
Multi- and Transgenerational Histological and Transcriptomic Outcomes of Developmental TCDD Exposure in Zebrafish (Danio rerio) Ovary

 and
and
Abstract
1. Introduction
2. Results and Discussion
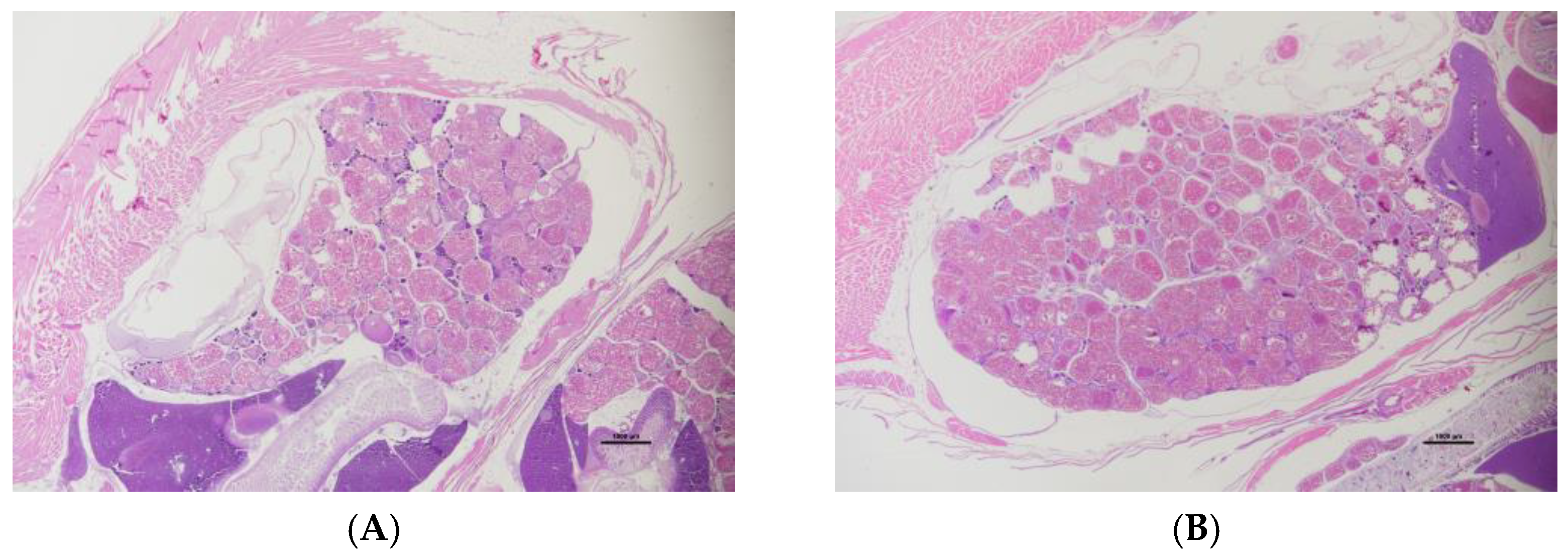
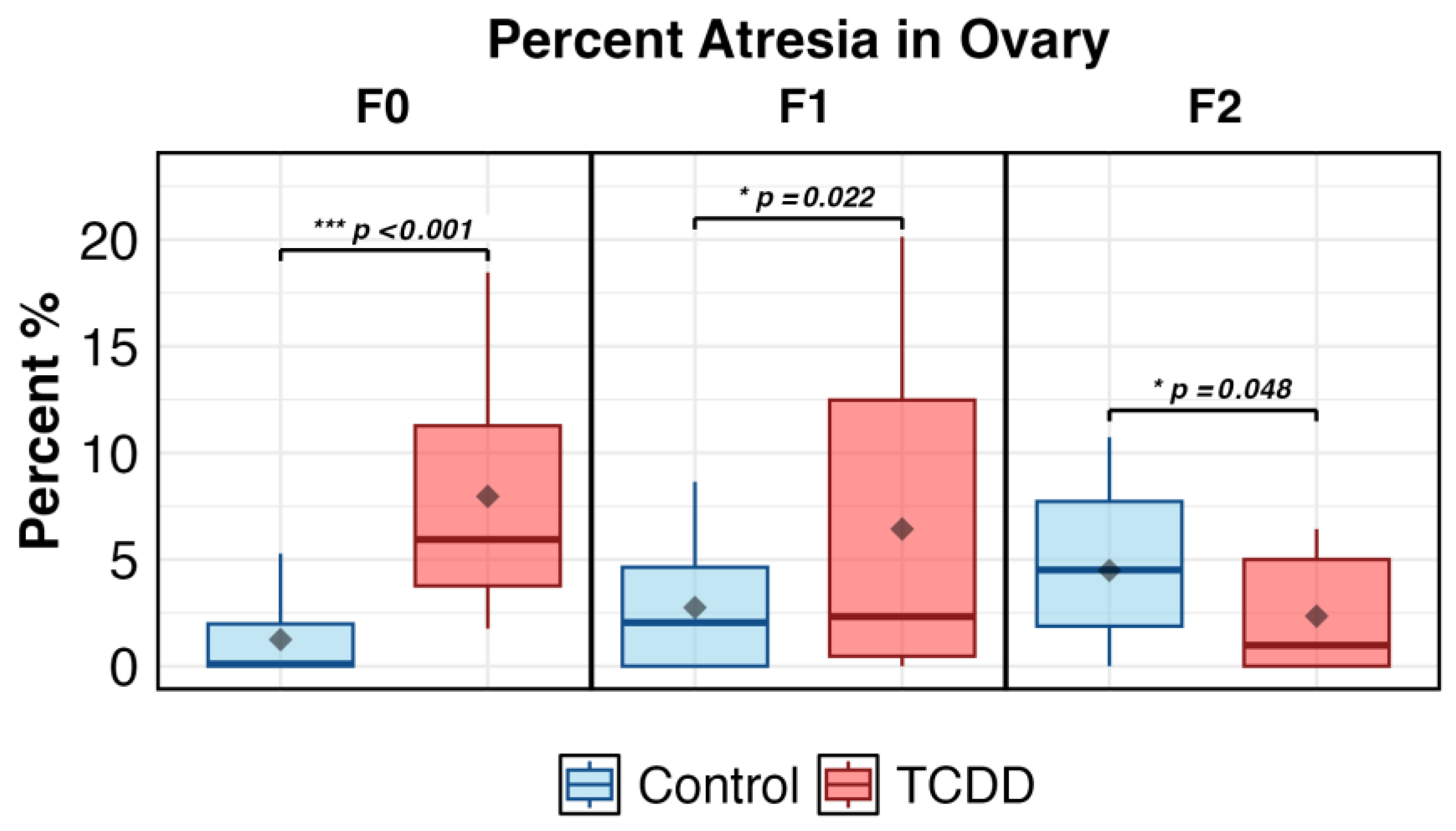
2.1. Ovarian Histology
2.2. Differentially Expressed Genes and Pathway Analysis
2.2.1. F0 Generation
2.2.2. F1 Generation
2.2.3. F2 Generation
2.2.4. Transgenerational Comparisons
2.3. Hormone Analysis
3. Materials and Methods
3.1. Fish Husbandry
3.2. Exposure
3.3. Ovarian Histology
3.4. Histopathological Endpoints
3.5. RNA Isolation
3.6. Differential Expression Analysis
3.7. Pathway Analysis
3.8. Hormone Analysis
3.8.1. Sample Homogenization
3.8.2. Vitellogenin ELISA Assay
3.8.3. Steroid/Thyroid Hormone Assay
3.8.4. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roseboom, T.J.; van der Meulen, J.H.P.; Ravelli, A.C.J.; Osmond, C.; Barker, D.J.P.; Bleker, O.P. Effects of prenatal exposure to the Dutch famine on adult disease in later life: An overview. Twin Res. 2001, 4, 293–298. [Google Scholar] [CrossRef]
- Zoeller, R.T.; Brown, T.R.; Doan, L.L.; Gore, A.C.; Skakkebaek, N.E.; Soto, A.M.; Woodruff, T.J.; Vom Saal, F.S. Endocrine-disrupting chemicals and public health protection: A statement of principles from The Endocrine Society. Endocrinology 2012, 153, 4097–4110. [Google Scholar] [CrossRef]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Dioxin (TCDD) induces epigenetic transgenerational inheritance of adult onset disease and sperm epimutations. PLoS ONE 2012, 7, e46249. [Google Scholar] [CrossRef]
- Skogen, J.C.; Øverland, S. The fetal origins of adult disease: A narrative review of the epidemiological literature. JRSM Short Rep. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Safe, S. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs), and related compounds: Environmental and mechanistic considerations which support the development of toxic equivalency factors (TEFs). Crit. Rev. Toxicol. 1990, 21, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Report on Human Biomonitoring of Environmental Chemicals in Pooled Samles: Results of the Canadian Health Measures Survey Cycles 1 (2007–2009), 3 (2012–2013), 4 (2014–2015) and 5 (2016–2017). Minister of Health. 2020. Available online: https://www.canada.ca/content/dam/hc-sc/images/services/environmental-workplace-health/reports-publications/environmental-contaminants/report-human-biomonitoring-environmental-chemicals-pooled-samples/27-20-2746-01-human-biomonitoring-final-en.pdf (accessed on 30 March 2025).
- Hofsteen, P.; Mehta, V.; Kim, M.-S.; Peterson, R.E.; Heideman, W. TCDD inhibits heart regeneration in adult zebrafish. Toxicol. Sci. 2012, 132, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.B.; Kerkvliet, N.I. Dioxin and immune regulation. Ann. N. Y. Acad. Sci. 2010, 1183, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.T.D.; Planchart, A.; Mattingly, C.J.; Winkler, C.; Reif, D.M.; Kullman, S.W. Embryonic exposure to tcdd impacts osteogenesis of the axial skeleton in Japanese medaka, Oryzias latipes. Toxicol. Sci. 2017, 155, 485–496. [Google Scholar] [CrossRef]
