
How Structural Variations Influence Crop Improvement

Abstract
1. Introduction
2. The Types and Detection of Structural Variations
3. The Naturally Occurring Causes of Structural Variations in Plants
4. The Impacts of Structural Variations
4.1. Crop Growth and Development
4.2. Crop Yield
4.3. Crop Quality
4.4. Crop Stress Resistance
4.5. Domestication and Evolution
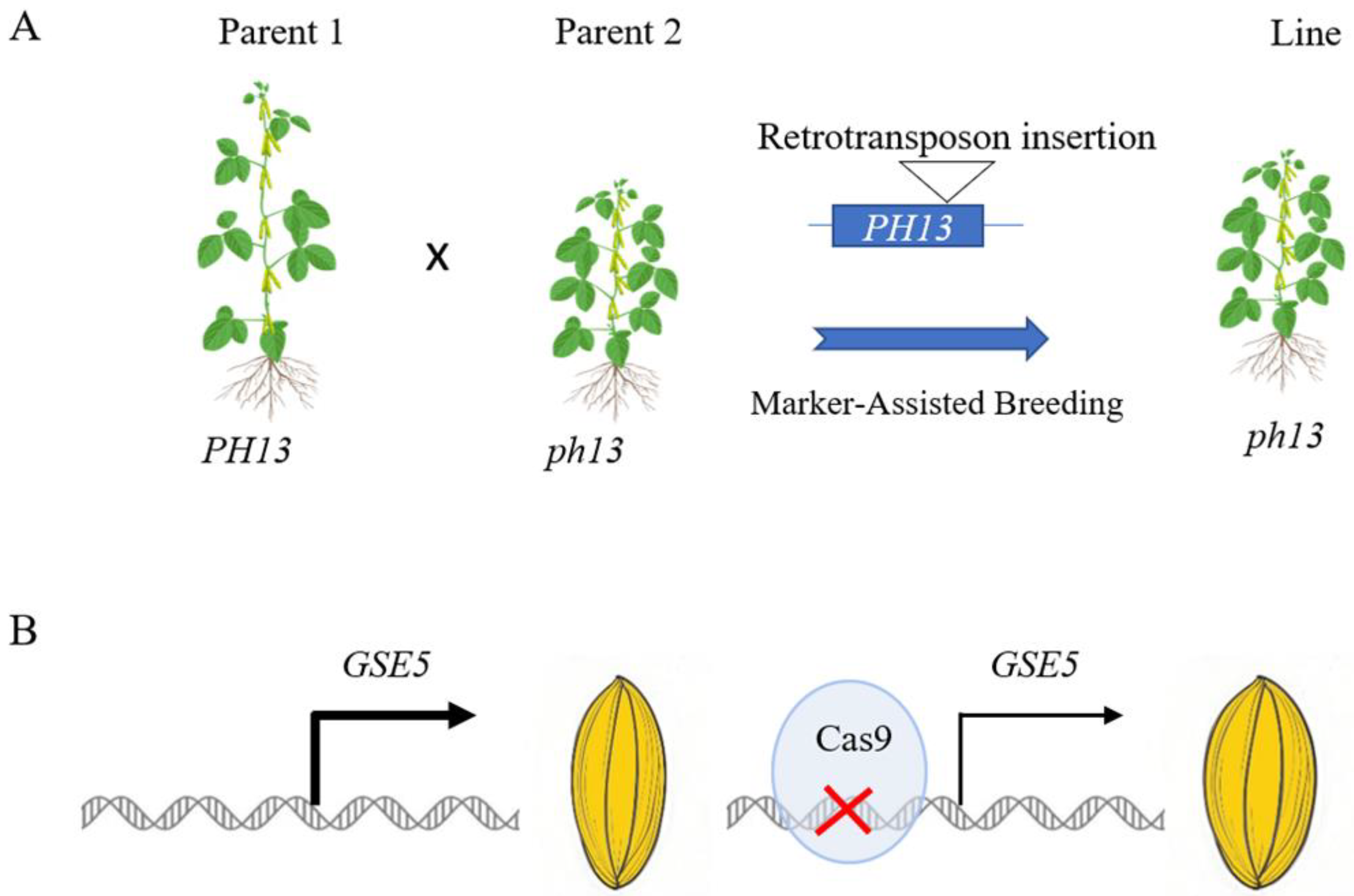
5. How to Apply Structural Variations to Crop Breeding
5.1. Molecular Marker-Assisted Breeding
5.2. Gene Editing Technology
6. Opportunities and Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Shi, J.; Lai, J. Patterns of genomic changes with crop domestication and breeding. Curr. Opin. Plant Biol. 2015, 24, 47–53. [Google Scholar] [CrossRef]
- Huang, X.; Huang, S.; Han, B.; Li, J. The integrated genomics of crop domestication and breeding. Cell 2022, 185, 2828–2839. [Google Scholar] [CrossRef] [PubMed]
- Escaramís, G.; Docampo, E.; Rabionet, R. A decade of structural variants: Description, history and methods to detect structural variation. Brief Funct. Genom. 2015, 14, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Feuk, L.; Carson, A.; Scherer, S. Structural variation in the human genome. Nat. Rev. Genet. 2006, 7, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Scott, A.J.; Davis, J.R.; Tsang, E.K.; Li, X.; Kim, Y.; Hadzic, T.; Damani, F.N.; Ganel, L.; Montgomery, S.B.; et al. The impact of structural variation on human gene expression. Nat. Genet. 2017, 49, 692–699. [Google Scholar] [CrossRef]
- Meyers, L.A.; Levin, D.A. On the abundance of polyploids in flowering plants. Evolution 2006, 60, 1198–1206. [Google Scholar]
- Yuan, Y.; Bayer, P.E.; Batley, J.; Edwards, D. Current status of structural variation studies in plants. Plant Biotechnol. J. 2021, 19, 2153–2163. [Google Scholar] [CrossRef]
- Rhoads, A.; Au, K.F. PacBio sequencing and its applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef]
- Jain, M.; Koren, S.; Miga, K.H.; Quick, J.; Rand, A.C.; Sasani, T.A.; Tyson, J.R.; Beggs, A.D.; Dilthey, A.T.; Fiddes, I.T.; et al. Nanopore sequencing and assembly of a human genome with ultra-long reads. Nat. Biotechnol. 2018, 36, 338–345. [Google Scholar] [CrossRef]
- Schiessl, S.V.; Katche, E.; Ihien, E.; Chawla, H.S.; Mason, A.S. The role of genomic structural variation in the genetic improvement of polyploid crops. Crop J. 2019, 7, 127–140. [Google Scholar] [CrossRef]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Chia, J.M.; Song, C.; Bradbury, P.J.; Costich, D.; de Leon, N.; Doebley, J.; Elshire, R.J.; Gaut, B.; Geller, L.; Glaubitz, J.C.; et al. Maize HapMap2 identifies extant variation from a genome in flux. Nat. Genet. 2012, 44, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Iovene, M.; Zhang, T.; Lou, Q.; Buell, C.R.; Jiang, J. Copy number variation in potato an asexually propagated autotetraploid species. Plant J. 2013, 75, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Zarrei, M.; MacDonald, J.R.; Merico, D.; Scherer, S.W. A copy number variation map of the human genome. Nat. Rev. Genet. 2015, 16, 172–183. [Google Scholar] [CrossRef]
