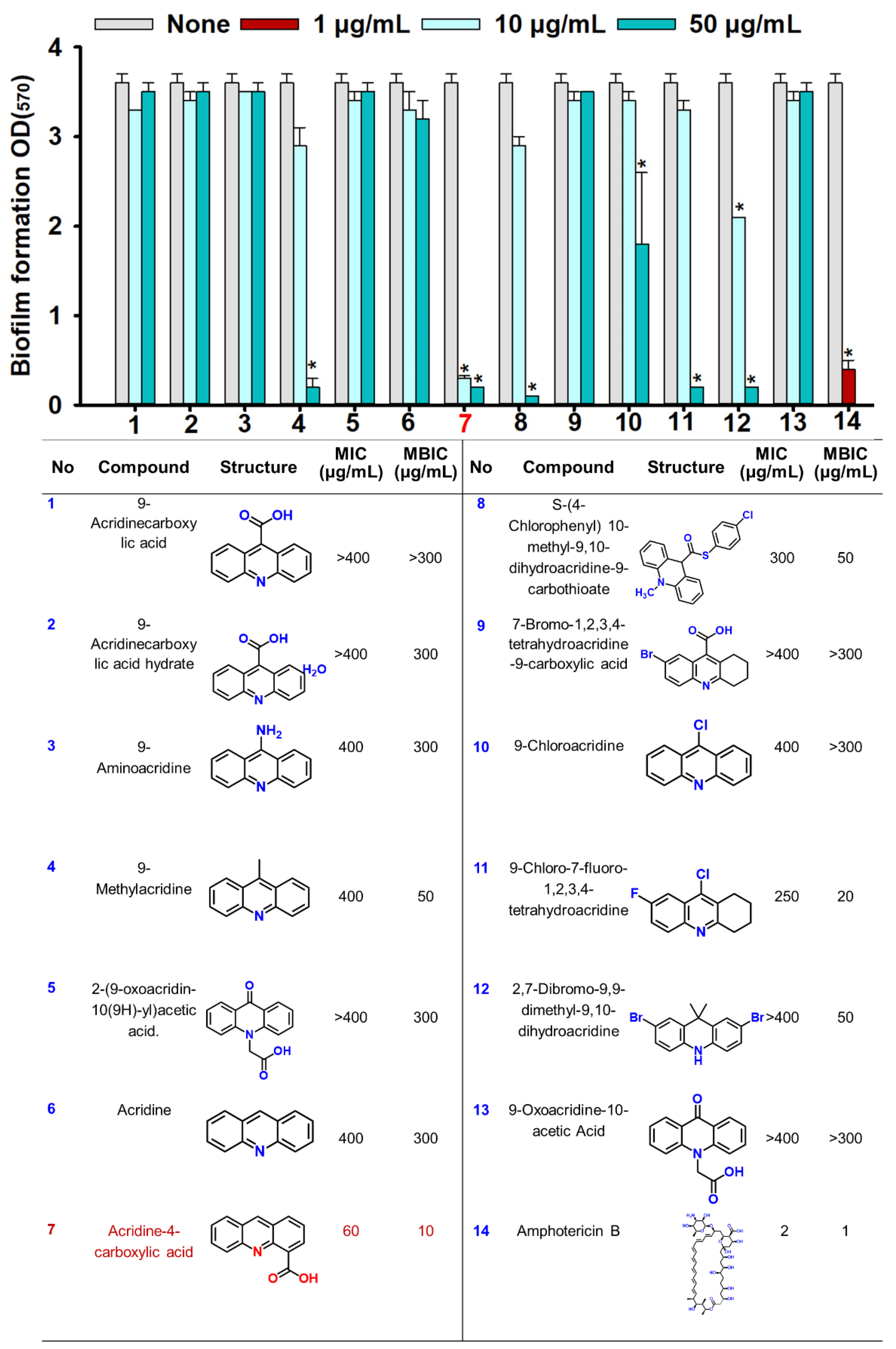
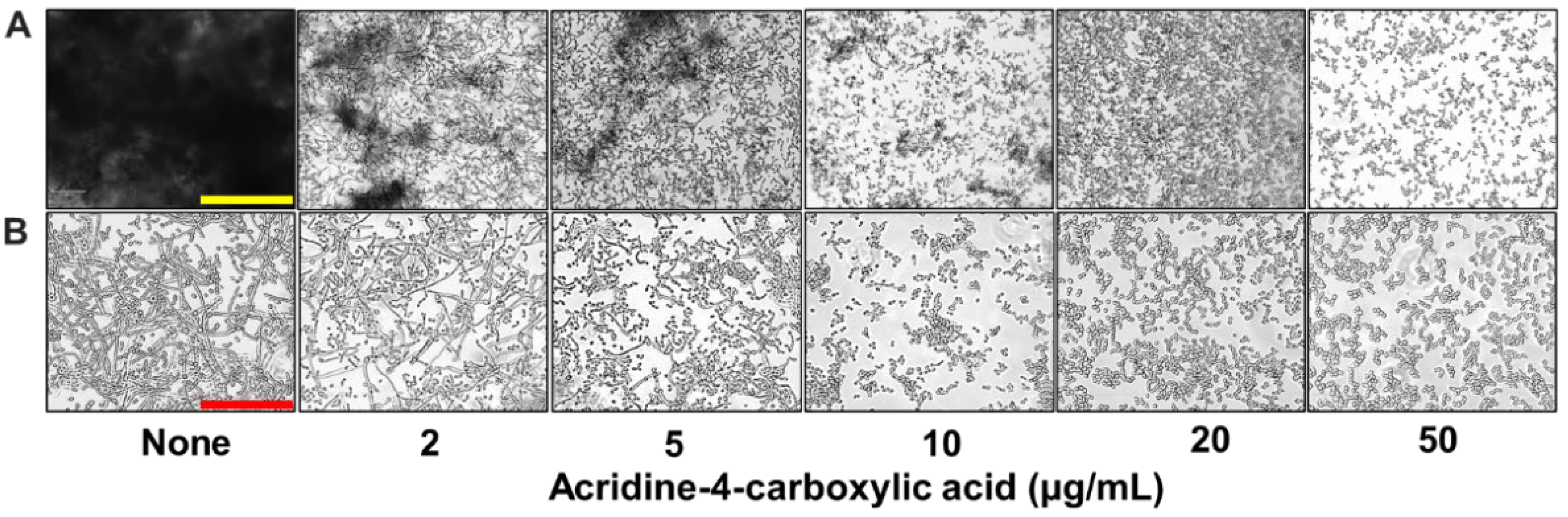
2.1. Effects of Acridine Derivatives on Biofilm Formation and Planktonic Cells of C. albicans
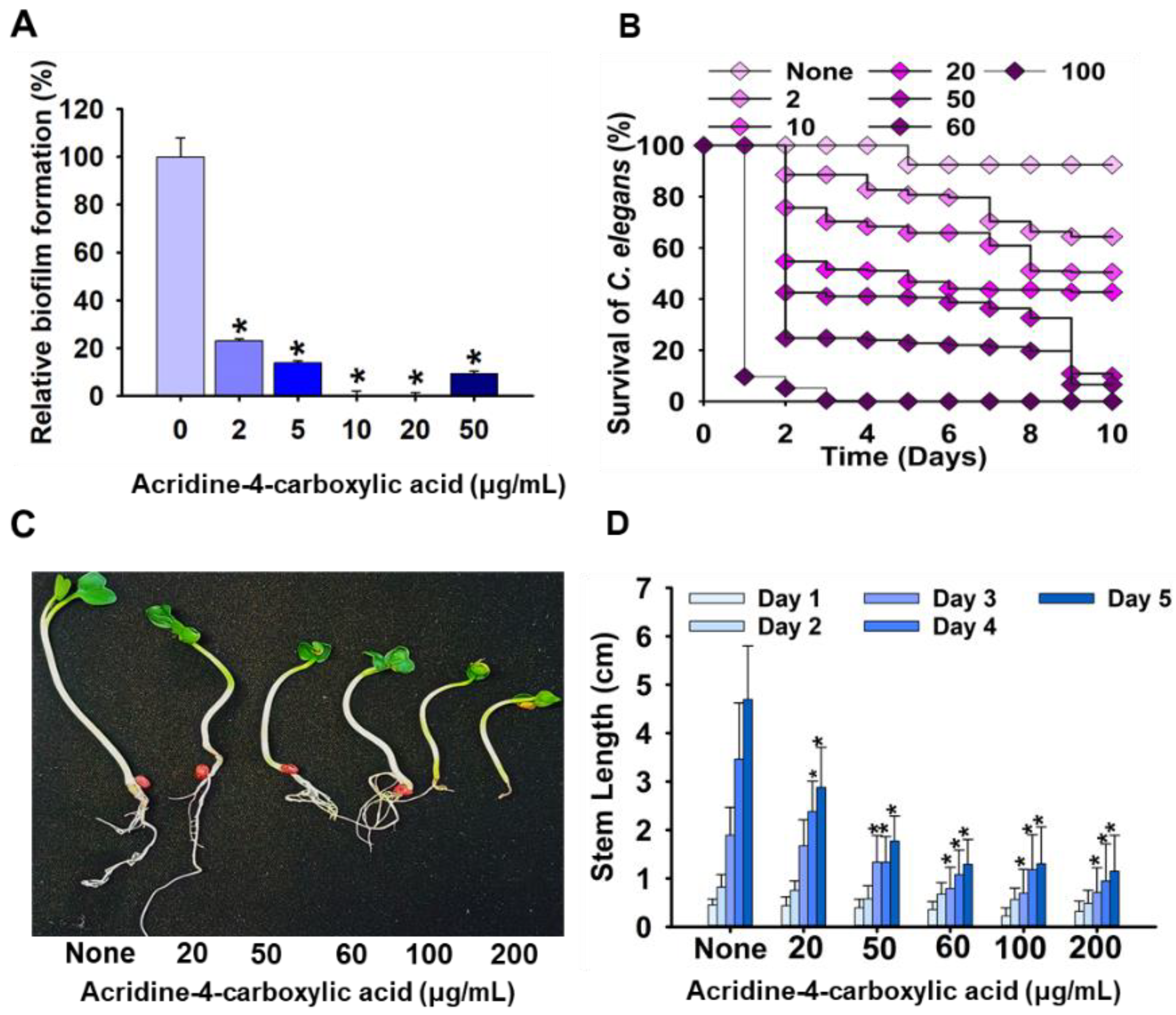
2.2. Effect of Acridine-4-carboxylic Acid on Plant Growth and Nematode Model
2.3. In Silico Absorption, Distribution, Metabolism, Excretion (ADME) Profiling of Acridine Derivatives
3. Discussion
4. Materials and Methods
4.1. Candida Strain, Growth Conditions, and Reagents
4.2. Crystal Violet Biofilm Inhibition Assay
4.3. XTT Assay for Assessing Inhibition of Early-Stage Biofilm Formation
4.4. Preformed Biofilm Dispersal Assay
4.5. Biofilm Visualization by Live Imaging and SEM
4.6. Hyphal Development and Cell Aggregation
4.7. Toxicity Studies of Acridine-4-carboxylic Acid Using Plant and Nematode Models
4.8. Predictions of Absorption, Distribution, Metabolism, and Excretion (ADME) Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.; Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. 2011, 49, 171–177. [Google Scholar] [CrossRef]
- Soriano, A.; Honore, P.M.; Puerta-Alcalde, P.; Garcia-Vidal, C.; Pagotto, A.; Gonçalves-Bradley, D.C.; Verweij, P.E. Invasive candidiasis: Current clinical challenges and unmet needs in adult populations. J. Antimicrob. Chemother. 2023, 78, 1569–1585. [Google Scholar] [CrossRef] [PubMed]
- Parslow, B.Y.; Thornton, C.R. Continuing shifts in epidemiology and antifungal susceptibility highlight the need for improved disease management of invasive candidiasis. Microorganisms 2022, 10, 1208. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans—The virulence factors and clinical manifestations of infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Karajacob, A.S.; Azizan, N.B.; Al-Maleki, A.R.M.; Goh, J.P.E.; Loke, M.F.; Khor, H.M.; Ho, G.F.; Ponnampalavanar, S.; Tay, S.T. Candida species and oral mycobiota of patients clinically diagnosed with oral thrush. PLoS ONE 2023, 18, e0284043. [Google Scholar] [CrossRef]
