

Human Dialyzable Leukocyte Extract Enhances Albendazole Efficacy and Promotes Th1/Th2-Biased Lymphocyte and Antibody Responses in Peritoneal Cavity of Murine Model of Mesocestoides vogae Infection
 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
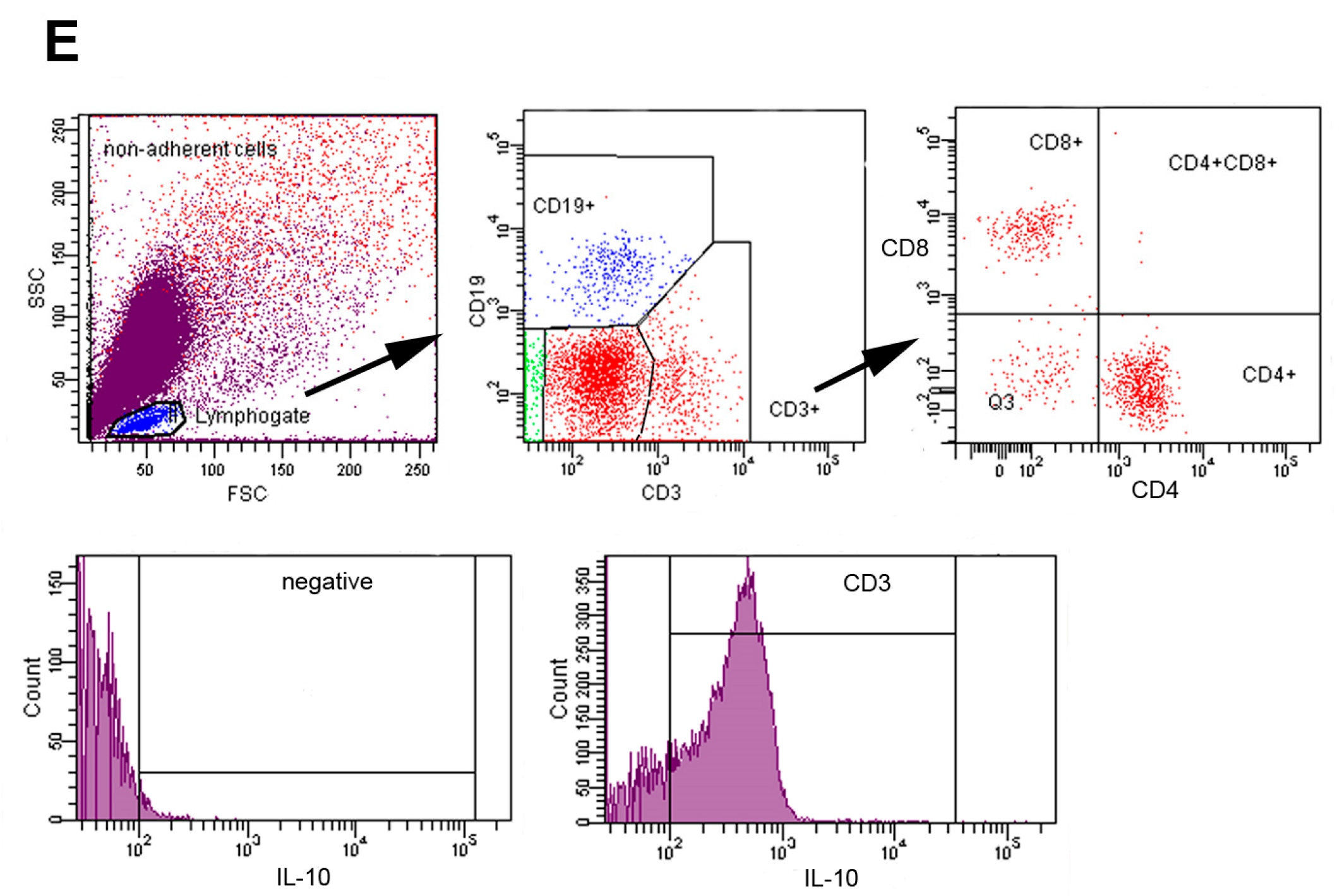
2.1. Phenotypic Analysis of Lymphoid Sub-Populations in the Peritoneal Cavities of Mice
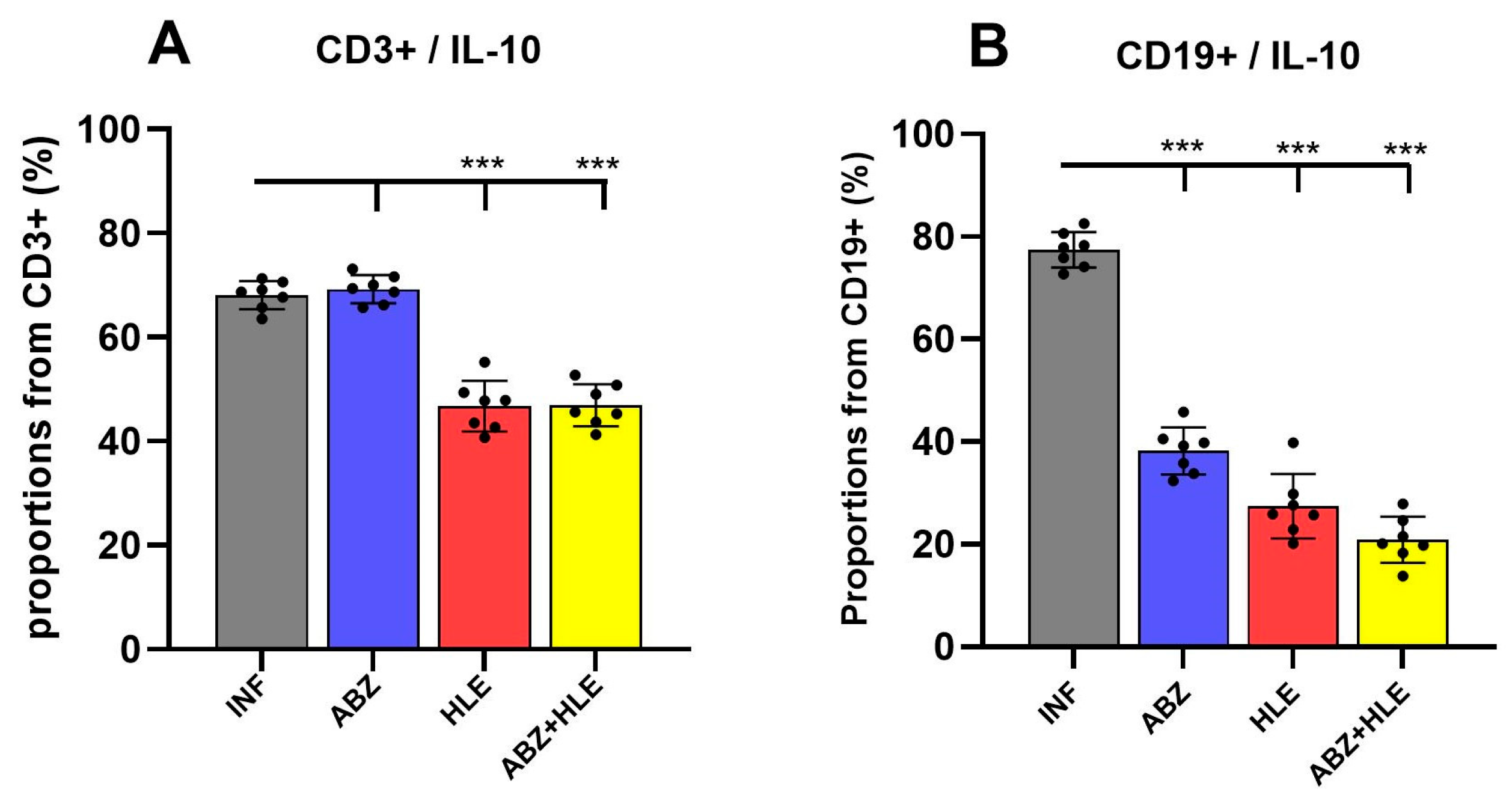
2.2. HLE and Combination Therapy Reduce the Frequency of IL-10-Producing T and B Lymphocytes
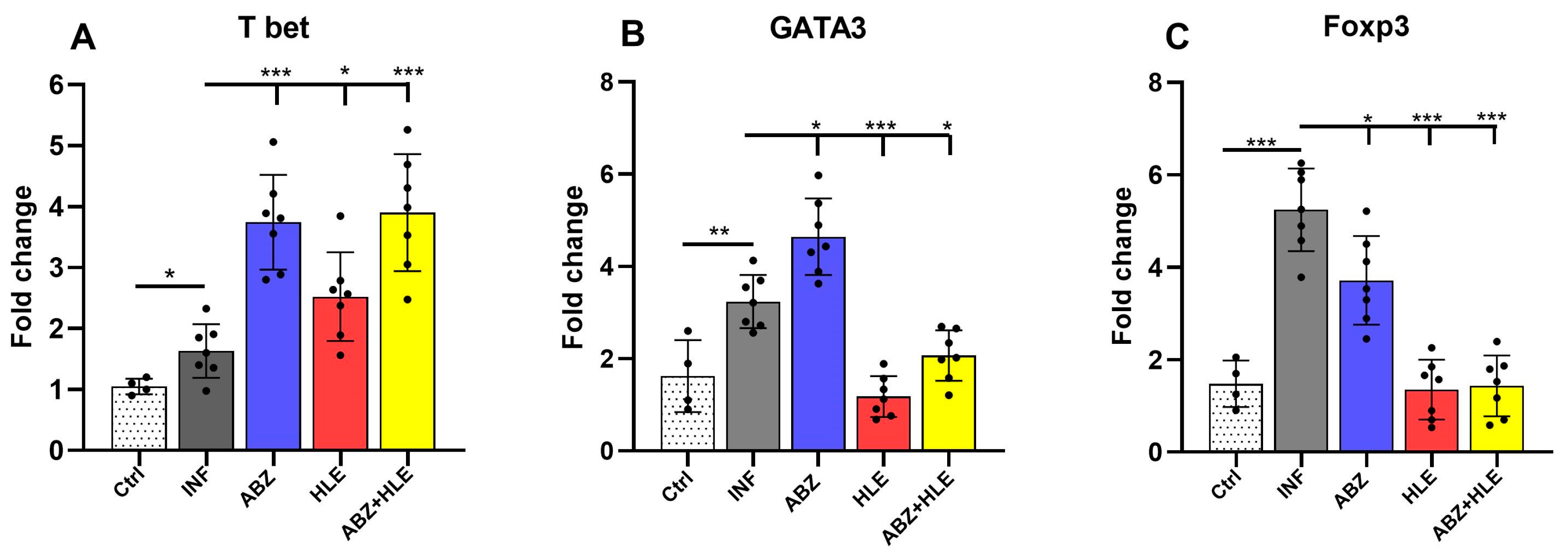
2.3. Gene Expression of Transcription Factors Regulating Th1/Th2/Treg Immunity
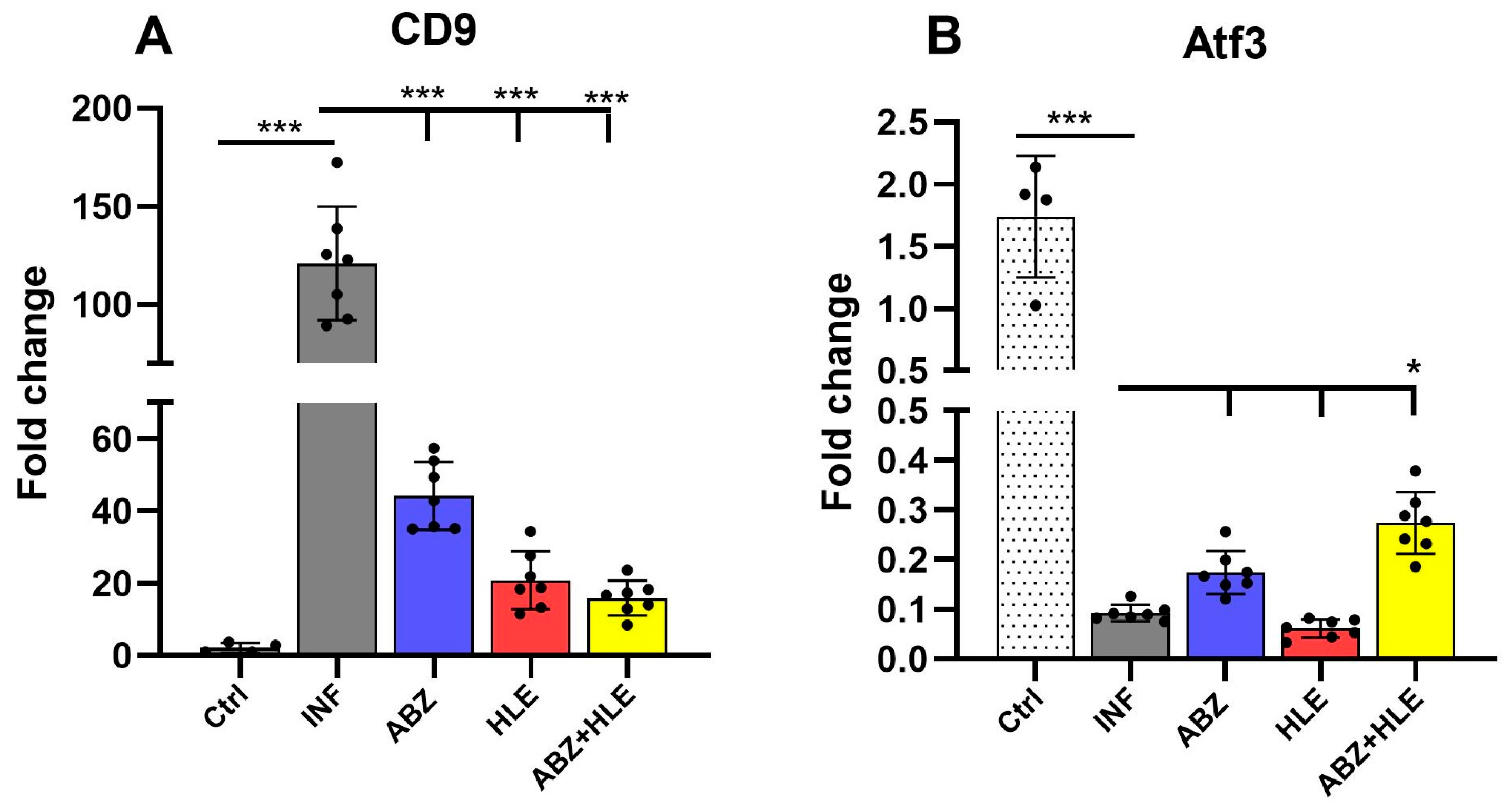
2.4. Gene Expression of Breg-Associated Markers
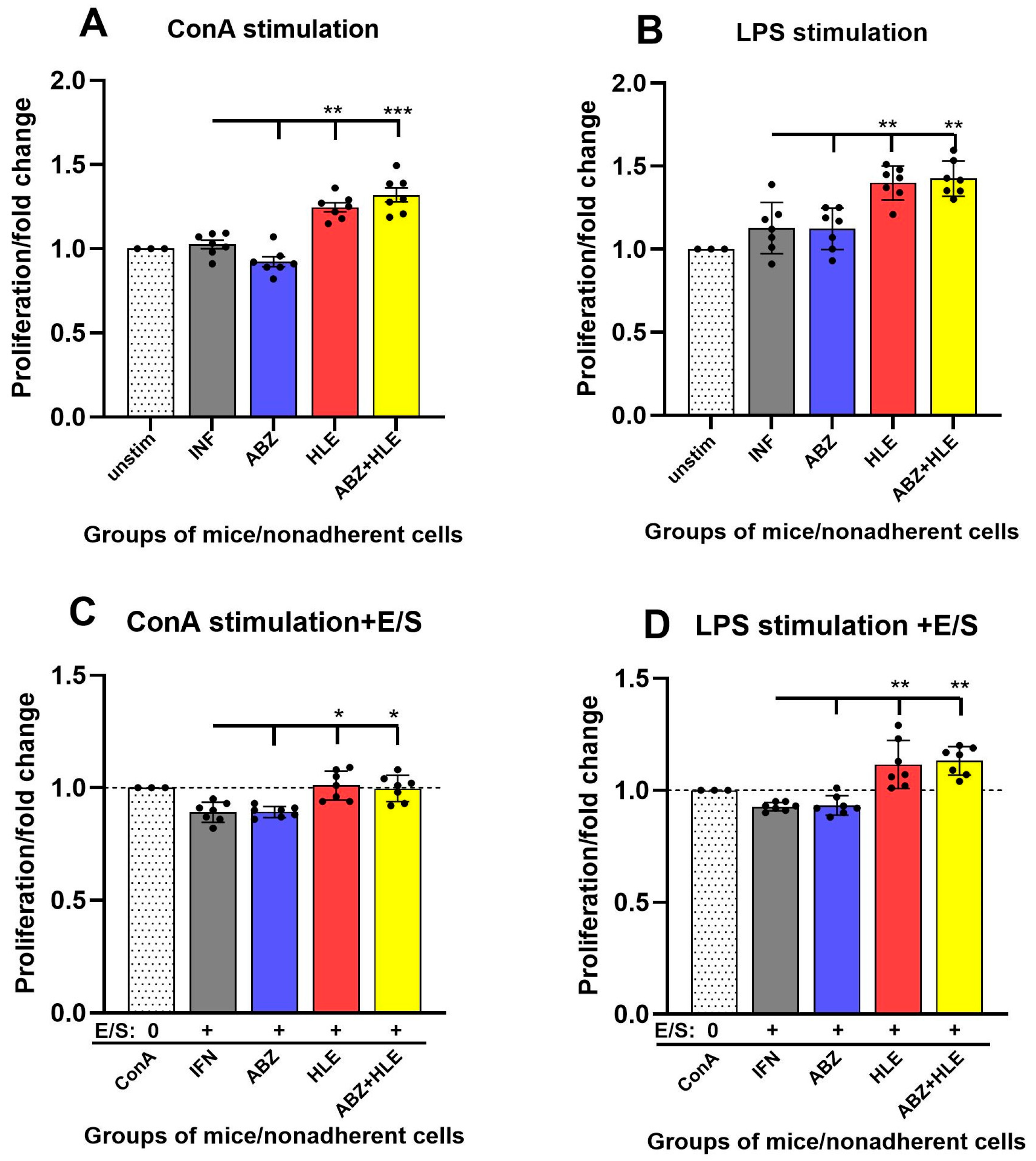
2.5. Proliferation of ConA- and LPS-Stimulated Lymphocytes and Response to E/S Antigens
2.6. Production of IFN-γ, IL-4, and IL-10 by Stimulated Peritoneal Lymphocytes
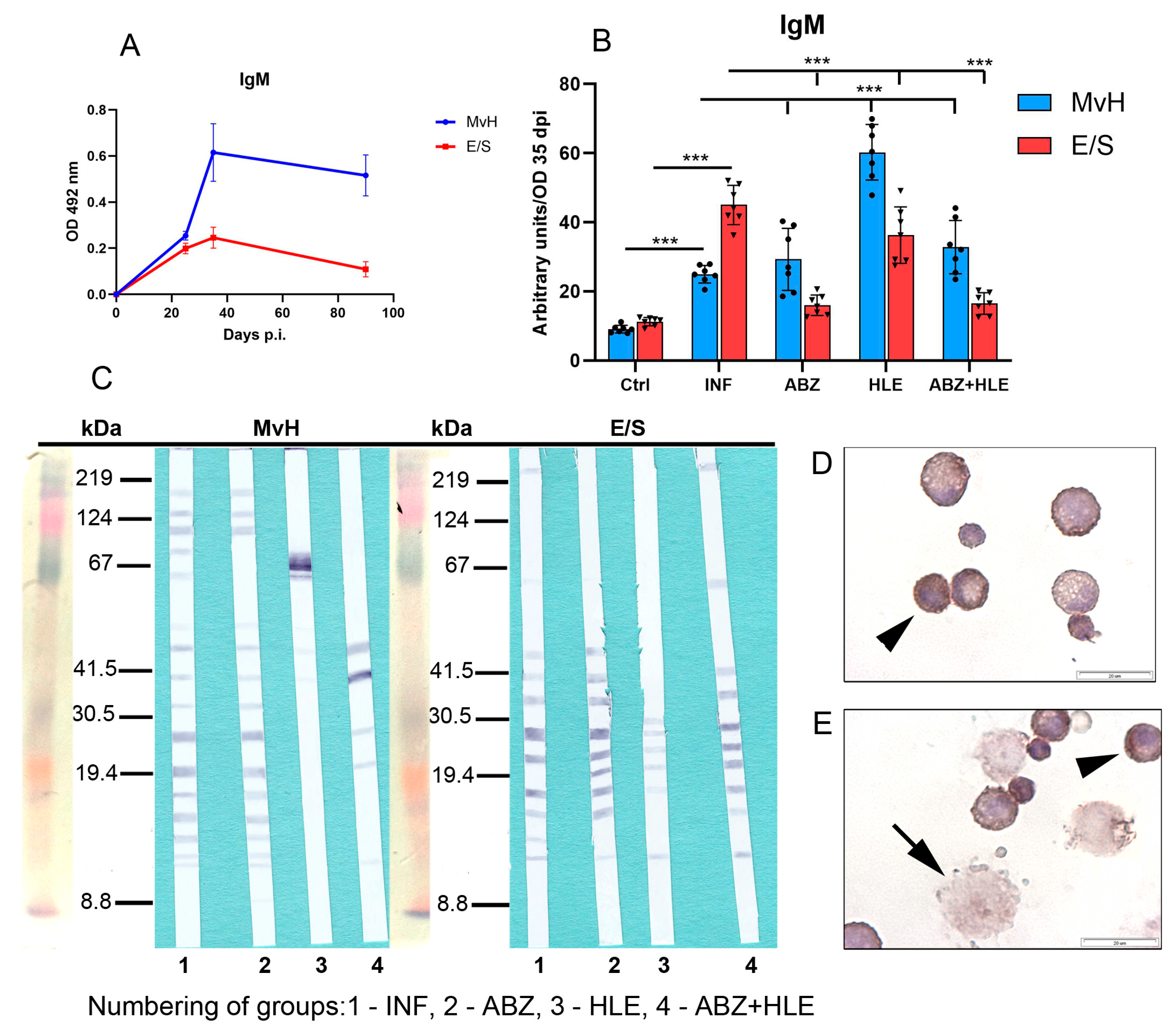
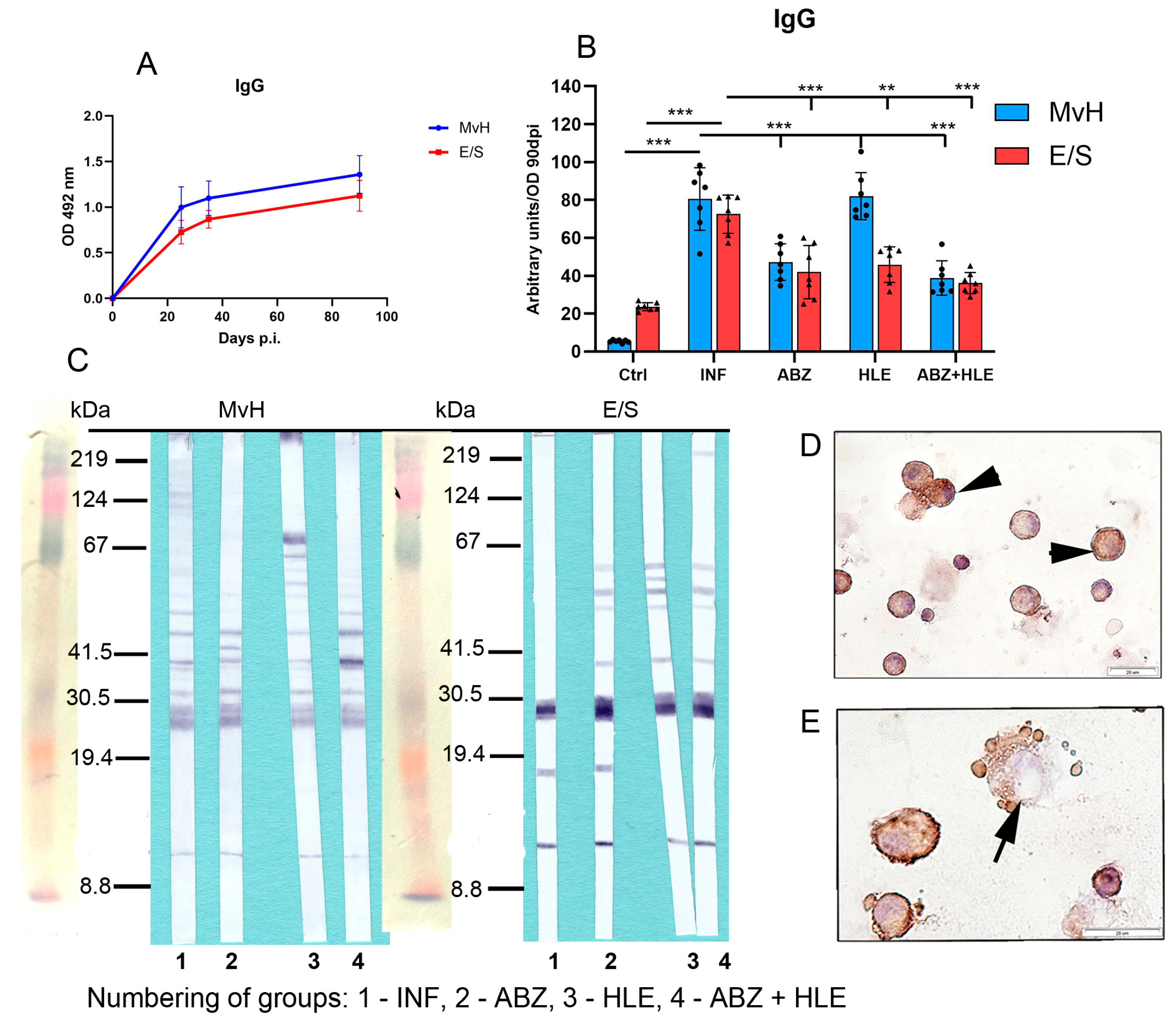
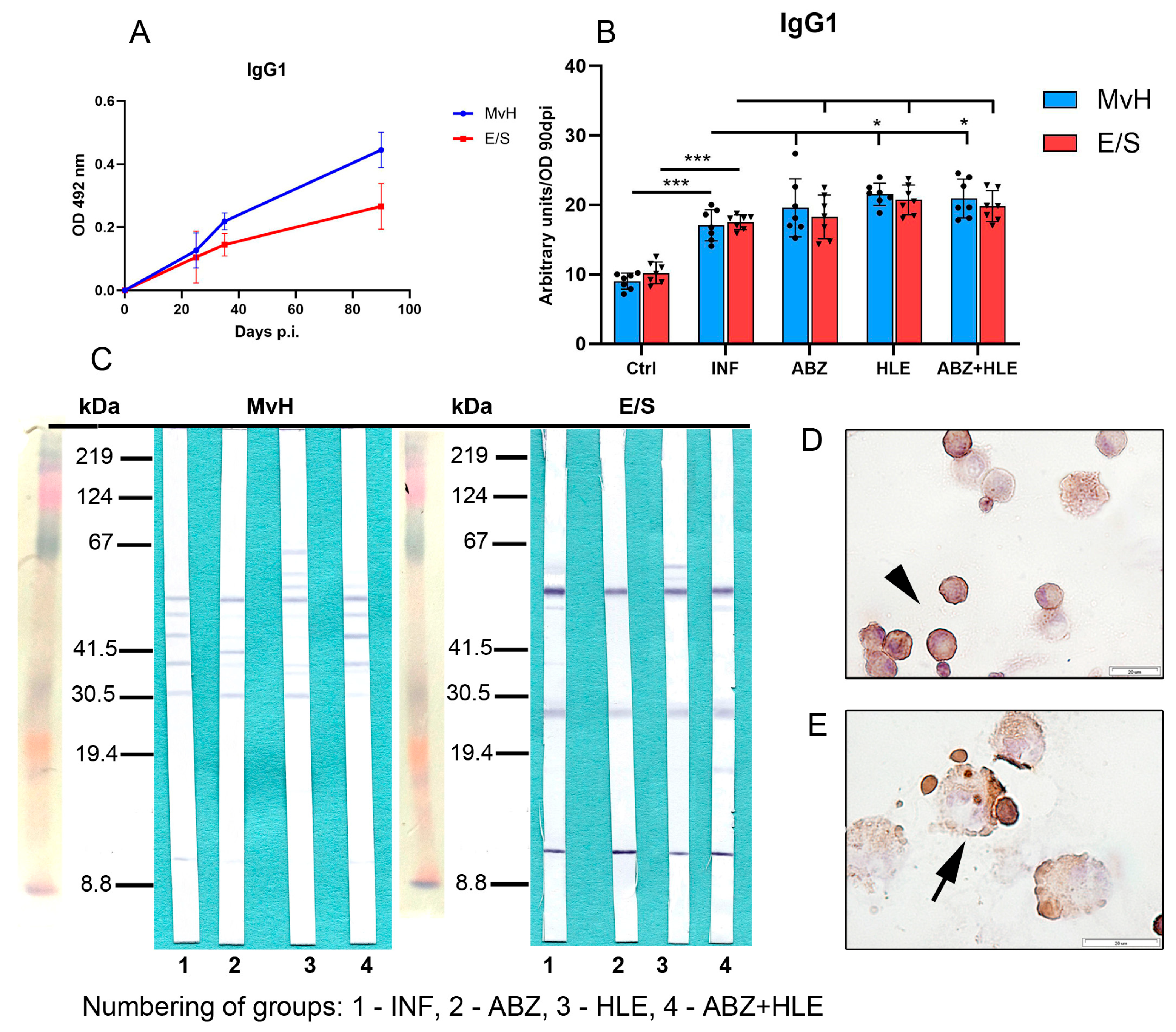
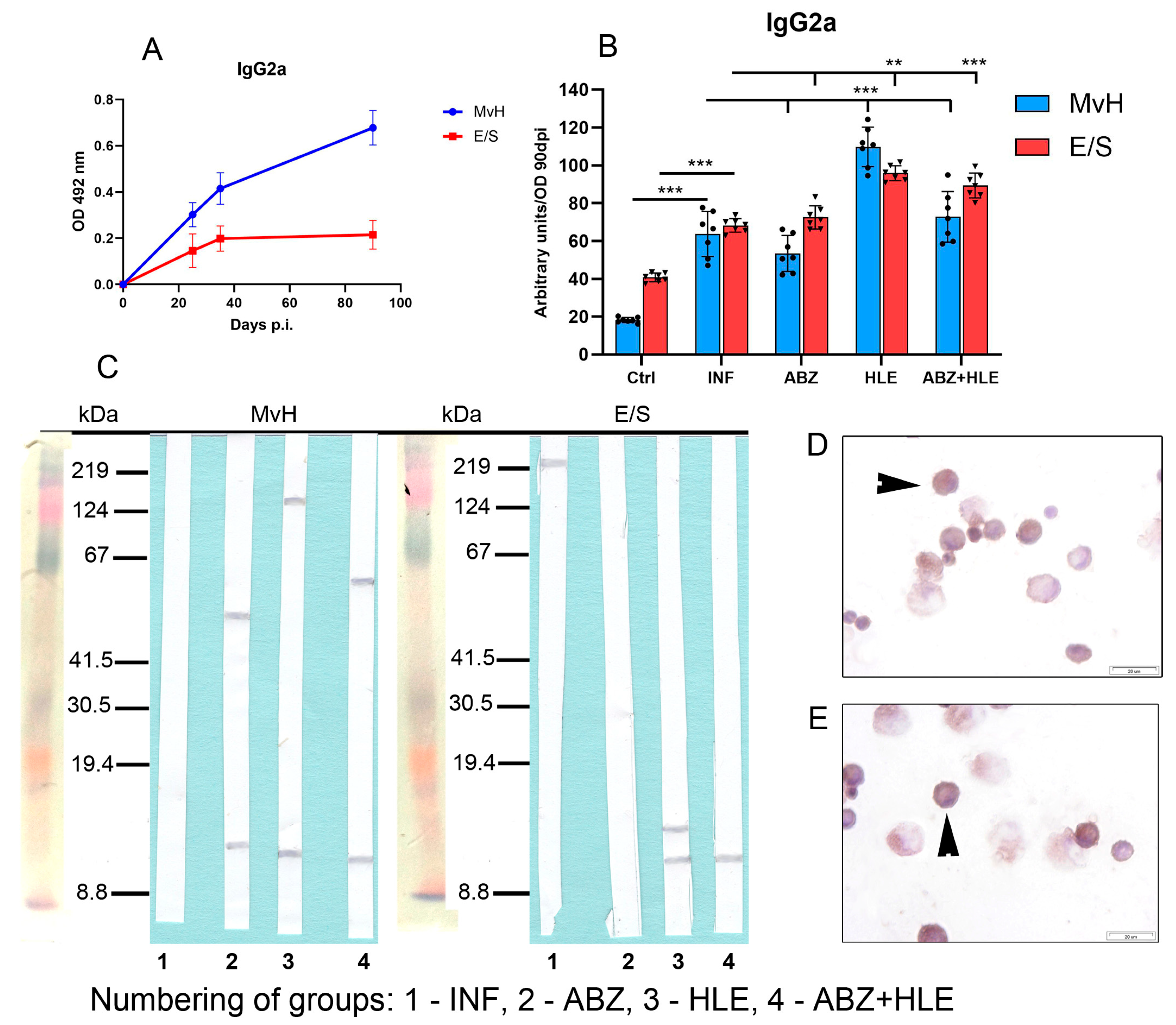
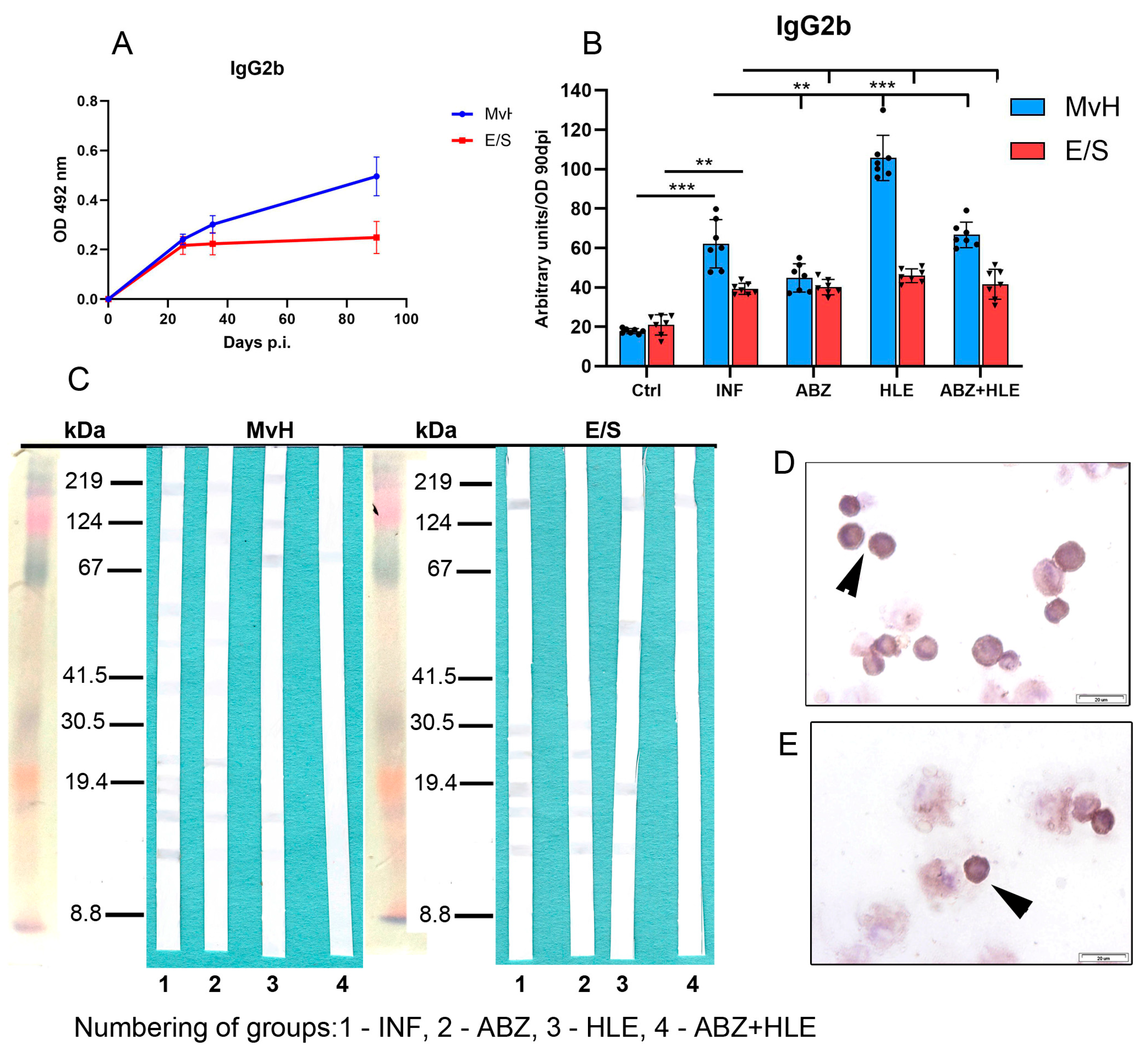
2.7. Analysis of Antibody Isotypes and Immunoreactive E/S and MvH Antigens in Peritoneal Exudates and Detection of Antibody Secreting Peritoneal Cells
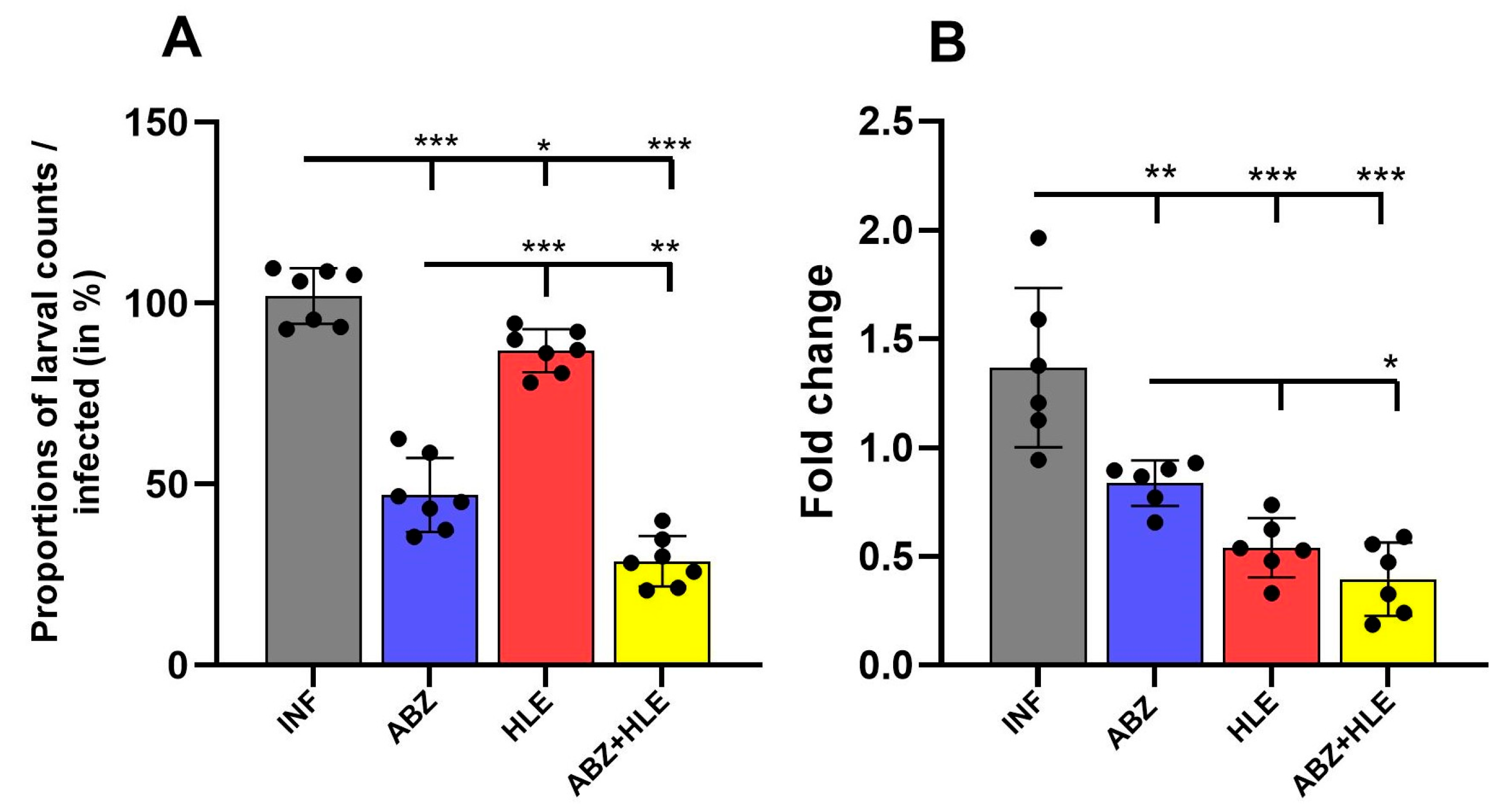
2.8. Efficacy of Treatments on Larval Burden and mRNA Levels of the Regulatory 14-3-3 Gene
3. Discussion
4. Materials and Methods
4.1. Albendazole and HLE
4.2. Infection and Experimental Design
4.3. Isolation of Peritoneal Non-Adherent Cell Populations and Exudates
4.4. Flow Cytometric Analysis
4.5. Cell Proliferation Assay
4.6. RNA Isolation and Real-Time qPCR
4.7. Preparation of Excretory/Secretory (E/S) and Larval Homogenate Antigens
4.8. Detection of Antibodies to E/S and MvH Antigens
4.9. Immunoblot Analysis
4.10. Immunocytochemical Staining
4.11. Larval Burden and Therapeutic Efficacy
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnaudov, A.; Kostova, Z. Dialysable leukocyte extracts in immunotherapy. Biotechnol. Biotechnol. Equip. 2015, 29, 1017–1023. [Google Scholar] [CrossRef]
- Estrada-Parra, S.; Nagaya, A.; Serrano, E.; Rodriguez, O.; Santamaria, V.; Ondarza, R.; Chavez, R.; Correa, B.; Monges, A.; Cabezas, R.; et al. Comparative study of transfer factor and acyclovir in the treatment of herpes zoster. Int. J. Immunopharmacol. 1998, 20, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ortega, C.; Dubed, M.; Ramos, Y.; Navea, L.; Alvarez, G.; Lobaina, L.; Lopez, L.; Casillas, D.; Rodriguez, L. Non-induced leukocyte extract reduces HIV replication and TNF secretion. Biochem. Biophys. Res. Commun. 