

Afucosylated IgG Promote Thrombosis in Mouse Injected with SARS-CoV-2 Spike Expressing Megakaryocytes

 , , , , ,
, , , , ,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Proplatelets Derived from SARS-CoV-2-Infected Human MKs Carry Spike Protein on Their Membrane During Their Maturation
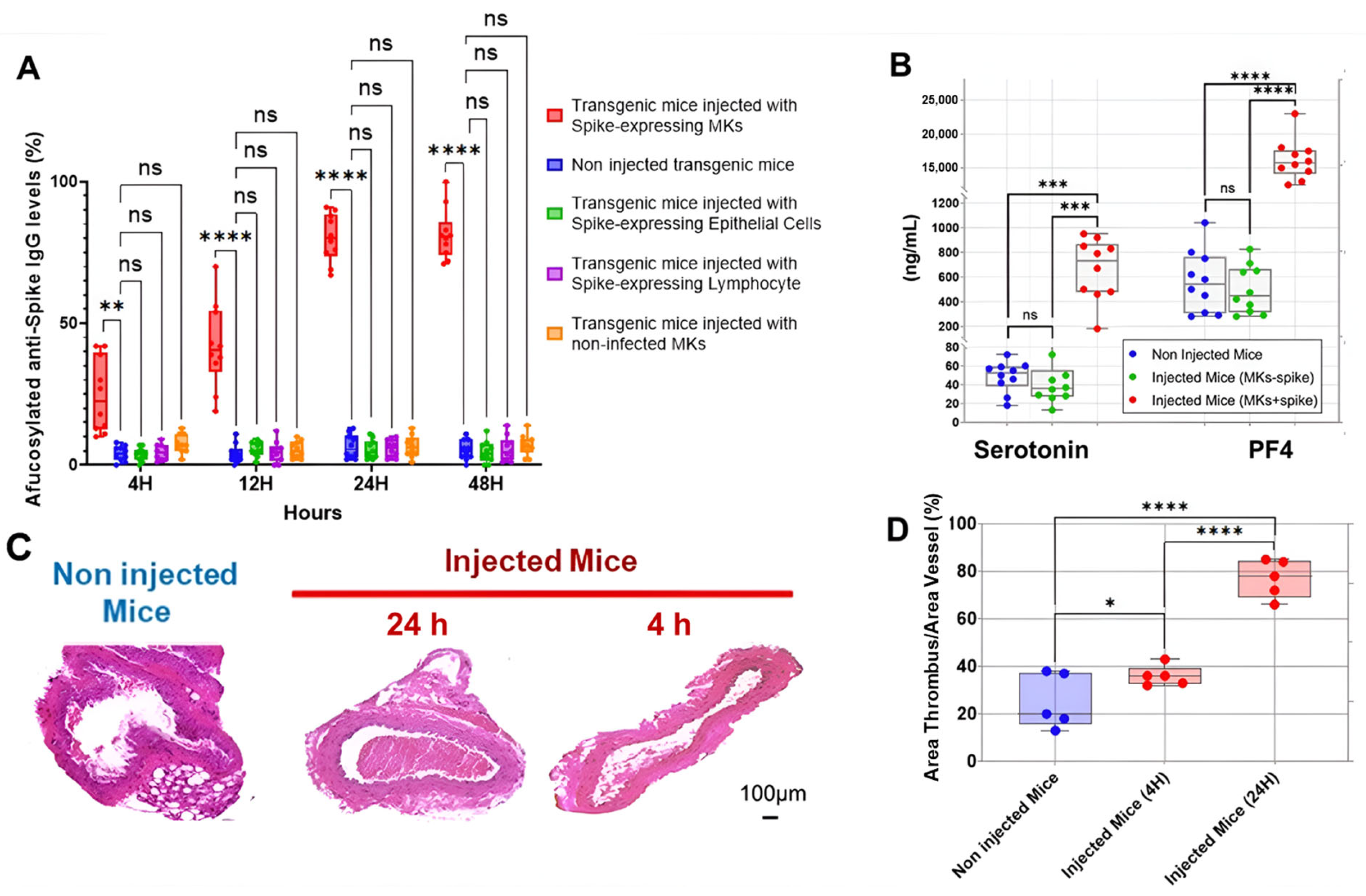
2.2. SARS-CoV-2-Infected Human MK Injection Triggers the Formation of Afucosylated Anti-Spike IgGs and Promotes Platelet Activation, Lung Injury, and Death in FcγRIIa-Transgenic Mice
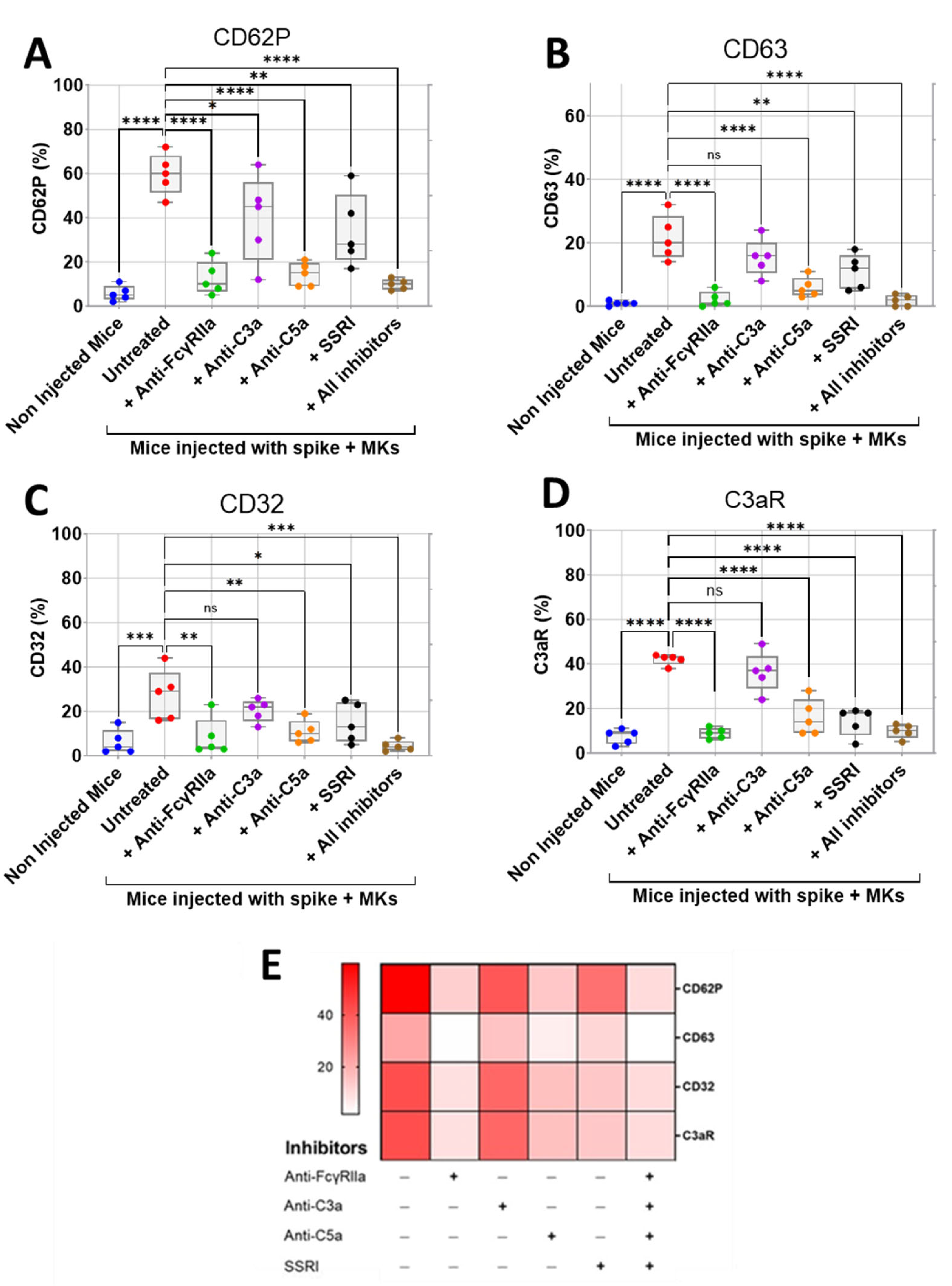
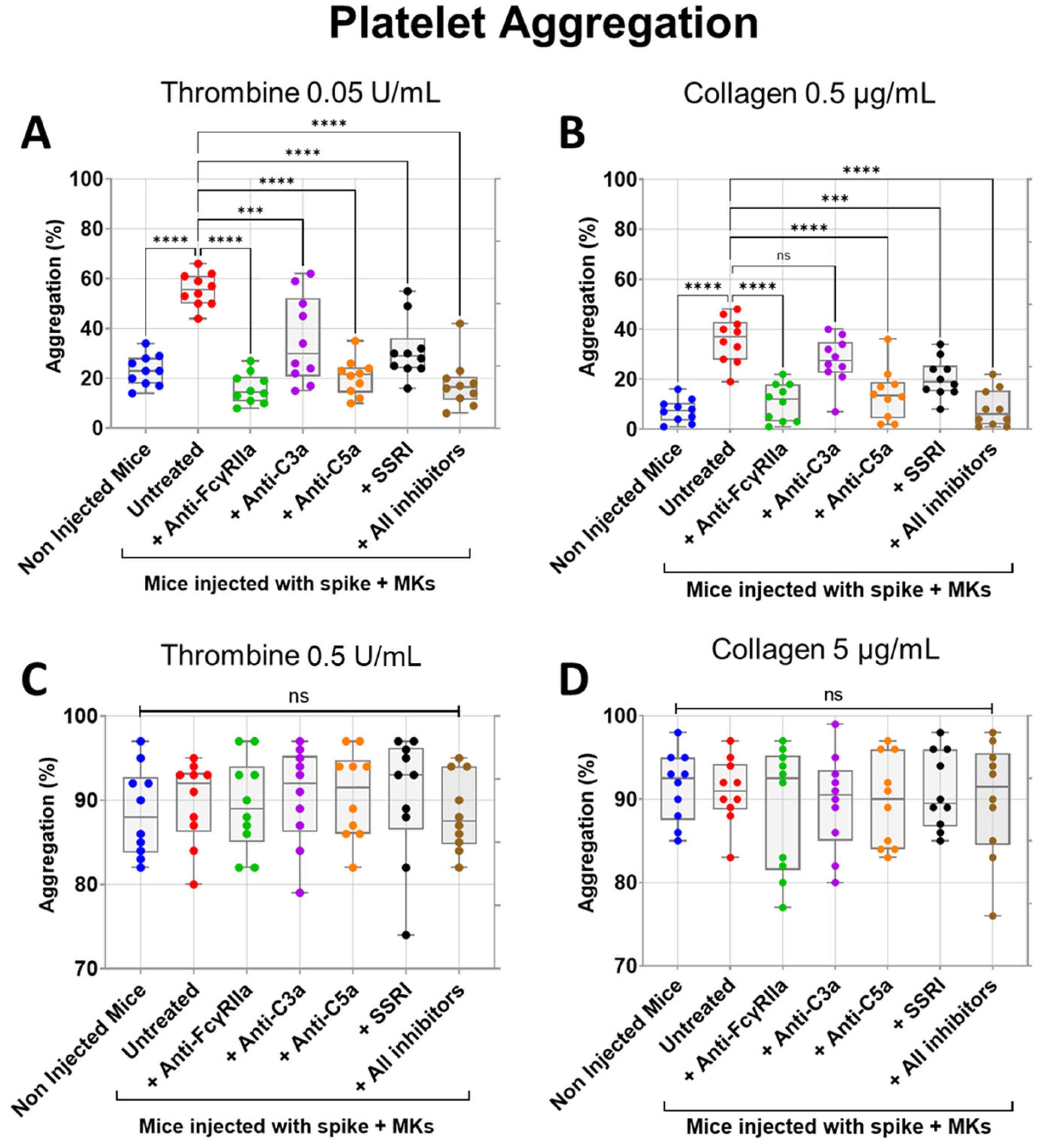
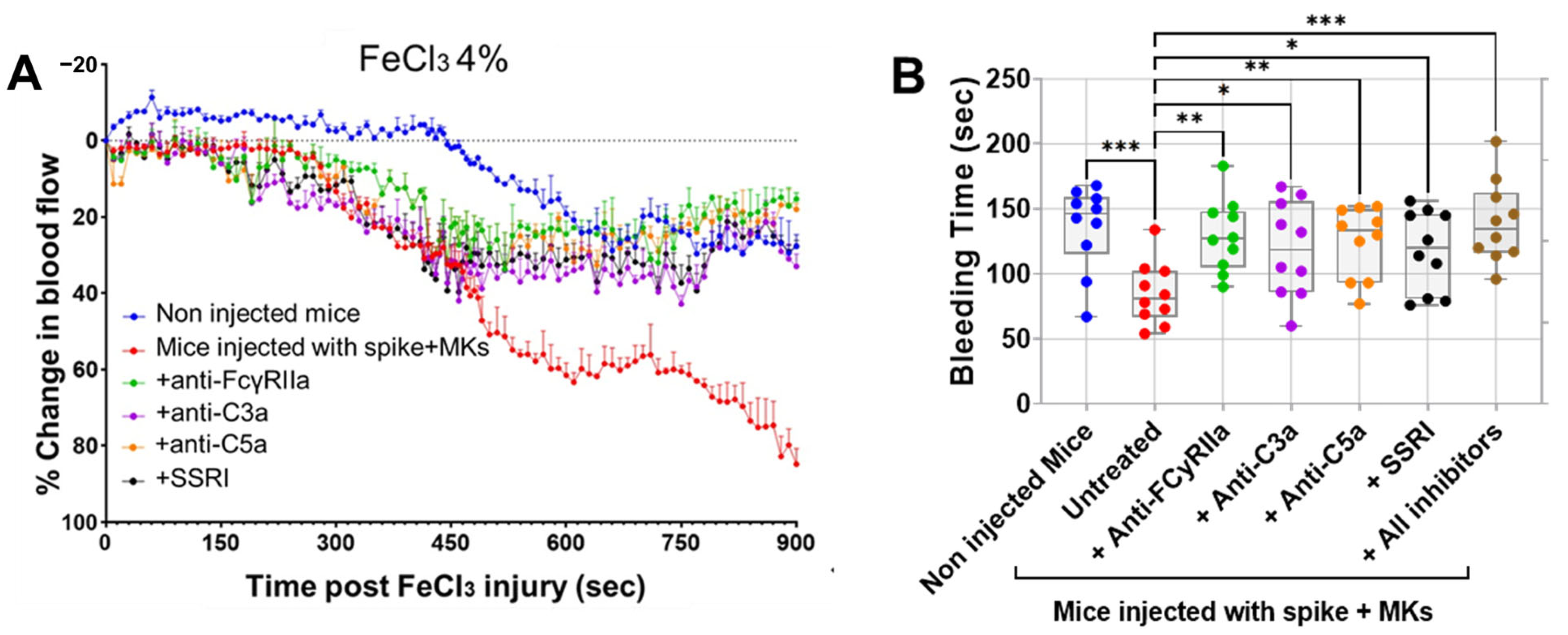
2.3. Inhibition of FcγRIIa Receptor, Serotonin Transporter, or Complement Anaphylatoxins Prevents Thrombosis by Reducing Platelet Activation and Aggregation in Injected FcγRIIa-Transgenic Mice with High Level of Afucosylated Anti-Spike IgGs
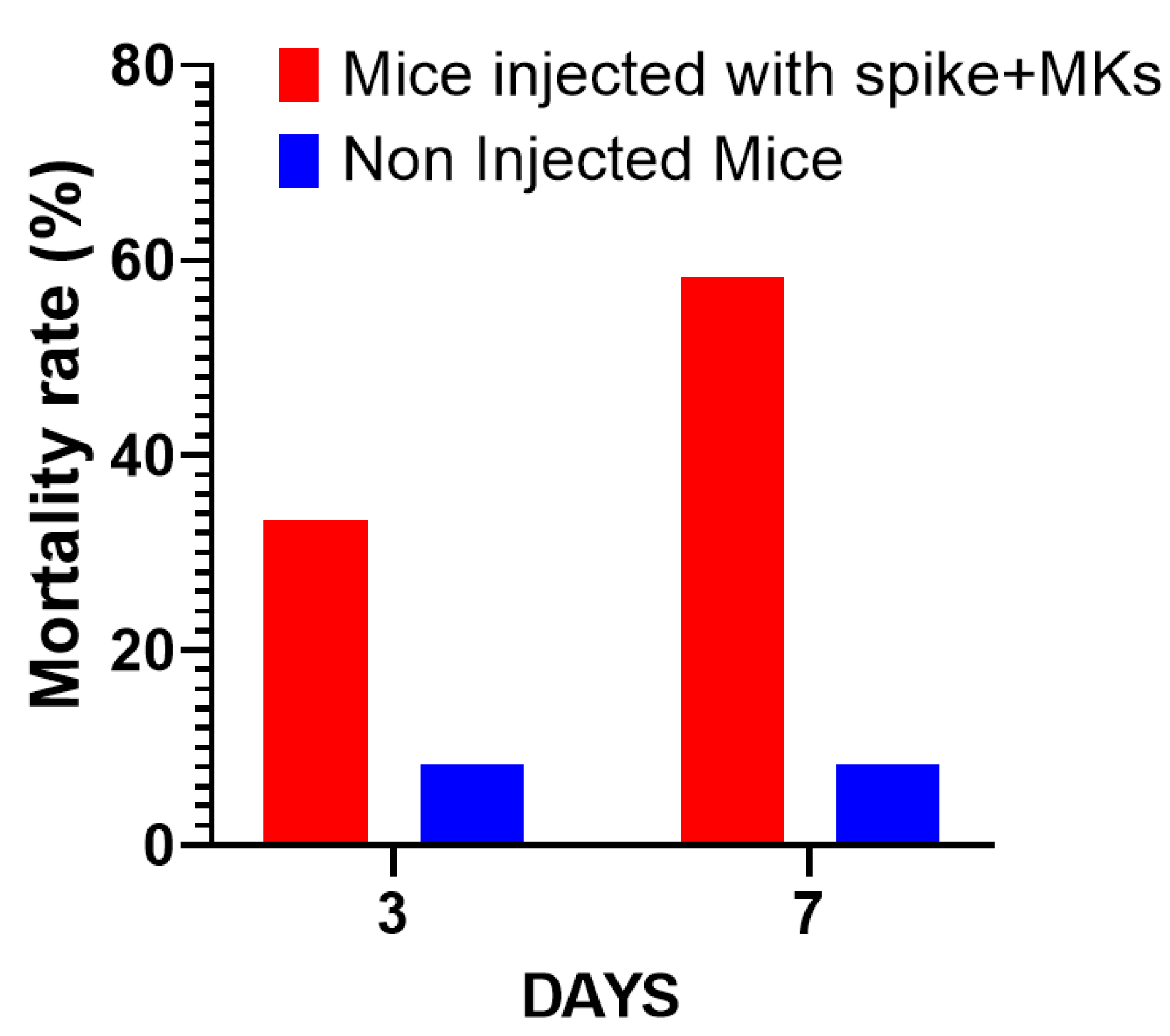
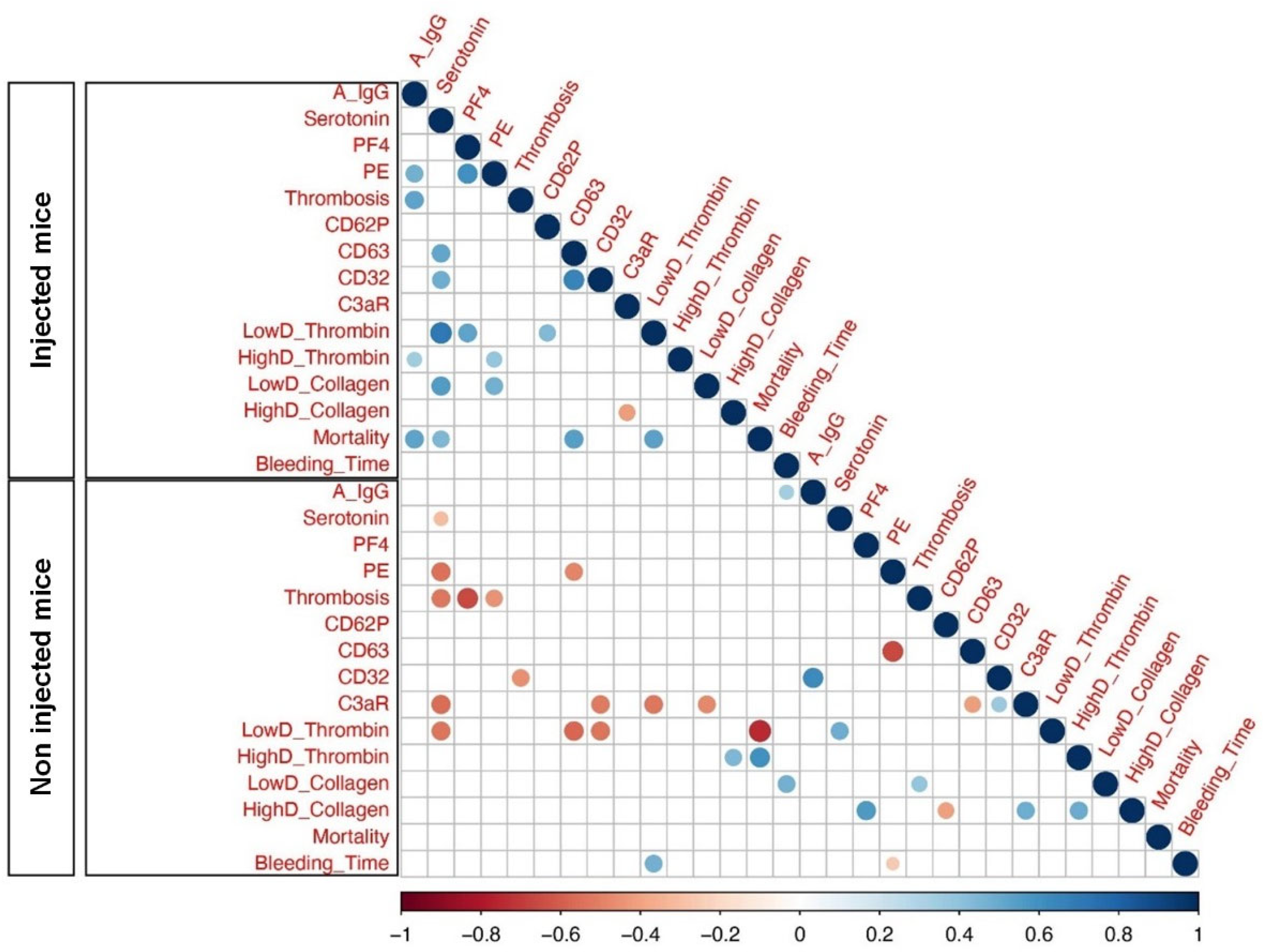
2.4. Afucosylated Anti-Spike IgG Levels Correlate with Pulmonary Thromboembolism, and Mortality in FcγRIIa-Transgenic Mice with Spike-Expressing Human MKs
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Human MK Isolation
4.3. Confocal Microscopy
4.4. Preparation of Cells for Injection
4.5. Preparation of Murine Platelets
4.6. Flow Cytometry Analysis
4.7. Platelet Aggregation Assay
4.8. Purification of Total IgG Antibodies
4.9. Mass Spectrometry for Afucosylated Anti-Spike IgG Quantification
4.10. PF4 and Serotonin Quantification in Plasma
4.11. Tail-Bleeding Time
4.12. Pulmonary Thrombo-Embolism
4.13. FeCl3-Induced Arterial Thrombosis
