
The Discovery of Potential Repellent Compounds for Zeugodacus cucuribitae (Coquillett) from Six Non-Favored Hosts
 ,
, 
Abstract
1. Introduction
2. Results
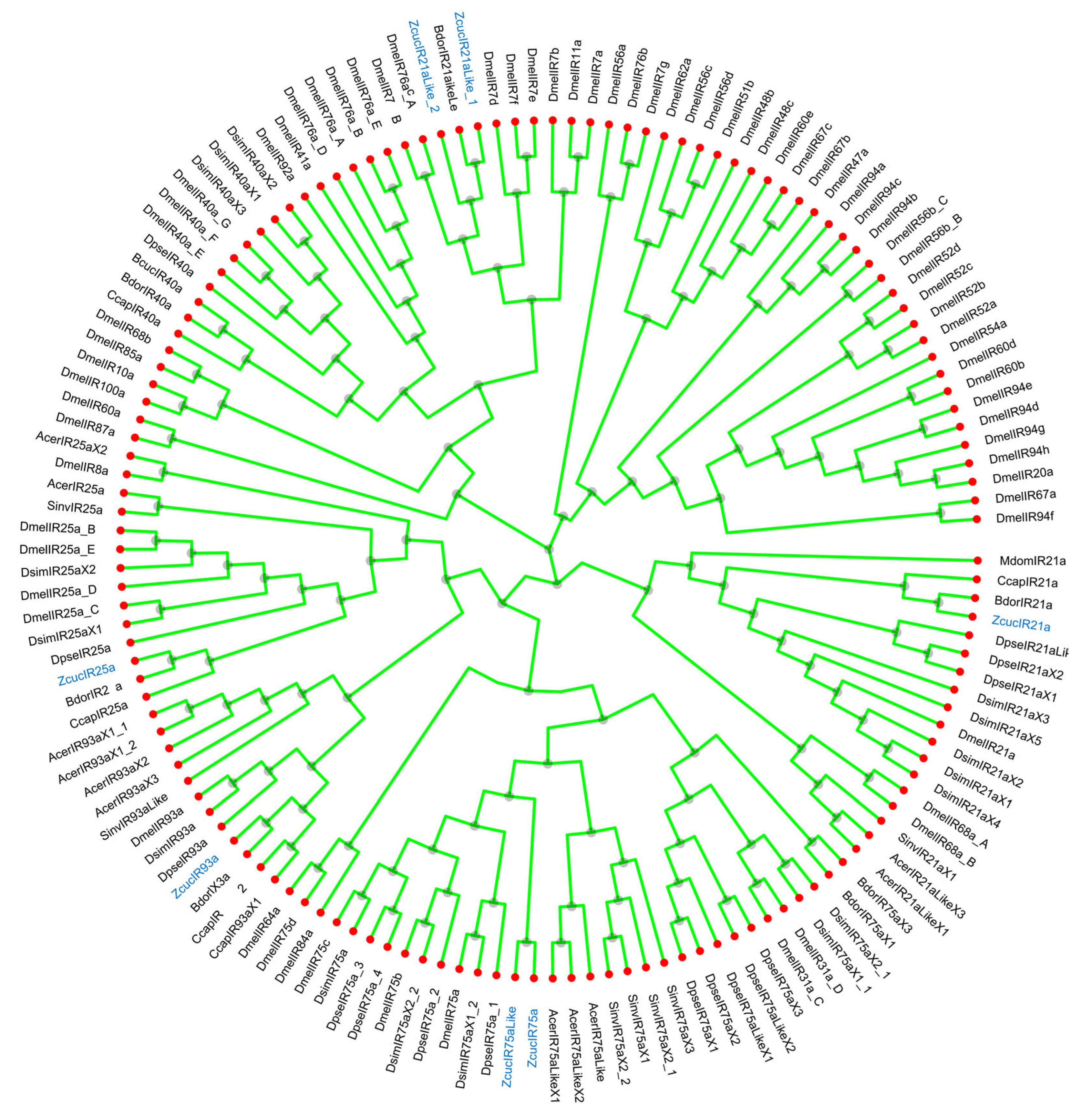
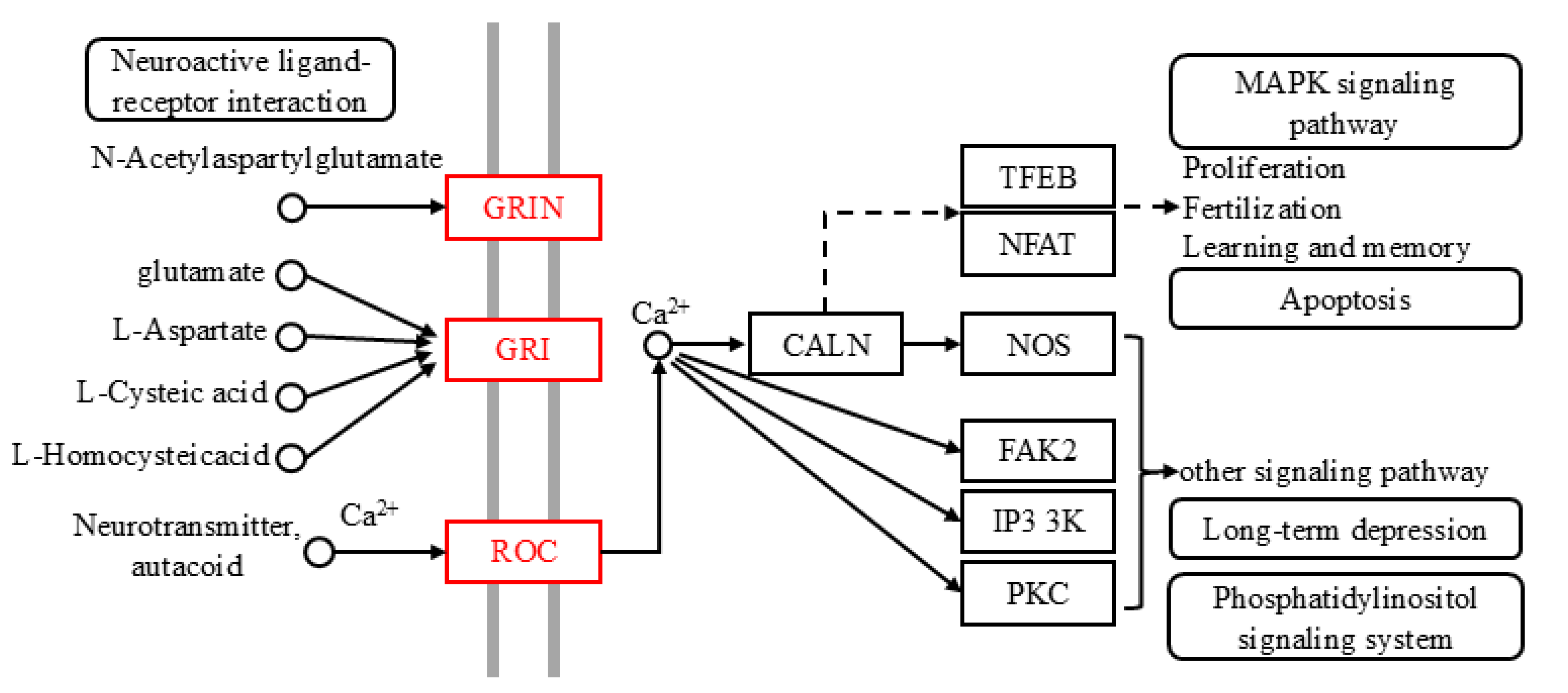
2.1. Identification and Enrichment of Olfactory Sensory Genes in Z. cucuribitae
2.1.1. Phylogenetic Analysis
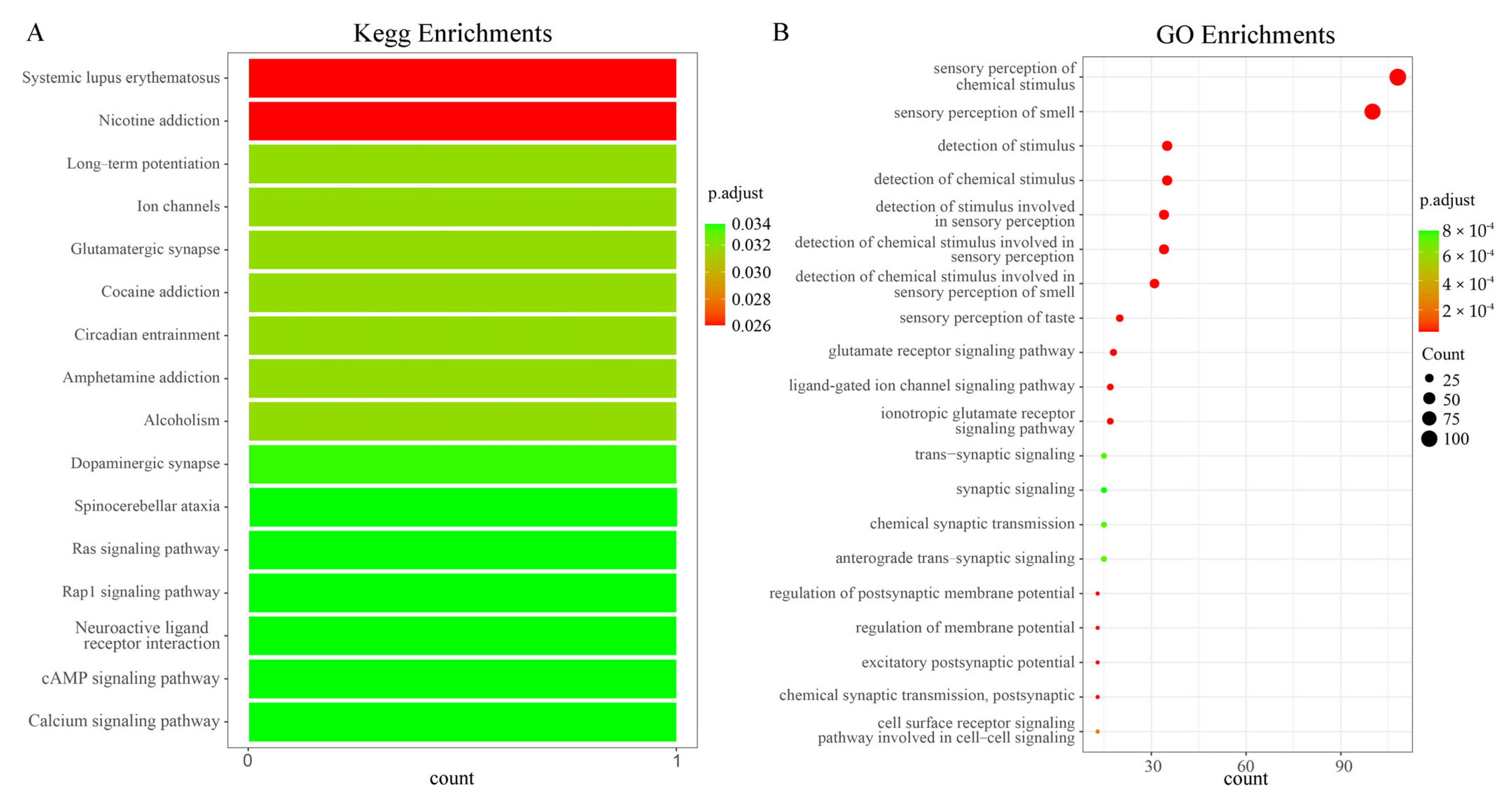
2.1.2. KEGG and GO Enrichment
2.1.3. Metabolite Analysis of L. acutangular, L. cylindrica, S. edule, B. oleracea, M. nana, and F. ananassa
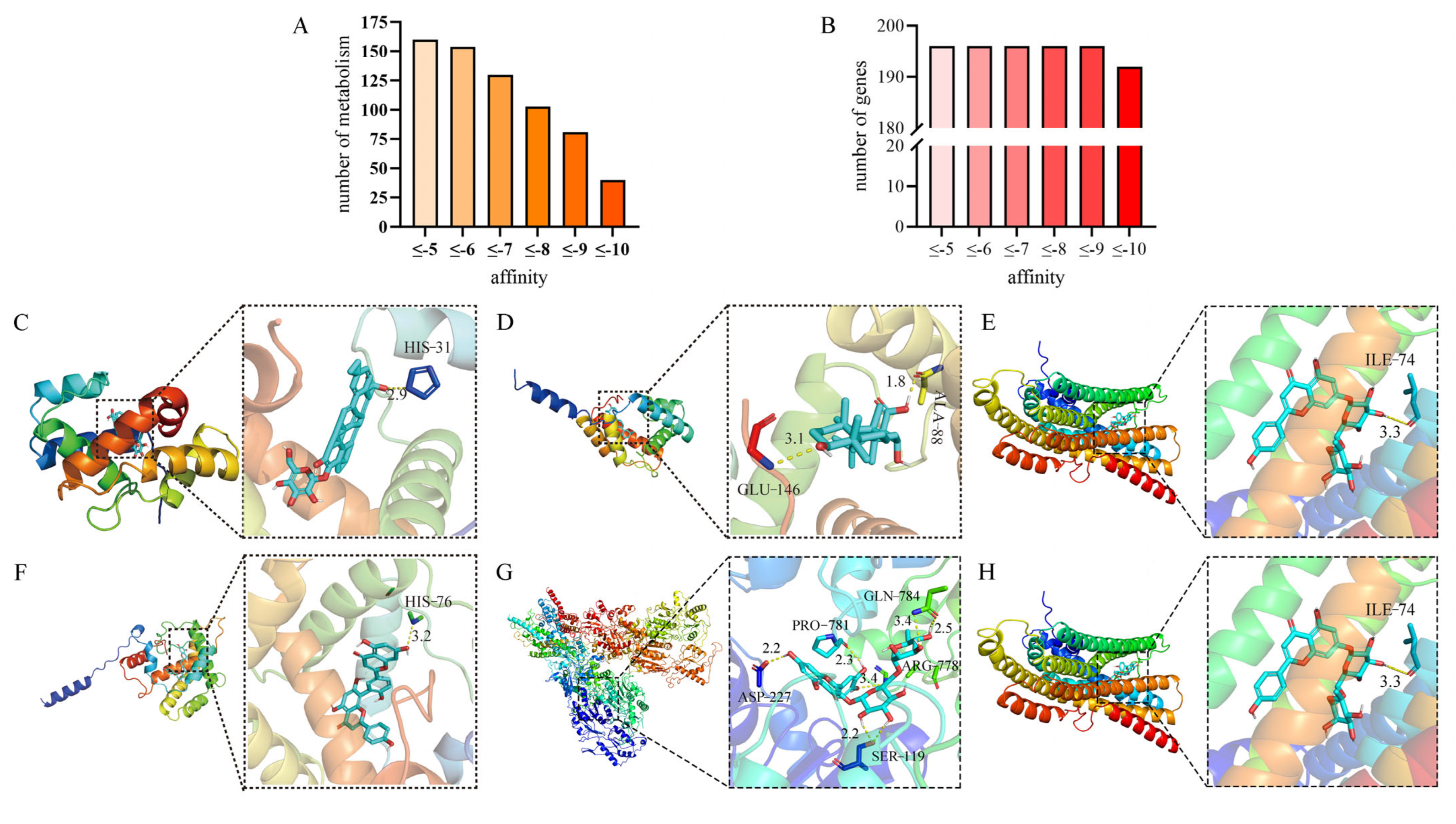
2.2. Molecular Docking
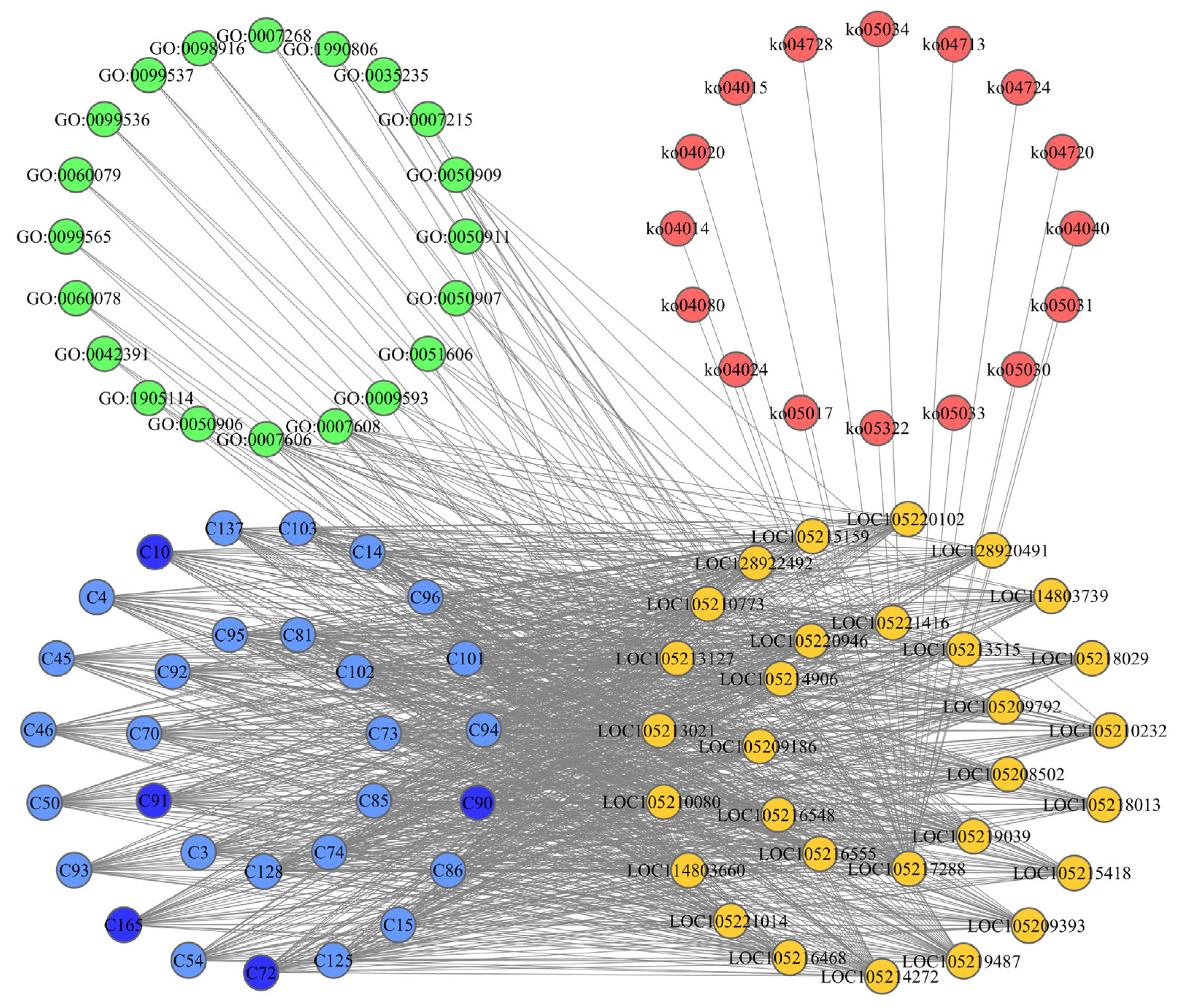
2.3. Construction of CPPG Networks and Screening of Core Functional Compounds
2.4. Behavior Determination of Z. cucuribitae
2.4.1. Results of Two-Way Selection Experiments
2.4.2. Results of Gastric Toxicity Experiments
2.5. Predicted Results of Molecular Mechanisms
3. Discussions
4. Materials and Methods
4.1. Insect Rearing
4.2. Identification and Functional Enrichment of Odor-Sensing Genes from Z. cucuribitae
4.3. Acquisition of Compound Models of L. acutangular, L. cylindrica, S. edule, B. oleracea, M. nana, and F. ananassa and Protein Models of Z. cucuribitae
4.4. CPPG Networks and Screening of Core Functional Compounds
4.4.1. Molecular Docking of Olfactory Sensory Genes and Metabolic Compounds
4.4.2. Construction of CPPG Networks and Screening of Core Functional Compounds
4.5. Effect of Core Compounds on Behavior of Z. cucuribitae
4.5.1. Two-Way Selection Experiment
4.5.2. Gastric Toxicity Experiment
4.6. Predicted Mechanism of Core Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| A. Cerana | Apis cerana |
| Acer | Apis cerana |
| Z. cucuribitae | Zeugodacus cucuribitae (Coquillett) |
| B. dorsalis | Bactrocera dorsalis |
| B. oleracea | Brassica oleracea var. botrytis |
| Bdor | Bactrocera dorsalis |
| Bole | Bactrocera oleae |
| C.pepo | Cucurbita pepo |
| CPPG | Genes vs. components vs. Kegg pathways vs. GO terms |
| D. melanogaster | Drosophila melanogaster |
| Dana | Drosophila ananassae |
| Dmel | Drosophila melanogaster |
| Dsim | Drosophila simulans |
| F. ananassa | Fragaria × ananassa |
| GR | Olfactory receptor |
| IR | Ionospheric receptor |
| L. cylindrica | Luffa cylindrica |
| L. acutangular | Luffa acutangular |
| M. domestica | Musca domestica |
| M. nana | Musa nana |
| Mdom | Musca domestica |
| OBPGOBP | Odor-binding protein and general odorant-binding proteins |
| OR | Olfactory receptor |
| PBP | Pheromone-binding protein |
| S. edule | Sechium edule |
| SNMP | Sensory neuron membrane protein |
| Bcuc | Zeugodacus cucuribitae (Coquillett) |
References
- Dhillon, M.K.; Singh, R.; Naresh, J.S.; Sharma, H.C. The Melon Fruit Fly, Bactrocera Cucurbitae: A Re-view of Its Biology and Management. J. Insect Sci. 2005, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Koyama, J.; Kakinohana, H.; Miyatake, T. Eradication of the Melon Fly, Bactrocera Cucurbitae, in Japan: Importance of Behavior, Ecology, Genetics, and Evolution. Annu. Rev. Entomol. 2004, 49, 331–349. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, M.; Delatte, H.; Mwatawala, M.; Quilici, S.; Vayssières, J.F.; Virgilio, M. A Review of the Current Knowledge on Zeugodacus Cucurbitae (Coquillett) (Diptera, Tephritidae) in Africa, with a List of Species Included in Zeugodacus. Zookeys 2015, 540, 539–557. [Google Scholar] [CrossRef]
- Dorla, E.; Gauvin-Bialecki, A.; Deuscher, Z.; Allibert, A.; Grondin, I.; Deguine, J.P.; Laurent, P. Insecticidal Activity of the Leaf Essential Oil of Peperomia Borbonensis Miq. (Piperaceae) and Its Major Components against the Melon Fly Bactrocera Cucurbitae (Diptera: Tephri-tidae). Chem. Biodivers. 2017, 14, e1600493. [Google Scholar] [CrossRef]
- Hsu, J.-C.; Haymer, D.S.; Chou, M.-Y.; Feng, H.-T.; Chen, H.-H.; Huang, Y.-B.; Mau, R.F.L. Monitoring Resistance to Spinosad in the Melon Fly (Bactrocera cucurbitae) in Hawaii and Taiwan. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Vargas, R.I.; Piñero, J.C.; Leblanc, L. An Overview of Pest Species of Bactrocera Fruit Flies (Diptera: Tephritidae) and the Integration of Biopesticides with Other Biological Approaches for Their Management with a Focus on the Pacific Region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef]