- Dühring, S.; Germerodt, S.; Skerka, C.; Zipfel, P.F.; Dandekar, T.; Schuster, S. Host-pathogen interactions between the human innate immune system and Candida albicans—Understanding and modeling defense and evasion strategies. Front. Microbiol. 2015, 6, 625. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.; Kumar, A. Systemic Candida albicans Infection in Mice Causes Endogenous Endophthalmitis via Breaching the Outer Blood-Retinal Barrier. Microbiol. Spectr. 2022, 10, e01658-22. [Google Scholar] [CrossRef]
- Evans, S.E. Coping with Candida infections. Proc. Am. Thorac. Soc. 2010, 7, 197–203. [Google Scholar] [CrossRef]
- Sims, C.R.; Ostrosky-Zeichner, L.; Rex, J.H. Invasive candidiasis in immunocompromised hospitalized patients. Arch. Med. Res. 2005, 36, 660–671. [Google Scholar] [CrossRef]
- Priya, A.; Pandian, S.K. Piperine Impedes Biofilm Formation and Hyphal Morphogenesis of Candida albicans. Front. Microbiol. 2020, 11, 756. [Google Scholar] [CrossRef]
- Wongsuk, T.; Pumeesat, P.; Luplertlop, N. Fungal quorum sensing molecules: Role in fungal morphogenesis and pathogenicity. J. Basic Microbiol. 2016, 56, 440–447. [Google Scholar] [CrossRef]
- Van Der Veen, S.; Tang, C.M. The BER necessities: The repair of DNA damage in human-adapted bacterial pathogens. Nat. Rev. Microbiol. 2015, 13, 83–94. [Google Scholar] [CrossRef]
- Wall, G.; Montelongo-Jauregui, D.; Bonifacio, B.V.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single-and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; Lopez-Ribot, J.L. Candida biofilms: An update. Eukaryot. Cell 2005, 4, 633–638. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, J.; Zhang, Q.; Song, Z.; Xin, C. Antifungal activities of Equol against Candida albicans in vitro and in vivo. Virulence 2024, 15, 2404256. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.V.; Mitchell, A.P.; Andes, D.R. Fungal biofilms, drug resistance, and recurrent infection. Cold Spring Harb. Perspect. Med. 2014, 4, a019729. [Google Scholar] [CrossRef] [PubMed]