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouadma, L.; Lescure, F.-X.; Lucet, J.-C.; Yazdanpanah, Y.; Timsit, J.-F. Severe SARS-CoV-2 infections: Practical considerations and management strategy for intensivists. Intensive Care Med. 2020, 46, 579–582. [Google Scholar] [CrossRef]
- Fortmann, S.D.; Patton, M.J.; Frey, B.F.; Tipper, J.L.; Reddy, S.B.; Vieira, C.P.; Hanumanthu, V.S.; Sterrett, S.; Floyd, J.L.; Prasad, R. Circulating SARS-CoV-2+ megakaryocytes are associated with severe viral infection in COVID-19. Blood Adv. 2023, 7, 4200–4214. [Google Scholar] [CrossRef]
- Zhu, A.; Real, F.; Capron, C.; Rosenberg, A.R.; Silvin, A.; Dunsmore, G.; Zhu, J.; Cottoignies-Callamarte, A.; Massé, J.-M.; Moine, P. Infection of lung megakaryocytes and platelets by SARS-CoV-2 anticipate fatal COVID-19. Cell. Mol. Life Sci. 2022, 79, 365. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.D.; de Graaf, E.L.; Sonneveld, M.E.; Plomp, H.R.; Nouta, J.; Hoepel, W.; Chen, H.-J.; Linty, F.; Visser, R.; Brinkhaus, M. Afucosylated IgG characterizes enveloped viral responses and correlates with COVID-19 severity. Science 2021, 371, eabc8378. [Google Scholar] [CrossRef] [PubMed]
- Hoepel, W.; Chen, H.-J.; Geyer, C.E.; Allahverdiyeva, S.; Manz, X.D.; de Taeye, S.W.; Aman, J.; Mes, L.; Steenhuis, M.; Griffith, G.R. High titers and low fucosylation of early human anti–SARS-CoV-2 IgG promote inflammation by alveolar macrophages. Sci. Transl. Med. 2021, 13, eabf8654. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gonzalez, J.C.; Sievers, B.L.; Mallajosyula, V.; Chakraborty, S.; Dubey, M.; Ashraf, U.; Cheng, B.Y.-L.; Kathale, N.; Tran, K.Q.T. Early non-neutralizing, afucosylated antibody responses are associated with COVID-19 severity. Sci. Transl. Med. 2022, 14, eabm7853. [Google Scholar] [CrossRef]