- Pathak, V.M.; Verma, V.K.; Rawat, B.S.; Kaur, B.; Babu, N.; Sharma, A.; Dewali, S.; Yadav, M.; Kumari, R.; Singh, S.; et al. Current Status of Pesticide Effects on Environ-ment, Human Health and It’s Eco-Friendly Management as Bioremediation: A Comprehensive Review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef]
- Wan, N.-F.; Fu, L.; Dainese, M.; Kiær, L.P.; Hu, Y.-Q.; Xin, F.; Goulson, D.; Woodcock, B.A.; Vanbergen, A.J.; Spurgeon, D.J.; et al. Pesticides Have Negative Effects on Non-Target Organisms. Nat. Commun. 2025, 16, 1360. [Google Scholar] [CrossRef]
- Nikolova, I. Pea weevil damage and chemical characteristics of pea cultivars determining their resistance to Bruchus pisorum L. Bull. Èntomol. Res. 2016, 106, 268–277. [Google Scholar] [CrossRef]
- Rubiales, D.; Barilli, E.; Rispail, N. Breeding for Biotic Stress Resistance in Pea. Agriculture 2023, 13, 1825. [Google Scholar] [CrossRef]
- Hisashi, K.-N. Defensive Molecules Momilactones a and B: Function, Biosynthesis, Induction and Occurrence. Toxins 2023, 15, 241. [Google Scholar] [CrossRef] [PubMed]
- Ramniwas, S.; Bilal, T.; Sharma, A. Repellent Activity of Salix Alba Bark Extract and Guava Oil-Based Formulation against the Oriental Fruit Fly, Bactrocera Dorsalis (Diptera: Tephritidae). Int. J. Trop. Insect Sci. 2024, 44, 831–842. [Google Scholar] [CrossRef]
- Akram, W.; Hussain, A.; Abbas, Q.; Abbas, A. Insecticidal Potential of Botanical Ex-tracts for Management of, the Oriental Fruit Fly, Bactrocera Dorsalis (Diptera: Tephritidae). J. Basic Appl. Zool. 2024, 85, 63. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant Secondary Metabolites as Defense Tools against Herbivores for Sustainable Crop Protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Hussain, B.; Buhroo, A.A.; Ignacimuthu, S.; Sharma, H.C. Effect of Plant Secondary Metabolites on Legume Pod Borer, Helicoverpa Armigera. J. Pest Sci. 2013, 86, 399–408. [Google Scholar] [CrossRef]
- Li, L.; Han, D.Y.; Niu, L.M.; Zhang, F.P.; Chen, J.Y.; Fu, Y.G. Performance Evaluation of Melon Fruit Fly, Zeugodacus Cucurbitae on 39 Hosts. J. Environ. Entomol. 2019, 41, 1057–1064. [Google Scholar]
- Farooq, M.; Baig, S.; Honey, S.F.; Bajwa, B.E.; Fazlullah; Shah, I.H. Evaluation of host susceptibility, preference and offspring performance of Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae) on different hosts. Int. J. Trop. Insect Sci. 2019, 40, 93–99. [Google Scholar] [CrossRef]
- Zeng, B.; Lian, Y.; Jia, J.; Liu, Y.; Wang, A.; Yang, H.; Li, J.; Yang, S.; Peng, S.; Zhou, S. Multigenerational Effects of Short-Term High Temperature on the Development and Reproduction of the Zeugodacus cucurbitae (Coquillett, 1899). Agriculture 2022, 12, 954. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; et al. Scientific opinion on the import of Musa fruits as a pathway for the entry of non-EU Tephritidae into the EU territory. EFSA J. 2021, 19, e06426. [Google Scholar] [CrossRef]
- Vieira, F.G.; Rozas, J. Comparative Genomics of the Odorant-Binding and Chemosensory Protein Gene Families across the Arthropoda: Origin and Evolutionary History of the Chemosensory System. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef]
- Hua, Y. Insect Olfactory Neurons: Receptors, Development, and Function. Curr. Opin. Insect Sci. 2025, 67, 101288. [Google Scholar]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: Ionotropic receptors and odorant receptors. Cell Tissue Res. 2021, 383, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, M.; Dong, K.; Zhang, J.; Wang, H.; Xie, M.; Wu, W.; Zhang, Y.-J.; Chen, Z. Structural Insights into the Ligand-Binding and -Releasing Mechanism of Helicoverpa armigera Pheromone-Binding Protein PBP1. Int. J. Mol. Sci. 2022, 23, 1190. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, J.; Zhang, Z.; Yan, M.; Zhang, B.; Zhu, C.; Yan, W.; Shi, B.; Wang, Y.; Zhao, C.; et al. A plant virus enhances odorant-binding protein 5 (OBP5) in the vector whitefly for more actively olfactory orientation to the host plant. Pest Manag. Sci. 2022, 79, 1410–1419. [Google Scholar] [CrossRef]
