
Targeting Vascular and Inflammatory Crosstalk: Cannabigerol as a Dual-Pathway Modulator in Rosacea


Abstract
1. Introduction
2. Results
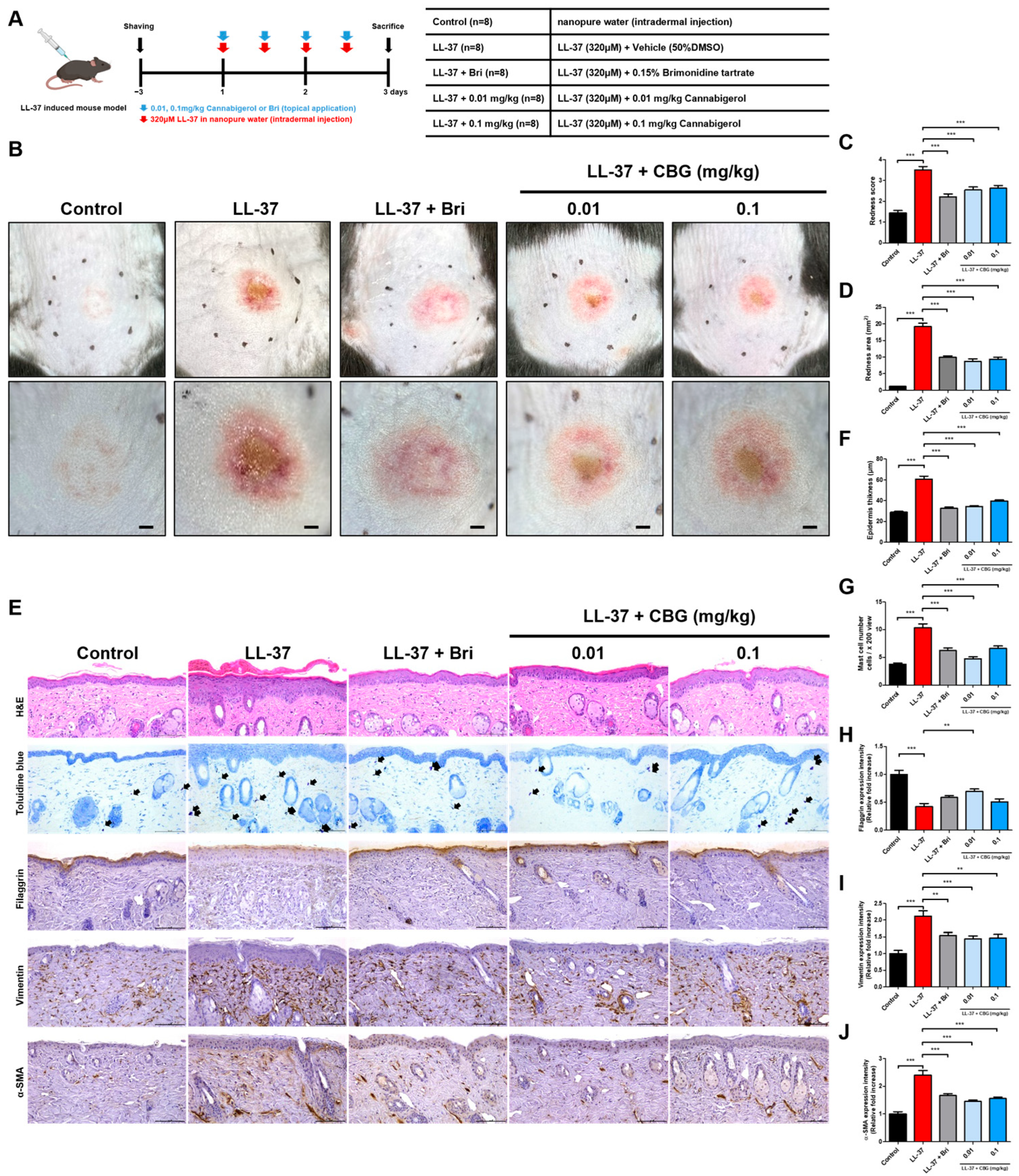
2.1. Cannabigerol Alleviates Redness and Inflammation in LL-37-Induced Rosacea Mouse Model
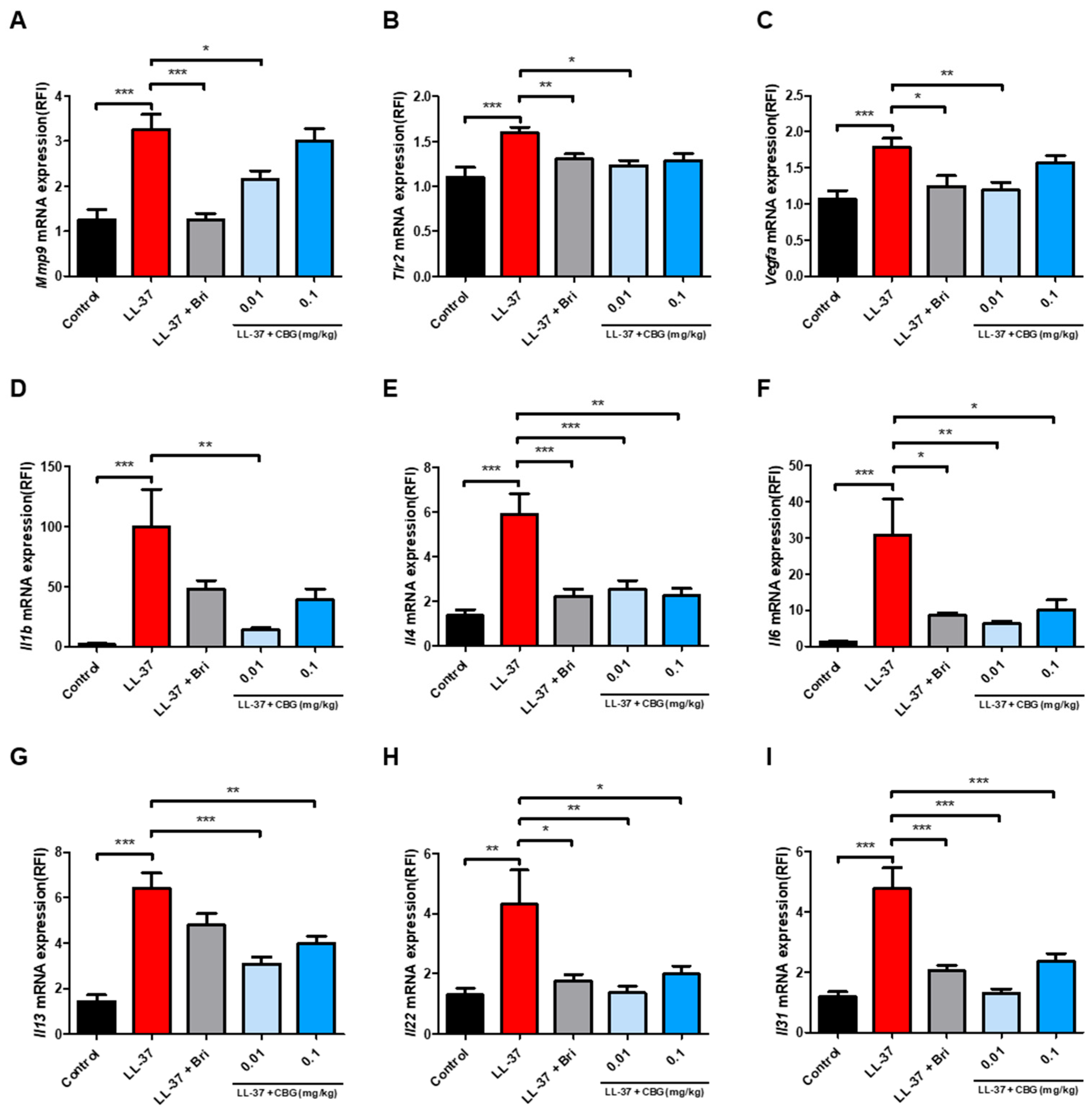
2.2. Cannabigerol Suppresses Inflammatory and Angiogenic Gene Expression
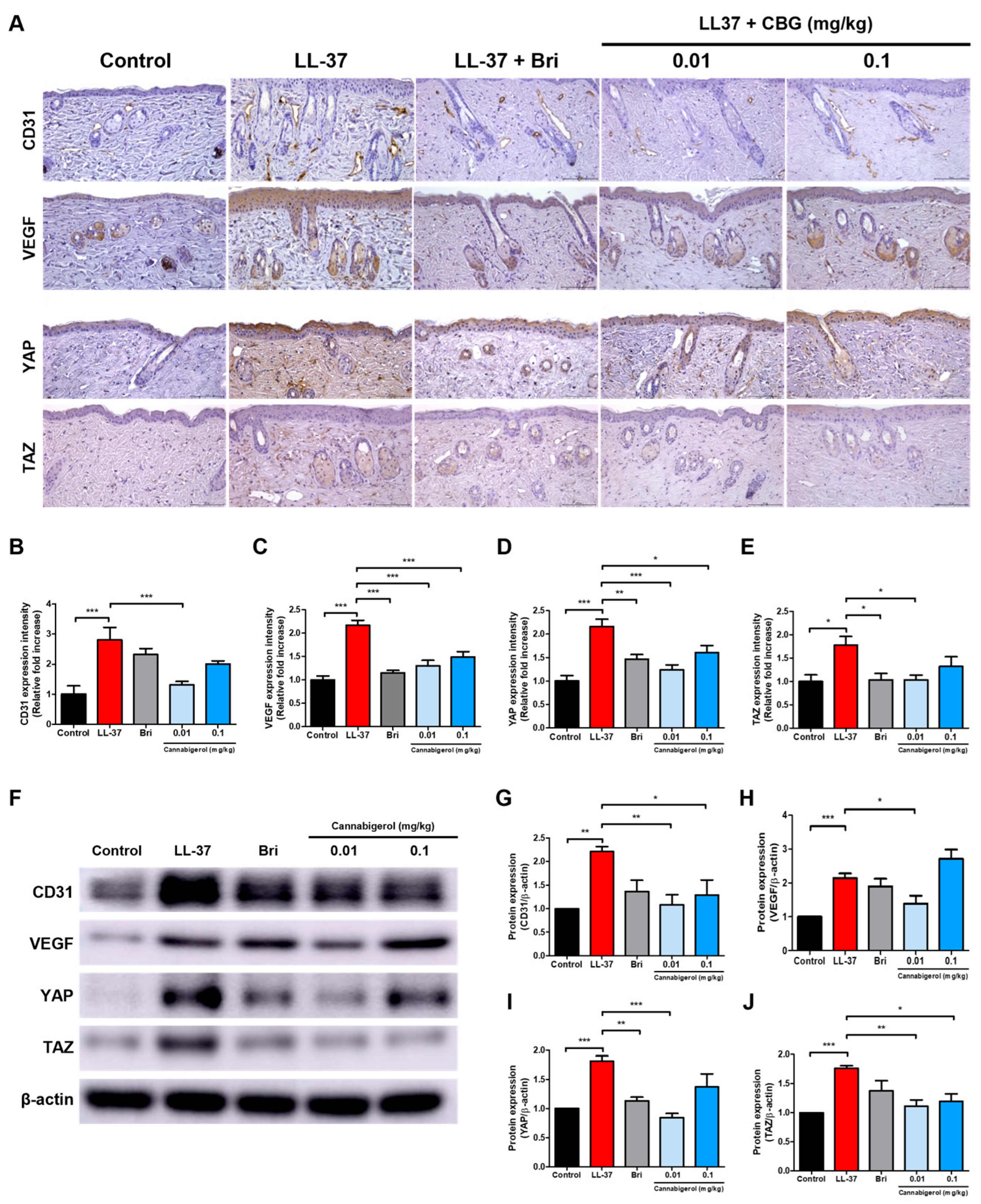
2.3. Cannabigerol Inhibits Vascular Activation and YAP/TAZ Signaling Pathway
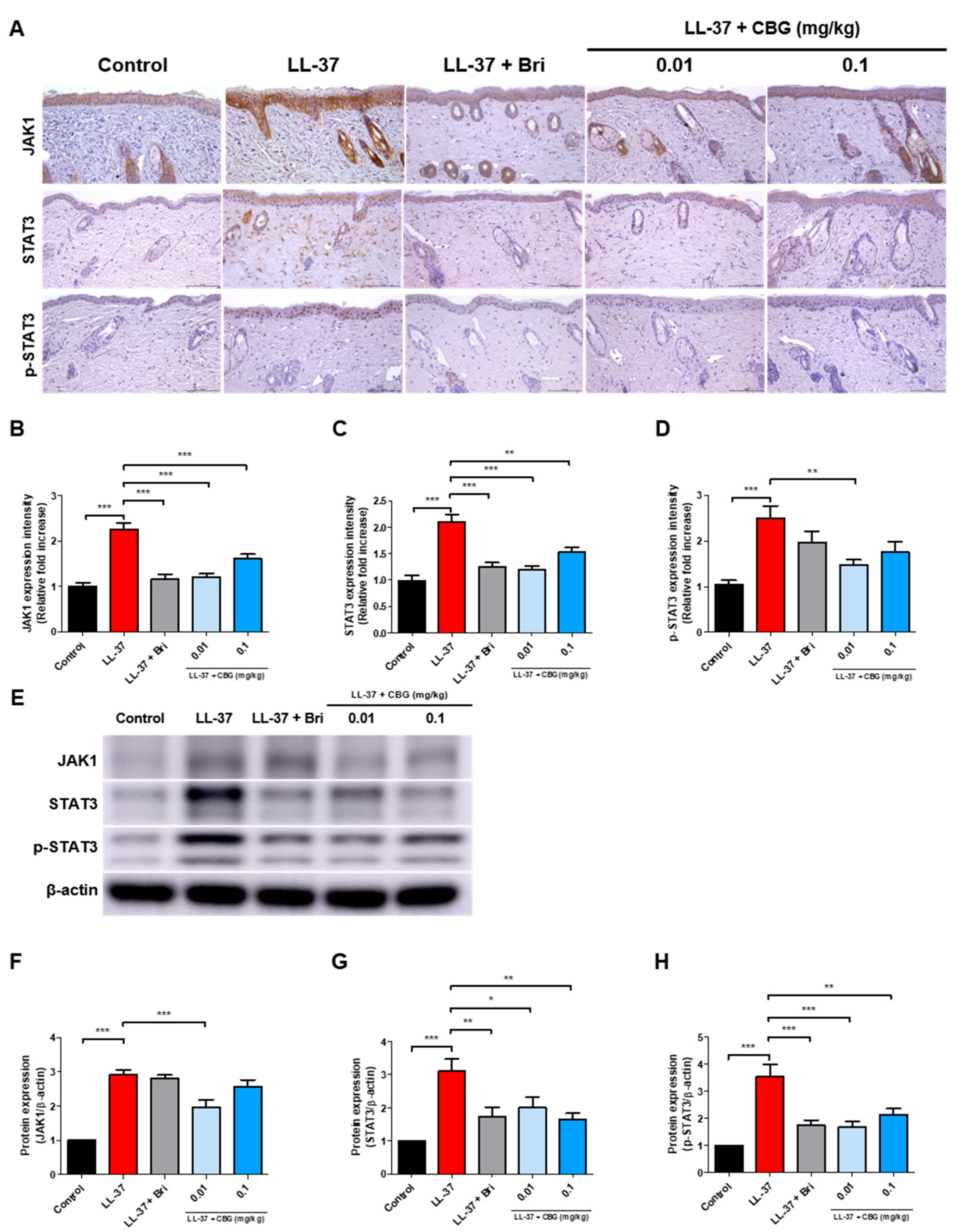
2.4. Cannabigerol Inhibits JAK/STAT Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Animal Study
4.2. Histological Analysis
4.3. Quantitative Real-Time PCR (qPCR) Analysis
4.4. Immunohistochemical Analysis
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Zuuren, E.J. Rosacea. N. Engl. J. Med. 2017, 377, 1754–1764. [Google Scholar] [CrossRef]
- Picardo, M.; Eichenfield, L.F.; Tan, J. Acne and Rosacea. Dermatol. Ther. 2017, 7, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Gallo, R.L. The molecular pathology of rosacea. J. Dermatol. Sci. 2009, 55, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Schwab, V.D.; Sulk, M.; Seeliger, S.; Nowak, P.; Aubert, J.; Mess, C.; Rivier, M.; Carlavan, I.; Rossio, P.; Metze, D.; et al. Neurovascular and neuroimmune aspects in the pathophysiology of rosacea. J. Investig. Dermatol. Symp. Proc. 2011, 15, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.S.; Huang, W.W. Rosacea Pathogenesis. Dermatol. Clin. 2018, 36, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Crawford, G.H.; Pelle, M.T.; James, W.D. Rosacea: I. Etiology, pathogenesis, and subtype classification. J. Am. Acad. Dermatol. 2004, 51, 327–341; quiz 342–324. [Google Scholar] [CrossRef] [PubMed]
- Engin, B.; Özkoca, D.; Kutlubay, Z.; Serdaroğlu, S. Conventional and Novel Treatment Modalities in Rosacea. Clin. Cosmet. Investig. Dermatol. 2020, 13, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Liu, Y.; Zhang, J.; Liu, Y.; Li, M.; Tao, M.; Zhang, Y.; Tang, Z.; Liu, W.; Xu, Y. Mast cell stabilization: New mechanism underlying the therapeutic effect of intense pulsed light on rosacea. Inflamm. Res. 2023, 72, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Dall’Oglio, F.; Nasca, M.R.; Gerbino, C.; Micali, G. Advances in pharmacotherapy for rosacea: What is the current state of the art? Expert. Opin. Pharmacother. 