- Solinas-Toldo, S.; Lampel, S.; Stilgenbauer, S.; Nickolenko, J.; Benner, A.; Döhner, H.; Cremer, T.; Lichter, P. Matrix-based comparative genomic hybridization: Biochips to screen for genomic imbalances. Genes Chromosomes Cancer 1997, 20, 399–407. [Google Scholar] [CrossRef]
- Pinkel, D.; Segraves, R.; Sudar, D.; Clark, S.; Poole, I.; Kowbel, D.; Collins, C.; Kuo, W.L.; Chen, C.; Zhai, Y.; et al. High resolution analysis of DNA copy number variation using comparative genomic hybridization to microarrays. Nat. Genet. 1998, 20, 207–211. [Google Scholar] [CrossRef]
- Miller, D.T.; Adam, M.P.; Aradhya, S.; Biesecker, L.G.; Brothman, A.R.; Carter, N.P.; Church, D.M.; Crolla, J.A.; Eichler, E.E.; Epstein, C.J.; et al. Consensus statement: Chromosomal microarray is a first-tier clinical diagnostic test for individuals with developmental disabilities or congenital anomalies. Am. J. Hum. Genet. 2010, 86, 749–764. [Google Scholar] [CrossRef]
- Alkan, C.; Coe, B.P.; Eichler, E.E. Genome structural variation discovery and genotyping. Nat. Rev. Genet. 2011, 12, 363–376. [Google Scholar] [CrossRef]
- Harewood, L.; Kishore, K.; Eldridge, M.D.; Wingett, S.; Pearson, D.; Schoenfelder, S.; Collins, V.P.; Fraser, P. Hi-C as a tool for precise detection and characterisation of chromosomal rearrangements and copy number variation in human tumours. Genome Biology 2017, 18, 125. [Google Scholar] [CrossRef]
- Huddleston, J.; Chaisson, M.J.P.; Steinberg, K.M.; Warren, W.; Hoekzema, K.; Gordon, D.; Graves-Lindsay, T.A.; Munson, K.M.; Kronenberg, Z.N.; Vives, L.; et al. Corrigendum: Discovery and genotyping of structural variation from long-read haploid genome sequence data. Genome Res. 2018, 28, 144. [Google Scholar] [CrossRef]
- Sedlazeck, F.J.; Rescheneder, P.; Smolka, M.; Fang, H.; Nattestad, M.; von Haeseler, A.; Schatz, M.C. Accurate detection of complex structural variations using single-molecule sequencing. Nat. Methods 2018, 15, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Heller, D.; Vingron, M. SVIM: Structural variant identification using mapped long reads. Bioinformatics 2019, 35, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhao, X.; Mace, E.; Henry, R.; Jordan, D. Exploring and exploiting pan-genomics for crop improvement. Mol. Plant 2019, 12, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Della Coletta, R.; Qiu, Y.; Ou, S.; Hufford, M.B.; Hirsch, C.N. How the pan-genome is changing crop genomics and improvement. Genome Biol. 2021, 22, 3. [Google Scholar] [CrossRef]
- Wei, C.; Gao, L.; Xiao, R.; Wang, Y.; Chen, B.; Zou, W.; Li, J.; Mace, E.; Jordan, D.; Tao, Y. Complete telomere-to-telomere assemblies of two sorghum genomes to guide biological discovery. Imeta 2024, 3, e193. [Google Scholar] [CrossRef]
- Jiao, C.; Xie, X.; Hao, C.; Chen, L.; Xie, Y.; Garg, V.; Zhao, L.; Wang, Z.; Zhang, Y.; Li, T.; et al. Pan-genome bridges wheat structural variations with habitat and breeding. Nature 2025, 637, 384–393. [Google Scholar] [CrossRef]