- Guo, R.; Zheng, Q.; Zhang, L.; Rajasingh, J. Identification and validation of diagnostic markers and drugs for pediatric bronchopulmonary dysplasia based on integrating bioinformatics and molecular docking analysis. PLoS ONE 2025, 20, e0323006. [Google Scholar] [CrossRef]
- Wang, Z.; Li, R.; Chen, X.; Ren, H.; Wang, C.; Min, R.; Zhang, X. Network pharmacology, molecular docking and experimental validation to elucidate the anti-T2DM mechanism of Lanxangia tsaoko. Fitoterapia 2024, 178, 106117. [Google Scholar] [CrossRef]
- Dan, Y.; Jin, Y.; Wang, J.; Wang, L.; Zheng, D. Combination of RNA-sequencing data analysis, network pharmacology, molecular docking techniques to investigate the mechanism of Prunella vulgaris L. in the treatment of non-small cell lung cancer. Discov. Oncol. 2025, 16, 726. [Google Scholar] [CrossRef]
- Bisht, A.; Tewari, D.; Kumar, S.; Chandra, S. Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation to Elucidate the Mechanism of Anti-Aging Action of Tino-spora Cordifolia. Mol. Divers. 2024, 28, 1743–1763. [Google Scholar] [CrossRef]
- Zou, Y.; Chen, Y.; Wang, D.; Xie, X.; Li, G.; Zheng, C.; Wen, J.; Su, H.; Liu, X.; Zeng, L.; et al. The Effects of Nine Compounds on Aldehyde-Oxidase-Related Genes in Bactrocera dorsalis (Hendel). Genes 2023, 15, 35. [Google Scholar] [CrossRef]
- Wang, J.J.; Ma, C.; Tian, Z.Y.; Zhou, Y.P.; Yang, J.F.; Gao, X.; Chen, H.S.; Ma, W.H.; Zhou, Z.S. Electroantennographic and Behavioral Responses of the Melon Fly, Zeugo-dacus Cucurbitae (Coquillett), to Volatile Compounds of Ridge Gourd, Luffa acutangular L. J. Chem. Ecol. 2024, 50, 1036–1045. [Google Scholar] [CrossRef]
- Ebeed, H.; Baz, M.; Habib, E.; Prabhu, S.; Ceasar, S.A. Integrated Metabolomic Analysis and Molecular Docking: Unveiling the Potential of Nephrolepis Exaltata (L.) Schott Phytocompounds for Mosquito Control Via Glutathione-S-Transferase Targeting. Int. J. Biol. Macromol. 2024, 273, 133072. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qiu, L.; Wang, B.; Guan, Z.; Dong, Z.; Zhang, J.; Cao, S.; Yang, L.; Wang, B.; Gong, Z.; et al. Structural basis for odorant recognition of the insect odorant receptor OR-Orco heterocomplex. Science 2024, 384, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Newman, Z.L.; Bakshinskaya, D.; Schultz, R.; Kenny, S.J.; Moon, S.; Aghi, K.; Stanley, C.; Marnani, N.; Li, R.; Bleier, J.; et al. Determinants of synapse diversity revealed by super-resolution quantal transmission and active zone imaging. Nat. Commun. 2022, 13, 229. [Google Scholar] [CrossRef]
- Jiang, X.; Dimitriou, E.; Grabe, V.; Sun, R.; Chang, H.; Zhang, Y.; Gershenzon, J.; Rybak, J.; Hansson, B.S.; Sachse, S. Ring-shaped odor coding in the antennal lobe of migratory locusts. Cell 2024, 187, 3973–3991.e24. [Google Scholar] [CrossRef]
- Patel, S.B.; Ghane, S.G. Phyto-Constituents Profiling of Luffa Echinata and in Vitro As-sessment of Antioxidant, Anti-Diabetic, Anticancer and Anti-Acetylcholine Esterase Activities. Saudi J. Biol. Sci. 2021, 28, 3835–3846. [Google Scholar] [CrossRef]
- Chanda, J.; Mukherjee, P.K.; Biswas, R.; Biswas, S.; Tiwari, A.K.; Pargaonkar, A. Uplc-Qtof-Ms Analysis of a Carbonic Anhydrase-Inhibiting Extract and Fractions of (L.) Roxb (Ridge Gourd). Phytochem. Anal. 2019, 30, 148–155. [Google Scholar] [CrossRef]
- Ceci, C.; Lacal, P.M.; Tentori, L.; De Martino, M.G.; Miano, R.; Graziani, G. Experimental Evidence of the Antitumor, Antimetastatic and Antiangiogenic Activity of Ellagic Ac-id. Nutrients 2018, 10, 1756. [Google Scholar] [CrossRef]
- Ben Hlel, T.; Belhadj, F.; Gül, F.; Altun, M.; Yağlıoğlu, A.Ş.; Demirtaş, I.; Marzouki, M.N. Variations in the Bioactive Compounds Composition and Biological Activities of Loofah (Luffa cylindrica) Fruits in Relation to Maturation Stages. Chem. Biodivers. 2017, 14, e1700178. [Google Scholar] [CrossRef]
