
In Silico and In Vitro Potential Antifungal Insights of Insect-Derived Peptides in the Management of Candida sp. Infections

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. In Silico Analysis and In Vitro Tests
2.1.1. Antifungal Insect Peptide Library Preparation from Existing Reports
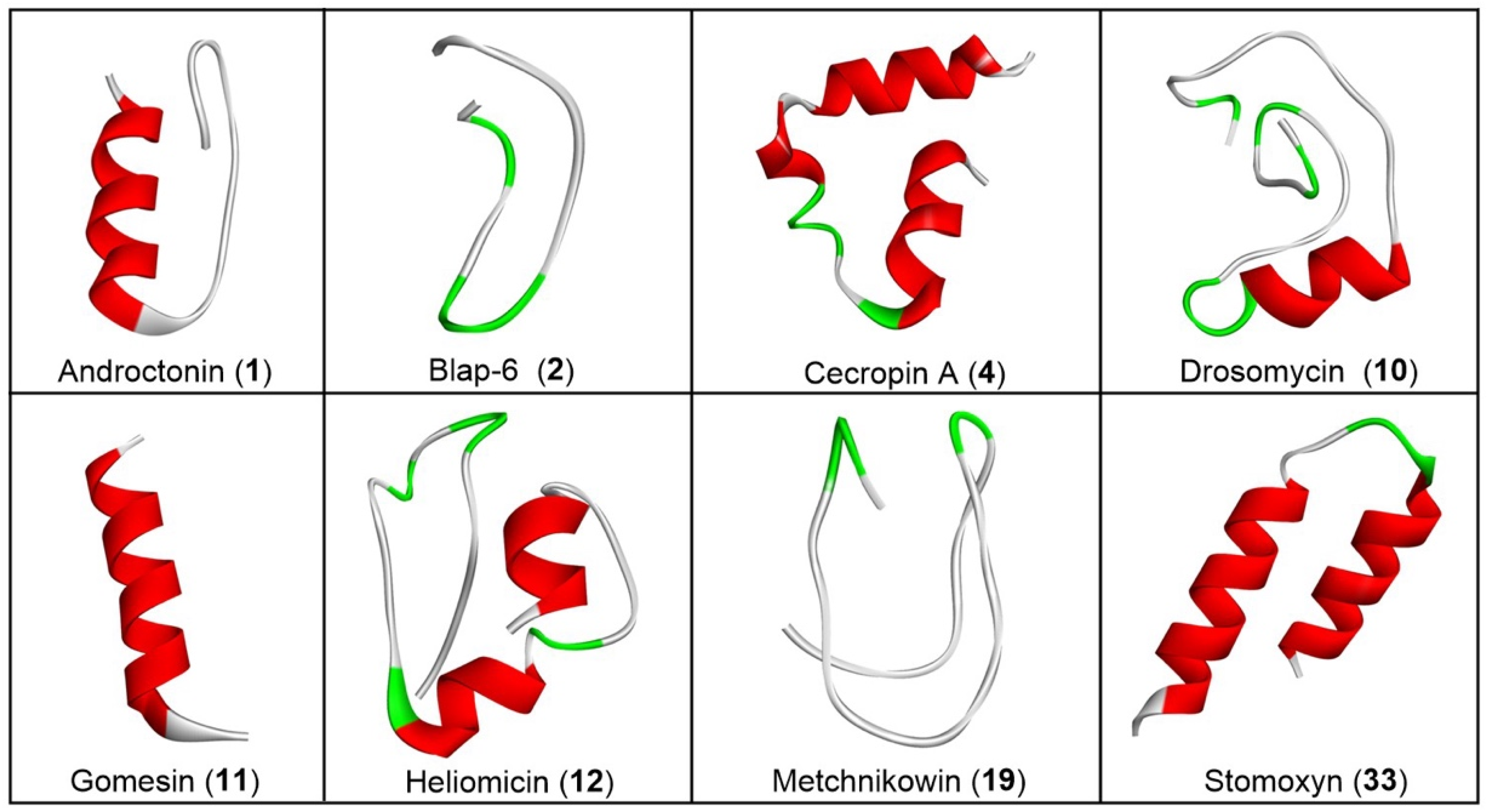
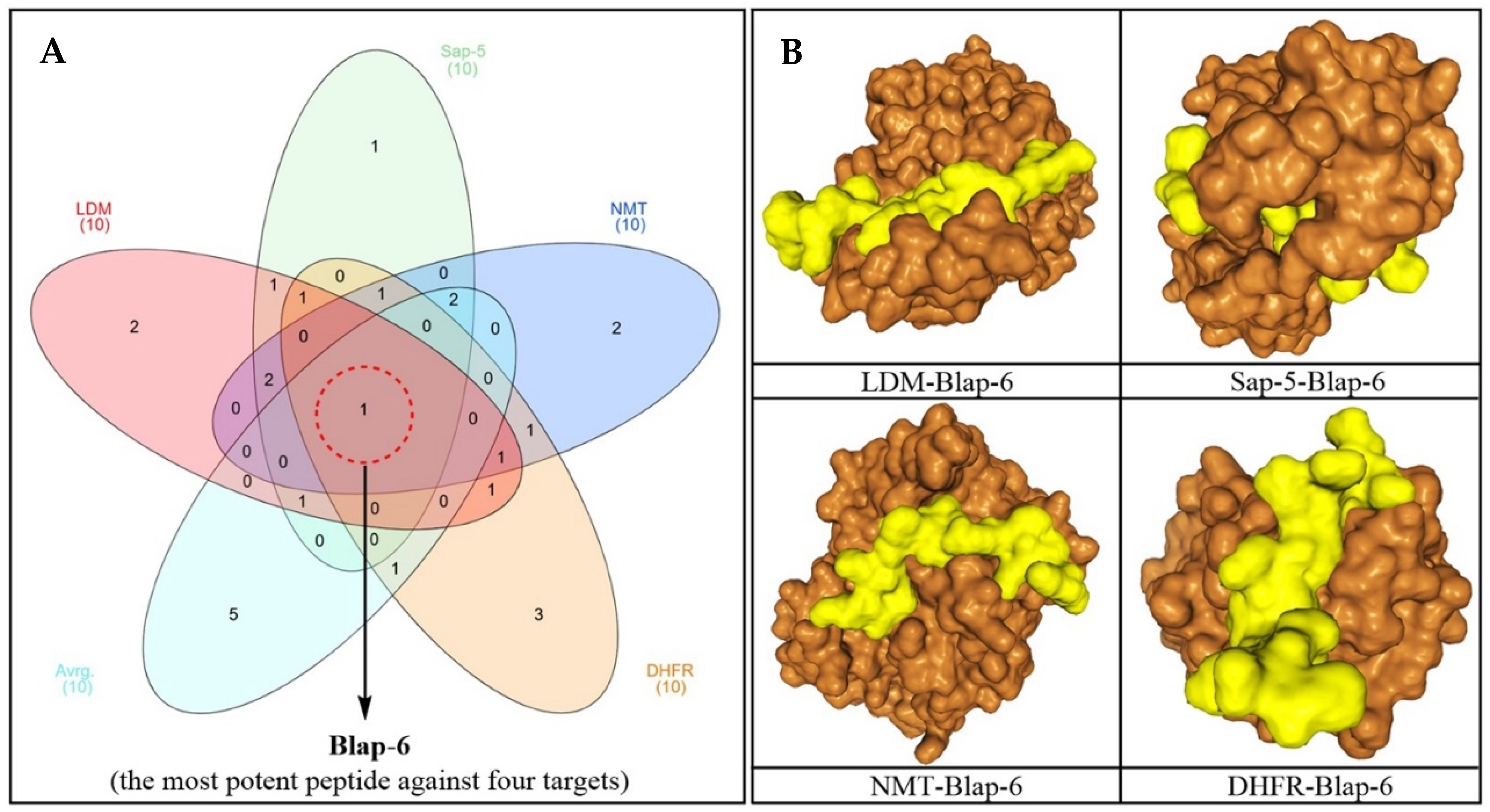
2.1.2. Three-Dimensional Peptide Structure Modelling and Molecular Docking Study
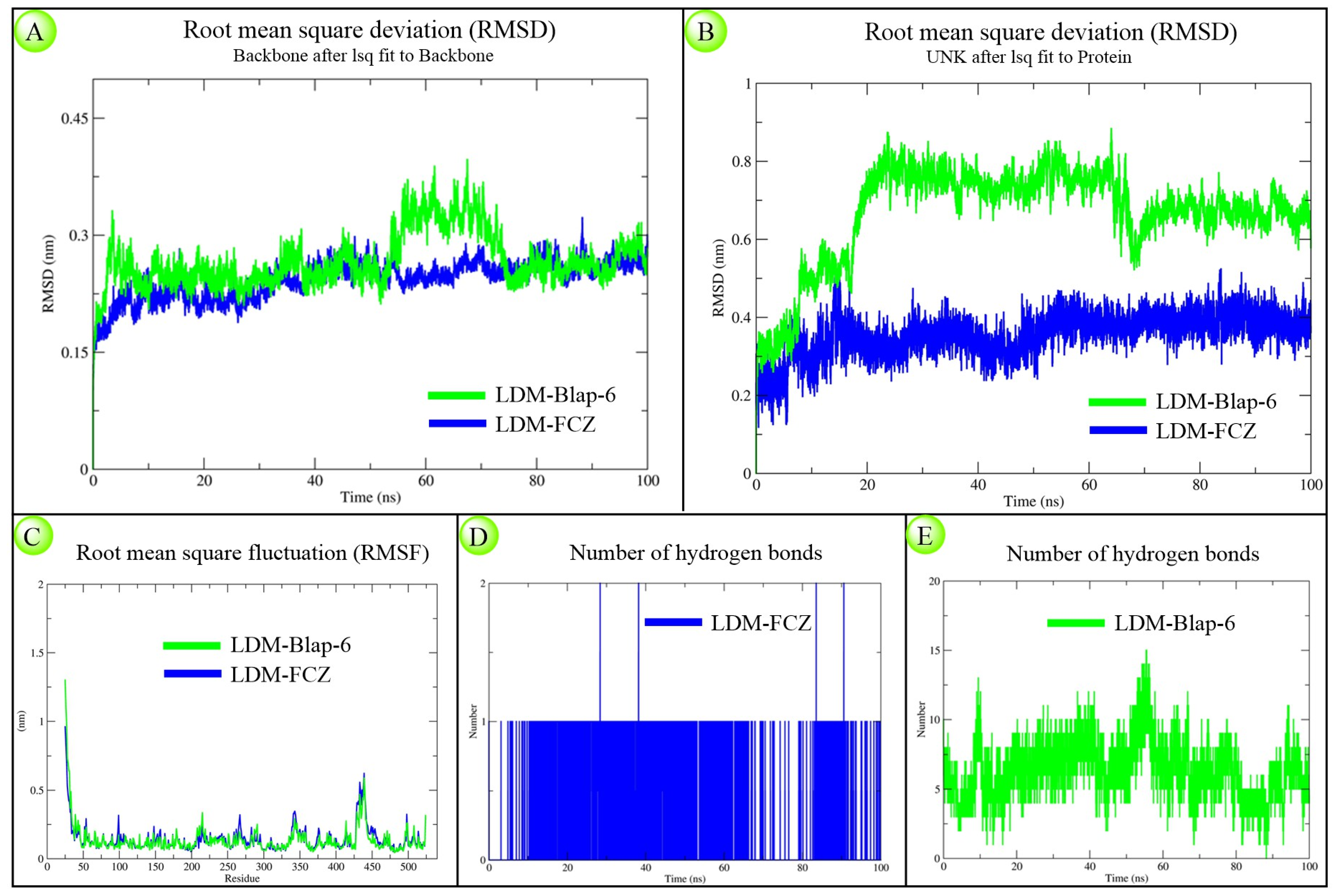
2.1.3. Molecular Dynamics Simulation Study
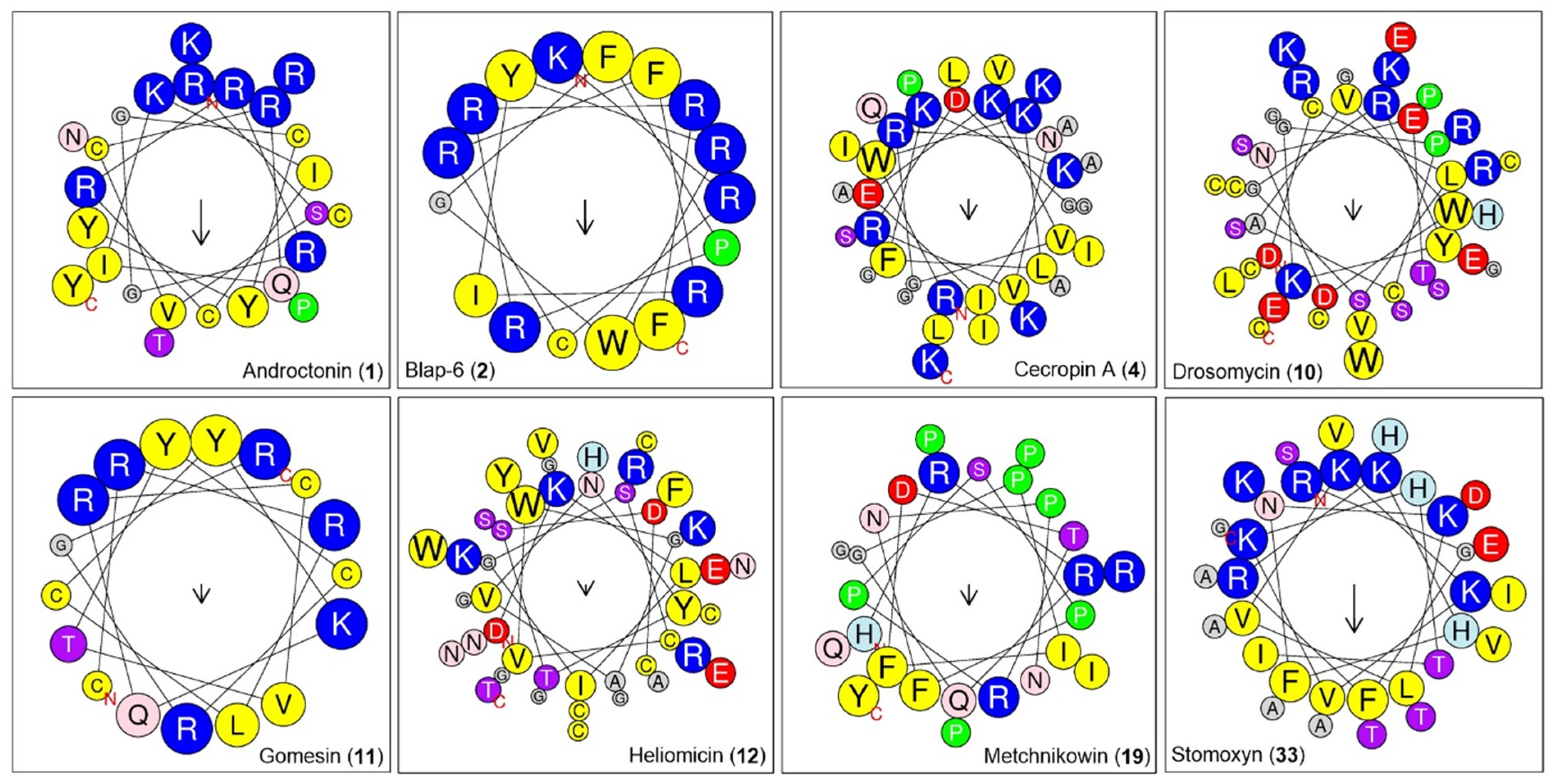
2.1.4. Physicochemical Property Analysis of Peptides
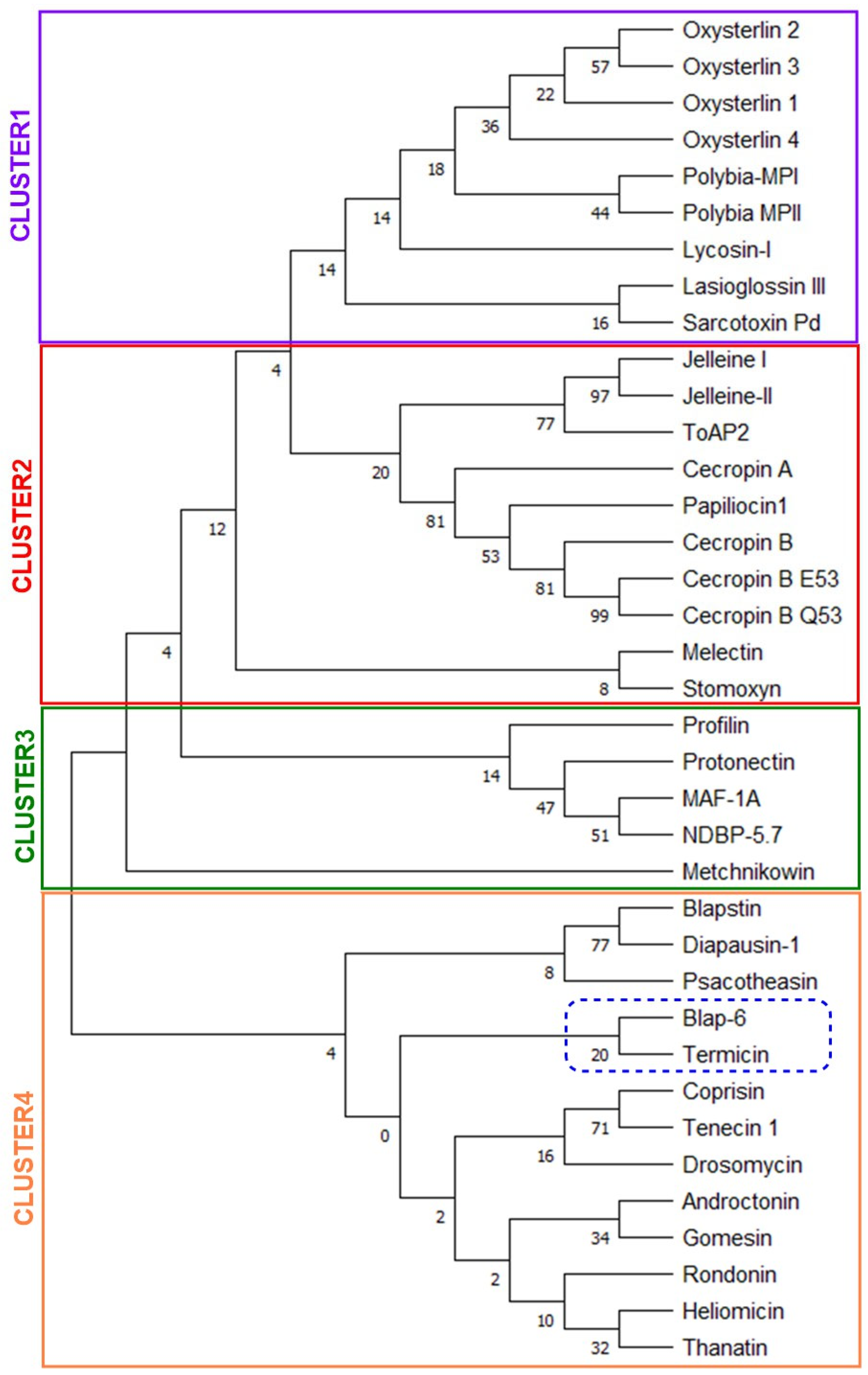
2.1.5. Homologues-Cum-Phylogenetic Tree Analysis of Peptides
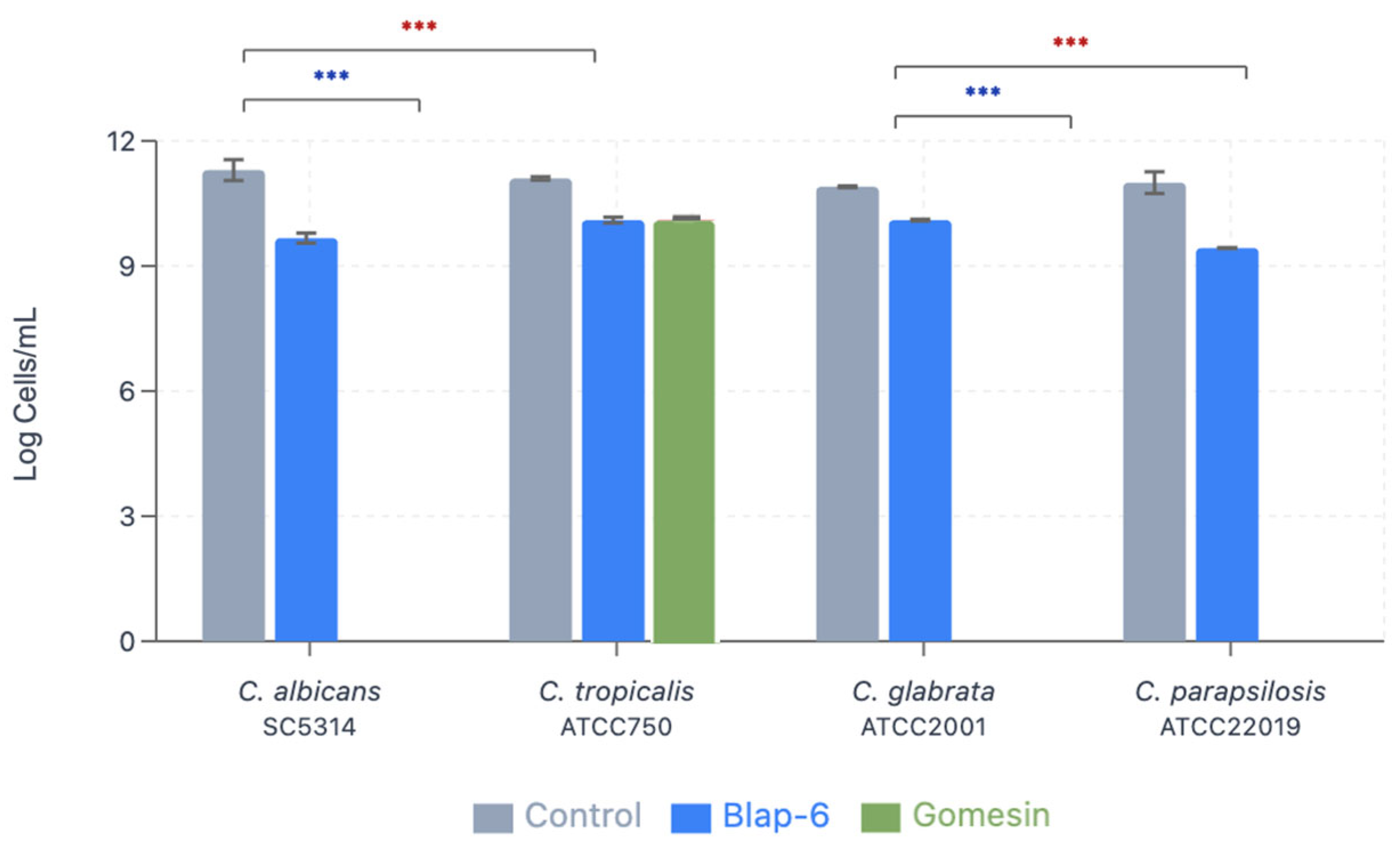
2.1.6. Minimum Inhibitory Concentration and Minimum Fungicidal Concentration
2.1.7. Minimum Biofilm Eradication Concentration
2.1.8. Biomass Production Quantification
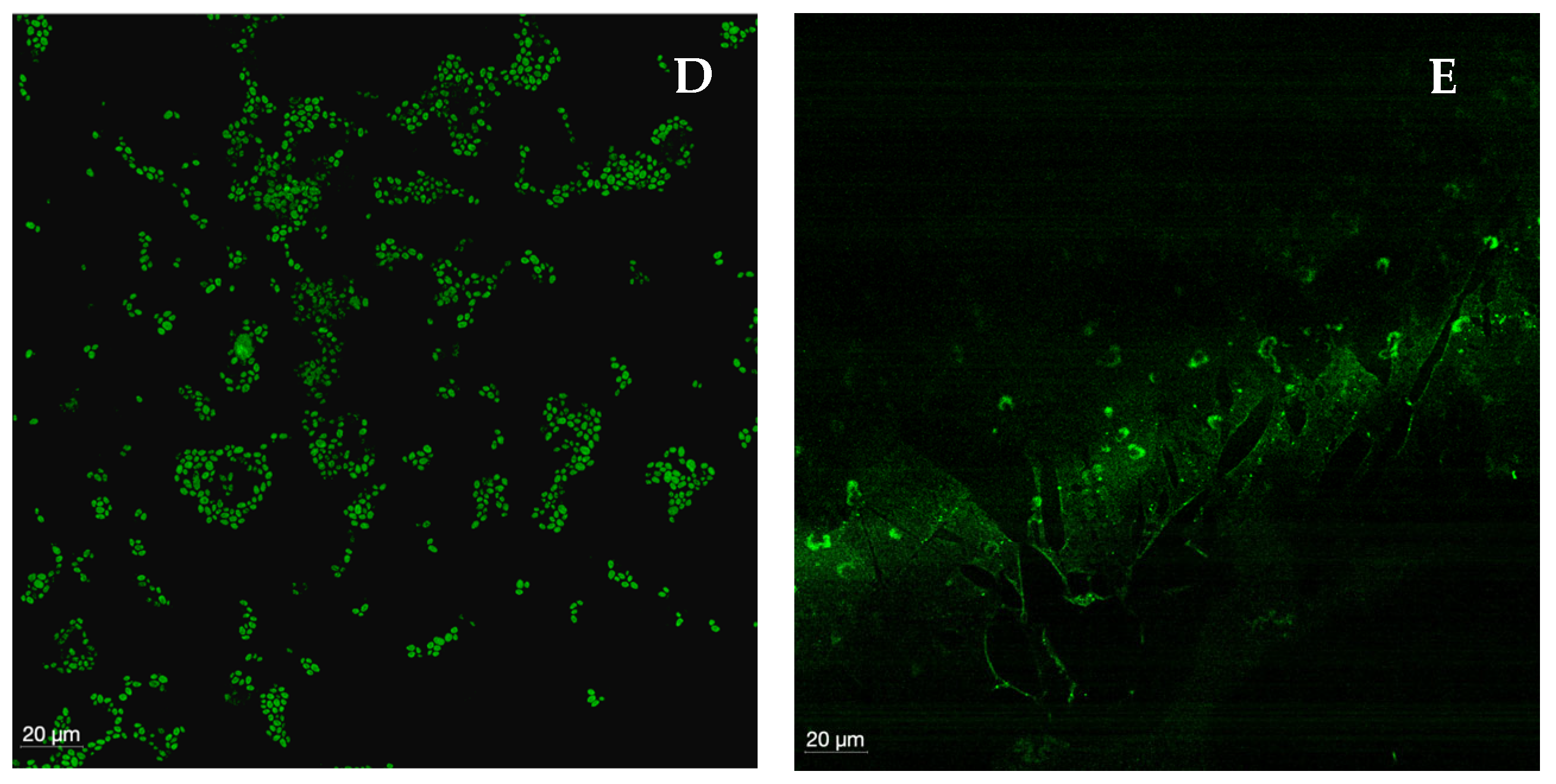
2.1.9. Confocal Laser Scanning Microscopy
3. Discussion
4. Materials and Methods
4.1. In Silico Analysis
4.1.1. Antifungal Insect Peptides Library Report Selection
4.1.2. Three-Dimensional Peptide Structure Modelling and Molecular Docking Study
4.1.3. Molecular Dynamics Simulation Study
4.1.4. Physicochemical Property Analysis of Peptides
4.1.5. Homologues-Cum-Phylogenetic Tree Analysis of Peptides
4.2. In Vitro Assays
4.2.1. Organisms and Growth Conditions
4.2.2. Insect Peptides
4.2.3. Minimum Inhibitory Concentration and Minimum Fungicidal Concentration
4.2.4. Minimum Biofilm Eradication Concentration
4.2.5. Biomass Production Quantification
4.2.6. Confocal Laser Scanning Microscopy (CLSM)