- Parks, M.M.; Lawrence, C.E.; Raphael, B.J. Detecting non-allelic homologous recombination from high-throughput sequencing data. Genome Biol. 2015, 16, 72. [Google Scholar] [CrossRef]
- Robberecht, C.; Voet, T.; Zamani Esteki, M.; Nowakowska, B.A.; Vermeesch, J.R. Nonallelic homologous recombination between retrotransposable elements is a driver of de novo unbalanced translocations. Genome Res. 2013, 23, 411–418. [Google Scholar] [CrossRef]
- Nenarokova, A.; Záhonová, K.; Krasilnikova, M.; Gahura, O.; McCulloch, R.; Zíková, A.; Yurchenko, V.; Lukeš, J. Causes and effects of loss of classical nonhomologous end joining pathway in Parasitic Eukaryotes. Microbiol. Biol. 2019, 10, e01541-19. [Google Scholar] [CrossRef]
- Ferguson, D.O.; Sekiguchi, J.M.; Chang, S.; Frank, K.M.; Gao, Y.; DePinho, R.A.; Alt, F.W. The nonhomologous end-joining pathway of DNA repair is required for genomic stability and the suppression of translocations. Proc. Natl. Acad. Sci. USA 2000, 97, 6630–6633. [Google Scholar] [CrossRef]
- Zhang, F.; Khajavi, M.; Connolly, A.M.; Towne, C.F.; Batish, S.D.; Lupski, J.R. The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nat. Genet. 2009, 41, 849–853. [Google Scholar] [CrossRef] [PubMed]
- McClintock, B. The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. USA 1950, 36, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Liu, J.; Zhang, T.; Su, T.; Li, S.; Cheng, Q.; Kong, L.; Li, X.; Bu, T.; Li, H.; et al. A recent retrotransposon insertion of J caused E6 locus facilitating soybean adaptation into low latitude. J. Integr. Plant Biol. 2021, 63, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, J.; Cai, Z.; Huang, Y.; Lv, M.; Du, H.; Gao, Q.; Zuo, Y.; Dong, Z.; Huang, W.; et al. Evolution and domestication footprints uncovered from the genomes of Coix. Mol. Plant 2020, 13, 295–308. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef]
- Qin, C.; Li, Y.H.; Li, D.; Zhang, X.; Kong, L.; Zhou, Y.; Lyu, X.; Ji, R.; Wei, X.; Cheng, Q.; et al. PH13 improves soybean shade traits and enhances yield for high-density planting at high latitudes. Nat. Commun. 2023, 14, 6813. [Google Scholar] [CrossRef]
- Tian, Z.; Chen, B.; Li, H.; Pei, X.; Sun, Y.; Sun, G.; Pan, Z.; Dai, P.; Gao, X.; Geng, X.; et al. Strigolactone-gibberellin crosstalk mediated by a distant silencer fine-tunes plant height in upland cotton. Mol. Plant 2024, 17, 1539–1557. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, J.; Wu, J.; Duan, J.; Liu, Y.; Ye, X.; Zhang, X.; Guo, X.; Gu, Y.; Zhang, L.; et al. A tandem segmental duplication (TSD) in green revolution gene Rht-D1b region underlies plant height variation. New Phytol. 2012, 196, 282–291. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Qin, P.; Lu, H.; Du, H.; Wang, H.; Chen, W.; Chen, Z.; He, Q.; Ou, S.; Zhang, H.; Li, X.; et al. Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 2021, 184, 3542–3558. [Google Scholar] [CrossRef]