2022, 23, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Norooznezhad, A.H.; Norooznezhad, F. Cannabinoids: Possible agents for treatment of psoriasis via suppression of angiogenesis and inflammation. Med. Hypotheses 2017, 99, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Wang, Z.; Zhang, R.; Zhu, Y.; Lin, G.; Li, R.; Zhang, J. The antinociceptive activity and mechanism of action of cannabigerol. Biomed. Pharmacother. 2023, 158, 114163. [Google Scholar] [CrossRef] [PubMed]
- Calapai, F.; Cardia, L.; Esposito, E.; Ammendolia, I.; Mondello, C.; Lo Giudice, R.; Gangemi, S.; Calapai, G.; Mannucci, C. Pharmacological Aspects and Biological Effects of Cannabigerol and Its Synthetic Derivatives. Evid.-Based Complement. Altern. Med. 2022, 2022, 3336516. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.H.; Kim, K.C.; Lee, J.H. Anti-Inflammatory Effects of Cannabigerol In Vitro and In Vivo Are Mediated Through the JAK/STAT/NFκB Signaling Pathway. Cells 2025, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Hong, Y.J.; Kim, M. Angiogenesis in Chronic Inflammatory Skin Disorders. Int. J. Mol. Sci. 2021, 22, 12035. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Werbel, T.; Wang, Z.; Wu, C.C.; Yaksh, T.L.; Di Nardo, A. Botulinum toxin blocks mast cells and prevents rosacea like inflammation. J. Dermatol. Sci. 2019, 93, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.J.; Hao, D.; Wen, X.; Du, D.; He, G.; Jiang, X. The Theranostics Role of Mast Cells in the Pathophysiology of Rosacea. Front. Med. 2019, 6, 324. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Sha, K.; Liu, Y.; Chen, M.; Xu, S.; Xie, H.; Deng, Z.; Li, J. mTORC1-Mediated Angiogenesis is Required for the Development of Rosacea. Front. Cell Dev. Biol. 2021, 9, 751785. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, H.; Deng, Z.; Wang, B.; Tang, Y.; Zhao, Z.; Yuan, X.; Zuo, Z.; Xu, S.; Zhang, Y.; et al. Tranexamic acid ameliorates rosacea symptoms through regulating immune response and angiogenesis. Int. Immunopharmacol. 2019, 67, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Lanier, V.B.; Braziel, R.M.; Falkenhagen, K.M.; White, C.; Rosenbaum, J.T. Expression of vascular endothelial growth factor and its receptors in rosacea. Br. J. Ophthalmol. 