
Mycogenic Silver Nanoparticles: Promising Antimicrobials with Fungistatic Properties

 , , , and
, , , and
Abstract
1. Introduction
2. Results
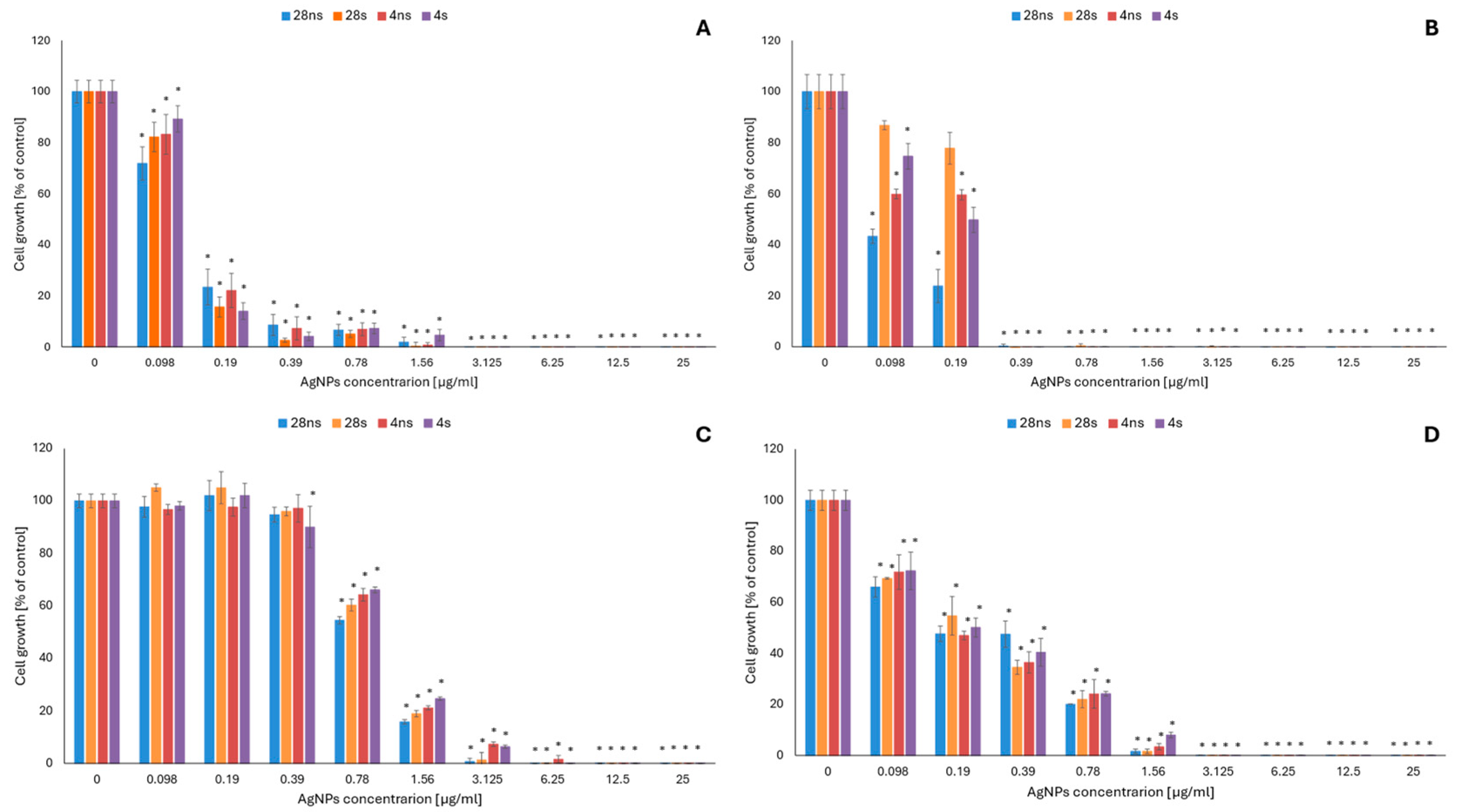
2.1. Evaluation of the Antifungal Activity of Mycogenic AgNPs
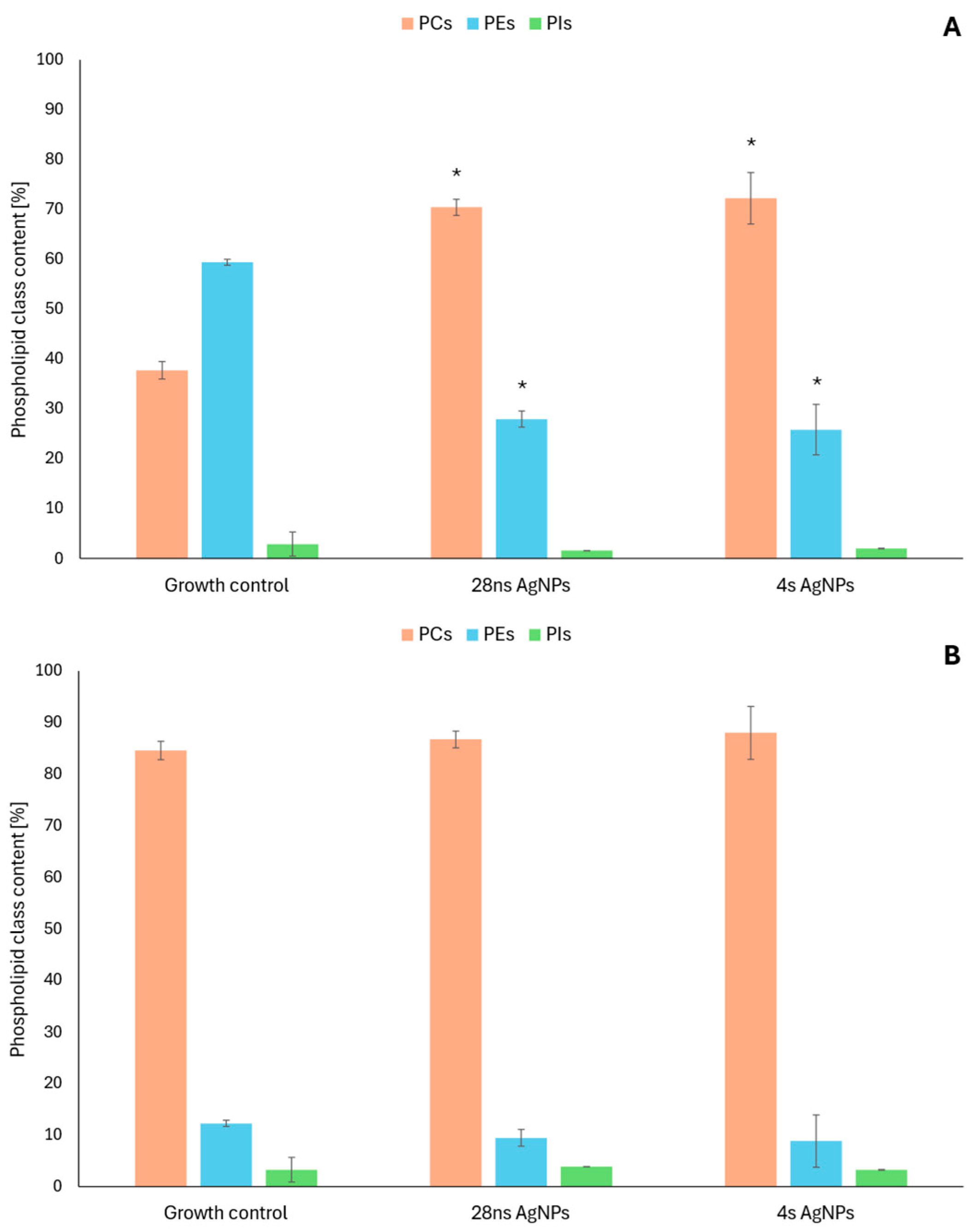
2.2. Changes in the Phospholipid Profiles of Fungal Cells in the Presence of Mycogenic AgNPs
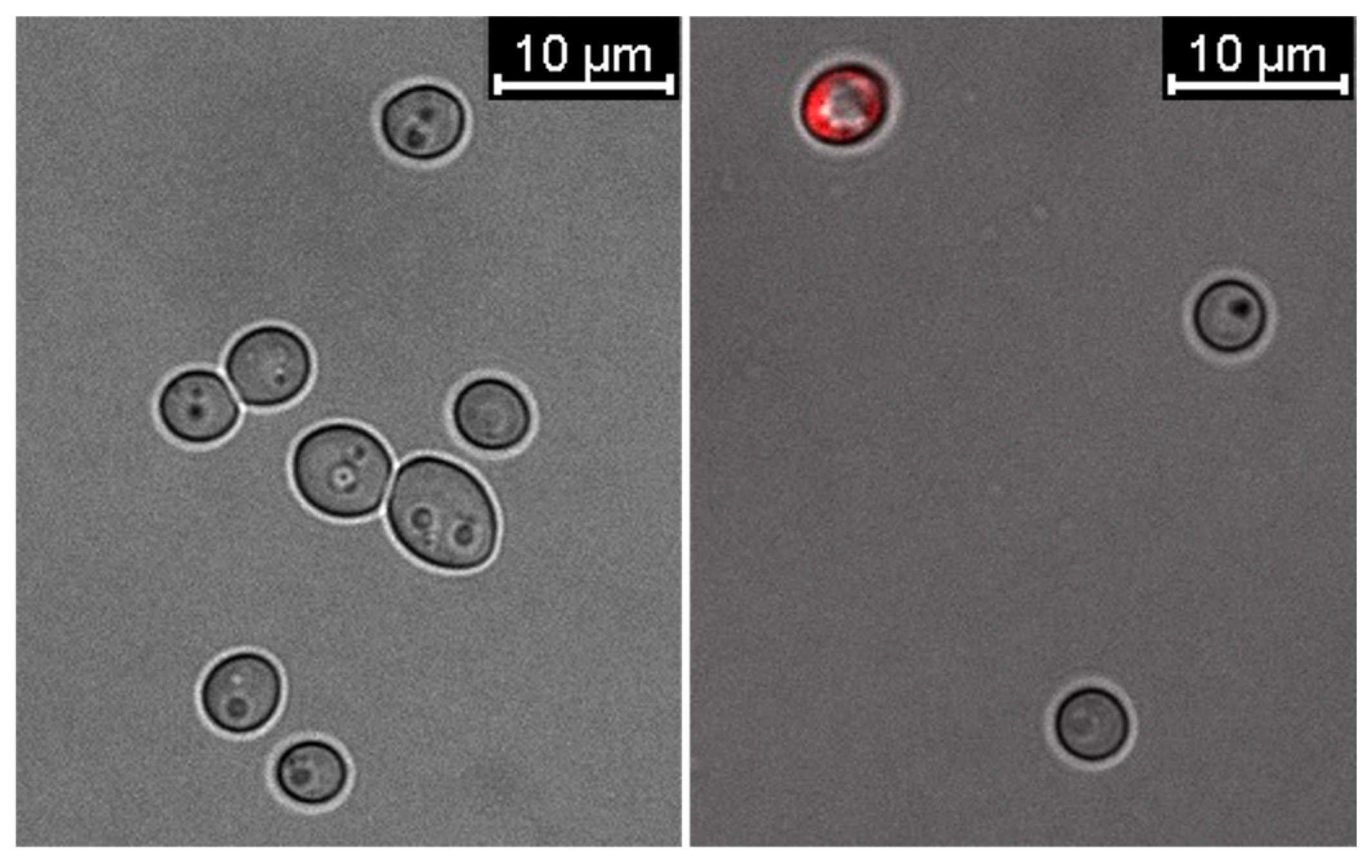
2.3. Changes in the Cell Membrane Properties of C. albicans in the Presence of Mycogenic AgNPs
2.3.1. Cell Membrane Fluidity
2.3.2. Cell Membrane Permeability
3. Discussion
4. Materials and Methods
4.1. Materials
AgNP Synthesis Using Gloeophyllum striatum DSM 9592
4.2. Methods
4.2.1. Evaluation of the Antifungal Activity of Mycogenic AgNPs
4.2.2. Changes in the Phospholipid Profiles of Fungal Cells in the Presence of Mycogenic AgNPs
4.2.3. Changes in C. albicans Cell Membrane Fluidity in the Presence of Mycogenic AgNPs
4.2.4. Confocal Microscopy Analysis
4.2.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dashora, A.; Rathore, K.; Raj, S.; Sharma, K. Synthesis of silver nanoparticles employing Polyalthia longifolia leaf extract and their in vitro antifungal activity against phytopathogen. Biochem. Biophys. Rep. 2022, 31, 101320. [Google Scholar] [CrossRef]
- Al-Otibi, F.; Perveen, K.; Al-Saif, N.A.; Alharbi, R.I.; Bokhari, N.A.; Albasher, G.; Al-Otaibi, R.M.; Al-Mosa, M.A. Biosynthesis of silver nanoparticles using Malva parviflora and their antifungal activity. Saudi J. Biol. Sci. 2021, 28, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.G.; Fernández-Baldo, M.A.; Berni, E.; Camí, G.; Durán, N.; Raba, J.; Sanz, M.I. Production of silver nanoparticles using yeasts and evaluation of their antifungal activity against phytopathogenic fungi. Process Biochem. 2016, 51, 1306–1313. [Google Scholar] [CrossRef]