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and Virulence of Candida Albicans. Virulence 2022, 13, 89–121. [Google Scholar] [CrossRef]
- d’Enfert, C.; Janbon, G. Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches? FEMS Yeast Res. 2016, 16, fov111. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.; Honore, P.M.; Puerta-Alcalde, P.; Garcia-Vidal, C.; Pagotto, A.; Gonçalves-Bradley, D.C.; Verweij, P.E. Invasive Candidiasis: Current Clinical Challenges and Unmet Needs in Adult Populations. J. Antimicrob. Chemother. 2023, 78, 1569–1585. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Kanj, S.S.; Govender, N.P.; Thompson, G.R.; Ostrosky-Zeichner, L.; Govrins, M.A. Invasive Candidiasis. Nat. Rev. Dis. Primers 2024, 10, 1–18. [Google Scholar] [CrossRef]
- WHO. WHO Fungal Pathogens Priority List; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Beardsley, J.; Halliday, C.L.; Chen, S.C.A.; Sorrell, T.C. Responding to the Emergence of Antifungal Drug Resistance: Perspectives from the Bench and the Bedside. Future Microbiol. 2018, 13, 1175–1191. [Google Scholar] [CrossRef]
- Logan, A.; Wolfe, A.; Williamson, J.C. Antifungal Resistance and the Role of New Therapeutic Agents. Curr. Infect. Dis. Rep. 2022, 24, 105–116. [Google Scholar] [CrossRef]
- De Oliveira, H.C.; Bezerra, B.T.; Rodrigues, M.L. Antifungal Development and the Urgency of Minimizing the Impact of Fungal Diseases on Public Health. ACS Bio Med. Chem. Au 2022, 3, 137–146. [Google Scholar] [CrossRef]
- Pereira, R.; dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida Albicans: Formation, Regulation and Resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida Albicans Biofilms: Development, Regulation, and Molecular Mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]