- Sha, R.; Chen, Y.; Wang, Y.; Luo, Y.; Liu, Y.; Ma, Y.; Li, Y.; Xu, L.; Xie, H.Q.; Zhao, B. Gestational and lactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin in mice: Neurobehavioral effects on female offspring. Sci. Total Environ. 2020, 752, 141784. [Google Scholar] [CrossRef]
- Mocarelli, P.; Gerthoux, P.M.; Patterson, D.G.; Milani, S.; Limonta, G.; Bertona, M.; Signorini, S.; Tramacere, P.; Colombo, L.; Crespi, C.; et al. Dioxin exposure, from infancy through puberty, produces endocrine disruption and affects human semen quality. Environ. Health Perspect. 2008, 116, 70–77. [Google Scholar] [CrossRef]
- Mocarelli, P.; Gerthoux, P.M.; Needham, L.L.; Patterson, D.G.; Limonta, G.; Falbo, R.; Signorini, S.; Bertona, M.; Crespi, C.; Sarto, C.; et al. Perinatal exposure to low doses of dioxin can permanently impair human semen quality. Environ. Health Perspect. 2011, 119, 713–718. [Google Scholar] [CrossRef]
- Eskenazi, B.; Warner, M.; Marks, A.R.; Samuels, S.; Needham, L.; Brambilla, P.; Mocarelli, P. Serum dioxin concentrations and time to pregnancy. Epidemiology 2010, 21, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Mocarelli, P.; Warner, M.; Samuels, S.; Vercellini, P.; Olive, D.; Needham, L.L.; Patterson, D.G.; Brambilla, P.; Gavoni, N.; et al. Serum dioxin concentrations and endometriosis: A cohort study in Seveso, Italy. Environ. Health Perspect. 2002, 110, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Beytur, A.; Ciftci, O.; Aydin, M.; Cakir, O.; Timurkaan, N.; Yılmaz, F. Protocatechuic acid prevents reproductive damage caused by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in male rats. Andrologia 2011, 44, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Foster, W.G.; Maharaj-Briceño, S.; Cyr, D.G. Dioxin-induced changes in epididymal sperm count and spermatogenesis. Environ. Health Perspect. 2010, 118, 458–464. [Google Scholar] [CrossRef]
- Mai, X.-M.; Dong, Y.; Xiang, L.; Er, Z. Matern exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin suppresses male reproductive functions in their adulthood. Hum. Exp. Toxicol. 2020, 39, 890–905. [Google Scholar] [CrossRef]
- Gray, L.E.; Ostby, J.S. In utero 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) alters reproductive morphology and function in female rat offspring. Toxicol. Appl. Pharmacol. 1995, 133, 285–294. [Google Scholar] [CrossRef]
- Myllymäki, S.A.; Haavisto, T.E.; Brokken, S.; Viluksela, M.; Toppari, J.; Paranko, J. In utero and lactational exposure to TCDD; steroidogenic outcomes differ in male and female rat pups. Toxicol. Sci. 2005, 88, 534–544. [Google Scholar] [CrossRef]