- Oosterhoff, J.J.; Larsen, M.D.; van der Schoot, C.E.; Vidarsson, G. Afucosylated IgG responses in humans–structural clues to the regulation of humoral immunity. Trends Immunol. 2022, 43, 800–814. [Google Scholar] [CrossRef]
- Kapur, R.; Kustiawan, I.; Vestrheim, A.; Koeleman, C.A.; Visser, R.; Einarsdottir, H.K.; Porcelijn, L.; Jackson, D.; Kumpel, B.; Deelder, A.M. A prominent lack of IgG1-Fc fucosylation of platelet alloantibodies in pregnancy. Blood J. Am. Soc. Hematol. 2014, 123, 471–480. [Google Scholar] [CrossRef]
- Sonneveld, M.E.; Natunen, S.; Sainio, S.; Koeleman, C.A.; Holst, S.; Dekkers, G.; Koelewijn, J.; Partanen, J.; van der Schoot, C.E.; Wuhrer, M. Glycosylation pattern of anti-platelet IgG is stable during pregnancy and predicts clinical outcome in alloimmune thrombocytopenia. Br. J. Haematol. 2016, 174, 310–320. [Google Scholar] [CrossRef]
- Qiao, J.; Al-Tamimi, M.; Baker, R.I.; Andrews, R.K.; Gardiner, E.E. The platelet Fc receptor, FcγRIIa. Immunol. Rev. 2015, 268, 241–252. [Google Scholar] [CrossRef]
- Arman, M.; Krauel, K. Human platelet IgG Fc receptor FcγRIIA in immunity and thrombosis. J. Thromb. Haemost. 2015, 13, 893–908. [Google Scholar] [CrossRef]
- Patel, P.; Michael, J.V.; Naik, U.P.; McKenzie, S.E. Platelet FcγRIIA in immunity and thrombosis: Adaptive immunothrombosis. J. Thromb. Haemost. 2021, 19, 1149–1160. [Google Scholar] [CrossRef]