- Sroor, F.M.; Younis, A.; Abdelraof, M.; Abdelhamid, I.A. Synthesis, molecular docking and anti-biofilm activity of novel benzo[4,5]imidazo[2,1-a]quinazoline, 4H-chromene, and acridine derivatives as potent anti-candida agents. J. Mol. Struct. 2025, 1331, 141520. [Google Scholar] [CrossRef]
- Yonker, M. Secondary headaches in children and adolescents: What not to miss. Curr. Neurol. Neurosci. Rep. 2018, 18, 61. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.R. Lyme neuroborreliosis. Contin. Lifelong Learn. Neurol. 2015, 21, 1729–1744. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar] [CrossRef] [PubMed]
- Taff, H.T.; Mitchell, K.F.; Edward, J.A.; Andes, D.R. Mechanisms of Candida biofilm drug resistance. Future Microbiol. 2013, 8, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, I. ‘Acridines’ as new horizons in antifungal treatment. J. Mol. 2020, 25, 1480. [Google Scholar] [CrossRef]
- Jalab, M. The Development of Photo-Activated Antimicrobial Dyes Against Opportunistic Infections. Ph.D. Thesis, University of Central Lancashire, Preston, UK, 2020. [Google Scholar]
- Romo, J.A.; Pierce, C.G.; Esqueda, M.; Hung, C.-Y.; Saville, S.P.; Lopez-Ribot, J.L. In Vitro Characterization of a Biaryl Amide Anti-virulence Compound Targeting Candida albicans Filamentation and Biofilm Formation. Front. Cell. Infect. Microbiol. 2018, 8, 227. [Google Scholar] [CrossRef]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef]
- Ježek, J.; Hlaváček, J.; Šebestík, J. Biomedical Applications of Acridines; Springer: Prague, Czech Republic, 2017; Volume 72, XIII, 237. [Google Scholar]
- Dudek, A.; Szulc, N.; Pawlak, A.; Strugała-Danak, P.; Krawczyk-Łebek, A.; Perz, M.; Kostrzewa-Susłow, E.; Pruchnik, H. Structural investigation of interactions between halogenated flavonoids and the lipid membrane along with their role as cytotoxic agents. Sci. Rep. 2024, 14, 10561. [Google Scholar] [CrossRef]