- Li, X.; Dai, X.; He, H.; Lv, Y.; Yang, L.; He, W.; Liu, C.; Wei, H.; Liu, X.; Yuan, Q.; et al. A pan-TE map highlights transposable elements underlying domestication and agronomic traits in Asian rice. Natl. Sci. Rev. 2024, 11, 188. [Google Scholar] [CrossRef] [PubMed]
- He, W.; He, H.; Yuan, Q.; Zhang, H.; Li, X.; Wang, T.; Yang, Y.; Yang, L.; Yang, Y.; Liu, X.; et al. Widespread inversions shape the genetic and phenotypic diversity in rice. Sci. Bull. 2024, 69, 593–596. [Google Scholar] [CrossRef]
- Duan, P.; Xu, J.; Zeng, D.; Zhang, B.; Geng, M.; Zhang, G.; Huang, K.; Huang, L.; Xu, R.; Ge, S.; et al. Natural variation in the promoter of GSE5 contributes to grain size diversity in rice. Mol. Plant 2017, 10, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, M.; Sun, Y.; Zhao, P.; Liu, C.; Qing, K.; Hu, X.; Zhong, Z.; Cheng, J.; Wang, H.; et al. Glycine max NNL1 restricts symbiotic compatibility with widely distributed bradyrhizobia via root hair infection. Nat. Plants 2021, 7, 73–86. [Google Scholar] [CrossRef]
- Liu, T.; Liu, H.; Xian, W.; Liu, Z.; Yuan, Y.; Fan, J.; Xiang, S.; Yang, X.; Liu, Y.; Liu, S.; et al. Duplication and sub-functionalization of flavonoid biosynthesis genes plays important role in Leguminosae root nodule symbiosis evolution. J. Integr. Plant Biol. 2024, 66, 2191–2207. [Google Scholar] [CrossRef]
- Liu, Y.; Du, H.; Li, P.; Shen, Y.; Peng, H.; Liu, S.; Zhou, G.A.; Zhang, H.; Liu, Z.; Shi, M.; et al. Pan-genome of wild and cultivated soybeans. Cell 2020, 182, 162–176. [Google Scholar] [CrossRef]
- Xia, Z.; Watanabe, S.; Yamada, T.; Tsubokura, Y.; Nakashima, H.; Zhai, H.; Anai, T.; Sato, S.; Yamazaki, T.; Lü, S.; et al. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. Proc. Natl. Acad. Sci. USA 2012, 109, E2155–E2164. [Google Scholar] [CrossRef]
- Zhai, H.; Lü, S.; Wu, H.; Zhang, Y.; Zhang, X.; Yang, J.; Wang, Y.; Yang, G.; Qiu, H.; Cui, T.; et al. Diurnal expression pattern, allelic variation, and association analysis reveal functional features of the E1 gene in control of photoperiodic flowering in soybean. PLoS ONE 2015, 10, e0135909. [Google Scholar] [CrossRef]
- Tuteja, J.H.; Clough, S.J.; Chan, W.C.; Vodkin, L.O. Tissue-specific gene silencing mediated by a naturally occurring chalcone synthase gene cluster in Glycine max. Plant Cell 2004, 16, 819–835. [Google Scholar] [CrossRef]
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.G.; Diers, B.W.; Hudson, M.E. An efficient method for measuring copy number variation applied to improvement of nematode resistance in soybean. Plant J. 2016, 88, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Sun, H.; Xu, D.; Chen, Q.; Liang, Y.; Wang, X.; Xu, G.; Tian, J.; Wang, C.; Li, D.; et al. ZmCCT9 enhances maize adaptation to higher latitudes. Proc. Natl. Acad. Sci. USA 2018, 115, E334–E341. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Du, Y.; Shen, X.; Li, M.; Sun, W.; Huang, J.; Liu, Z.; Tao, Y.; Zheng, Y.; Yan, J.; et al. KRN4 controls quantitative variation in maize kernel row number. PLoS Genet. 2015, 11, e1005670. [Google Scholar] [CrossRef]