- Gedük, A.Ş.; Zengin, F. Lc–Ms/Ms Characterization, Antidiabetic, Antioxidative, and An-tibacterial Effects of Different Solvent Extracts of Anamur Banana (Musa Cavendishii). Food Sci. Biotechnol. 2021, 30, 1183–1193. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Wiczkowski, W.; Płatosz, N.; Bartoszek, A. Phytochemical Composition and Biological Activities of Differently Pigmented Cabbage (Bras-sica Oleracea Var. Capitata) and Cauliflower (Brassica Oleracea Var. Botrytis) Varieties. J. Sci. Food Agric. 2019, 99, 5499–5507. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lim, S.-H.; Ha, S.-H.; Yeo, Y.; Park, W.T.; Kwon, D.Y.; Park, S.U.; Kim, J.K. Metabolite Profiling Approach Reveals the Interface of Primary and Secondary Metabolism in Colored Cauliflowers (Brassica oleracea L. ssp. botrytis). J. Agric. Food Chem. 2013, 61, 6999–7007. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Rosario, A.C.R.S.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.S.; El-Hefny, M.; Ghoneim, I.M.; El-Lahot, M.S.R.A.; Akrami, M.; Al-Huqail, A.A.; Ali, H.M.; Abd-Elkader, D.Y. Assessing the Use of Aloe vera Gel Alone and in Combination with Lemongrass Essential Oil as a Coating Material for Strawberry Fruits: HPLC and EDX Analyses. Coatings 2022, 12, 489. [Google Scholar] [CrossRef]
- Maamoun, A.A.; El-Akkad, R.H.; Farag, M.A. Mapping metabolome changes in Luffa aegyptiaca Mill fruits at different maturation stages via MS-based metabolomics and chemometrics. J. Adv. Res. 2021, 29, 179–189. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Díaz-De-Cerio, E.; Verardo, V.; Fernández-Gutiérrez, A.; Gómez-Caravaca, A.M. New insight into phenolic composition of chayote (Sechium edule (Jacq.) Sw.). Food Chem. 2019, 295, 514–519. [Google Scholar] [CrossRef]
- Singh, A.S.; Vellapandian, C. Phytochemical Studies, Antioxidant Potential, and Identification of Bioactive Compounds Using Gc–Ms of the Ethanolic Extract of Luffa cylindrica (L.) Fruit. Appl. Biochem. Biotechnol. 2022, 194, 4018–4032. [Google Scholar]
- Hu, W.; Wang, X.; Wu, L.; Shen, T.; Ji, L.; Zhao, X.; Si, C.L.; Jiang, Y.; Wang, G. Apigenin-7-O-Β-D-Glucuronide Inhibits Lps-Induced Inflammation through the Inac-tivation of Ap-1 and Mapk Signaling Pathways in Raw 264.7 Macrophages and Protects Mice against En-dotoxin Shock. Food Funct. 2016, 7, 1002–1013. [Google Scholar] [CrossRef]
- Nath, L.R.; Gorantla, J.N.; Joseph, S.M.; Antony, J.; Thankachan, S.; Menon, D.B.; Sankar, S.; Lankalapalli, R.S.; Anto, R.J. Kaempferide, the most active among the four flavonoids isolated and characterized from Chromolaena odorata, induces apoptosis in cervical cancer cells while being pharmacologically safe. RSC Adv. 2015, 5, 100912–100922. [Google Scholar] [CrossRef]
- Arab, H.H.; Salama, S.A.; Omar, H.A.; Arafa, E.-S.A.; Maghrabi, I.A.; Sen, U. Diosmin Protects against Ethanol-Induced Gastric Injury in Rats: Novel Anti-Ulcer Actions. PLoS ONE 2015, 10, e0122417. [Google Scholar] [CrossRef]
- Siciliano, T.; De Tommasi, N.; Morelli, I.; Braca, A. Study of Flavonoids of Sechium edule (Jacq) Swartz (Cucurbitaceae) Different Edible Organs by Liquid Chromatography Photodiode Array Mass Spectrometry. J. Agric. Food Chem. 2004, 52, 6510–6515. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Yang, H.; Son, G.W.; Park, H.R.; Park, C.S.; Jin, Y.H.; Park, Y.S. Eriodictyol Protects Endothelial Cells against Oxidative Stress-Induced Cell Death through Modu-lating Erk/Nrf2/Are-Dependent Heme Oxygenase-1 Expression. Int. J. Mol. Sci. 2015, 16, 14526–14539. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A Review of the Most Recent Research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef]
- Isemura, M. Catechin in Human Health and Disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, T.; Zhu, X.; Yang, C.; Wang, Y.; Zhou, N.; Ju, B.; Zhou, T.; Deng, G.; Qiu, C. Hyperoside Induces Breast Cancer Cells Apoptosis via ROS-Mediated NF-κB Signaling Pathway. Int. J. Mol. Sci. 2019, 21, 131. [Google Scholar] [CrossRef]
- Govindarasu, M.; Thangaraj, K.; Murugesan, V.; Vaiyapuri, M. Kaempferitrin Cause Cell Cycle Arrest at G2/M Phase and Reactive Oxygen Species Mediated Apoptosis in Human Colon Cancer Ht-29 Cells. Int. J. Recent Technol. Eng. Int. J. Recent Technol. Eng. 2020, 8, 2277–3878. [Google Scholar]