2007, 91, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, Y.; Jeong, S.W.; Jeong, G.H.; Moon, G.T.; Kim, M. Inhibition of Hippo Signaling Improves Skin Lesions in a Rosacea-Like Mouse Model. Int. J. Mol. Sci. 2021, 22, 931. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, Y.H.; Kim, J.; Park, D.Y.; Bae, H.; Lee, D.H.; Kim, K.H.; Hong, S.P.; Jang, S.P.; Kubota, Y.; et al. YAP/TAZ regulates sprouting angiogenesis and vascular barrier maturation. J. Clin. Investig. 2017, 127, 3441–3461. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Freire Valls, A.; Schermann, G.; Shen, Y.; Moya, I.M.; Castro, L.; Urban, S.; Solecki, G.M.; Winkler, F.; Riedemann, L.; et al. YAP/TAZ Orchestrate VEGF Signaling during Developmental Angiogenesis. Dev. Cell 2017, 42, 462–478.e7. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Liu, C.; Liu, S.; Zhou, X.; Lu, J.; Li, M.; Zhu, L. Inhibition of JAK1/STAT3 pathway by 2-methoxyestradiol ameliorates psoriatic features in vitro and in an imiquimod-induced psoriasis-like mouse model. Eur. J. Pharmacol. 2022, 933, 175276. [Google Scholar] [CrossRef] [PubMed]
- Routt, E.T.; Levitt, J.O. Rebound erythema and burning sensation from a new topical brimonidine tartrate gel 0.33%. J. Am. Acad. Dermatol. 2014, 70, e37–e38. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.S.; Nadkarni, A.; Cardwell, L.A.; Alinia, H.; Feldman, S.R. Spotlight on brimonidine topical gel 0.33% for facial erythema of rosacea: Safety, efficacy, and patient acceptability. Patient Prefer. Adherence 2017, 11, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis Rosa Franco, G.; Smid, S.; Viegas, C. Phytocannabinoids: General Aspects and Pharmacological Potential in Neurodegenerative Diseases. Curr. Neuropharmacol. 2021, 19, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Verde, R.; Vaia, M.; Allara, M.; Iuvone, T.; Di Marzo, V. Anti-inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. J. Pharmacol. Exp. Ther. 2018, 365, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, J.H.; Kim, S.W.; Bang, C.H. Cannabidiol Alleviates Imiquimod-Induced Psoriasis by Inhibiting JAK2-STAT3 in a Mouse Model. Biomedicines 2024, 12, 2084. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.M.; Gomes, A.L.; Vilas Boas, I.; Marto, J.; Ribeiro, H.M. Cannabis-Based Products for the Treatment of Skin Inflammatory Diseases: A Timely Review. Pharmaceuticals 2022, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.C.; Jeong, G.H.; Bang, C.H.; Lee, J.H. Cannabichromene as a Novel Inhibitor of Th2 Cytokine and JAK/STAT Pathway Activation in Atopic Dermatitis Models. Int. J. Mol. Sci. 2024, 25, 13539. [Google Scholar] [CrossRef] [PubMed]