- Barabadi, H.; Hosseini, O.; Jounaki, K.; Sadeghian-Abadi, S.; Ashouri, F.; Alrikabi, A.M.A.; Vahidi, H.; Amidi, S.; Mojab, F.; Mohammadi, N.; et al. Bioinspired green-synthesized silver nanoparticles: In vitro physicochemical, antibacterial, biofilm inhibitory, genotoxicity, antidiabetic, antioxidant, and anticoagulant performance. Mater. Adv. 2023, 4, 3037. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, L.; Rajoka, M.S.R.; Yan, L.; Jiang, C.; Shao, D.; Zhu, J.; Shi, J.; Huang, Q.; Yang, H.; et al. Fungal silver nanoparticles: Synthesis, application and challenges. Crit. Rev. Biotechnol. 2017, 38, 817–835. [Google Scholar] [CrossRef]
- Yassin, M.A.; Elgorban, A.M.; El-Samawaty, A.E.-R.M.; Almunqedhi, B.M. Biosynthesis of silver nanoparticles using Penicillium verrucosum and analysis of their antifungal activity. Saudi J. Biol. Sci. 2021, 28, 2123–2127. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Mali, S.C.; Sharma, S.; Trivedi, R. A review on biological synthesis of silver nanoparticles and their potential applications. Results Chem. 2023, 6, 101108. [Google Scholar] [CrossRef]
- Hoseini-Nilaki, S.F.; Ashengroph, M.; Zorab, M.M. Eco-friendly synthesis of silver nanoparticles using the fungus Alternaria sp. OP242500: Optimization through box-Behnken design. Results Chem. 2025, 15, 102265. [Google Scholar] [CrossRef]
- Malik, M.A.; Wani, A.H.; Bhat, M.Y.; Siddiqui SAlamri, S.A.M.; Alrumman, S.A. Fungal-mediated synthesis of silver nanoparticles: A novel strategy for plant disease management. Front. Microbiol. 2024, 15, 1399331. [Google Scholar] [CrossRef]
- El deeb, B.A.; Faheem, G.G.; Bakhit, M.S. Biosynthesis of silver nanoparticles by Talaromyces funiculosus for therapeutic applications and safety evaluation. Sci. Rep. 2025, 15, 13750. [Google Scholar] [CrossRef]
- Waszczuk, U.; Zapora, E.; Berezovska, D.; Stocki, M.; Wołkowycki, M.; Malewski, T.; Hsiang, T.; Oszako, T.; Borowik, P. Use of Secondary Metabolites of Wood-Decaying Fungi to Reduce Damping off Disease. Forests 2022, 13, 1208. [Google Scholar] [CrossRef]