- Sun, X.; Xiang, Y.; Dou, N.; Zhang, H.; Pei, S.; Franco, A.V.; Menon, M.; Monier, B.; Ferebee, T.; Liu, T.; et al. The role of transposon inverted repeats in balancing drought tolerance and yield-related traits in maize. Nat. Biotechnol. 2023, 41, 120–127. [Google Scholar] [CrossRef]
- Francia, E.; Morcia, C.; Pasquariello, M.; Mazzamurro, V.; Milc, J.A.; Rizza, F.; Terzi, V.; Pecchioni, N. Copy number variation at the HvCBF4-HvCBF2 genomic segment is a major component of frost resistance in barley. Plant Mol. Biol. Rep. 2016, 92, 161–175. [Google Scholar] [CrossRef]
- Armour, J.A.; Sismani, C.; Patsalis, P.C.; Cross, G. Measurement of locus copy number by hybridisation with amplifiable probes. Nucleic Acids Res. 2000, 28, 605–609. [Google Scholar] [CrossRef]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.J.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-toxicity tolerance in barley arising from efflux transporter amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Nitcher, R.; Distelfeld, A.; Tan, C.; Yan, L.; Dubcovsky, J. Increased copy number at the HvFT1 locus is associated with accelerated flowering time in barley. Mol. Plant Genom. 2013, 288, 261–275. [Google Scholar] [CrossRef]
- Yin, D.; Ji, C.; Song, Q.; Zhang, W.; Zhang, X.; Zhao, K.; Chen, C.Y.; Wang, C.; He, G.; Liang, Z.; et al. Comparison of Arachis monticola with diploid and cultivated tetraploid genomes reveals asymmetric subgenome evolution and improvement of peanut. Adv. Sci. 2019, 7, 1901672. [Google Scholar] [CrossRef]
- Li, N.; He, Q.; Wang, J.; Wang, B.; Zhao, J.; Huang, S.; Yang, T.; Tang, Y.; Yang, S.; Aisimutuola, P.; et al. Super-pangenome analyses highlight genomic diversity and structural variation across wild and cultivated tomato species. Nat. Genet. 2023, 55, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Jiang, N.; Schaffner, E.; Stockinger, E.J.; van der Knaap, E. A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit. Science 2008, 319, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, L.; Li, H.; He, J.; Chao, H.; Yan, S.; Yin, Y.; Zhao, W.; Li, M. 3D genome structural variations play important roles in regulating seed oil content of Brassica napus. Plant Commun. 2024, 5, 100666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Z.; He, Y.; Liu, D.; Liu, Y.; Liang, C.; Xie, M.; Jia, Y.; Ke, Q.; Zhou, Y.; et al. Structural variation reshapes population gene expression and trait variation in 2105 Brassica napus accessions. Nat. Genet. 2024, 56, 2538–2550. [Google Scholar] [CrossRef]
- Su, H.; Cao, L.; Ren, Z.; Sun, W.; Zhu, B.; Ma, S.; Sun, C.; Zhang, D.; Liu, Z.; Zeng, H.; et al. ZmELF6-ZmPRR37 module regulates maize flowering and salt response. Plant Biotechnol. J. 2024, 22, 929–945. [Google Scholar] [CrossRef]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major impacts of widespread structural variation on gene expression and crop improvement in Tomato. Cell 2020, 182, 145–161.e23. [Google Scholar] [CrossRef]
- Harberd, N.P.; Bartels, D.; Thompson, R.D. Analysis of the gliadin multigene loci in bread wheat using nullisomic-tetrasomic lines. Mol. Gen. Genet. 1985, 198, 234–242. [Google Scholar] [CrossRef]