- Nynca, A.; Sadowska, A.; Paukszto, L.; Molcan, T.; Ruszkowska, M.; Swigonska, S.; Orlowska, K.; Myszczynski, K.; Jastrzebski, J.P.; Ciereszko, R.E. Temporal changes in the transcriptomic profile of granulosa cells of pigs treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Anim. Reprod. Sci. 2019, 207, 83–94. [Google Scholar] [CrossRef]
- Yu, K.; Zhang, X.; Tan, X.; Ji, M.; Chen, Y.; Tao, Y.; Liu, M.; Yu, Z. Transgenerational impairment of ovarian induced by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) associated with Igf2 and H19 in adult female rat. Toxicology 2019, 428, 152311. [Google Scholar] [CrossRef]
- Tsutsumi, O.; Uechi, H.; Sone, H.; Yonemoto, J.; Takai, Y.; Momoeda, M.; Tohyama, C.; Hashimoto, S.; Morita, M.; Taketani, Y. Presence of dioxins in human follicular fluid: Their possible stage-specific action on the development of preimplantation mouse embryos. Biochem. Biophys. Res. Commun. 1998, 250, 498–501. [Google Scholar] [CrossRef]
- Ye, M.; Chen, Y. Zebrafish as an emerging model to study gonad development. Comput. Struct. Biotechnol. J. 2020, 18, 2373–2380. [Google Scholar] [CrossRef]
- Orban, L.; Sreenivasan, R.; Olsson, P.-E. Long and winding roads: Testis differentiation in zebrafish. Mol. Cell. Endocrinol. 2009, 312, 35–41. [Google Scholar] [CrossRef]
- Baker, T.R.; Peterson, R.E.; Heideman, W. Early dioxin exposure causes toxic effects in adult zebrafish. Toxicol. Sci. 2013, 135, 241–250. [Google Scholar] [CrossRef]
- Baker, T.R.; Peterson, R.E.; Heideman, W. Using zebrafish as a model system for studying the transgenerational effects of dioxin. Toxicol. Sci. 2014, 138, 403–411. [Google Scholar] [CrossRef]
- Baker, B.B.; Yee, J.S.; Meyer, D.N.; Yang, D.; Baker, T.R. Histological and transcriptomic changes in male zebrafish testes due to early life exposure to low level 2,3,7,8-tetrachlorodibenzo-p-dioxin. Zebrafish 2016, 13, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.N.; Baker, B.B.; Baker, T.R. Ancestral TCDD exposure induces multigenerational histologic and transcriptomic alterations in gonads of male zebrafish. Toxicol. Sci. 2018, 164, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Akemann, C.; Meyer, D.N.; Gurdziel, K.; Baker, T.R. Developmental dioxin exposure alters the methylome of adult male zebrafish gonads. Front. Genet. 2019, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Akemann, C.; Meyer, D.N.; Gurdziel, K.; Baker, T.R. TCDD-induced multi- and transgenerational changes in the methylome of male zebrafish gonads. Curr. Zool. 2020, 6, dvaa010. [Google Scholar] [CrossRef]
- Johnson, R.; Wolf, J.; Braunbeck, T. Guidance Document on the Diagnosis of Endocrine-Related Histopathology in Fish Gonads. Organisation for Economic Co-Operation and Development (OECD). 2010. Available online: https://www.oecd.org/en/publications/guidance-document-on-the-diagnosis-of-endocrine-related-histopathology-in-fish-gonads_8f7cf3b5-en.html (accessed on 4 March 2025).