- Kobashigawa, J.M.; Robles, C.A.; Ricci, M.L.M.; Carmarán, C.C. Influence of strong bases on the synthesis of silver nanoparticles (AgNPs) using the ligninolytic fungi Trametes trogii. Saudi J. Biol. Sci. 2019, 26, 1331–1337. [Google Scholar] [CrossRef]
- Osorio-Echevarría, J.; Osorio-Echevarría, J.; Ossa-Orozco, C.P.; Gomez-Vanegas, N.A. Synthesis of silver nanoparticles using white-rot fungus Anamorphous Bjerkandera sp. R1: Influence of silver nitrate concentration and fungus growth time. Sci. Rep. 2021, 11, 3842. [Google Scholar] [CrossRef]
- Gudikandula, K.; Vadapally, P.; Charya, M.A.S. Biogenic synthesis of silver nanoparticles from white rot fungi: Their characterization and antibacterial studies. OpenNano 2017, 2, 64–78. [Google Scholar] [CrossRef]
- Purnomo, A.S.; Mauliddawati, V.T.; Khoirudin, M.; Yonda, A.F.; Nawfa, R.; Putra, R. Bio-decolorization and novel bio-transformation of methyl orange by brown-rot fungi. Int. J. Environ. Sci. Technol. 2019, 16, 7555–7564. [Google Scholar] [CrossRef]
- Purnomo, A.S.; Sariwati, A.; Kamei, I. Synergistic interaction of a consortium of the brown-rot fungus Fomitopsis pinicola and bacterium Ralstonia pickettii for DDT biodegradation. Heliyon 2020, 6, e04027. [Google Scholar] [CrossRef]
- Oliveria, M.A.M.; Calado, M.L.; Guerreiro, S.F.C.; Dias, J.R.; Campos, M.J.; Murray, P.G.; Novais, S.C. Improved biosynthesis and characteristic of silver nanoparticles using marine endophytic fungi exposed to hypo-osmotic stress. Sci. Rep. 2025, 15, 16327. [Google Scholar] [CrossRef]
- Tończyk, A.; Niedziałkowska, K.; Lisowska, K. Optimizing the microbial synthesis of silver nanoparticles using Gloeophyllum striatum and their antimicrobial potential evaluation. Sci. Rep. 2023, 13, 21124. [Google Scholar] [CrossRef]
- Zawadzka, K.; Felczak, A.; Nowak, M.; Kowalczyk, A.; Piwoński, I.; Lisowska, K. Antimicrobial activity and toxicological risk assessment of silver nanoparticles synthesized using an eco-friendly method with Gloeophyllum striatum. J. Hazard. Mater. 2021, 418, 126316. [Google Scholar] [CrossRef]