- Rząd, K.; Paluszkiewicz, E.; Gabriel, I. A new 1-nitro-9-aminoacridine derivative targeting yeast topoisomerase II able to overcome fluconazole-resistance. Bioorg. Med. Chem. Lett. 2021, 35, 127815. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. 2011, 75, 583–609. [Google Scholar] [CrossRef]
- Abirami, G.; Alexpandi, R.; Durgadevi, R.; Kannappan, A.; Veera Ravi, A. Inhibitory Effect of Morin Against Candida albicans Pathogenicity and Virulence Factor Production: An in vitro and in vivo Approaches. Front. Microbiol. 2020, 11, 561298. [Google Scholar] [CrossRef]
- Shareck, J.; Belhumeur, P. Modulation of morphogenesis in Candida albicans by various small molecules. Eukaryot. Cell 2011, 10, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Jothi, R.; Sangavi, R.; Kumar, P.; Pandian, S.K.; Gowrishankar, S. Catechol thwarts virulent dimorphism in Candida albicans and potentiates the antifungal efficacy of azoles and polyenes. Sci. Rep. 2021, 11, 21049. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, C.d.R.E.; Galvão, D.S.; do Desterro Cunha, S.; Fonseca de Freitas, H.; Barros, T.F. Thiosemicarbazones and analogues as potential biofilm inhibitors of Candida albicans. Biofouling 2025, 41, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Subramenium, G.A.; Swetha, T.K.; Iyer, P.M.; Balamurugan, K.; Pandian, S.K. 5-hydroxymethyl-2-furaldehyde from marine bacterium Bacillus subtilis inhibits biofilm and virulence of Candida albicans. Microbiol. Res. 2018, 207, 19–32. [Google Scholar] [CrossRef]
- Rajasekharan, S.K.; Kamalanathan, C.; Ravichandran, V.; Ray, A.K.; Satish, A.S.; Mohanvel, S.K. Mannich base limits Candida albicans virulence by inactivating Ras-cAMP-PKA pathway. Sci. Rep. 2018, 8, 14972. [Google Scholar] [CrossRef]
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic understanding of Candida albicans biofilm formation and approaches for its inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef]