- Bye, A.P.; Hoepel, W.; Mitchell, J.L.; Jégouic, S.; Loureiro, S.; Sage, T.; Vidarsson, G.; Nouta, J.; Wuhrer, M.; de Taeye, S. Aberrant glycosylation of anti-SARS-CoV-2 spike IgG is a prothrombotic stimulus for platelets. Blood J. Am. Soc. Hematol. 2021, 138, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat. Immunol. 2021, 22, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Sun, S.-H.; Chen, Q.; Gu, H.-J.; Yang, G.; Wang, Y.-X.; Huang, X.-Y.; Liu, S.-S.; Zhang, N.-N.; Li, X.-F.; Xiong, R. A mouse model of SARS-CoV-2 infection and pathogenesis. Cell Host Microbe 2020, 28, 124–133.e124. [Google Scholar] [CrossRef]
- Knight, A.C.; Montgomery, S.A.; Fletcher, C.A.; Baxter, V.K. Mouse models for the study of SARS-CoV-2 infection. Comp. Med. 2021, 71, 383–397. [Google Scholar] [CrossRef]
- Mercado, C.P.; Kilic, F. Molecular mechanisms of SERT in platelets: Regulation of plasma serotonin levels. Mol. Interv. 2010, 10, 231. [Google Scholar] [CrossRef]
- Bruhns, P.; Jönsson, F. Mouse and human FcR effector functions. Immunol. Rev. 2015, 268, 25–51. [Google Scholar] [CrossRef]
- McKenzie, S.E.; Taylor, S.M.; Malladi, P.; Yuhan, H.; Cassel, D.L.; Chien, P.; Schwartz, E.; Schreiber, A.D.; Surrey, S.; Reilly, M.P. The role of the human Fc receptor FcγRIIA in the immune clearance of platelets: A transgenic mouse model. J. Immunol. 1999, 162, 4311–4318. [Google Scholar] [CrossRef]
- Chauhan, A.J.; Wiffen, L.J.; Brown, T.P. COVID-19: A collision of complement, coagulation and inflammatory pathways. J. Thromb. Haemost. 2020, 18, 2110–2117. [Google Scholar] [CrossRef]
- Apostolidis, S.A.; Sarkar, A.; Giannini, H.M.; Goel, R.R.; Mathew, D.; Suzuki, A.; Baxter, A.E.; Greenplate, A.R.; Alanio, C.; Abdel-Hakeem, M. Signaling through FcγRIIA and the C5a-C5aR pathway mediate platelet hyperactivation in COVID-19. Front. Immunol. 2022, 13, 834988. [Google Scholar] [CrossRef] [PubMed]