- Nachnani, R.; Raup-Konsavage, W.M.; Vrana, K.E. The Pharmacological Case for Cannabigerol. J. Pharmacol. Exp. Ther. 2021, 376, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Wroński, A.; Jarocka-Karpowicz, I.; Surażyński, A.; Gęgotek, A.; Zarkovic, N.; Skrzydlewska, E. Modulation of Redox and Inflammatory Signaling in Human Skin Cells Using Phytocannabinoids Applied after UVA Irradiation: In Vitro Studies. Cells 2024, 13, 965. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.; Fernandez, J.R.; Fitzgerald, C.; Rouzard, K.; Tamura, M.; Savile, C. In Vitro and Clinical Evaluation of Cannabigerol (CBG) Produced via Yeast Biosynthesis: A Cannabinoid with a Broad Range of Anti-Inflammatory and Skin Health-Boosting Properties. Molecules 2022, 27, 491. [Google Scholar] [CrossRef] [PubMed]
- Luz-Veiga, M.; Mendes, A.; Tavares-Valente, D.; Amorim, M.; Conde, A.; Pintado, M.E.; Moreira, H.R.; Azevedo-Silva, J.; Fernandes, J. Exploring Cannabidiol (CBD) and Cannabigerol (CBG) Safety Profile and Skincare Potential. Int. J. Mol. Sci. 2024, 25, 12224. [Google Scholar] [CrossRef] [PubMed]
- Muto, Y.; Wang, Z.; Vanderberghe, M.; Two, A.; Gallo, R.L.; Di Nardo, A. Mast cells are key mediators of cathelicidin-initiated skin inflammation in rosacea. J. Investig. Dermatol. 2014, 134, 2728–2736. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.A.; Buhren, B.A.; Steinhoff, M.; Homey, B. Rosacea: The cytokine and chemokine network. J. Investig. Dermatol. Symp. Proc. 2011, 15, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, L.; Song, D.; Zhang, L.; Wang, X.; Du, D.; Jiang, X. Signaling pathways and targeted therapy for rosacea. Front. Immunol. 2024, 15, 1367994. [Google Scholar] [CrossRef] [PubMed]
- Maia, J.; Fonseca, B.M.; Teixeira, N.; Correia-da-Silva, G. Unveiling the angiogenic effects of cannabinoids: Enhancers or inhibitors? Biochem. Pharmacol. 2023, 215, 115686. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.H.; Yaar, M.; Eyada, M.M.; Bhawan, J. Lymphangiogenesis and angiogenesis in non-phymatous rosacea. J. Cutan. Pathol. 2007, 34, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.A.; Obaid, Z.M.; Fouda, I. Topical Brimonidine Tartrate 0.33% Gel versus 577-nm Pro-yellow Laser for the Treatment of Post-acne Erythema: A Comparative Study. J. Clin. Aesthet. Dermatol. 2025, 18, 35–39. [Google Scholar] [PubMed]