- Gupta, P.; Pruthi, P.A.; Pruthi, V. Role of exopolysaccharides in biofilm formation. In Introduction to Biofilm Engineering; ACS Publications: Washington, DC, USA, 2019; pp. 17–57. [Google Scholar]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Pierce, C.G.; Chaturvedi, A.K.; Lazzell, A.L.; Powell, A.T.; Saville, S.P.; McHardy, S.F.; Lopez-Ribot, J.L. A novel small molecule inhibitor of Candida albicans biofilm formation, filamentation and virulence with low potential for the development of resistance. npj Biofilms Microbiomes 2015, 1, 15012. [Google Scholar] [CrossRef]
- Sharma, J.; Rosiana, S.; Razzaq, I.; Shapiro, R.S. Linking cellular morphogenesis with antifungal treatment and susceptibility in Candida pathogens. J. Fungi 2019, 5, 17. [Google Scholar] [CrossRef]
- Ferenc, T.; Janik-Spiechowicz, E.; Bratkowska, W.; Łopaczyńska, D.; Stróżyński, H.; Denys, A.; Mordalska, A. Genotoxicity assessment of new synthesized acridine derivative—3,6-diamino-10-methyl-9, 10-dihydroacridine. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1999, 444, 463–470. [Google Scholar] [CrossRef]
- Hsu, K.-H.; Su, B.-H.; Tu, Y.-S.; Lin, O.A.; Tseng, Y.J. Mutagenicity in a molecule: Identification of core structural features of mutagenicity using a scaffold analysis. PLoS ONE 2016, 11, e0148900. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, C.; Nikoyan, A.; Decome, L.; Botta, C.; Robin, M.; Reboul, J.-P.; Sabatier, A.-S.; Matta, A.; De Méo, M. DNA-damaging activity and mutagenicity of 16 newly synthesized thiazolo[5,4-a]acridine derivatives with high photo-inducible cytotoxicity. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2008, 650, 104–114. [Google Scholar] [CrossRef] [PubMed]
- PubChem. Compound Summary for CID 11138739, Acridine-4-Carboxylic Acid. PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Acridine-4-carboxylic-Acid (accessed on 10 July 2025).