- Aldeli, N.; Soukkarie, C.; Hanano, A. Transcriptional, hormonal and histological alterations in the ovaries of BALB/c mice exposed to TCDD in connection with multigenerational female infertility. Ecotoxicol. Environ. Saf. 2023, 258, 114990. [Google Scholar] [CrossRef]
- Baker, T.R.; King-Heiden, T.C.; Peterson, R.E.; Heideman, W. Dioxin induction of transgenerational inheritance of disease in zebrafish. Mol. Cell. Endocrinol. 2014, 398, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.R.; Peterson, R.E.; Heideman, W. Adverse effects in adulthood resulting from low-level dioxin exposure in juvenile zebrafish. Endocr. Disruptors 2014, 2, e28309. [Google Scholar] [CrossRef] [PubMed]
- Wannemacher, R.; Rebstock, A.; Kulzer, E.; Schrenk, D.; Bock, K.W. Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on reproduction and oogenesis in zebrafish (Brachydanio rerio). Chemosphere 1992, 24, 1361–1368. [Google Scholar] [CrossRef]
- Heiden, T.K.; Hutz, R.J.; Carvan, M.J. Accumulation, tissue distribution, and maternal transfer of dietary 2,3,7,8,-tetrachlorodibenzo-p-dioxin: Impacts on reproductive success of zebrafish. Toxicol. Sci. 2005, 87, 497–507. [Google Scholar] [CrossRef]
- King Heiden, T.C.; Spitsbergen, J.; Heideman, W.; Peterson, R.E. Persistent adverse effects on health and reproduction caused by exposure of zebrafish to 2,3,7,8-tetrachlorodibenzo-p-dioxin during early development and gonad differentiation. Toxicol. Sci. 2009, 109, 75–87. [Google Scholar] [CrossRef]
- King-Heiden, T.C.; Mehta, V.; Xiong, K.M.; Lanham, K.A.; Antkiewicz, D.S.; Ganser, A.; Heideman, W.; Peterson, R.E. Reproductive and developmental toxicity of dioxin in fish. Mol. Cell. Endocrinol. 2012, 354, 121–138. [Google Scholar] [CrossRef]
- Liu, Q.; Spitsbergen, J.M.; Cariou, R.; Huang, C.-Y.; Jiang, N.; Goetz, G.; Hutz, R.J.; Tonellato, P.J.; Carvan, M.J. Histopathologic alterations associated with global gene expression due to chronic dietary TCDD exposure in juvenile zebrafish. PLoS ONE 2014, 9, e100910. [Google Scholar] [CrossRef]
- Heimler, I.; Trewin, A.L.; Chaffin, C.L.; Rawlins, R.G.; Hutz, R.J. Modulation of ovarian follicle maturation and effects on apoptotic cell death in Holtzman rats exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD) in utero and lactationally. Reprod. Toxicol. 1998, 12, 69–73. [Google Scholar] [CrossRef]
- Karman, B.N.; Basavarajappa, M.S.; Craig, Z.R.; Flaws, J.A. 2,3,7,8-Tetrachlorodibenzo-p-dioxin activates the aryl hydrocarbon receptor and alters sex steroid hormone secretion without affecting growth of mouse antral follicles in vitro. Toxicol. Appl. Pharmacol. 2012, 261, 88–96. [Google Scholar] [CrossRef]
- Hernández-Ochoa, I.; Karman, B.N.; Flaws, J.A. The role of the aryl hydrocarbon receptor in the female reproductive system. Biochem. Pharmacol. 2009, 77, 547–559. [Google Scholar] [CrossRef]
- Hombach-Klonisch, S.; Pocar, P.; Kietz, S.; Klonisch, T. Molecular actions of polyhalogenated arylhydrocarbons (PAHs) in female reproduction. Curr. Med. Chem. 2005, 12, 599–616. [Google Scholar] [CrossRef]
- Soave, I.; Caserta, D.; Wenger, J.-M.; Dessole, S.; Perino, A.; Marci, R. Environment and Endometriosis: A toxic relationship. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1964–1972. [Google Scholar] [PubMed]
- Bruner-Tran, K.L.; Yeaman, G.R.; Crispens, M.A.; Igarashi, T.; Osteen, K.G. Dioxin may promote inflammation-related development of endometriosis. Fertil. Steril. 2008, 89, 1287–1298. [Google Scholar] [CrossRef]
- Birnbaum, L.S.; Cummings, A.M. Dioxins and endometriosis: A plausible hypothesis. Environ. Health Perspect. 2002, 110, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kotkamp, K.; Mössner, R.; Allen, A.; Onichtchouk, D.; Driever, W. A Pou5f1/Oct4 dependent Klf2a, Klf2b, and Klf17 regulatory sub-network contributes to EVL and ectoderm development during zebrafish embryogenesis. Dev. Biol. 2014, 385, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Rawnsley, D.R.; Goddard, L.M.; Pan, W.; Cao, X.-J.; Jakus, Z.; Zheng, H.; Yang, J.; Arthur, J.S.C.; Whitehead, K.J.; et al. The cerebral cavernous malformation pathway controls cardiac development via regulation of endocardial MEKK3 signaling and KLF expression. Dev. Cell 2015, 32, 168–180. [Google Scholar] [CrossRef]
- Alexander, D.L.; Ganem, L.G.; Fernandez-Salguero, P.; Gonzalez, F.; Jefcoate, C.R. Aryl-hydrocarbon receptor is an inhibitory regulator of lipid synthesis and of commitment to adipogenesis. J. Cell Sci. 1998, 111 Pt 22, 3311–3322. [Google Scholar] [CrossRef]
- Banerjee, S.S.; Feinberg, M.E.; Watanabe, M.; Gray, S.H.; Haspel, R.L.; Denkinger, D.J.; Kawahara, R.S.; Hauner, H.; Jain, M.K. The Krüppel-like factor KLF2 inhibits peroxisome proliferator-activated receptor-γ expression and adipogenesis. J. Biol. Chem. 2003, 278, 2581–2584. [Google Scholar] [CrossRef]
- Basu, P.; Morris, P.E.; Haar, J.L.; Wani, M.A.; Lingrel, J.B.; Gaensler, K.M.L.; Lloyd, J.A. KLF2 is essential for primitive erythropoiesis and regulates the human and murine embryonic β-like globin genes in vivo. Blood 2005, 106, 2566–2571. [Google Scholar] [CrossRef]
- Belair, C.D.; Peterson, R.E.; Heideman, W. Disruption of erythropoiesis by dioxin in the zebrafish. Dev. Dyn. 2001, 222, 581–594. [Google Scholar] [CrossRef]