2004, 325, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Fabre, R.A.; Pérez, T.M.; Aguilar, L.D.; Rangel, M.J.; Estrada-Garcìa, I.; Hernández-Pando, R.; Parra, S.E. Transfer factors as immunotherapy and supplement of chemotherapy in experimental pulmonary tuberculosis. Clin. Exp. Immunol. 2004, 136, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Espinosa, O.; Moreno-García, S.; Arce-Paredes, P.; Becerril-Villanueva, E.; Juárez-Ortega, M. Effect of dialyzable leukocyte extract, sodium butyrate, and valproic acid in the development of anergy in murine leprosy. Int. J. Mycobacteriol. 2020, 9, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Ciglanová, D.; Jurčacková, Z.; Mudroňová, D.; Dvorožňaková, E.; Hrčková, G. Differential Activity of Human Leukocyte Extract on Systemic Immune Response and Cyst Growth in Mice with Echinococcus Multilocularis Infection After Oral, Subcutaneous and Intraperitoneal Routes of Administration. Helminthologia 2022, 59, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Kubašková, T.M.; Mudroňová, D.; Velebný, S.; Hrčková, G. The utilisation of human dialyzable leukocyte extract (IMMODIN) as adjuvant in albendazole therapy on mouse model of larval cestode infection: Immunomodulatory and hepatoprotective effects. Int. Immunopharmacol. 2018, 65, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Viza, D.; Fudenberg, H.H.; Palareti, A.; Ablashi, D.; De Vinci, C.; Pizza, G. Transfer Factor: An Overlooked Potential for the Prevention and Treatment of Infectious Diseases. Folia Biol. (Prague) 2013, 59, 53–67. [Google Scholar] [CrossRef]
- Demečková, V.; Solar, P.; Hrčková, G.; Mudroňová, D.; Bojková, B.; Kassayová, M.; Gancarčiková, S. Immodin and its immune system supportive role in paclitaxel therapy of 4T1 mouse breast cancer. Biomed. Pharmacother. 2017, 89, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Esquivel, M.A.; Perez-Torres, A.; Romero-Romero, L.; Reyes-Matute, A.; Loaiza, B.; Mellado-Sanchez, G.; Pavon, L.; Medina-Rivero, E.; Pestell, R.G.; Perez-Tapia, S.M.; et al. The dialyzable leukocyte extract Transferon(TM) inhibits tumor growth and brain metastasis in a murine model of prostate cancer. Biomed. Pharmacother. 2018, 101, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Castrejon Vazquez, M.I.; Resendiz-Albor, A.A.; Ynga-Durand, M.A.; Martinez, I.M.A.; Orellana-Villazon, V.I.; Lopez, C.A.G.; Noguera, M.L.L.; Camano, M.E.V. Dialyzable Leukocyte Extract (Transferon) Administration in Sepsis: Experience from a Single Referral Pediatric Intensive Care Unit. BioMed Res. Int. 2019, 2019, 8980506. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, L.G.; Panameno-Cruz, V.; Luna-Arias, J.P.; Reyna-Beltran, E. Implementation of Dialyzable Leukocyte Extract to Treat Multiple Sclerosis: A Clinical Case. Clin. Case Rep. 2024, 12, e70016. [Google Scholar] [CrossRef] [PubMed]
- Hana, I.; Vrubel, J.; Pekarek, J.; Cech, K. The influence of age on transfer factor treatment of cellular immunodeficiency, chronic fatigue syndrome and/or chronic viral infections. Biotherapy 1996, 9, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Homberg, T.A.; Lara, I.; Andaluz, C.; Cervantes-Trujano, E.; Hernandez-Martinez, P.M.; Perez-Tapia, S.M.; Jimenez-Martinez, M.C. Quality of life in adult patients using dialyzable leukocyte extract for allergic rhinitis. Medicine (Baltimore) 2023, 102, e34186. [Google Scholar] [CrossRef] [PubMed]
- Budke, C.M.; White, A.C., Jr.; Garcia, H.H. Zoonotic larval cestode infections: Neglected, neglected tropical diseases? PLoS Neglected Trop. Dis. 2009, 3, e319. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.V.; Galán-Puchades, M.T.; Malone, J.B. Short report: A new case report of human Mesocestoides infection in the United States. Am. J. Trop. Med. Hyg. 2003, 68, 566–567. [Google Scholar] [CrossRef] [PubMed]
- Eckert, J.; Deplazes, P. Biological, epidemiological, and clinical aspects of echinococcosis, a zoonosis of increasing concern. Clin. Microbiol. Rev. 2004, 17, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Filho, P.T.H.; Rodriguez-Rivas, R.; Fleury, A. Neurocysticercosis: A Review into Treatment Options, Indications, and Their Efficacy. Res. Rep. Trop. Med. 2022, 13, 67–79. [Google Scholar] [CrossRef] [PubMed]
- White, T.R.; Thompson, R.C.; Penhale, W.J. A comparative study of the susceptibility of inbred strains of mice to infection with Mesocestoides corti. Int. J. Parasitol. 1982, 12, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Hemphill, A.; Stadelmann, B.; Rufener, R.; Spiliotis, M.; Boubaker, G.; Müller, J.; Müller, N.; Gorgas, D.; Gottstein, B. Treatment of echinococcosis: Albendazole and mebendazole—What else? Parasite 2014, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Popova, G.; Vuchev, D.; Anichina, K. Treatment of hepatic and pulmonary hydatidosis with albendazole and praziquantel. Helminthologia 2023, 60, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Vuitton, D.A. Benzimidazoles for the treatment of cystic and alveolar echinococcosis: What is the consensus? Expert Rev. Anti-Infect. Ther. 2009, 7, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Kern, P. Medical treatment of echinococcosis under the guidance of Good Clinical Practice (GCP/ICH). Parasitol. Int. 2006, 55, S273–S282. [Google Scholar] [CrossRef] [PubMed]