- Zhang, Y.; Guo, Y.; Wang, M.; Dong, H.; Zhang, J.; Zhang, L. Quercetrin from Toona sinensis leaves induces cell cycle arrest and apoptosis via enhancement of oxidative stress in human colorectal cancer SW620 cells. Oncol. Rep. 2017, 38, 3319–3326. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Locatelli, M.; Granica, S.; Cacciagrano, F.; Tomczyk, M. A Review on the Dietary Flavonoid Tiliroside. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1395–1421. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, Y.J.; Shin, M.-S.; Kim, H.-R.; Kim, M.J.; Lee, S.H.; Yun, S.P.; Kwon, S.-H. Acacetin inhibits neuronal cell death induced by 6-hydroxydopamine in cellular Parkinson’s disease model. Bioorganic Med. Chem. Lett. 2017, 27, 5207–5212. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, Y.; Liu, H.; Li, P.; Zhang, H.; Cheng, G. Fortunellin Pro-tects against High Fructose-Induced Diabetic Heart Injury in Mice by Suppressing Inflammation and Oxi-dative Stress Via Ampk/Nrf-2 Pathway Regulation. Biochem. Biophys. Res. Commun. 2017, 490, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, B.; Lu, F.; Hu, X.; Tang, J.; Huang, L. Inhibitory activity of linarin on osteoclastogenesis through receptor activator of nuclear factor κB ligand-induced NF-κB pathway. Biochem. Biophys. Res. Commun. 2018, 495, 2133–2138. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Hu, X.; Gong, D.; Zhang, G. Inhibitory mechanism of vitexin on α-glucosidase and its synergy with acarbose. Food Hydrocoll. 2020, 105, 105824. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, T.; Li, M.-F.; Yang, Y.-S.; Li, R.; Tan, J.; Tang, S.-H.; Jiang, Z.-T. Composition, cytotoxicity and antioxidant activities of polyphenols in the leaves of star anise (Illicium verum Hook. f.). ScienceAsia 2019, 45, 532–537. [Google Scholar] [CrossRef]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic Acid and Its De-rivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef]
- Yu, Q.; Li, X.; Cao, X. Linarin Could Protect Myocardial Tissue from the Injury of Ische-mia-Reperfusion through Activating Nrf-2. Biomed. Pharmacother. 2017, 90, 1–7. [Google Scholar] [CrossRef]
- Karayildirim, T.; Ozmen, A.; Emirdag-Ozturk, S.; Capaci-Karagoez, A.; Poyrazoglu-Coban, E. Synthesis, Antimicrobial and Cytotoxic Activities, and Structure-Activity Relationships of Gyp-sogenin Derivatives against Human Cancer Cells. Eur. J. Med. Chem. Chim. 2014, 82, 565–573. [Google Scholar]
- Ren, Y.; Shen, L.; Zhang, D.W.; Dai, S.J. Two New Sesquiterpenoids from Solanum Lyratum with Cytotoxic Activities. Chem. Pharm. Bull. 2009, 57, 408–410. [Google Scholar] [CrossRef]
- Subagio, A.; Morita, N.; Sawada, S. Carotenoids and Their Fatty-Acid Esters in Bana-na Peel. J. Nutr. Sci. Vitaminol. 1996, 42, 553–566. [Google Scholar] [CrossRef]
- Cozzolino, R.; Pace, B.; Palumbo, M.; Laurino, C.; Picariello, G.; Siano, F.; De Giulio, B.; Pelosi, S.; Cefola, M. Profiles of Volatile and Phenolic Compounds as Markers of Ripening Stage in Candonga Strawberries. Foods 2021, 10, 3102. [Google Scholar] [CrossRef]
- Hu, F.; Chen, J.; Zhang, Y.; Sun, Y.; Liu, Y.; Yu, Y.; Xu, K.; Cai, H. Novel Bio-transformation of Maslinic Acid to Ma-2-O-Β-D-Glucoside by Udp-Glycosyltransferases from Bacillus Sub-tilis. Catalysts 2022, 12, 884. [Google Scholar] [CrossRef]
- Ryu, S.; Shin, J.S.; Jung, J.Y.; Cho, Y.W.; Kim, S.J.; Jang, D.S.; Lee, K.T. Echinocystic Acid Isolated from Eclipta Prostrata Suppresses Lipopolysaccharide-Induced Inos, Tnf-A, and Il-6 Expressions Via Nf-Κb Inac-tivation in Raw 264.7 Macrophages. Planta Med. 2013, 79, 1031–1037. [Google Scholar] [PubMed]
- Riddick, E.W. Evaluating the Effects of Flavonoids on Insects: Implications for Managing Pests Without Harming Beneficials. Insects 2024, 15, 956. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.; Wang, X.W.; Elkahky, M. Transitioning Towards Dynamic, Na-ture-Based Crop Defenses. J. Biosci. 2024, 49, 99. [Google Scholar] [CrossRef]
- Stec, K.; Kordan, B.; Gabryś, B. Quercetin and Rutin as Modifiers of Aphid Probing Behavior. Molecules 2021, 26, 3622. [Google Scholar] [CrossRef]
- Scott-Fordsmand, J.J.; Fraceto, L.F.; Amorim, M.J.B. Nano-pesticides: The lunch-box principle—Deadly goodies (semio-chemical functionalised nanoparticles that deliver pesticide only to target species). J. Nanobiotechnology 2022, 20, 13. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Campos, J.M.; da Silva Rolim, G.; Martínez, L.C.; Dos Santos, M.; Fernandes, F.L.; Serrão, J.E.; Zanuncio, J.C. Terpenoid constituents of cinnamon and clove essential oils cause toxic effects and behavior repellency response on granary weevil, Sitophilus granarius. Ecotoxicol. Environ. Saf. 2018, 156, 263–270. [Google Scholar] [CrossRef]