- Beom, H.R.; Kim, J.S.; Jang, Y.R.; Lim, S.H.; Kim, C.K.; Lee, C.K.; Lee, J.Y. Proteomic analysis of low-molecular-weight glutenin subunits and relationship with their genes in a common wheat variety. 3 Biotech 2018, 8, 56. [Google Scholar] [CrossRef]
- Jiao, Y.; Deng, X.W. A genome-wide transcriptional activity survey of rice transposable element-related genes. Genome Biol. 2007, 8, R28. [Google Scholar] [CrossRef]
- Yosuke, K.; Yukio, T.; Saori, S.; Ryohei, S.; Motoaki, K.; Tetsuo, S.; Shigeyuki, B.; Yukiko, E.; Hitoshi, N.; Shigeyuki, M. OARE-1, a Ty1-copia retrotransposon in oat activated by abiotic and biotic stresses. Plant Cell Physiol. 2001, 42, 1345–1354. [Google Scholar]
- Takeda, S.; Sugimoto, K.; Otsuki, H.; Hirochika, H. A 13-bp cis-regulatory element in the LTR promoter of the tobacco retrotransposon Tto1 is involved in responsiveness to tissue culture, wounding, methyl jasmonate and fungal elicitors. Plant J. 1999, 18, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Knox, A.K.; Dhillon, T.; Cheng, H.; Tondelli, A.; Pecchioni, N.; Stockinger, E.J. CBF gene copy number variation at Frost Resistance-2 is associated with levels of freezing tolerance in temperate-climate cereals. Theor. Appl. Genet. 2010, 121, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.; Zhu, J.; Boldizsár, Á.; Vágújfalvi, A.; Burke, A.; Garland-Campbell, K.; Galiba, G.; Dubcovsky, J. Large deletions in the CBF gene cluster at the Fr-B2 locus are associated with reduced frost tolerance in wheat. Theor. Appl. Genet. 2013, 126, 2683–2697. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.; Zikhali, M.; Turner, A.S.; Isaac, P.; Laurie, D.A. Copy number variation affecting the Photoperiod-B1 and Vernalization-A1 genes is associated with altered flowering time in wheat (Triticum aestivum). PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, W.; Meng, Z.; Braz, G.T.; Li, Y.; Wang, K.; Wang, H.; Lai, J.; Jiang, J.; Dong, Z.; et al. Megabase-scale presence-absence variation with Tripsacum origin was under selection during maize domestication and adaptation. Genome Biol. 2021, 22, 237. [Google Scholar] [CrossRef]
- Lou, H.; Li, S.; Shi, Z.; Zou, Y.; Zhang, Y.; Huang, X.; Yang, D.; Yang, Y.; Li, Z.; Xu, C. Engineering source-sink relations by prime editing confers heat-stress resilience in tomato and rice. Cell 2025, 188, 530–549. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | Gene | Type | Traits | References |
|---|---|---|---|---|
| Rice | SD1 | Deletion | Semi-dwarf | Sasaki et al., 2002 [35] |
| Gn1 | Deletion | Grain number | Ashikari et al., 2005 [39] | |
| OsWAK112d | Deletion | Blast disease | Qin et al., 2021 [40] | |
| LIP19 | Insertion | Cold tolerance | Li et al., 2024 [41] | |
| MADS56 | Inversion | Heat tolerance | He et al., 2024 [42] | |
| GSE5 | Deletion | Grain size | Duan et al., 2017 [43] | |
| GL7 | Copy number | Grain size | Wang et al., 2015 [44] | |