- Benedetti, A.; Turco, C.; Gallo, E.; Daralioti, T.; Sacconi, A.; Pulito, C.; Donzelli, S.; Tito, C.; Dragonetti, M.; Perracchio, L.; et al. ID4-dependent secretion of VEGFA enhances the invasion capability of breast cancer cells and activates YAP/TAZ via integrin beta3-VEGFR2 interaction. Cell Death Dis. 2024, 15, 113. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Xian, Y.; Chen, X.L. YAP/TAZ Signaling Enhances Angiogenesis of Retinal Microvascular Endothelial Cells in a High-Glucose Environment. Curr. Eye Res. 2024, 49, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.H.; Lee, J.H. Dysregulated Hippo Signaling Pathway and YAP Activation in Atopic Dermatitis: Insights from Clinical and Animal Studies. Int. J. Mol. Sci. 2023, 24, 17322. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Wang, Y.; Chen, L.; Wang, L.; Qin, X.; Xu, J.; Li, L.; Tu, Y.; Zhou, T.; et al. YAP via interacting with STAT3 regulates VEGF-induced angiogenesis in human retinal microvascular endothelial cells. Exp. Cell Res. 2018, 373, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Qureshy, Z.; Johnson, D.E.; Grandis, J.R. Targeting the JAK/STAT pathway in solid tumors. J. Cancer Metastasis Treat. 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ma, Z.; Song, L.; Zhu, C.; Li, J.; Su, Z.; Liu, H. Expression of interleukin-17 in oral tongue squamous cell carcinoma and its effect on biological behavior. Sci. Rep. 2025, 15, 3195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, B.; Huang, Y.; Li, Y.; Yan, S.; Xie, H.; Zhang, Y.; Li, J. Multi-Transcriptomic Analysis and Experimental Validation Implicate a Central Role of STAT3 in Skin Barrier Dysfunction Induced Aggravation of Rosacea. J. Inflamm. Res. 2022, 15, 2141–2156. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, Y.; Zhang, Z.; Jiang, Y.; Wang, R.; Zhang, H.; Su, Y. POSTN Silencing Ameliorates LL37-Induced Rosacea and Inhibits the JAK2/STAT3 and NF-κB Pathways. FASEB J. 2025, 39, e70643. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhong, Y.; Kuang, X.; Zhang, Y.; Yang, L.; Cai, Y.; Wang, F.; He, F.; Xie, H.; Wang, B.; et al. Targeting the STAT3/IL-36G signaling pathway can be a promising approach to treat rosacea. J. Adv. Res. 2025, 71, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, B.; Hu, R.; Huang, C.; Huang, K. A Review on the Mechanisms and Clinical Advances of JAK Inhibitors in the Treatment of Rosacea. J. Cutan. Med. Surg. 2025. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wang, X.; Liu, Y.; Singhal, M.; Gürkaşlar, C.; Valls, A.F.; Lei, Y.; Hu, W.; Schermann, G.; Adler, H.; et al. STAT3-YAP/TAZ signaling in endothelial cells promotes tumor angiogenesis. Sci. Signal 2021, 14, eabj8393. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Hwang, I.; Lee, E.; Cho, H.J.; Ryu, J.H.; Kim, T.G.; Yu, J.W. Antimicrobial Peptide LL-37 Drives Rosacea-Like Skin Inflammation in an NLRP3-Dependent Manner. J. Investig. Dermatol. 2021, 141, 2885–2894.e2885. [Google Scholar] [CrossRef] [PubMed]
- Buhl, T.; Sulk, M.; Nowak, P.; Buddenkotte, J.; McDonald, I.; Aubert, J.; Carlavan, I.; Déret, S.; Reiniche, P.; Rivier, M.; et al. Molecular and Morphological Characterization of Inflammatory Infiltrate in Rosacea Reveals Activation of Th1/Th17 Pathways. J. Investig. Dermatol. 2015, 135, 2198–2208. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Furue, M. Interleukin-22 and keratinocytes; pathogenic implications in skin inflammation. Explor. Immunol. 2021, 1, 37–47. [Google Scholar]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; Man, X.Y.; Xuan, X.Y.; Huang, C.Z.; Shen, Y.; Lao, L.M. Tofacitinib for the treatment of erythematotelangiectatic and papulopustular rosacea: A retrospective case series. Dermatol. Ther. 2022, 35, e15848. [Google Scholar] [CrossRef] [PubMed]
- Erickson, S.; Heul, A.V.; Kim, B.S. New and emerging treatments for inflammatory itch. Ann. Allergy Asthma Immunol. 2021, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Trier, A.M.; Mack, M.R.; Fredman, A.; Tamari, M.; Ver Heul, A.M.; Zhao, Y.; Guo, C.J.; Avraham, O.; Ford, Z.K.; Oetjen, L.K.; et al. IL-33 signaling in sensory neurons promotes dry skin itch. J. Allergy Clin. Immunol. 2022, 149, 1473–1480.e1476. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, W.; Malhi, N.K.; Huang, J.; Li, Q.; Zhou, Z.; Wang, R.; Peng, J.; Yin, T.; Wang, H. Cannabigerol (CBG): A Comprehensive Review of Its Molecular Mechanisms and Therapeutic Potential. Molecules 2024, 29, 5417. [Google Scholar] [CrossRef] [PubMed]