- Alosaimi, B.; Mubarak, A.; Hamed, M.E.; Almutairi, A.Z.; Alrashed, A.A.; AlJuryyan, A.; Enani, M.; Alenzi, F.Q.; Alturaiki, W. Complement anaphylatoxins and inflammatory cytokines as prognostic markers for COVID-19 severity and in-hospital mortality. Front. Immunol. 2021, 12, 668725. [Google Scholar] [CrossRef] [PubMed]
- Sonneveld, M.E.; Koelewijn, J.; de Haas, M.; Admiraal, J.; Plomp, R.; Koeleman, C.A.; Hipgrave Ederveen, A.L.; Ligthart, P.; Wuhrer, M.; van der Schoot, C.E. Antigen specificity determines anti-red blood cell IgG-Fc alloantibody glycosylation and thereby severity of haemolytic disease of the fetus and newborn. Br. J. Haematol. 2017, 176, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.D.; Lopez-Perez, M.; Dickson, E.K.; Ampomah, P.; Tuikue Ndam, N.; Nouta, J.; Koeleman, C.A.; Ederveen, A.L.H.; Mordmüller, B.; Salanti, A. Afucosylated Plasmodium falciparum-specific IgG is induced by infection but not by subunit vaccination. Nat. Commun. 2021, 12, 5838. [Google Scholar] [CrossRef]
- Wang, T.T.; Sewatanon, J.; Memoli, M.J.; Wrammert, J.; Bournazos, S.; Bhaumik, S.K.; Pinsky, B.A.; Chokephaibulkit, K.; Onlamoon, N.; Pattanapanyasat, K. IgG antibodies to dengue enhanced for FcγRIIIA binding determine disease severity. Science 2017, 355, 395–398. [Google Scholar] [CrossRef]
- Xiao, F.; Rui, K.; Shi, X.; Wu, H.; Cai, X.; Lui, K.O.; Lu, Q.; Ballestar, E.; Tian, J.; Zou, H. Epigenetic regulation of B cells and its role in autoimmune pathogenesis. Cell. Mol. Immunol. 2022, 19, 1215–1234. [Google Scholar] [CrossRef]