- You, C.-X.; Zhang, W.-J.; Guo, S.-S.; Wang, C.-F.; Yang, K.; Liang, J.-Y.; Wang, Y.; Geng, Z.-F.; Du, S.-S.; Deng, Z.-W. Chemical composition of essential oils extracted from six Murraya species and their repellent activity against Tribolium castaneum. Ind. Crop. Prod. 2015, 76, 681–687. [Google Scholar] [CrossRef]
- Hazarika, M.; Nath, P.; Begum, T.; Lal, M.; Kalita, J.; Dutta, P. In-sights into Insecticidal Efficacy of Cymbopogon Essential Oils against Callosobruchus Chinensis: An Inte-grated Approach through Bioassays and in-Silico Molecular Docking for Sustainable Pest Management. J. Stored Prod. Res. 2025, 112, 102655. [Google Scholar] [CrossRef]
- de Oliveira, P.M.C.; Sousa, J.P.B.; Albernaz, L.C.; Coelho-Ferreira, M.; Espindola, L.S. Bioprospection for new larvicides against Aedes aegypti based on ethnoknowledge from the Amazonian São Sebastião de Marinaú riverside community. J. Ethnopharmacol. 2022, 293, 115284. [Google Scholar] [CrossRef]
- Mallavadhani, U.V.; Mahapatra, A.; Raja, S.S.; Manjula, C. Antifeedant Activity of Some Pentacyclic Triterpene Acids and Their Fatty Acid Ester Analogues. J. Agric. Food Chem. 2003, 51, 1952–1955. [Google Scholar] [CrossRef] [PubMed]
- Cen, Y.; Zou, X.; Li, L.; Chen, S.; Lin, Y.; Liu, L.; Zheng, S. Inhibition of the glutathione biosynthetic pathway increases phytochemical toxicity to Spodoptera litura and Nilaparvata lugens. Pestic. Biochem. Physiol. 2020, 168, 104632. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Xue, L.; Li, Y.; Cui, G.; Sun, R.; Hu, M.; Zhong, G. Rote-none-Induced Necrosis in Insect Cells Via the Cytoplasmic Membrane Damage and Mitochondrial Dys-function. Pestic. Biochem. Physiol. 2021, 173, 104801. [Google Scholar] [CrossRef]
- Chen, J.; Mu, X.; Liu, H.; Yong, Q.; Ouyang, X.; Liu, Y.; Zheng, L.; Chen, H.; Zhai, Y.; Ma, J.; et al. Rotenone impairs brain glial energetics and locomotor behavior in bumblebees. Sci. Total. Environ. 2023, 907, 167870. [Google Scholar] [CrossRef]
- Wei, F.; Wang, D.; Li, H.; Xia, P.; Ran, Y.; You, J. Toxicogenomics Provides Insights to Toxicity Pathways of Neonicotinoids to Aquatic Insect, Chironomus Dilutus. Environ. Pollut. 2020, 260, 114011. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, F.; Pan, H.; Ye, J.; Dong, X.; Li, C.; Lin, F. Identifying Virulence-Associated Genes Using Transcriptomic and Proteomic Association Analyses of the Plant Parasitic Nematode Bursaphelenchus mucronatus. Int. J. Mol. Sci. 2016, 17, 1492. [Google Scholar] [CrossRef]
- Dua, V.K.; Pandey, A.C.; Raghavendra, K.; Gupta, A.; Sharma, T.; Dash, A.P. Larvicidal activity of neem oil (Azadirachta indica) formulation against mosquitoes. Malar. J. 2009, 8, 124. [Google Scholar] [CrossRef]
- Ayinde, A.; Morakinyo, O.; Sridhar, M. Repellency and larvicidal activities of Azadirachta indica seed oil on Anopheles gambiae in Nigeria. Heliyon 2020, 6, e03920. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, J.; Huang, Y.; Liu, Y.; Hu, G.; Yan, W.; Yan, G.; Guo, Q.; Shi, J.; Han, R.; et al. Sustainable pest management using plant secondary metabolites regulated azadirachtin nano-assemblies. Nat. Commun. 2025, 16, 1721. [Google Scholar] [CrossRef]
- Bae, M.; Lewis, A.; Liu, S.; Arcot, Y.; Lin, Y.-T.; Bernal, J.S.; Cisneros-Zevallos, L.; Akbulut, M. Novel Biopesticides Based on Nanoencapsulation of Azadirachtin with Whey Protein to Control Fall Armyworm. J. Agric. Food Chem. 2022, 70, 7900–7910. [Google Scholar] [CrossRef]
- Iqbal, N.; Hazra, D.K.; Purkait, A.; Agrawal, A.; Kumar, J. Bioengineering of neem nano-formulation with adjuvant for better adhesion over applied surface to give long term insect control. Colloids Surf. B Biointerfaces 2022, 209, 112176. [Google Scholar] [CrossRef] [PubMed]
- Nazir, N.; Fatima, S.; Aasim, M.; Yaqoob, F.; Mahmood, K.; Ali, S.A.; Awan, S.I.; Haq, I.U. Zeugodacus fruit flies (Diptera: Tephritidae) host preference analysis by machine learning-based approaches. Comput. Electron. Agric. 2024, 222, 109095. [Google Scholar] [CrossRef]