- Cui, X.; Wang, L.; Lü, Y.; Yue, C. Development and Research Progress of Anti-Drug Resistant Fungal Drugs. J. Infect. Public Health 2022, 15, 986–1000. [Google Scholar] [CrossRef]
- Augostine, C.R.; Avery, S.V. Discovery of Natural Products with Antifungal Potential Through Combinatorial Synergy. Front. Microbiol. 2022, 13, 866840. [Google Scholar] [CrossRef]
- Shariati, A.; Didehdar, M.; Razavi, S.; Heidary, M.; Soroush, F.; Chegini, Z. Natural Compounds: A Hopeful Promise as an Antibiofilm Agent Against Candida Species. Front. Pharmacol. 2022, 13, 917787. [Google Scholar] [CrossRef]
- Roe, K. Treatment Alternatives for Multidrug-Resistant Fungal Pathogens. Drug Discov. Today 2023, 28, 103596. [Google Scholar] [CrossRef] [PubMed]
- Najafian, L.; Babji, A.S. A Review of Fish-Derived Antioxidant and Antimicrobial Peptides: Their Production, Assessment, and Applications. Peptides 2012, 33, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the Role of Antimicrobial Peptides in Insects. Int. J. Mol. Sci. 2023, 24, 5753. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Ahmed, I.; Asgher, M.; Sher, F.; Hussain, S.M.; Nazish, N.; Joshi, N.; Sharma, A.; Parra-Saldívar, R.; Bilal, M.; Iqbal, H.M.N. Exploring Marine as a Rich Source of Bioactive Peptides: Challenges and Opportunities from Marine Pharmacology. Mar. Drugs 2022, 20, 208. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant Antimicrobial Peptides: An Overview about Classification, Toxicity and Clinical Applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Balandin, S.V.; Belogurova-Ovchinnikova, O.Y.; Ovchinnikova, T.V. Marine Invertebrate Antimicrobial Peptides and Their Potential as Novel Peptide Antibiotics. Mar. Drugs 2023, 21, 503. [Google Scholar] [CrossRef]