- Bock, K.W. Aryl hydrocarbon receptor (AHR)-mediated inflammation and resolution: Non-genomic and genomic signaling. Biochem. Pharmacol. 2020, 182, 114220. [Google Scholar] [CrossRef]
- Jha, P.; Das, H. KLF2 in regulation of NF-κB-mediated immune cell function and inflammation. Int. J. Mol. Sci. 2017, 18, 2383. [Google Scholar] [CrossRef] [PubMed]
- Heiden, T.C.K.; Struble, C.A.; Rise, M.L.; Hessner, M.J.; Hutz, R.J.; Carvan, M.J. Molecular targets of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) within the zebrafish ovary: Insights into TCDD-induced endocrine disruption and reproductive toxicity. Reprod. Toxicol. 2008, 25, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Cintrón-Rivera, L.G.; Oulette, G.; Prakki, A.; Burns, N.M.; Patel, R.; Cyr, R.; Plavicki, J. Exposure to the persistent organic pollutant 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD, dioxin) disrupts development of the zebrafish inner ear. Aquat. Toxicol. 2023, 259, 106539. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.N.; Silva, I.; Vo, B.; Paquette, A.; Blount, J.R.; George, S.E.; Gonzalez, G.; Cavaneau, E.; Khalaf, A.; Petriv, A.-M.; et al. Juvenile exposure to low-level 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) alters behavior and longitudinal morphometrics in zebrafish and F1 offspring. J. Dev. Orig. Health Dis. 2024, 15, e22. [Google Scholar] [CrossRef]
- Marakulina, D.; Vorontsov, I.E.; Kulakovskiy, I.V.; Lennartsson, A.; Drabløs, F.; Medvedeva, Y. EpiFactors 2022: Expansion and enhancement of a curated database of human epigenetic factors and complexes. Nucleic Acids Res. 2022, 51, D564–D570. [Google Scholar] [CrossRef]
- Patrizi, B.; de Cumis, M.S. TCDD toxicity mediated by epigenetic mechanisms. Int. J. Mol. Sci. 2018, 19, 4101. [Google Scholar] [CrossRef]
- Good, S.; Yegorov, S.; Martijn, J.; Franck, J.; Bogerd, J. New insights into ligand-receptor pairing and coevolution of relaxin family peptides and their receptors in teleosts. Int. J. Evol. Biol. 2012, 2012, 310278. [Google Scholar] [CrossRef]
- Crespo, D.; Assis, L.H.C.; Zhang, Y.T.; Safian, D.; Furmanek, T.; Skaftnesmo, K.O.; Norberg, B.; Ge, W.; Choi, Y.-C.; den Broeder, M.J.; et al. Insulin-like 3 affects zebrafish spermatogenic cells directly and via Sertoli cells. Commun. Biol. 2021, 4, 204. [Google Scholar] [CrossRef]
- Ivell, R.; Anand-Ivell, R. Insulin-like peptide 3 (INSL3) is a major regulator of female reproductive physiology. Hum. Reprod. Update 2018, 24, 639–651. [Google Scholar] [CrossRef]
- Lefèvre, P.L.C.; Berger, R.G.; Ernest, S.R.; Gaertner, D.W.; Rawn, D.F.K.; Wade, M.G.; Robaire, B.; Hales, B.F. Exposure of female rats to an environmentally relevant mixture of brominated flame retardants targets the ovary, affecting folliculogenesis and steroidogenesis. Biol. Reprod. 2016, 94, 9. [Google Scholar] [CrossRef]
- Lim, H.N.; Hawkins, J.R. Genetic control of gonadal differentiation. Bailliere’s Clin. Endocrinol. Metab. 1998, 12, 1–16. [Google Scholar] [CrossRef]
- Ledig, S.; Schippert, C.; Strick, R.; Beckmann, M.W.; Oppelt, P.G.; Wieacker, P. Recurrent aberrations identified by array-CGH in patients with Mayer-Rokitansky-Küster-Hauser syndrome. Fertil. Steril. 2011, 95, 1589–1594. [Google Scholar] [CrossRef]
- Bernardini, L.; Gimelli, S.; Gervasini, C.; Carella, M.; Baban, A.; Frontino, G.; Barbano, G.; Divizia, M.T.; Fedele, L.; Novelli, A.; et al. Recurrent microdeletion at 17q12 as a cause of Mayer-Rokitansky-Kuster-Hauser (MRKH) syndrome: Two case reports. Orphanet J. Rare Dis. 2009, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Sandbacka, M.; Laivuori, H.; Freitas, É.; Halttunen, M.; Jokimaa, V.; Morin-Papunen, L.; Rosenberg, C.; Aittomäki, K. TBX6, LHX1 and copy number variations in the complex genetics of Müllerian aplasia. Orphanet J. Rare Dis. 2013, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Sanker, S.; MCecilia Cirio Vollmer, L.L.; Goldberg, N.D.; McDermott, L.A.; Hukriede, N.A.; Vogt, A. Development of high-content assays for kidney progenitor cell expansion in transgenic zebrafish. SLAS Discov. 2013, 18, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Diep, C.Q.; Ma, D.; Deo, R.C.; Holm, T.M.; Naylor, R.W.; Arora, N.; Wingert, R.A.; Bollig, F.; Djordjevic, G.; Lichman, B.; et al. Identification of adult nephron progenitors capable of kidney regeneration in zebrafish. Nature 2011, 470, 95–100. [Google Scholar] [CrossRef]
- Henry, T.R.; Spitsbergen, J.M.; Hornung, M.W.; Abnet, C.C.; Peterson, R.E. Early life stage toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in zebrafish (Danio rerio). Toxicol. Appl. Pharmacol. 1997, 142, 56–68. [Google Scholar] [CrossRef]
- Tovo-Rodrigues, L.; Rohde, L.A.; Menezes, A.M.B.; Polanczyk, G.V.; Kieling, C.; Genro, J.P.; Anselmi, L.; Hutz, M.H. DRD4 rare variants in attention-deficit/hyperactivity disorder (ADHD): Further evidence from a birth cohort study. PLoS ONE 2013, 8, e85164. [Google Scholar] [CrossRef]
- Ek, F.; Malo, M.; Andersson, M.Å.; Wedding, C.; Kronborg, J.; Svensson, P.; Waters, S.; Petersson, P.; Olsson, R. Behavioral analysis of dopaminergic activation in zebrafish and rats reveals similar phenotypes. ACS Chem. Neurosci. 2016, 7, 633–646. [Google Scholar] [CrossRef]