- Cao, Y.; Song, Z.; Guo, Z.; Zhao, X.; Gong, Y.; Zhao, K.; Qu, C.; Huang, Y.; Li, Y.; Gao, Y. Cytokines in the immune microenvironment change the glycosylation of IgG by regulating intracellular glycosyltransferases. Front. Immunol. 2022, 12, 724379. [Google Scholar] [CrossRef]
- Ozturkler, Z.; Kalkan, R. A new perspective of COVID-19 infection: An epigenetics point of view. Glob. Med. Genet. 2022, 9, 004–006. [Google Scholar] [CrossRef]
- Guilliams, M.; Bruhns, P.; Saeys, Y.; Hammad, H.; Lambrecht, B.N. The function of Fcγ receptors in dendritic cells and macrophages. Nat. Rev. Immunol. 2014, 14, 94–108. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 2020, 181, 1036–1045, e1039. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, Q.; Wang, D.C.; Ingbar, D.H.; Wang, X. Acute lung injury in patients with COVID-19 infection. Clin. Transl. Med. 2020, 10, 20–27. [Google Scholar] [CrossRef]
- Becker, R.C. COVID-19 update: COVID-19-associated coagulopathy. J. Thromb. Thrombolysis 2020, 50, 54–67. [Google Scholar] [CrossRef]
- Zaid, Y.; Dore, E.; Dubuc, I.; Archambault, A.-S.; Flamand, O.; Laviolette, M.; Flamand, N.; Boilard, E.; Flamand, L. Chemokines and eicosanoids fuel the hyperinflammation within the lungs of patients with severe COVID-19. J. Allergy Clin. Immunol. 2021, 148, 368–380, e363. [Google Scholar] [CrossRef]
- Zaid, Y.; Puhm, F.; Allaeys, I.; Naya, A.; Oudghiri, M.; Khalki, L.; Limami, Y.; Zaid, N.; Sadki, K.; Ben El Haj, R. Platelets can associate with SARS-CoV-2 RNA and are hyperactivated in COVID-19. Circ. Res. 2020, 127, 1404–1418. [Google Scholar] [CrossRef]