- Dai, M.; Yang, J.; Liu, X.; Gu, H.; Li, F.; Li, B.; Wei, J. Parasitism by the Tachinid Parasitoid Exorista japonica Leads to Suppression of Basal Metabolism and Activation of Immune Response in the Host Bombyx mori. Insects 2022, 13, 792. [Google Scholar] [CrossRef] [PubMed]
- Eggleston, H.; Adelman, Z.N. Transcriptomic analyses of Aedes aegypti cultured cells and ex vivo midguts in response to an excess or deficiency of heme: A quest for transcriptionally-regulated heme transporters. BMC Genom. 2020, 21, 604. [Google Scholar] [CrossRef]
- Marshall, K.L.; Moran, C.; Chen, Y.; Herron, G.A. Detection of Kdr Pyre-throid Resistance in the Cotton Aphid, Aphis Gossypii (Hemiptera: Aphididae), Using a Pcr-Rflp Assay. J. Pestic. Sci. 2012, 37, 169–172. [Google Scholar] [CrossRef]
- Lixiang, C.; Zhenya, T.; Weihua, M.; Jingjing, W.; Qiaofen, H.; Yongping, Z.; Xuyuan, G.; Hongsong, C.; Zhongshi, Z. Comparison of bacterial diversity in Bactrocera cucurbitae (Coquillett) ovaries and eggs based on 16S rRNA sequencing. Sci. Rep. 2023, 13, 1–15. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. Clusterprofiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Li, J.; Fu, A.; Zhang, L. An Overview of Scoring Functions Used for Protein–Ligand Interactions in Molecular Docking. Interdiscip. Sci. Comput. Life Sci. 2019, 11, 320–328. [Google Scholar] [CrossRef]
- Elhenawy, A.A.; Al-Harbi, L.M.; El-Gazzar, M.A.; Khowdiary, M.M.; Alosaimi, A.M.; elhamid Salim, A. Naproxenylamino Acid Derivatives: Design, Synthesis, Docking, Qsar and Anti-Inflammatory and Analgesic Activity. Biomed. Pharmacother. 2019, 116, 109024. [Google Scholar] [CrossRef]
- Wu, B.; Lan, X.; Chen, X.; Wu, Q.; Yang, Y.; Wang, Y. Researching the molecular mechanisms of Taohong Siwu Decoction in the treatment of varicocele-associated male infertility using network pharmacology and molecular docking: A review. Medicine 2023, 102, e34476. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | ID | CAS | Host | References |
|---|---|---|---|---|
| cinncassiol E | C1 | L. acutangular | [35] | |
| hymenoxon | C2 | 57377-32-9 | L. acutangular | [35] |
| 1-(alpha-methyl-4-(2-methylpropyl)benzeneacetate)-beta-D-Glucopyranuronic acid | C3 | 115075-59-7 | L. acutangular | [35] |
| cinncassiol C3 | C4 | L. acutangular | [35] | |
| clerodin | C5 | 464-71-1 | L. acutangular | [35] |
| nigakilactone E | C6 | 28360-79-4 | L. acutangular | [35] |
| shogaol | C7 | 555-66-8 | L. acutangular | [35] |
| maritimetin | C8 | 576-02-3 | L. acutangular | [35] |
| gartanin | C9 | 33390-42-0 | L. acutangular | [35] |
| rotenone | C10 | 83-79-4 | L. acutangular | [35] |
| apiin | C11 | 26544-34-3 | L. acutangular | [35] |
| galangin | C12 | 548-83-4 | L. acutangular | [35] |
| hispidulin | C13 | 1447-88-7 | L. acutangular | [35] |
| berbamunine | C14 | 485-18-7 | L. acutangular | [35] |
| daphnoline | C15 | 479-36-7 | L. acutangular | [35] |
| ellagic acid | C16 | 476-66-4 | L. acutangular, L. cylindrica | [36,37] |
| chlorogenic acid | C17 | 202650-88-2 | L. acutangular, L. cylindrica, M. nana, B. oleracea | [36,38,39,40] |
| sinapaldehyde | C18 | 4206-58-0 | L. acutangular | [36] |
| mandelic acid | C19 | 90-64-2 | L. acutangular | [36] |
| scopoletin | C20 | 92-61-5 | L. acutangular | [36] |
| salicylic acid | C21 | 69-72-7 | L. acutangular, L. cylindrica, M. nana, B. oleracea | [36,38,39,41] |
| gallic acid | C22 | 149-91-7 | L. acutangular, L. cylindrica, M. nana, B. oleracea | [36,38,39,40] |
| 4-hydroxycoumarin | C23 | 1076-38-6 | L. acutangular | [36] |
| phenylacetic acid | C24 | 103-82-2 | L. acutangular | [36] |
| guaiacol | C25 | 32994 | L. acutangular | [36] |
| 3,4-dihydroxyphenylacetic acid | C26 | 102-32-9 | L. acutangular | [36] |
| hydroxycaffeic acid | C27 | L. acutangular, L. cylindrica, B. oleracea, F. ananassa | [36,40,42,43] | |
| 2,5-dihydroxybenzoic acid | C28 | L. acutangular, L. cylindrica, M. nana | [36,38,39] | |
| 4-ethylguaiacol-d5 | C29 | L. acutangular | [36] | |
| 3-(4-hydroxy-3-methoxyphenyl)propionic acid | C30 | 1135-23-5 | L. acutangular | [36] |
| caffeic acid | C31 | 501-16-6 | L. acutangular, M. nana | [36,39] |
| pinocembrin | C32 | 480-39-7 | L. acutangular | [36] |