- Wen, H.; Shi, H.; Jiang, N.; Qiu, J.; Lin, F.; Kou, Y. Antifungal mechanisms of silver nanoparticles on mycotoxin producing rice false smut fungus. iScience 2023, 26, 105763. [Google Scholar] [CrossRef]
- Ghojavand, S.; Madani, M.; Karimi, J. Green Synthesis, Characterization and Antifungal Activity of Silver Nanoparticles Using Stems and Flowers of Felty Germander. J. Inorg. Organomet. Polym. Mater. 2020, 30, 2987–2997. [Google Scholar] [CrossRef]
- Mussin, J.; Gussiano, G. Biogenic silver nanoparticles as antifungal agents. Front. Chem. 2022, 10, 1023542. [Google Scholar] [CrossRef] [PubMed]
- Rozhin, A.; Batasheva, S.; Iskuzhina, L.; Gomzikova, M.; Kryuchkova, M. Antimicrobial and Antifungal Action of Biogenic Silver Nanoparticles in Combination with Antibiotics and Fungicides Against Opportunistic Bacteria and Yeast. Int. J. Mol. Sci. 2024, 25, 12494. [Google Scholar] [CrossRef]
- Hossain, C.M.; Ryan, L.K.; Gera, M.; Choudhuri, S.; Lyle, N.; Ali, K.A.; Diamond, G. Antifungals and Drug Resistance. Encyclopedia 2022, 2, 1722–1737. [Google Scholar] [CrossRef]
- Garg, A.; Sharma, G.S.; Goyal, A.K.; Ghosh, G.; Si, S.C.; Rath, G. Recent advances in topical carriers of anti-fungal agents. Heliyon 2020, 6, e04663. [Google Scholar] [CrossRef]
- Jacobsen, I.D. The Role of Host and Fungal Factors in the Commensal-to-Pathogen Transition of Candida albicans. Curr. Clin. Microbiol. Rep. 2023, 10, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Yu, B.S.; Heo, Y.M.; Kyung, S.; Lee, K.-E.; Kim, S.; Kang, S.; Han, K.; Kim, D.H. Characteristic of Malassezia furfur at various pH and effects of Malassezia lipids on skin cells. Appl. Microb. Cell Physiol. 2024, 108, 455. [Google Scholar] [CrossRef] [PubMed]