- Krzyzewska, A.; Kloza, M.; Kozlowska, H. Comprehensive mini-review: Therapeutic potential of cannabigerol-focus on the cardiovascular system. Front. Pharmacol. 2025, 16, 1561385. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2007, 13, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, T.; Zhao, H.; Xiao, X.; Hu, X.; Wang, B.; Huang, Y.; Yin, Z.; Zhong, Y.; Li, Y.; et al. High-sensitive sensory neurons exacerbate rosacea-like dermatitis in mice by activating gammadelta T cells directly. Nat. Commun. 2024, 15, 7265. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Chen, M.; Liu, Y.; Xu, S.; Ouyang, Y.; Shi, W.; Jian, D.; Wang, B.; Liu, F.; Li, J.; et al. A positive feedback loop between mTORC1 and cathelicidin promotes skin inflammation in rosacea. EMBO Mol. Med. 2021, 13, e13560. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Boikess, R.S.; Schwartz, R.A.; Cohen, P.J. Dimethyl sulfoxide (DMSO): A solvent that may solve selected cutaneous clinical challenges. Arch. Dermatol. Res. 2023, 315, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.M.; Shimabuko, D.Y.; Sipert, C.R. Dimethyl sulfoxide affects the viability and mineralization activity of apical papilla cells in vitro. Braz. Dent. J. 2024, 35, e246054. [Google Scholar] [CrossRef] [PubMed]
- Coda, A.B.; Hata, T.; Miller, J.; Audish, D.; Kotol, P.; Two, A.; Shafiq, F.; Yamasaki, K.; Harper, J.C.; Del Rosso, J.Q.; et al. Cathelicidin, kallikrein 5, and serine protease activity is inhibited during treatment of rosacea with azelaic acid 15% gel. J. Am. Acad. Dermatol. 2013, 69, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Wienholtz, N.K.F.; Thyssen, J.P.; Christensen, C.E.; Thomsen, S.F.; Karmisholt, K.E.; Jemec, G.B.E.; Lomholt, H.B.; Heidenheim, M.; Simonsen, A.B.; Sand, C.; et al. Validity and reliability of the Rosacea Area and Severity Index: A novel scoring system for clinical assessment of rosacea severity. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence (5′-3′) | |
|---|---|---|
| Mmp9 | Forward | GCCCGGAACTCACACGACA |
| Reverse | TTGGAAACTCACACGCGAGAAG | |
| Tlr2 | Forward | CTCTTCAGCAAACGCTGTTCT |
| Reverse | GGCGTCTCCCTCTATTGTATTG | |
| Vegfa | Forward | TATTCAGCGGACTCACCAGC |
| Reverse | AACCAACCTCCTCAAACCGT | |
| Il1b | Forward | TGC CAC CTT TTG ACA GTG AT |
| Reverse | AGT GAT ACT GCC TGC CTG AA | |
| Il4 | Forward | TCTCGAATGTACCAGGAGCCATATC |