- Denny, W.A. Acridine-4-carboxamides and the concept of minimal DNA intercalators. In Small Molecule DNA/RNA Binders: From Synthesis to Nucleic Acid Complexes; Wiley: Hoboken, NJ, USA, 2002; pp. 482–502. [Google Scholar] [CrossRef]
- Sabarees, G.; Tamilarasi, G.P.; Alagarsamy, V.; Kandhasamy, S.; Gouthaman, S.; Solomon, V.R. An overview of acridine analogs: Pharmacological significance and recent developments. Curr. Med. Chem. 2025, 32, 5226–5259. [Google Scholar] [CrossRef]
- Rupar, J.; Dobričić, V.; Aleksić, M.; Brborić, J.; Čudina, O. A review of published data on acridine derivatives with different biological activities. Kragujev. J. Sci. 2018, 40, 83–101. [Google Scholar] [CrossRef]
- Bartoszek, A.; Dackiewicz, P.; Składanowski, A.; Konopa, J. In vitro DNA crosslinking by Ledakrin, an antitumor derivative of 1-nitro-9-aminoacridine. Chem. Biol. Interact. 1997, 103, 141–151. [Google Scholar] [CrossRef]
- Sharma, V.; Sharma, P.C.; Kumar, V. A mini review on pyridoacridines: Prospective lead compounds in medicinal chemistry. J. Adv. Res. 2015, 6, 63–71. [Google Scholar] [CrossRef]
- Lipinski, C.A. Lead-and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Sousa, G.; de Almeida, M.C.; Lócio, L.L.; Santos, V.L.d.; Bezerra, D.P.; Silva, V.R.; Almeida, S.; Simon, A.; Honório, T.d.S.; Cabral, L.M. Synthesis and evaluation of antiproliferative activity, topoisomerase II inhibition, DNA binding and non-clinical toxicity of new acridine–thiosemicarbazone derivatives. Pharmaceuticals 2022, 15, 1098. [Google Scholar] [CrossRef]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef]
- Aydin, M.; Unusan, N.; Sumlu, E.; Korucu, E.N. Rosmarinic acid exhibits antifungal and antibiofilm activities against Candida albicans: Insights into gene expression and morphological changes. J. Fungi 2024, 10, 751. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Khadke, S.K.; Yamano, A.; Watanabe, A.; Lee, J. Inhibition of biofilm formation by Candida albicans and polymicrobial microorganisms by nepodin via hyphal-growth suppression. ACS Infect. Dis. 2019, 5, 1177–1187. [Google Scholar] [CrossRef]
- Wayne, P. Clinical and laboratory Standards Institute (CLSI) (2020) Performance standards for antimicrobial susceptibility testing: Background, Organization, Functions, and Processes. J. Clin. Microbiol. 2020. [Google Scholar]
- Fayed, B.; El-Sayed, H.S.; Luo, S.; Reda, A.E. Comparative evaluation of biologically and chemically synthesized zinc oxide nanoparticles for preventing Candida auris biofilm. Biometals 2025, 38, 817–830. [Google Scholar] [CrossRef]
- Sharifi, A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P.; Nourian, A. Cuminum cyminum L. essential oil: A promising antibacterial and antivirulence agent against multidrug-resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 667833. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; De Luca, A.; Arroyo, J.; Blanco, N.; Servillo, G.; Sanglard, D.; Reichard, U.; Palmer, G.E.; Latgè, J.-P. Sensing of mammalian IL-17A regulates fungal adaptation and virulence. Nat. Commun. 2012, 3, 683. [Google Scholar] [CrossRef]
- Sathiyamoorthi, E.; Faleye, O.S.; Lee, J.-H.; Lee, J. Hydroquinone derivatives attenuate biofilm formation and virulence factor production in Vibrio spp. Int. J. Food Microbiol. 2023, 384, 109954. [Google Scholar] [CrossRef]
- Bezerra Filho, C.M.; da Silva, L.C.N.; da Silva, M.V.; Løbner-Olesen, A.; Struve, C.; Krogfelt, K.A.; Correia, M.T.d.S.; Vilela Oliva, M.L. Antimicrobial and Antivirulence Action of Eugenia brejoensis Essential Oil in vitro and in vivo Invertebrate Models. Front. Microbiol. 2020, 11, 424. [Google Scholar] [CrossRef]