- Ganeshkumar, A.; Gonçale, J.C.; Rajaram, R.; Junqueira, J.C. Anti-Candidal Marine Natural Products: A Review. J. Fungi 2023, 9, 800. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y.; Makovitzky, A.; Avrahami, D.; Makovitzki, A. Host Defense Peptides and Lipopeptides: Modes of Action and Potential Candidates for the Treatment of Bacterial and Fungal Infections. Curr. Protein Pept. Sci. 2006, 7, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Prusty, J.S.; Kumar, A.; Kumar, A. Anti-Fungal Peptides: An Emerging Category with Enthralling Therapeutic Prospects in the Treatment of Candidiasis. Crit. Rev. Microbiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Zida, A.; Bamba, S.; Yacouba, A.; Ouedraogo-Traore, R.; Guiguemdé, R.T. Anti-Candida Albicans Natural Products, Sources of New Antifungal Drugs: A Review. J. Mycol. Med. 2017, 27, 1–19. [Google Scholar] [CrossRef]
- Rodríguez-Castaño, G.P.; Rosenau, F.; Ständker, L.; Firacative, C. Antimicrobial Peptides: Avant-Garde Antifungal Agents to Fight against Medically Important Candida Species. Pharmaceutics 2023, 15, 789. [Google Scholar] [CrossRef]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A Review on Antimicrobial Peptides Databases and the Computational Tools. Database 2022, 2022, baac011. [Google Scholar] [CrossRef]
- Sahoo, A.; Swain, S.S.; Panda, S.K.; Hussain, T.; Panda, M.; Rodrigues, C.F. In Silico Identification of Potential Insect Peptides against Biofilm-Producing Staphylococcus Aureus. Chem. Biodivers. 2022, 19, e202200494. [Google Scholar] [CrossRef]