- Son, D.S.; Lee, E.S.; Adunyah, S.E. The Antitumor Potentials of Benzimidazole Anthelmintics as Repurposing Drugs. Immune Netw. 2020, 20, e29. [Google Scholar] [CrossRef] [PubMed]
- Kubašková, T.M.; Mudroňová, D.; Vargová, M.; Reiterová, K.; Hrčková, G. Cellular and humoral peritoneal immunity to Mesocestoides vogae metacestode infection in mice. Parasites Vectors 2021, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Vendelova, E.; Lutz, M.B.; Hrčková, G. Immunity and immune modulation elicited by the larval cestode Mesocestoides vogae and its products. Parasite Immunol. 2015, 37, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Smith-Garvin, J.E.; Koretzky, G.A.; Jordan, M.S. T cell activation. Annu. Rev. Immunol. 2009, 27, 591–619. [Google Scholar] [CrossRef] [PubMed]
- Lund, F.E. Cytokine-producing B lymphocytes-key regulators of immunity. Curr. Opin. Immunol. 2008, 20, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Fillatreau, S. Regulatory functions of B cells and regulatory plasma cells. Biomed. J. 2019, 42, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Nono, J.K.; Lutz, M.B.; Brehm, K. Expansion of Host Regulatory T Cells by Secreted Products of the Tapeworm Echinococcus multilocularis. Front. Immunol. 2020, 11, 798. [Google Scholar] [CrossRef] [PubMed]
- Kallies, A.; Good-Jacobson, K.L. Transcription Factor T-bet Orchestrates Lineage Development and Function in the Immune System. Trends Immunol. 2017, 38, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y. GATA3: A master of many trades in immune regulation. Trends Immunol. 2014, 35, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Long, J.; Huang, M.X.; Luo, P.Y.; Bian, Z.H.; Xu, Y.F.; Wang, C.B.; Yang, S.H.; Li, L.; Selmi, C.; et al. Characterization of Organ-Specific Regulatory B Cells Using Single-Cell RNA Sequencing. Front. Immunol. 2021, 12, 711980. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Smits, H.H.; McSorley, H.J. Modulation of Host Immunity by Helminths: The Expanding Repertoire of Parasite Effector Molecules. Immunity 2018, 49, 801–818. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.M. IgG subclass co-expression brings harmony to the quartet model of murine IgG function. Immunol. Cell Biol. 2016, 94, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Snapper, C.M.; Mond, J.J. Towards a comprehensive view of immunoglobulin class switching. Immunol. Today 1993, 14, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Siles-Lucas Mdel, M.; Gottstein, B. The 14-3-3 protein: A key molecule in parasites as in other organisms. Trends Parasitol. 2003, 19, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Hrčková, G.; Kubašková, T.M.; Mudroňová, D.; Jurčacková, Z.; Ciglanová, D. Co-Treatment with Human Leukocyte Extract and Albendazole Stimulates Drug’s Efficacy and Th1 Biased Immune Response in Mesocestoides vogae (Cestoda) Infection via Modulation of Transcription Factors, Macrophage Polarization, and Cytokine Profiles. Pharmaceutics 2023, 15, 541. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, A.E.; Kerepesi, L.A.; Vandergrift, G.L.; Herbert, D.R.; Van Winkle, T.J.; Hooper, D.C.; Pearce, E.J.; Abraham, D. IL-4-/- mice with lethal Mesocestoides corti infections—reduced Th2 cytokines and alternatively activated macrophages. Parasite Immunol. 2009, 31, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Peon, A.N.; Espinoza-Jimenez, A.; Terrazas, L.I. Immunoregulation by Taenia crassiceps and its antigens. BioMed Res. Int. 2013, 2013, 498583. [Google Scholar] [CrossRef] [PubMed]
- Vendelova, E.; de Lima, J.C.; Lorenzatto, K.R.; Monteiro, K.M.; Mueller, T.; Veepaschit, J.; Grimm, C.; Brehm, K.; Hrčková, G.; Lutz, M.B.; et al. Proteomic Analysis of Excretory-Secretory Products of Mesocestoides corti Metacestodes Reveals Potential Suppressors of Dendritic Cell Functions. PLoS Neglected Trop. Dis. 2016, 10, e0005061. [Google Scholar] [CrossRef] [PubMed]
- Dorshkind, K.; Montecino-Rodriguez, E. Fetal B-cell lymphopoiesis and the emergence of B-1-cell potential. Nat. Rev. Immunol. 2007, 7, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.I.; Catalan-Dibene, J.; Zlotnik, A. B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine 2015, 74, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. Regulatory B Cells: Origin, Phenotype, and Function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Oishi, S.; Takano, R.; Tamura, S.; Tani, S.; Iwaizumi, M.; Hamaya, Y.; Takagaki, K.; Nagata, T.; Seto, S.; Horii, T.; et al. M2 polarization of murine peritoneal macrophages induces regulatory cytokine production and suppresses T-cell proliferation. Immunology 2016, 149, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Dvorožnáková, E.; Porubcová, J.; Ševcíková, Z. Immune response of mice with alveolar echinococcosis to therapy with transfer factor, alone and