- Reimer, M.M.; Norris, A.; Ohnmacht, J.; Patani, R.; Zhong, Z.; Dias, T.B.; Kuscha, V.; Scott, A.L.; Chen, Y.-C.; Rozov, S.; et al. Dopamine from the Brain Promotes Spinal Motor Neuron Generation during Development and Adult Regeneration. Dev. Cell 2013, 25, 478–491. [Google Scholar] [CrossRef]
- Knecht, A.L.; Truong, L.; Simonich, M.T.; Tanguay, R.L. Developmental benzo[a]pyrene (B[a]P) exposure impacts larval behavior and impairs adult learning in zebrafish. Neurotoxicology Teratol. 2017, 59, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Tanida, T.; Tasaka, K.; Akahoshi, E.; Ishihara-Sugano, M.; Saito, M.; Kawata, S.; Danjo, M.; Tokumoto, J.; Mantani, Y.; Nagahara, D.; et al. Fetal exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin transactivates aryl hydrocarbon receptor-responsive element III in the tyrosine hydroxylase immunoreactive neurons of the mouse midbrain. J. Appl. Toxicol. 2013, 34, 117–126. [Google Scholar] [CrossRef] [PubMed]
- La Salle, S.; Palmer, K.; O’BRien, M.; Schimenti, J.C.; Eppig, J.; Handel, M.A. Spata22, a novel vertebrate-specific gene, is required for meiotic progress in mouse germ cells. Biol. Reprod. 2012, 86, 45. [Google Scholar] [CrossRef]
- Hays, E.; Majchrzak, N.; Daniel, V.; Ferguson, Z.; Brown, S.; Hathorne, K.; La Salle, S. Spermatogenesis associated 22 is required for DNA repair and synapsis of homologous chromosomes in mouse germ cells. Andrology 2017, 5, 299–312. [Google Scholar] [CrossRef]
- Yao, C.; Hou, D.; Ji, Z.; Pang, D.; Li, P.; Tian, R.; Zhang, Y.; Ou, N.; Bai, H.; Zhi, E.; et al. Bi-allelic SPATA22 variants cause premature ovarian insufficiency and nonobstructive azoospermia due to meiotic arrest. Clin. Genet. 2022, 101, 507–516. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Y.; Murtaza, G.; Zhou, J.; Jiao, Y.; Gong, C.; Hu, C.; Han, Q.; Zhang, H.; Zhang, Y.; et al. Whole-exome sequencing of consanguineous families with infertile men and women identifies homologous mutations in SPATA22 and MEIOB. Hum. Reprod. 2021, 36, 2793–2804. [Google Scholar] [CrossRef]
- Haimbaugh, A.; Wu, C.-C.; Akemann, C.; Meyer, D.N.; Connell, M.; Abdi, M.; Khalaf, A.; Johnson, D.; Baker, T.R. Multi- and transgenerational effects of developmental exposure to environmental levels of PFAS and PFAS mixture in zebrafish (Danio rerio). Toxics 2022, 10, 334. [Google Scholar] [CrossRef]
- King, S.E.; Schatz, N.A.; Babenko, O.; Yaroslav Ilnytskyy Kovalchuk, I.; Metz, S. Prenatal maternal stress in rats alters the epigenetic and transcriptomic landscape of the maternal-fetal interface across four generations. Commun. Biol. 2025, 8, 1–13. [Google Scholar] [CrossRef]
- Desantis, D.; Yang, Y.; Lai, K.P.; Wu, R.S.S.; Schunter, C. Sex-specific neurotoxicity and transgenerational effects of an emerging pollutant, tris(1,3-dichloro-2-propyl)phosphate (TDCIPP). Sci. Total Environ. 2024, 957, 177520. [Google Scholar] [CrossRef]
- Sample to Insight. IPA Training: Maximizing the Biological Interpretation of Gene, Transcript & Protein Expression Data with IPA. Available online: https://chhe.research.ncsu.edu/wordpress/wp-content/uploads/2015/10/IPA-Data-Analysis-training-slides-2016_04.pdf (accessed on 4 March 2025).
- Yoshioka, W.; Tohyama, C. Mechanisms of developmental toxicity of dioxins and related compounds. Int. J. Mol. Sci. 2019, 20, 617. [Google Scholar] [CrossRef]
- Piwarski, S.A.; Salisbury, T.B. The effects of environmental aryl hydrocarbon receptor ligands on signaling and cell metabolism in cancer. Biochem. Pharmacol. 2023, 216, 115771. [Google Scholar] [CrossRef]
- Libertini, M.; Hallin, M.; Khin Thway Noujaim, J.; Benson, C.; van Jones, R.L. Gynecological sarcomas: Molecular characteristics, behavior, and histology-driven therapy. Int. J. Surg. Pathol. 2020, 29, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Astapova, O.; Minor, B.M.N.; Hammes, S.R. Physiological and Pathological Androgen Actions in the Ovary. Endocrinology 2019, 160, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Craig, Z.R.; Wang, W.; Flaws, J.A. Endocrine-disrupting chemicals in ovarian function: Effects on steroidogenesis, metabolism and nuclear receptor signaling. Reproduction 2011, 142, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, K.; Walker, N.J.; Nyska, A.; Kissling, G.E.; Jokinen, M.P.; Brix, A.E.; Sells, D.M.; Wyde, M.E. Thyroid follicular lesions induced by oral treatment for 2 years with 2,3,7,8-tetrachlorodibenzo-p-dioxin and dioxin-like compounds in female Harlan Sprague-Dawley rats. Toxicol. Pathol. 2010, 38, 1037–1050. [Google Scholar] [CrossRef]
- Katarzyńska, D.; Hrabia, A.; Kowalik, K.; Sechman, A. Comparison of the in vitro effects of TCDD, PCB 126 and PCB 153 on thyroid-restricted gene expression and thyroid hormone secretion by the chicken thyroid gland. Environ. Toxicol. Pharmacol. 2015, 39, 496–503. [Google Scholar] [CrossRef]
- Chevrier, J.; Warner, M.; Gunier, R.B.; Brambilla, P.; Eskenazi, B.; Mocarelli, P. Serum dioxin concentrations and thyroid hormone levels in the Seveso Women’s Health Study. Am. J. Epidemiol. 2014, 180, 490–498. [Google Scholar] [CrossRef]
- Johnson, E.; Shorter, C.; Bestervelt, L.; Patterson, D.; Needham, L.; Piper, W.; Lucier, G.; Nolan, C. Serum hormone levels in humans with low serum concentrations of 2,3,7,8-TCDD. Toxicol. Ind. Health 2001, 17, 105–112. [Google Scholar] [CrossRef]