- Sama-ae, I.; Pattaranggoon, N.C.; Tedasen, A. In Silico Prediction of Antifungal Compounds from Natural Sources towards Lanosterol 14-Alpha Demethylase (CYP51) Using Molecular Docking and Molecular Dynamic Simulation. J. Mol. Graph. Model. 2023, 121, 108435. [Google Scholar] [CrossRef]
- Woodburn, K.W.; Edward Clemens, L.; Jaynes, J.; Joubert, L.M.; Botha, A.; Nazik, H.; Stevens, D.A. Designed Antimicrobial Peptides for Recurrent Vulvovaginal Candidiasis Treatment. Antimicrob. Agents Chemother. 2019, 63, e02690-18. [Google Scholar] [CrossRef]
- Datta, A.; Ghosh, A.; Airoldi, C.; Sperandeo, P.; Mroue, K.H.; Jimenez-Barbero, J.; Kundu, P.; Ramamoorthy, A.; Bhunia, A. Antimicrobial Peptides: Insights into Membrane Permeabilization, Lipopolysaccharide Fragmentation and Application in Plant Disease Control. Sci. Rep. 2015, 5, 11951. [Google Scholar] [CrossRef]
- Swain, S.S.; Singh, S.R.; Sahoo, A.; Panda, P.K.; Hussain, T.; Pati, S. Integrated Bioinformatics-Cheminformatics Approach toward Locating Pseudo-Potential Antiviral Marine Alkaloids against SARS-CoV-2-Mpro. Proteins 2022, 90, 1617–1633. [Google Scholar] [CrossRef] [PubMed]
- Zomorodbakhsh, S.; Abbasian, Y.; Naghinejad, M.; Sheikhpour, M. The Effects Study of Isoniazid Conjugated Multi-Wall Carbon Nanotubes Nanofluid on Mycobacterium tuberculosis. Int. J. Nanomed. 2020, 15, 5901–5909. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic Peptides: Current Applications and Future Directions. Signal Transduct. Target Ther. 2022, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.F.; Poh, C.L. Strategies to Improve the Physicochemical Properties of Peptide-Based Drugs. Pharm. Res. 2023, 40, 617–632. [Google Scholar] [CrossRef]