in combination with albendazole. Parasitol. Res. 2009, 105, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Ramirez, D.; Vadillo, E.; Arriaga-Pizano, L.A.; Mayani, H.; Estrada-Parra, S.; Velasco-Velazquez, M.A.; Perez-Tapia, S.M.; Pelayo, R. Early Differentiation of Human CD11c(+)NK Cells with gammadelta T Cell Activation Properties Is Promoted by Dialyzable Leukocyte Extracts. J. Immunol. Res. 2016, 2016, 4097642. [Google Scholar] [CrossRef] [PubMed]
- Ricken, F.J.; Nell, J.; Grüner, B.; Schmidberger, J.; Kaltenbach, T.; Kratzer, W.; Hillenbrand, A.; Henne-Bruns, D.; Deplazes, P.; Moller, P.; et al. Albendazole increases the inflammatory response and the amount of Em2-positive small particles of Echinococcus multilocularis (spems) in human hepatic alveolar echinococcosis lesions. PLoS Neglected Trop. Dis. 2017, 11, e0005636. [Google Scholar] [CrossRef] [PubMed]
- Tiemessen, M.M.; Jagger, A.L.; Evans, H.G.; van Herwijnen, M.J.C.; John, S.; Taams, L.S. CD4-CD25-Foxp3 regulatory T cells induce alternative activation of human monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 19446–19451. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Müller, S.; Lin, R.Y.; Siffert, M.; Vuitton, D.A.; Wen, H.; Gottstein, B. Depletion of FoxP3+Tregs improves control of larval Echinococcus multilocularis infection by promoting co-stimulation and Th1/17 immunity. Immun. Inflamm. Dis. 2017, 5, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Portugall, I.; Layland, L.E. TLRs, Treg, and B cells, an interplay of regulation during helminth infection. Front. Immunol. 2012, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, H.; Ni, Y.; Li, C.; Xu, X.; Chang, H.; Xu, Z.; Hou, M.; Ji, M. Helminth-induced CD9(+) B-cell subset alleviates obesity-associated inflammation via IL-10 production. Int. J. Parasitol. 2022, 52, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.C.; Cheng, C.F. Master Regulator Activating Transcription Factor 3 (ATF3) in Metabolic Homeostasis and Cancer. Front. Endocrinol. 2020, 11, 556. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.Y.; Chang, C.P. Lectin of Concanavalin A as an anti-hepatoma therapeutic agent. J. Biomed. Sci. 2009, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.H. T cell anergy. Annu. Rev. Immunol. 2003, 21, 305–334. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Li, L.; Dong, D.; Wang, L.; Wang, X.; Yang, K.; Xu, X.; Chen, C.; Wu, X.; Chen, X. Glycomolecules in Echinococcus granulosus cyst fluid inhibit TLR4-mediated inflammatory responses via c-Raf. Cell Mol. Immunol. 