- de Taeye, S.W.; Schriek, A.I.; Umotoy, J.C.; Grobben, M.; Burger, J.A.; Sanders, R.W.; Vidarsson, G.; Wuhrer, M.; Falck, D.; Kootstra, N.A. Afucosylated broadly neutralizing antibodies enhance clearance of HIV-1 infected cells through cell-mediated killing. Commun. Biol. 2024, 7, 964. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Wang, X.; Yang, L.; Li, H.; Wang, Y.; Liu, M.; Zhao, X.; Xie, Y.; Yang, Y. SARS-CoV-2 binds platelet ACE2 to enhance thrombosis in COVID-19. J. Hematol. Oncol. 2020, 13, 120. [Google Scholar] [CrossRef]
- Agbani, E.O.; Schneider, P.; McDonald, B.; Skeith, L.; Poon, M.-C.; Lee, A. Activated platelets harbor SARS-CoV-2 during severe COVID-19. Thromb. Haemost. 2021, 122, 308–309. [Google Scholar] [CrossRef]
- Zaid, Y.; Guessous, F.; Puhm, F.; Elhamdani, W.; Chentoufi, L.; Morris, A.C.; Cheikh, A.; Jalali, F.; Boilard, E.; Flamand, L. Platelet reactivity to thrombin differs between patients with COVID-19 and those with ARDS unrelated to COVID-19. Blood Adv. 2021, 5, 635–639. [Google Scholar] [CrossRef]
- Fritz, B.A.; Hoertel, N.; Lenze, E.J.; Jalali, F.; Reiersen, A.M. Association between antidepressant use and ED or hospital visits in outpatients with SARS-CoV-2. Transl. Psychiatry 2022, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Oskotsky, T.; Marić, I.; Tang, A.; Oskotsky, B.; Wong, R.J.; Aghaeepour, N.; Sirota, M.; Stevenson, D.K. Mortality risk among patients with COVID-19 prescribed selective serotonin reuptake inhibitor antidepressants. JAMA Netw. Open 2021, 4, e2133090. [Google Scholar] [CrossRef]
- Jalali, F.; Rezaie, S.; Rola, P.; Kyle-Sidell, C. COVID-19 Pathophysiology: Are Platelets and Serotonin Hiding in Plain Sight? 2021. Available online: https://ssrn.com/abstract=3800402 (accessed on 20 December 2024).