- Petrucelli, R.; Cosio, T.; Camicia, V.; Fiorilla, C.; Gaziano, R.; D’Agostini, C. Malassezia furfur bloodstream infection: Still a diagnostic challenge in clinical practice. Med. Mycol. Case Rep. 2024, 45, 100657. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.; Xu, W.; Han, X.; Zhao, J. Primary Cutaneous Aspergillosis Due to Aspergillus fumigatus in an Immunocompetent Patient with Diabetes Mellitus After Tattooing: A Case Report and Revier of Literature. Infect. Drug Resist. 2023, 16, 791–797. [Google Scholar] [CrossRef]
- Merad, Y.; Derrar, H.; Belmokhtar, Z.; Belkacemi, M. Aspergillus Genus and Its Various Human Superficial and Cutaneous Features. Pathogens 2021, 10, 643. [Google Scholar] [CrossRef]
- Frippiat, T.; Art, T.; Delguste, C. Silver Nanoparticles as Antimicrobial Agents in Veterinary Medicine: Current Applications and Future Perspectives. Nanomaterials 2025, 15, 202. [Google Scholar] [CrossRef]
- Guliger-Casagrande, M.; de Lima, R. Synthesis of Silver Nanoparticles Mediated by Fungi: A Review. Front. Bioeng. Biotechnol. 2019, 7, 287. [Google Scholar] [CrossRef]
- Naveen, K.V.; Sathiyaseelan, A.; Mariadoss, A.V.A.; Xiaowen, H.; Saravanakumar, K.; Wang, M.-H. Fabrication of Mycogenic silver nanoparticles using endophytic fungal extract and their characterization, antibacterial and cytotoxicactivities. Inorg. Chem. Commun. 2021, 128, 108575. [Google Scholar] [CrossRef]
- Barabadi, H.; Mobaraki, K.; Jounaki, K.; Sadeghian-Abadi, S.; Vahidi, H.; Jahani, R.; Noqani, H.; Hosseini, O.; Ashouri, F.; Amidi, S. Exploring the biological application of Penicillium fimorum-derived silver nanoparticles: In vitro physicochemical, antifungal, biofilm inhibitory, antioxidant, anticoagulant, and thrombolytic performance. Heliyon 2023, 9, e16853. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, P.; Preethi, J.; Vijayan, R.; Yusoff, A.R.M.; Ameen, F.; Suresh, S.; Balagurunathan, R.; Palvannan, T. Anti-acne, anti-dandruff and anti-breast cancer efficacy of green synthesised silver nanoparticles using Coriandrum sativum leaf extract. J. Photochem. Photobiol. B Biol. 2016, 163, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.H.; Saied, E.; Amin, B.H.; Alotibi, F.O.; Al-Askar, A.A.; Arishi, A.A.; Elkady, F.M.; Elbahnasawy, M.A. Antifungal Activity of Biosynthesized Silver Nanoparticles (AgNPs) against Aspergilli Causing Aspergillosis: Ultrastructure Study. J. Funct. Biomater. 2022, 13, 242. [Google Scholar] [CrossRef] [PubMed]