2020, 17, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Gruner, B.; Peters, L.; Hillenbrand, A.; Vossberg, P.; Schweiker, J.; Rollmann, E.G.; Rodriguez, L.H.; Blumhardt, J.; Burkert, S.; Kern, P.; et al. Echinococcus multilocularis specific antibody, systemic cytokine, and chemokine levels, as well as antigen-specific cellular responses in patients with progressive, stable, and cured alveolar echinococcosis: A 10-year follow-up. PLoS Neglected Trop. Dis. 2022, 16, e0010099. [Google Scholar] [CrossRef] [PubMed]
- Rigáno, R.; Profumo, E.; Ioppolo, S.; Notargiacomo, S.; Teggi, A.; Siracusano, A. Serum cytokine detection in the clinical follow up of patients with cystic echinococcosis. Clin. Exp. Immunol. 1999, 115, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Rawat, J.; Dixon, J.B.; Macintyre, A.R.; McGarry, H.F.; Taylor, M.J. IL-4 dependent resistance to the tapeworm Mesocestoides corti (Cestoda) in mice. Parasite Immunol. 2003, 25, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, P.; Dixon, J.B.; Haywood, S.; Rakha, N.K.; Carter, S.D. Differential Regulation of Murine Mesocestoides corti Infection by Bacterial Lipopolysaccharide and Interferon-Gamma. Parasitology 1991, 102, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Abraham, K.M.; Teale, J.M. Isotype Restriction during Infection of Mice with the Cestode Mesocestoides corti—Role of Immune Suppression. J. Immunol. 1987, 138, 1699–1704. [Google Scholar] [CrossRef] [PubMed]
- Zakroff, S.G.H.; Beck, L.; Platzer, E.G.; Spiegelberg, H.L. The Ige and Igg Subclass Responses of Mice to 4 Helminth-Parasites. Cell Immunol. 1989, 119, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Zuniga-Navarrete, F.; Zavala-Meneses, S.G.; Zelnik, V.; Kopacek, J.; Skultety, L. Initial proteomic characterization of IMMODIN, commercially available dialysable leukocytes extract. Chem. Pap. 2021, 75, 1959–1968. [Google Scholar] [CrossRef]
- Matsumoto, J.; Muller, N.; Hemphill, A.; Oku, Y.; Kamiya, M.; Gottstein, B. 14-3-3- and II/3-10-gene expression as molecular markers to address viability and growth activity of Echinococcus multilocularis metacestodes. Parasitology 2006, 132 Pt 1, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lighvani, A.A.; Frucht, D.M.; Jankovic, D.; Yamane, H.; Aliberti, J.; Hissong, B.D.; Nguyen, B.V.; Gadina, M.; Sher, A.; Paul, W.E.; et al. T-bet is rapidly induced by interferon-γ in lymphoid and myeloid cells. Proc. Natl. Acad. Sci. USA 2001, 98, 15137–15142. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.P.; Flavell, R.A. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell 1997, 89, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Ichiyama, K.; Yoshida, H.; Wakabayashi, Y.; Chinen, T.; Saeki, K.; Nakaya, M.; Takaesu, G.; Hori, S.; Yoshimura, A.; Kobayashi, T. Foxp3 inhibits RORγt-mediated IL-17A mRNA transcription through direct interaction with RORγt. J. Biol. Chem. 2008, 283, 17003–17008. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Chen, J.; Hai, T. The regulation of ATF3 gene expression by mitogen-activated protein kinases. Biochem. J. 2007, 401, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Won, W.J.; Kearney, J.F. CD9 is a unique marker for marginal zone B cells, B1 cells, and plasma cells in mice. J. Immunol. 2002, 168, 5605–5611. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Geng, J.; Gao, J.H.; Zhao, H.; Li, J.H.; Shi, Y.R.; Yang, B.Y.; Xiao, C.; Linghu, Y.Y.; Sun, X.F.; et al. Macrophage achieves self-protection against oxidative stress-induced ageing through the Mst-Nrf2 axis. Nat. Commun. 2019, 10, 755. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hrčková, G.; Mudroňová, D.; Reiterová, K.; Cavallero, S.; Bellini, I. Human Dialyzable Leukocyte Extract Enhances Albendazole Efficacy and Promotes Th1/Th2-Biased Lymphocyte and Antibody Responses in Peritoneal Cavity of Murine Model of Mesocestoides vogae Infection. Int. J. Mol. Sci. 2025, 26, 6994. https://doi.org/10.3390/ijms26146994
Hrčková G, Mudroňová D, Reiterová K, Cavallero S, Bellini I. Human Dialyzable Leukocyte Extract Enhances Albendazole Efficacy and Promotes Th1/Th2-Biased Lymphocyte and Antibody Responses in Peritoneal Cavity of Murine Model of Mesocestoides vogae Infection. International Journal of Molecular Sciences. 2025; 26(14):6994. https://doi.org/10.3390/ijms26146994
Chicago/Turabian StyleHrčková, Gabriela, Dagmar Mudroňová, Katarína Reiterová, Serena Cavallero, and Ilaria Bellini. 2025. "Human Dialyzable Leukocyte Extract Enhances Albendazole Efficacy and Promotes Th1/Th2-Biased Lymphocyte and Antibody Responses in Peritoneal Cavity of Murine Model of Mesocestoides vogae Infection" International Journal of Molecular Sciences 26, no. 14: 6994. https://doi.org/10.3390/ijms26146994
APA StyleHrčková, G., Mudroňová, D., Reiterová, K., Cavallero, S., & Bellini, I. (2025). Human Dialyzable Leukocyte Extract Enhances Albendazole Efficacy and Promotes Th1/Th2-Biased Lymphocyte and Antibody Responses in Peritoneal Cavity of Murine Model of Mesocestoides vogae Infection. International Journal of Molecular Sciences, 26(14), 6994. https://doi.org/10.3390/ijms26146994