- Stratz, C.; Trenk, D.; Bhatia, H.S.; Valina, C.; Neumann, F.-J.; Fiebich, B.L. Identification of 5-HT3 receptors on human platelets: Increased surface immunoreactivity after activation with adenosine diphosphate (ADP) and thrombin receptor-activating peptide (TRAP). Thromb. Haemost. 2008, 99, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Cervia-Hasler, C.; Brüningk, S.C.; Hoch, T.; Fan, B.; Muzio, G.; Thompson, R.C.; Ceglarek, L.; Meledin, R.; Westermann, P.; Emmenegger, M. Persistent complement dysregulation with signs of thromboinflammation in active Long Covid. Science 2024, 383, eadg7942. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Livada, A.C.; Morrell, C.N. Platelet and megakaryocyte roles in innate and adaptive immunity. Circ. Res. 2022, 130, 288–308. [Google Scholar] [CrossRef]
- Fletcher-Sandersjöö, A.; Bellander, B.-M. Is COVID-19 associated thrombosis caused by overactivation of the complement cascade? A literature review. Thromb. Res. 2020, 194, 36–41. [Google Scholar] [CrossRef]
- Sauter, R.J.; Sauter, M.; Reis, E.S.; Emschermann, F.N.; Nording, H.; Ebenhöch, S.; Kraft, P.; Münzer, P.; Mauler, M.; Rheinlaender, J. Functional relevance of the anaphylatoxin receptor C3aR for platelet function and arterial thrombus formation marks an intersection point between innate immunity and thrombosis. Circulation 2018, 138, 1720–1735. [Google Scholar] [CrossRef]
- Campbell, C.M.; Kahwash, R. Will complement inhibition be the new target in treating COVID-19–related systemic thrombosis? Circulation 2020, 141, 1739–1741. [Google Scholar] [CrossRef]
- Vitiello, A.; La Porta, R.; D’Aiuto, V.; Ferrara, F. Pharmacological approach for the reduction of inflammatory and prothrombotic hyperactive state in COVID-19 positive patients by acting on complement cascade. Hum. Immunol. 2021, 82, 264–269. [Google Scholar] [CrossRef]
- Afzali, B.; Noris, M.; Lambrecht, B.N.; Kemper, C. The state of complement in COVID-19. Nat. Rev. Immunol. 2022, 22, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Butov, K.R.; Osipova, E.Y.; Mikhalkin, N.B.; Trubina, N.M.; Panteleev, M.A.; Machlus, K.R. In vitro megakaryocyte culture from human bone marrow aspirates as a research and diagnostic tool. Platelets 2021, 32, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Di Lullo, A.M.; Scorza, M.; Amato, F.; Comegna, M.; Raia, V.; Maiuri, L.; Ilardi, G.; Cantone, E.; Castaldo, G.; Iengo, M. An “ex vivo model” contributing to the diagnosis and evaluation of new drugs in cystic fibrosis. Acta Otorhinolaryngol. Ital. 2017, 37, 207. [Google Scholar] [CrossRef]
- Yacoub, D.; Hachem, A.; Théorêt, J.-F.o.; Gillis, M.-A.; Mourad, W.; Merhi, Y. Enhanced levels of soluble CD40 ligand exacerbate platelet aggregation and thrombus formation through a CD40-dependent tumor necrosis factor receptor–associated factor-2/Rac1/p38 mitogen-activated protein kinase signaling pathway. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2424–2433. [Google Scholar] [CrossRef]
- Sodemann, E.B.; Dähling, S.; Klopfleisch, R.; Boiarina, E.; Cataldo, D.; Alhasan, M.M.; Yildirim, A.Ö.; Witzenrath, M.; Tabeling, C.; Conrad, M.L. Maternal asthma is associated with persistent changes in allergic offspring antibody glycosylation. Clin. Exp. Allergy 2020, 50, 520–531. [Google Scholar] [CrossRef]
- Barrientos, G.; Habazin, S.; Novokmet, M.; Almousa, Y.; Lauc, G.; Conrad, M.L. Changes in subclass-specific IgG Fc glycosylation associated with the postnatal maturation of the murine immune system. Sci. Rep. 2020, 10, 15243. [Google Scholar] [CrossRef]
- Zaytseva, O.O.; Jansen, B.C.; Hanić, M.; Mrčela, M.; Razdorov, G.; Stojković, R.; Erhardt, J.; Brizić, I.; Jonjić, S.; Pezer, M. MIgGGly (mouse IgG glycosylation analysis)—A high-throughput method for studying Fc-linked IgG N-glycosylation in mice with nanoUPLC-ESI-MS. Sci. Rep. 2018, 8, 13688. [Google Scholar] [CrossRef]
- Mustafa, M.H.; Mispireta, L.A.; Pierce, L.E. Occult pulmonary embolism presenting with thrombocytopenia and elevated fibrin split products. Am. J. Med. 1989, 86, 490–491. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mabrouk, M.; Atifi, F.; Wahnou, H.; Allaoui, A.; Zaid, N.; Naya, A.; Agbani, E.O.; Khalki, L.; Khyatti, M.; Tijani, Y.; et al. Afucosylated IgG Promote Thrombosis in Mouse Injected with SARS-CoV-2 Spike Expressing Megakaryocytes. Int. J. Mol. Sci. 2025, 26, 7002. https://doi.org/10.3390/ijms26147002
Mabrouk M, Atifi F, Wahnou H, Allaoui A, Zaid N, Naya A, Agbani EO, Khalki L, Khyatti M, Tijani Y, et al. Afucosylated IgG Promote Thrombosis in Mouse Injected with SARS-CoV-2 Spike Expressing Megakaryocytes. International Journal of Molecular Sciences. 2025; 26(14):7002. https://doi.org/10.3390/ijms26147002
Chicago/Turabian StyleMabrouk, Meryem, Farah Atifi, Hicham Wahnou, Afaf Allaoui, Nabil Zaid, Abdallah Naya, Ejaife O. Agbani, Loubna Khalki, Meriem Khyatti, Youssef Tijani, and et al. 2025. "Afucosylated IgG Promote Thrombosis in Mouse Injected with SARS-CoV-2 Spike Expressing Megakaryocytes" International Journal of Molecular Sciences 26, no. 14: 7002. https://doi.org/10.3390/ijms26147002
APA StyleMabrouk, M., Atifi, F., Wahnou, H., Allaoui, A., Zaid, N., Naya, A., Agbani, E. O., Khalki, L., Khyatti, M., Tijani, Y., Akarid, K., Arnoult, D., Abou-Saleh, H., El Faqer, O., Labied, S., Ammara, M., Guessous, F., Jalali, F., & Zaid, Y. (2025). Afucosylated IgG Promote Thrombosis in Mouse Injected with SARS-CoV-2 Spike Expressing Megakaryocytes. International Journal of Molecular Sciences, 26(14), 7002. https://doi.org/10.3390/ijms26147002