- Lotfy, W.A.; Alkersh, B.M.; Sabry, S.A.; Ghozlan, H.A. Biosynthesis of Silver Nanoparticles by Aspergillus terreus: Characterization, Optimization, and Biological Activities. Front. Bioeng. Biotechnol. 2021, 9, 633468. [Google Scholar] [CrossRef]
- Ansari, M.A.; Kalam, A.; Al-Sehemi, A.G.; Alomary, M.N.; AlYahya, S.; Aziz, M.K.; Srivastava, S.; Alghamdi, S.; Akhtar, S.; Almalki, H.D.; et al. Counteraction of Biofilm Formation and Antimicrobial Potential of Terminalia catappa Functionalized Silver Nanoparticles against Candida albicans and Multidrug-Resistant Gram-Negative and Gram-Positive Bacteria. Antibiotics 2021, 10, 725. [Google Scholar] [CrossRef] [PubMed]
- Różalska, B.; Sadowska, B.; Budzyńska, A.; Bernat, P.; Różalska, S. Biogenic nanosilver synthesized in Metharizium robertsii waste mycelium extract–As a modulator of Candida albicans morphogenesis, membrane lipidome and biofilm. PLoS ONE 2018, 13, e0194254. [Google Scholar] [CrossRef]
- Mussin, J.E.; Roldan, M.V.; Rojas, F.; de los Angeles Sosa, M.; Pellegri, N.; Giusiano, G. Antifungal activity of silver nanoparticles in combination with ketoconazole against Malassezia furfur. AMB Express 2019, 9, 131. [Google Scholar] [CrossRef]
- Ramirez, A.M.C.; Amezquita, A.; Jaramillo, J.E.C.C.; Matiz-Ceron, L.F.; Andrande-Martinez, J.S.; Triana, S.; Mantilla, M.J.; Restrepo, S.; Barrios, A.F.G.; de Cock, H. Analysis of Malassezia Lipidome Disclosed Differences Among Species and Reveals Presence of Unusual Yeast Lipids. Front. Cell. Infect. Microbiol. 2020, 10, 338. [Google Scholar] [CrossRef]
- Matras, E.; Gorczyca, A.; Przemieniecki, S.W.; Oćwieja, M. Surface properties-dependent antifunga lactivity of silver nanoparticles. Sci. Rep. 2022, 12, 18046. [Google Scholar] [CrossRef] [PubMed]