- Pavuk, M.; Schecter, A.; Akhtar, F.Z.; Michalek, J.E. Serum 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) levels and thyroid function in air force veterans of the Vietnam War. Ann. Epidemiol. 2003, 13, 335–343. [Google Scholar] [CrossRef]
- Pohjanvirta, R. Target tissue morphology and serum biochemistry following 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) exposure in a TCDD-susceptible and a TCDD-resistant rat strain*1. Fundam. Appl. Toxicol. 1989, 12, 698–712. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program. NTP Technical report on the toxicology and carcinogenesis studies of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) (CAS no. 1746-01-6) in female Harlan Sprague-Dawley rats (gavage studies). Natl. Toxicol. Program 2006, 521, 4–232. [Google Scholar]
- Kohn, M.C. Effects of TCDD on thyroid hormone homeostasis in the rat. Drug Chem. Toxicol. 2000, 23, 259–277. [Google Scholar] [CrossRef]
- Chen, X.; Ma, X.; Ma, S.-M.; Coenraads, P.J.; Zhang, C.; Liu, J.; Zhao, L.; Sun, M.; Tang, N. Proteomic analysis of the rat ovary following chronic low-dose exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). J. Toxicol. Environ. Health 2009, 72, 717–726. [Google Scholar] [CrossRef]
- Heiden, T.K.; Carvan, M.J.; Hutz, R.J. Inhibition of follicular development, vitellogenesis, and serum 17β-estradiol concentrations in zebrafish following chronic, sublethal dietary exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Sci. 2005, 90, 490–499. [Google Scholar] [CrossRef]
- Enan, E.; Moran, F.; VandeVoort, C.A.; Stewart, D.R.; Overstreet, J.W.; Lasley, B.L. Mechanism of toxic action of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in cultured human luteinized granulosa cells. Reprod. Toxicol. 1996, 10, 497–508. [Google Scholar] [CrossRef]
- Morán, F.M.; Conley, A.J.; Corbin, C.J.; Enan, E.; VandeVoort, C.; Overstreet, J.W.; Lasley, B.L. 2,3,7,8-tetrachlorodibenzo-p-dioxin decreases estradiol production without altering the enzyme activity of cytochrome P450 aromatase of human luteinized granulosa cells in vitro. Biol. Reprod. 2000, 62, 1102–1108. [Google Scholar] [CrossRef]
- Baldridge, M.G.; Marks, G.T.; Rawlins, R.G.; Hutz, R.J. Very low-dose (femtomolar) 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) disrupts steroidogenic enzyme mRNAs and steroid secretion by human luteinizing granulosa cells. Reprod. Toxicol. 2015, 52, 57–61. [Google Scholar] [CrossRef]
- Grochowalski, A.; Chrza̧szcz, R.; Pieklo, R.; Gregoraszczuk, E.L. Estrogenic and antiestrogenic effect of in vitro treatment of follicular cells with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Chemosphere 2001, 43, 823–827. [Google Scholar] [CrossRef]
- Son, D.S.; Ushinohama, K.; Gao, X.; Taylor, C.C.; Roby, K.F.; Rozman, K.K.; Terranova, P.F. 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) blocks ovulation by a direct action on the ovary without alteration of ovarian steroidogenesis: Lack of a direct effect on ovarian granulosa and thecal-interstitial cell steroidogenesis in vitro. Reprod. Toxicol. 1999, 13, 521–530. [Google Scholar] [CrossRef]
- Bugel, S.M.; White, L.A.; Cooper, K.R. Inhibition of vitellogenin gene induction by 2,3,7,8-tetrachlorodibenzo-p-dioxin is mediated by aryl hydrocarbon receptor 2 (AHR2) in zebrafish (Danio rerio). Aquat. Toxicol. 2013, 126, 1–8. [Google Scholar] [CrossRef]
- Smeets, J.M.W.; Rankouhi, T.R.; Komen, J.; Kaminski, J.P.; Giesy, J.P.; van den Berg, M. In vitro vitellogenin production by carp (Cyprinus carpio) hepatocytes as a screening method for determining (anti)estrogenic activity of xenobiotics. Toxicol. Appl. Pharmacol. 1999, 157, 68–76. [Google Scholar] [CrossRef]
- Kossack, M.E.; Manz, K.E.; Martin, N.R.; Pennell, K.D.; Plavicki, J. Environmentally relevant uptake, elimination, and metabolic changes following early embryonic exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin in zebrafish. Chemosphere 2023, 310, 136723. [Google Scholar] [CrossRef] [PubMed]
- Eini, F.; Kutenaei, M.A.; Foroutan, T.; Salehi, E. High levels of follicular fluid testosterone could impair oocyte developmental competency via affecting aryl hydrocarbon receptor pathway in PCOS patients. BMC Mol. Cell Biol. 2022, 23, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, H.; Tian, Z.; Sun, A.; Dong, Y.; Dong, T.; Hu, H. Effects of 11-ketotestosterone on development of the previtellogenic ovary in the sterlet, Acipenser ruthenus. Front. Endocrinol. 2020, 11, 115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ye, D.; Wang, H.; Wang, Y.; Hu, W.; Sun, Y. Zebrafish cyp11c1 knockout reveals the roles of 11-ketotestosterone and cortisol in sexual development and reproduction. Endocrinology 2020, 161, bqaa048. [Google Scholar] [CrossRef]
- Aluru, N.; Vijayan, M.M. Aryl hydrocarbon receptor activation impairs cortisol response to stress in rainbow trout by disrupting the rate-limiting steps in steroidogenesis. Endocrinology 2006, 147, 1895–1903. [Google Scholar] [CrossRef]
- Nesan, D.; Vijayan, M.M. Role of glucocorticoid in developmental programming: Evidence from zebrafish. Gen. Comp. Endocrinol. 2013, 181, 35–44. [Google Scholar] [CrossRef]