- Zarroug, S.H.; Bajaman, J.S.; Hamza, F.N.; Saleem, R.A.; Abdalla, H.K. Caenorhabditis elegans as an in vivo model for the discovery and development of natural plant-based antimicrobial compounds. Pharmaceuticals 2023, 16, 1070. [Google Scholar] [CrossRef]
- Dulsat, J.; López-Nieto, B.; Estrada-Tejedor, R.; Borrell, J.I. Evaluation of free online ADMET tools for academic or small biotech environments. Molecules 2023, 28, 776. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Acridine | Acridine-4-carboxylic Acid | 2,7-Dibromo-9,9-dimethyl-9,10-dihydroacridine |
|---|---|---|---|
| Lipinski (Pfizer) rule of five | Suitable | Suitable | Suitable |
| Lipinski rule of five violations | 0 violation | 0 violation | 1 violation |
| Veber (GSK) rule | Suitable | Suitable | Suitable |
| Muegge (Bayer) rule | Likely Suitable | Likely Suitable | Likely Suitable |
| Plasma protein binding | 97.8 | 92.8 | 98.8 |
| Blood–brain barrier permeability | 3.0 | 2.3 | 15.1 |
| Lipophilicity (iLOGP) | Not Available | Not Available | Not Available |
| Water Solubility (Log S) | Moderately Soluble | Moderately Soluble | Poorly Soluble |
| Pgp Substrate_inhibition | Non-Inhibitor | Non-Inhibitor | Inhibitor |
| CYP1A2 inhibitor | Non-Inhibitor | Non-Inhibitor | Non-Inhibitor |
| CYP3A4 inhibitor | Non-Inhibitor | Non-Inhibitor | Non-Inhibitor |
| Skin_Permeability (logKp, cm/hour) | −2.7 | −3.30 | −1.5 |
| Gastrointestinal intestinal absorption | Moderate | Low | Low |
| Caco2 | 30.3 | 2.9 | 55.1 |
| Mouse carcinogenicity | Positive | Positive | Positive |
| Rat carcinogenicity | Negative | Negative | Negative |
| Acute algae toxicity | 0.10 | 0.09 | 0.01 |
| Acute fish toxicity (medaka) | 0.02 | 0.01 | 0 |
| Acute fish toxicity (minnow) | 0.01 | 0.01 | 0 |
| In vitro hERG inhibition | Medium_risk | Medium_risk | Medium_risk |
| miLogP | 3.1 | 2.69 | 6 |
| Mol volume | 167.8 | 194.8 | 242.4 |
| TPSA | 12.8 | 50.1 | 12 |
| GPCR ligand | Inactive | Inactive | Inactive |
| Ion channel modulator | Inactive | Inactive | Inactive |
| Kinase inhibitor | Potential Inhibitor | Potential Inhibitor | Inactive |
| Nuclear receptor ligand | Inactive | Inactive | Inactive |
| Protease inhibitor | Inactive | Inactive | Inactive |
| Enzyme inhibitor | Possible | Possible | Inactive |
| Rat IP LD50 classification | Class IV in AD | Class V in AD | Class V in AD |
| Rat IV LD50 classification | Class IV in AD | Class IV in AD | Non-Toxic |
| Rat oral LD50 classification | Class IV in AD | Class IV in AD | Class IV in AD |
| Rat SC LD50 classification | Class IV in AD | Class V in AD | Class V in AD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunus, A.; Faleye, O.O.; Lee, J.-H.; Lee, J. Acridine Derivatives as Antifungal and Antivirulence Agents Against Candida albicans. Int. J. Mol. Sci. 2025, 26, 7228. https://doi.org/10.3390/ijms26157228
Yunus A, Faleye OO, Lee J-H, Lee J. Acridine Derivatives as Antifungal and Antivirulence Agents Against Candida albicans. International Journal of Molecular Sciences. 2025; 26(15):7228. https://doi.org/10.3390/ijms26157228
Chicago/Turabian StyleYunus, Amra, Oluwatosin Oluwaseun Faleye, Jin-Hyung Lee, and Jintae Lee. 2025. "Acridine Derivatives as Antifungal and Antivirulence Agents Against Candida albicans" International Journal of Molecular Sciences 26, no. 15: 7228. https://doi.org/10.3390/ijms26157228
APA StyleYunus, A., Faleye, O. O., Lee, J.-H., & Lee, J. (2025). Acridine Derivatives as Antifungal and Antivirulence Agents Against Candida albicans. International Journal of Molecular Sciences, 26(15), 7228. https://doi.org/10.3390/ijms26157228