- Prasher, P.; Singh, M.; Mudila, H. Green Synthesis of Silver Nanoparticles and their Antifungal Properties. BioNanoScience 2017, 8, 254–263. [Google Scholar] [CrossRef]
- Felczak, A.; Zawadzka, K.; Bernat, P.; Nowak-Lange, M.; Lisowska, K. Effect of Quinoline on the Phospholipid Profile of Curvularia lunata and Its Microbial Detoxification. Molecules 2022, 27, 2081. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Zhang, C.; Wu, T.; Ma, Z.; Chen, Y. Phospholipid homeostasis plays an important role in fungal development, fungicide resistance and virulence in Fusarium graminearum. Phytopathol. Res. 2019, 1, 16. [Google Scholar] [CrossRef]
- Ball, W.B.; Neff, J.K.; Gohil, V.M. The role of nonbilayer phospholipids in mitochondrial structure and function. FEBS Lett. 2017, 592, 1273–1290. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Huang, G.; Feng, F.; Liu, X.; Guo, R.; Gu, F.; Zhong, X.; Mei, X. Dynamic changes in membrane lipid composition of leaves of winter wheat seedlings in response to PEG-induced water stress. BMC Plant Biol. 2020, 20, 84. [Google Scholar] [CrossRef]
- Yu, L.; Zhou, C.; Fan, J.; Shanklin, J.; Xu, C. Mechanisms and functions of membrane lipid remodeling in plants. Plant J. 2021, 107, 37–53. [Google Scholar] [CrossRef]
- Gao, B.; Yu, B.; Huang, X.; Li, H.; Jia, Y.; Wang, M.; Lu, Y.; Zhang, X.; Li, W. Cadmium and calcium ions’ effects on the growth of Pleurotus straceus mycelia are related to phosphatidylethanolamine content. Fungal Biol. 2024, 128, 2190–2196. [Google Scholar] [CrossRef]
- Choy, H.L.; Gaylord, E.A.; Doering, T.L. Ergosterol distribution controls surface structure formation and fungal pathogenicity. mBio 2023, 14, e0135323. [Google Scholar] [CrossRef]
- Bernat, P.; Jasińska, A.; Niedziałkowska, K.; Słaba, M.; Różalska, S.; Paraszkiewicz, K.; Sas-Paszt, L.; Heipieper, H.J. Adaptation of the metolachlor-degrading fungus Trichoderma harzianum to the simultaneous presence of low-density polyethylene (LDPE) microplastics. Ecotoxicol. Environ. Saf. 2023, 267, 115656. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC | MFC | |||||||