| Soybean | PH13 | Translocation | Plant hight | Qin et al., 2023 [36] |
| SINE1 | Insertion | Nitrogenase activity of nodule | Zhang et al., 2021 [45] | |
| CHI | Copy number | Nodule | Liu et al., 2024 [46] | |
| E4 | Translocation | Flowering time | Liu et al., 2020 [47] | |
| E1 | Deletion | Flowering time | Xia et al., 2012 [48] | |
| E1 | Deletion | Flowering time | Zhai et al., 2015 [49] | |
| E6 | Translocation | Flowering time | Fang et al., 2015 [33] | |
| CHS | Copy number | Seed coat color | Tuteja et al., 2004 [50] | |
| CHS | Copy number | Seed coat color | Zhou et al., 2015 [51] | |
| CHS | Copy number | Seed coat color | Liu et al., 2020 [47] | |
| Rhg1 | Copy number | Soybean cyst nematode resistance | Lee et al., 2016 [52] | |
| Maize | ZmCCT9 | Translocation | Flowering time | Huang et al., 2018 [53] |
| ZmCCT10 | Translocation | Flowering time | Huang et al., 2018 [53] | |
| KRN4 | Translocation | Kernel row number | Liu et al., 2015 [54] | |
| ZmMYBR38 | Translocation | Drought response | Sun et al., 2023 [55] | |
| Wheat | Rht-D1c | Copy number | Plant height | Tian et al., 2024; Li et al., 2012 [37,38] |
| FR-2 | Copy number | Low-temperature tolerance | Francia et al., 2016 [56] | |
| Vrn-A1 | Copy number | Flowering time | Jiao et al., 2025; Armour et al., 2000 [26,57] | |
| barley | Bot1 | Copy number | Boron toxicity | Sutton et al., 2007 [58] |
| FT1 | Copy number | Flowering time | Nitcher et al., 2013 [59] | |
| Peanut | ARF2 | Deletion | Seed size | Yin et al., 2019 [60] |
| Tomato | Cytochrome P450 | Deletion | Plant architecture | Li et al., 2023 [61] |
| SUN | Translocation | Grain size | Xiao et al., 2008 [62] | |
| Brassica napus | BnaA09g48250D | Insertion | oil content | Zhang et al., 2024 [63] |
| BnaA03.MAMf | Deletion | glucosinolates ratio | Zhang et al., 2024 [64] | |
| BnaA09.MYB28 | Insertion | glucosinolate content | Zhang et al., 2024 [64] | |
| Upland cotton | GhPH1 | Insertion | Plant height | Tian et al., 2024 [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Liu, C.; Sun, X.; Sun, G.; Zong, C.; Qi, Y.; Bai, Y.; Li, W.; Kong, F.; Li, H.; et al. How Structural Variations Influence Crop Improvement. Int. J. Mol. Sci. 2025, 26, 6635. https://doi.org/10.3390/ijms26146635
Wang X, Liu C, Sun X, Sun G, Zong C, Qi Y, Bai Y, Li W, Kong F, Li H, et al. How Structural Variations Influence Crop Improvement. International Journal of Molecular Sciences. 2025; 26(14):6635. https://doi.org/10.3390/ijms26146635
Chicago/Turabian StyleWang, Xiaomei, Changyuan Liu, Xiaohuan Sun, Guohong Sun, Chunmei Zong, Yuxin Qi, Yanfeng Bai, Wen Li, Fanjiang Kong, Haiyang Li, and et al. 2025. "How Structural Variations Influence Crop Improvement" International Journal of Molecular Sciences 26, no. 14: 6635. https://doi.org/10.3390/ijms26146635
APA StyleWang, X., Liu, C., Sun, X., Sun, G., Zong, C., Qi, Y., Bai, Y., Li, W., Kong, F., Li, H., & Wang, Y. (2025). How Structural Variations Influence Crop Improvement. International Journal of Molecular Sciences, 26(14), 6635. https://doi.org/10.3390/ijms26146635