| Reverse | AGCACCTTGGAAGCCTACAGA | |
| Il6 | Forward | CCC CAA TTT CCA ATG CTC TCC |
| Reverse | AGG CAT AAC GCA CTA GGT TT | |
| Il13 | Forward | CTGCTACCTCACTGTAGCCT |
| Reverse | TATTTCATGGCTGAGGGCTG | |
| Il22 | Forward | TTCCGAGGAGTCAGTGCTAA |
| Reverse | GAGTTTGGTCAGGAAAGGCA | |
| Il31 | Forward | ATACAGCTGCCGTGTTTCAG |
| Reverse | AGCCATCTTATCACCCAAGAA | |
| Actb | Forward | TGTGATGGTGGGAATGGGTCAGAA |
| Reverse | TGTGGTGCCAGATCTTCTCCATGT |
| Assay | Antibody | Dilution | Cat No. | Source |
|---|---|---|---|---|
| WB | β-actin | 1:2500 | #3700 | Cell Signaling Technology®, Danvers, MA, USA |
| WB/IHC | CD31 | 1:1000/ 1:200 | #77699 | Cell Signaling Technology® |
| WB/IHC | VEGF | 1:1000/1:500 | AB416154 | Abcam, Cambridge, UK |
| WB/IHC | YAP | 1:500/1:100 | sc-376830 | Santa Cruz, Dallas, TX, USA |
| WB/IHC | TAZ | 1:500/1:50 | sc-518026 | Santa Cruz |
| WB/IHC | JAK1 | 1:400/1:100 | #3344 | Cell Signaling Technology® |
| WB/IHC | STAT3 | 1:1000/1:500 | #9139 | Cell Signaling Technology® |
| WB/IHC | p-STAT3 | 1:1000/1:200 | #9145 | Cell Signaling Technology® |
| IHC | Filaggrin | 1:500 | 905804 | BioLegend, San Diego, CA, USA |
| IHC | Vimentin | 1:500 | AB92547 | Abcam |
| IHC | α-SMA | 1:500 | 14-9760-82 | Invitrogen |
| WB | Goat Anti-Mouse IgG antibody (HRP) | 1:4000 | GRX213111-01 | GeneTex, Irvine, CA, USA |
| WB | Goat Anti-Rabbit IgG antibody (HRP) | 1:4000 | GRX213110-01 | GeneTex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Lee, J.H. Targeting Vascular and Inflammatory Crosstalk: Cannabigerol as a Dual-Pathway Modulator in Rosacea. Int. J. Mol. Sci. 2025, 26, 6840. https://doi.org/10.3390/ijms26146840
Kim S, Lee JH. Targeting Vascular and Inflammatory Crosstalk: Cannabigerol as a Dual-Pathway Modulator in Rosacea. International Journal of Molecular Sciences. 2025; 26(14):6840. https://doi.org/10.3390/ijms26146840
Chicago/Turabian StyleKim, Suji, and Ji Hyun Lee. 2025. "Targeting Vascular and Inflammatory Crosstalk: Cannabigerol as a Dual-Pathway Modulator in Rosacea" International Journal of Molecular Sciences 26, no. 14: 6840. https://doi.org/10.3390/ijms26146840
APA StyleKim, S., & Lee, J. H. (2025). Targeting Vascular and Inflammatory Crosstalk: Cannabigerol as a Dual-Pathway Modulator in Rosacea. International Journal of Molecular Sciences, 26(14), 6840. https://doi.org/10.3390/ijms26146840