|---|---|---|---|---|---|---|---|---|
| 28 ns | 28 s | 4 ns | 4 s | 28 ns | 28 s | 4 ns | 4 s | |
| A. fumigatus ATCC 204305 | 1.56 | 1.56 | 3.125 | 3.125 | >25 | >25 | >25 | >25 |
| A. flavus ATCC 9643 | 3.125 | 3.125 | 6.25 | 12.5 | >25 | >25 | >25 | >25 |
| C. albicans ATCC 10231 | 1.56 | 1.56 | 1.56 | 3.125 | >25 | >25 | >25 | 25 |
| M. furfur DSM 6170 | 0.39 | 0.39 | 0.39 | 0.39 | >25 | >25 | >25 | >25 |
| A. flavus ATCC 9643 | C. albicans ATCC 10231 | |||||
|---|---|---|---|---|---|---|
| Growth Control | 28 ns AgNPs | 4 s AgNPs | Growth Control | 28 ns AgNPs | 4 s AgNPs | |
| PC/PE | 0.64 | 2.52 | 2.87 | 6.92 | 9.24 | 10.09 |
| DBI | 0.95 | 1.20 | 1.17 | 1.08 | 1.10 | 1.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tończyk, A.; Niedziałkowska, K.; Nowak-Lange, M.; Bernat, P.; Lisowska, K. Mycogenic Silver Nanoparticles: Promising Antimicrobials with Fungistatic Properties. Int. J. Mol. Sci. 2025, 26, 6639. https://doi.org/10.3390/ijms26146639
Tończyk A, Niedziałkowska K, Nowak-Lange M, Bernat P, Lisowska K. Mycogenic Silver Nanoparticles: Promising Antimicrobials with Fungistatic Properties. International Journal of Molecular Sciences. 2025; 26(14):6639. https://doi.org/10.3390/ijms26146639
Chicago/Turabian StyleTończyk, Aleksandra, Katarzyna Niedziałkowska, Marta Nowak-Lange, Przemysław Bernat, and Katarzyna Lisowska. 2025. "Mycogenic Silver Nanoparticles: Promising Antimicrobials with Fungistatic Properties" International Journal of Molecular Sciences 26, no. 14: 6639. https://doi.org/10.3390/ijms26146639
APA StyleTończyk, A., Niedziałkowska, K., Nowak-Lange, M., Bernat, P., & Lisowska, K. (2025). Mycogenic Silver Nanoparticles: Promising Antimicrobials with Fungistatic Properties. International Journal of Molecular Sciences, 26(14), 6639. https://doi.org/10.3390/ijms26146639