- Warner, M.; Rauch, S.; Ames, J.; Mocarelli, P.; Brambilla, P.; Signorini, S.; Eskenazi, B. Prenatal dioxin exposure and thyroid hormone levels in the Seveso second generation study. Environ. Res. 2020, 183, 109280. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingenuity Canonical Pathways | −log(p-Value) | z-Score |
|---|---|---|
| RHO GTPase cycle | 3.96 | 3.16 |
| Neutrophil degranulation | 4.45 | 2.71 |
| Molecular Mechanisms of Cancer | 1.88 | 2.53 |
| Post-translational protein phosphorylation | 4.72 | 2.45 |
| Regulation of Insulin-like Growth Factor (IGF) transport and uptake by IGFBPs | 4.36 | 2.45 |
| RHOGDI Signaling | 3.85 | −2.45 |
| Categories | Functions | Diseases or Functions Annotation | p-Value | Bias-Corrected z-Score |
|---|---|---|---|---|
| Cancer, Organismal Injury and Abnormalities | cancer | Cancer of cells | 5.33 × 10−11 | 2.154 |
| Cancer, Organismal Injury and Abnormalities, Respiratory Disease | lung tumor | Lung tumor | 2.32 × 10−14 | 2.021 |
| Inflammatory Disease, Inflammatory Response, Organismal Injury and Abnormalities, Respiratory Disease | inflammation | Inflammation of lung | 0.000121 | −2.068 |
| Cell Death and Survival, Organismal Injury and Abnormalities | apoptosis | Apoptosis of carcinoma cell lines | 6.21 × 10−9 | −2.14 |
| Cardiovascular Disease, Organismal Injury and Abnormalities | size | Size of infarct | 0.00000019 | −2.298 |
| Inflammatory Response, Organismal Injury and Abnormalities, Respiratory Disease | inflammation | Inflammation of respiratory system component | 0.000000239 | −2.326 |
| Ingenuity Canonical Pathways | −log(p-Value) | z-Score |
|---|---|---|
| RHO GTPase cycle | 5.14 | 4.47 |
| Neutrophil degranulation | 3.31 | 4.12 |
| Phagosome Formation | 2.24 | 3.44 |
| Integrin Signaling | 6.47 | 3.36 |
| Actin Cytoskeleton Signaling | 6.41 | 3.32 |
| Heme signaling | 3.29 | −2 |
| Mitochondrial Dysfunction | 2.87 | −2.31 |
| Granzyme A Signaling | 3.17 | −2.45 |
| RHOGDI Signaling | 6.97 | −2.53 |
| Categories | Functions | Diseases or Functions Annotation | p-Value | Bias-Corrected z-Score |
|---|---|---|---|---|
| Cellular Movement | cell movement | Cell movement | 2.73 × 10−23 | 2.30 |
| Cell-To-Cell Signaling and Interaction, Inflammatory Response | immune response | Immune response of leukocytes | 0.000000424 | 2.12 |
| Cell Morphology | sprouting | Sprouting | 1.37 × 10−14 | 2.11 |
| Cellular Movement | invasion | Invasion of cells | 2.41 × 10−19 | 2.09 |
| Cell-To-Cell Signaling and Interaction, Hematological System Development and Function | activation | Activation of myeloid cells | 5.64 × 10−10 | 2.08 |
| Cardiovascular Disease, Organismal Injury and Abnormalities | infarction | Infarction | 1.82 × 10−11 | −2.04 |
| Cell Death and Survival, Organismal Injury and Abnormalities | apoptosis | Apoptosis of tumor cell lines | 5.3 × 10−10 | −2.06 |
| Cancer, Organismal Injury and Abnormalities | incidence | Incidence of tumor | 2.03 × 10−26 | −2.09 |
| Cancer, Gastrointestinal Disease, Organismal Injury and Abnormalities | colon tumor | Colon tumor | 1.75 × 10−16 | −2.17 |
| Cancer, Gastrointestinal Disease, Organismal Injury and Abnormalities | colorectal neoplasia | Colorectal tumor | 1.32 × 10−17 | −2.21 |
| Zebrafish Gene ID | Human Ortholog | Log2FC |
|---|---|---|
| tet2 | TET2 | 1.86 |
| chd3 | CHD3 | 1.71 |
| sfmbt1 | SFMBT1 | 1.68 |
| celf3a | CELF3 | 1.63 |
| phip | PHIP | 1.61 |
| chd5 | CHD5 | 1.56 |
| ywhae2 | YWHAE | 1.56 |
| zgpat | ZGPAT | 1.43 |
| zmynd8 | ZMYND8 | 1.39 |
| foxp4 | FOXP4 | 1.32 |
| prdm11 | PRDM11 | 1.29 |
| top2b | TOP2B | 1.26 |
| srsf1b | SRSF1 | 1.14 |
| kmt2e | KMT2E | 1.13 |
| smarcad1b | SMARCAD1 | 1.11 |
| cbx6 | CBX6 | 1.10 |
| foxo1a | FOXO1 | 1.04 |
| srsf10b | SRSF10 | −1.11 |
| ubr7 | UBR7 | −1.19 |
| taf4a | TAF4 | −1.43 |
| dnmt3bb.1 | DNMT3B | −1.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paquette, A.; Cavaneau, E.; Haimbaugh, A.; Meyer, D.N.; Akemann, C.; Dennis, N.; Baker, T.R. Multi- and Transgenerational Histological and Transcriptomic Outcomes of Developmental TCDD Exposure in Zebrafish (Danio rerio) Ovary. Int. J. Mol. Sci. 2025, 26, 6839. https://doi.org/10.3390/ijms26146839
Paquette A, Cavaneau E, Haimbaugh A, Meyer DN, Akemann C, Dennis N, Baker TR. Multi- and Transgenerational Histological and Transcriptomic Outcomes of Developmental TCDD Exposure in Zebrafish (Danio rerio) Ovary. International Journal of Molecular Sciences. 2025; 26(14):6839. https://doi.org/10.3390/ijms26146839
Chicago/Turabian StylePaquette, Amelia, Emma Cavaneau, Alex Haimbaugh, Danielle N. Meyer, Camille Akemann, Nicole Dennis, and Tracie R. Baker. 2025. "Multi- and Transgenerational Histological and Transcriptomic Outcomes of Developmental TCDD Exposure in Zebrafish (Danio rerio) Ovary" International Journal of Molecular Sciences 26, no. 14: 6839. https://doi.org/10.3390/ijms26146839
APA StylePaquette, A., Cavaneau, E., Haimbaugh, A., Meyer, D. N., Akemann, C., Dennis, N., & Baker, T. R. (2025). Multi- and Transgenerational Histological and Transcriptomic Outcomes of Developmental TCDD Exposure in Zebrafish (Danio rerio) Ovary. International Journal of Molecular Sciences, 26(14), 6839. https://doi.org/10.3390/ijms26146839