- Fuchs, J.E.; Wellenzohn, B.; Weskamp, N.; Liedl, K.R. Matched Peptides: Tuning Matched Molecular Pair Analysis for Biopharmaceutical Applications. J. Chem. Inf. Model. 2015, 55, 2315–2323. [Google Scholar] [CrossRef]
- Saleem, A.; Rajput, S. Insights from the in Silico Structural, Functional and Phylogenetic Characterization of Canine Lysyl Oxidase Protein. J. Genet. Eng. Biotechnol. 2020, 18, 20. [Google Scholar] [CrossRef]
- Cao, W.; Wu, L.Y.; Xia, X.Y.; Chen, X.; Wang, Z.X.; Pan, X.M. A Sequence-Based Evolutionary Distance Method for Phylogenetic Analysis of Highly Divergent Proteins. Sci. Rep. 2023, 13, 20304. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, A.; Roudbary, M.; Mohammadi, R.; Černáková, L.; Rodrigues, C.F. Overview on the Infections Related to Rare Candida Species. Pathogens 2022, 11, 963. [Google Scholar] [CrossRef]
- Rocha, W.R.V.d.; Nunes, L.E.; Neves, M.L.R.; Ximenes, E.C.P.d.A.; Albuquerque, M.C.P.d.A. Candida Genus—Virulence Factores, Epidemiology, Candidiasis and Resistance Mechanisms. Res. Soc. Dev. 2021, 10, e43910414283. [Google Scholar] [CrossRef]
- Zhang, L.M.; Zhou, S.W.; Huang, X.S.; Chen, Y.F.; Mwangi, J.; Fang, Y.Q.; Du, T.; Zhao, M.; Shi, L.; Lu, Q.M. Blap-6, a Novel Antifungal Peptide from the Chinese Medicinal Beetle Blaps Rhynchopetera against Cryptococcus Neoformans. Int. J. Mol. Sci. 2024, 25, 5336. [Google Scholar] [CrossRef]
- WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action. Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 19 May 2025).
- Odchimar, N.M.O.; Dulay, A.N.G.; Orosco, F.L. Molecular Modelling and Optimization of a High-Affinity Nanobody Targeting the Nipah Virus Fusion Protein through in Silico Site-Directed Mutagenesis. Comput. Biol. Chem. 2025, 115, 108354. [Google Scholar] [CrossRef]
- Sahoo, A.; Swain, S.S.; Paital, B.; Panda, M. Combinatorial Approach of Vitamin C Derivative and Anti-HIV Drug-Darunavir against SARS-CoV-2. Front. Biosci. (Landmark Ed.) 2022, 27, 10. [Google Scholar] [CrossRef]
- Chen, M.; Huang, W.K.; Yao, Y.; Wu, S.M.; Yang, Y.X.; Liu, W.X.; Luo, G.; Wei, S.F.; Zhang, H.; Liu, H.M.; et al. Heterologous Expression of the Insect SVWC Peptide WHIS1 Inhibits Candida Albicans Invasion into A549 and HeLa Epithelial Cells. Front. Microbiol. 2024, 15, 1358752. [Google Scholar] [CrossRef]
- Schaefer, S.; Vij, R.; Sprague, J.L.; Austermeier, S.; Dinh, H.; Judzewitsch, P.R.; Müller-Loennies, S.; Lopes Silva, T.; Seemann, E.; Qualmann, B.; et al. A Synthetic Peptide Mimic Kills Candida Albicans and Synergistically Prevents Infection. Nat. Commun. 2024, 15, 6818. [Google Scholar] [CrossRef]
- Tancer, R.J.; Pawar, S.; Wang, Y.; Ventura, C.R.; Wiedman, G.; Xue, C. Improved Broad Spectrum Antifungal Drug Synergies with Cryptomycin, a Cdc50-Inspired Antifungal Peptide. ACS Infect. Dis. 2024, 10, 3973–3993. [Google Scholar] [CrossRef]
- Spitzer, M.; Robbins, N.; Wright, G.D. Combinatorial Strategies for Combating Invasive Fungal Infections. Virulence 2017, 8, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Revie, N.M.; Iyer, K.R.; Maxson, M.E.; Zhang, J.; Yan, S.; Fernandes, C.M.; Meyer, K.J.; Chen, X.; Skulska, I.; Fogal, M.; et al. Targeting Fungal Membrane Homeostasis with Imidazopyrazoindoles Impairs Azole Resistance and Biofilm Formation. Nat. Commun. 2022, 13, 3634. [Google Scholar] [CrossRef] [PubMed]
- dos Reis, T.F.; de Castro, P.A.; Bastos, R.W.; Pinzan, C.F.; Souza, P.F.N.; Ackloo, S.; Hossain, M.A.; Drewry, D.H.; Alkhazraji, S.; Ibrahim, A.S.; et al. A Host Defense Peptide Mimetic, Brilacidin, Potentiates Caspofungin Antifungal Activity against Human Pathogenic Fungi. Nat. Commun. 2023, 14, 2052. [Google Scholar] [CrossRef]
- de Ullivarri, M.F.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell. Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef]
- Ratcliffe, N.A.; Mello, C.B.; Garcia, E.S.; Butt, T.M.; Azambuja, P. Insect Natural Products and Processes: New Treatments for Human Disease. Insect Biochem. Mol. Biol. 2011, 41, 747–769. [Google Scholar] [CrossRef]
- Rossi, D.C.; Muñoz, J.E.; Carvalho, D.D.; Belmonte, R.; Faintuch, B.; Borelli, P.; Miranda, A.; Taborda, C.P.; Daffre, S. Therapeutic Use of a Cationic Antimicrobial Peptide from the Spider Acanthoscurria Gomesiana in the Control of Experimental Candidiasis. BMC Microbiol. 2012, 12, 28. [Google Scholar] [CrossRef]
- Guevara-Lora, I.; Bras, G.; Juszczak, M.; Karkowska-Kuleta, J.; Gorecki, A.; Manrique-Moreno, M.; Dymek, J.; Pyza, E.; Kozik, A.; Rapala-Kozik, M. Cecropin D-Derived Synthetic Peptides in the Fight against Candida Albicans Cell Filamentation and Biofilm Formation. Front. Microbiol. 2023, 13, 1045984. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, E.; de Alteriis, E.; De Natale, A.; D’Alterio, A.; Siciliano, A.; Guida, M.; Lombardi, L.; Falanga, A.; Galdiero, S. Eradication of Candida Albicans Persister Cell Biofilm by the Membranotropic Peptide GH625. Sci. Rep. 2020, 10, 5780. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Contursi, P.; Vollaro, A.; Fusco, S.; Notomista, E.; Catania, M.R. Antifungal and Anti-Biofilm Activity of the First Cryptic Antimicrobial Peptide from an Archaeal Protein against Candida Spp. Clinical Isolates. Sci. Rep. 2018, 8, 17570. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.M.C.V.; Lopes, B.O.; Leite, A.C.R.d.M.; Cruz, G.S.; Brito, É.H.S.d.; Lima, L.F.d.; Černáková, L.; Azevedo, N.F.; Rodrigues, C.F. Characterization of Oral Candida Spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America. Antibiotics 2023, 12, 797. [Google Scholar] [CrossRef]
- do Nascimento Dias, J.; de Souza Silva, C.; de Araújo, A.R.; Souza, J.M.T.; de Holanda Veloso Júnior, P.H.; Cabral, W.F.; da Glória da Silva, M.; Eaton, P.; de Souza de Almeida Leite, J.R.; Nicola, A.M.; et al. Mechanisms of Action of Antimicrobial Peptides ToAP2 and NDBP-5.7 against Candida Albicans Planktonic and Biofilm Cells. Sci. Rep. 2020, 10, 10327. [Google Scholar] [CrossRef]
- Lamiable, A.; Thevenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tuffery, P. PEP-FOLD3: Faster de Novo Structure Prediction for Linear Peptides in Solution and in Complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A Web Server for Blind Peptide-Protein Docking Based on a Hierarchical Algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef]
- Yan, Y.; Tao, H.; He, J.; Huang, S.Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Swain, S.S.; Sahoo, A.; Singh, S.R.; Sahoo, J.; Paidesetty, S.K. Synthesis, Spectroscopic Analysis, and Computational-Based Investigations on “azo-Coumarin-Co(II)-Galangin” Hybrids Exhibit Multipotential Activities. J. Biomol. Struct. Dyn. 2024. [Google Scholar] [CrossRef]
- Sahoo, A.; Paidesetty, S.K.; Panda, M. Target-Specific High-Throughput Screening of Anti-Inflammatory Phytosteroids for Autoimmune Diseases: A Molecular Docking-Dynamics Simulation Approach. Steroids 2025, 217, 109601. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Zaman, W.; Ye, J.; Saqib, S.; Liu, Y.; Shan, Z.; Hao, D.; Chen, Z.; Xiao, P. Predicting Potential Medicinal Plants with Phylogenetic Topology: Inspiration from the Research of Traditional Chinese Medicine. J. Ethnopharmacol. 2021, 281, 114515. [Google Scholar] [CrossRef] [PubMed]
- Chase, K.; Watkins, M.; Safavi-Hemami, H.; Olivera, B.M. Integrating Venom Peptide Libraries Into a Phylogenetic and Broader Biological Framework. Front. Mol. Biosci. 2022, 9, 784419. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. Definitive Document, E.Def 7.4. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts; EUCAST: Växjö, Sweden, 2003. [Google Scholar]
- Maziere, M.; Rompante, P.; Andrade, J.C.; De Oliveira, B.S.F.; Alves, M.C.; Rodrigues, C.F. Repurposing Mouthwashes: Antifungal and Antibiofilm Abilities of Commercially Available Mouthwashes Against Candida spp. Antibiotics 2025, 14, 483. [Google Scholar] [CrossRef] [PubMed]
- Rajão, A.; Silva, J.P.N.; Almeida-Nunes, D.L.; Rompante, P.; Rodrigues, C.F.; Andrade, J.C. Limosilactobacillus Reuteri AJCR4: A Potential Probiotic in the Fight Against Oral Candida Spp. Biofilms. Int. J. Mol. Sci. 2025, 26, 638. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Boas, D.V.; Haynes, K.; Henriques, M. The MNN2 Gene Knockout Modulates the Antifungal Resistance of Biofilms of Candida Glabrata. Biomolecules 2018, 8, 130. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log Cells/mL ± SD | |||
|---|---|---|---|
| Control | Blap-6 (200 mg/L) | Gomesin (265 mg/L) | |
| C. albicans SC5314 | 11.30 ± 0.25 | 9.67 ± 0.12 (14.4% reduction) | 0.00 ± 0.00 (100% reduction) Complete eradication |
| C. tropicalis ATCC750 | 11.10 ± 0.04 | 10.10 ± 0.07 (9.0% reduction) | 10.10 ± 0.01 (9.0% reduction) |
| C. glabrata ATCC2001 | 10.90 ± 0.02 | 10.10 ± 0.02 (7.3% reduction) | 0.00 ± 0.00 (100% reduction) Complete eradication |
| C. parapsilosis ATCC20019 | 11.00 ± 0.26 | 9.43 ± 0.02 (14.3% reduction) | 0.00 ± 0.00 (100% reduction) Complete eradication |
| % Biomass | ||
|---|---|---|
| Blap-6 (200 mg/L) | Gomesin (265 mg/L) | |
| C. albicans SC5314 | +47% | −49% |
| C. tropicalis ATCC750 | +21% | −35% |
| C. glabrata ATCC2001 | +16% | −19% |
| C. parapsilosis ATCC20019 | +46% | −16% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, C.; Sahoo, A.; Swain, S.S.; Gupta, P.; Silva, F.; Azevedo, A.S.; Rodrigues, C.F. In Silico and In Vitro Potential Antifungal Insights of Insect-Derived Peptides in the Management of Candida sp. Infections. Int. J. Mol. Sci. 2025, 26, 7449. https://doi.org/10.3390/ijms26157449
Sousa C, Sahoo A, Swain SS, Gupta P, Silva F, Azevedo AS, Rodrigues CF. In Silico and In Vitro Potential Antifungal Insights of Insect-Derived Peptides in the Management of Candida sp. Infections. International Journal of Molecular Sciences. 2025; 26(15):7449. https://doi.org/10.3390/ijms26157449
Chicago/Turabian StyleSousa, Catarina, Alaka Sahoo, Shasank Sekhar Swain, Payal Gupta, Francisco Silva, Andreia S. Azevedo, and Célia Fortuna Rodrigues. 2025. "In Silico and In Vitro Potential Antifungal Insights of Insect-Derived Peptides in the Management of Candida sp. Infections" International Journal of Molecular Sciences 26, no. 15: 7449. https://doi.org/10.3390/ijms26157449
APA StyleSousa, C., Sahoo, A., Swain, S. S., Gupta, P., Silva, F., Azevedo, A. S., & Rodrigues, C. F. (2025). In Silico and In Vitro Potential Antifungal Insights of Insect-Derived Peptides in the Management of Candida sp. Infections. International Journal of Molecular Sciences, 26(15), 7449. https://doi.org/10.3390/ijms26157449