| benzoic acid | C33 | 65-85-0 | L. acutangular | [36] |
| cirsimaritin | C34 | 6601-62-3 | L. acutangular | [36] |
| eugenol | C35 | 97-53-0 | L. acutangular | [36] |
| p-coumaric acid | C36 | 501-98-4 | L. acutangular, L. cylindrica, M. nana, B. oleracea, F. ananassa | [36,38,39,40,43] |
| sakuranetin | C37 | 2957-21-3 | L. acutangular | [36] |
| gardenin B | C38 | 2798-20-1 | L. acutangular | [36] |
| chrysin | C39 | 480-40-0 | L. acutangular, M. nana | [36,39] |
| scutellarein | C40 | 529-53-3 | L. acutangular | [36] |
| tetramethylscutellarein | C41 | 1168-42-9 | L. acutangular | [36] |
| mellein | C42 | 1200-93-7 | L. acutangular | [36] |
| geraldone | C43 | 21583-32-4 | L. acutangular | [36] |
| umbelliferone | C44 | 202-240-3 | L. acutangular | [44] |
| lucyoside A | C45 | L. acutangular | [44] | |
| lucyoside J | C46 | 100156-31-8 | L. acutangular | [44] |
| luteolin | C47 | 491-70-3 | L. acutangular, L. cylindrica, S. edule, M. nana | [39,44,45,46] |
| apigenin | C48 | 520-36-5 | L. acutangular, M. nana | [39,44] |
| diosmetin | C49 | 520-34-3 | L. acutangular, S. edule | [44] |
| lucyoside G | C50 | L. acutangular | [44] | |
| 13-trihydroxy-octadecenoic acid | C51 | L. acutangular | [44] | |
| lucyoside H | C52 | L. acutangular | [44] | |
| lucyoside I | C53 | 99543-11-0 | L. acutangular | [44] |
| Maslinic acid 3-O-b-D-glucoside | C54 | 163634-06-8 | L. acutangular | [44] |
| lucyin A | C55 | 152845-76-6 | L. acutangular | [44] |
| 4-hydroxybenzoic acid | C56 | 99-96-7 | L. cylindrica | [38] |
| catechol | C57 | 120-80-9 | L. cylindrica, B. oleracea | [38,40] |
| vanillic acid | C58 | 121-34-6 | L. cylindrica, M. nana, B. oleracea | [38,39,40] |
| rutin | C59 | 153-18-4 | L. cylindrica, M. nana, B. oleracea | [38,39,40] |
| ferulic acid | C60 | 537-98-4 | L. cylindrica, M. nana, B. oleracea, F. ananassa | [38,39,40,43] |
| naringenin | C61 | 480-41-1 | L. cylindrica, M. nana, B. oleracea | [38,39,40] |
| 2-hydroxy-4-methylbenzaldehyde | C62 | 698-27-1 | L. cylindrica | [47] |
| 4-acetoxy-2-azetidinone | C63 | 28562-53-0 | L. cylindrica | [47] |
| mahaleboside | C64 | L. cylindrica | [47] | |
| crotanecine | C65 | 5096-50-4 | L. cylindrica | [47] |
| perlolyrine | C66 | 29700-20-7 | L. cylindrica | [47] |
| dihydrocapsaicin | C67 | 19408-84-5 | L. cylindrica | [47] |
| morindone | C68 | 478-29-5 | L. cylindrica | [47] |
| 4-aminosalicylic acid | C69 | 65-49-6 | L. cylindrica | [38] |
| apigenin 7-glucuronide | C70 | 29741-09-1 | L. cylindrica | [48] |
| kaempferide | C71 | 491-54-3 | L. cylindrica, | [49] |
| diosmin | C72 | 520-27-4 | L. cylindrica S. edule | [50,51] |
| 5-hydroxy-2-(3-hydroxy-4-methoxyphenyl)-4-oxo-4H-1-benzopyran-7-yl 2-O-(6-deoxy-alpha-L-mannopyranosyl)-beta-D-glucopyranoside | C73 | 38665-01-9 | L. cylindrica | [52] |
| eriodictyol-7-O-glucoside | C74 | 38965-51-4 | L. cylindrica | [52] |
| quercetin | C75 | 117-39-5 | L. cylindrica, M. nana, B. oleracea | [39,40,53] |
| myricetin | C76 | 529-44-2 | L. cylindrica | [54] |
| cianidanol | C77 | 7295-85-4 | L. cylindrica, M. nana, B. oleracea | [39,40,55] |
| hyperoside | C78 | 482-36-0 | L. cylindrica | [56] |
| lespedin | C79 | 482-38-2 | L. cylindrica | [57] |
| quercitrin | C80 | 522-12-3 | L. cylindrica, M. nana | [39,58] |
| tiliroside | C81 | 20316-62-5 | L. cylindrica | [59] |
| acacetin | C82 | 480-44-4 | L. cylindrica, M. nana | [39,60] |
| saponarin | C83 | 20310-89-8 | L. cylindrica | [60] |
| datiscin | C84 | L. cylindrica | [61] | |
| fortunellin | C85 | 20633-93-6 | L. cylindrica | [61] |
| linarin | C86 | 480-36-4 | L. cylindrica | [62] |
| vitexin | C87 | 3681-93-4 | L. cylindrica, S. edule | [51,63] |
| vitexin 2″-O-rhamnoside | C88 | 64820-99-1 | L. cylindrica | [64] |
| 2-hydroxycinnamic acid,(2E)- | C89 | 614-60-8 | L. cylindrica, M. nana | [39,64] |
| oleanolic acid | C90 | 508-02-1 | L. cylindrica | [65] |
| echinocystic acid | C91 | 510-30-5 | L. cylindrica | [66] |
| gypsogenin | C92 | 639-14-5 | L. cylindrica | [67] |
| 3-O-[beta-D-glucopyranosyl]-28-O-[alpha-L-rhamnopyranosyl-(1->2)-beta-D-glucopyranosyl]maslinic acid | C93 | 1268696-94-1 | L. cylindrica | [68] |
| luteolin 7-rutinoside | C94 | 20633-84-5 | S. edule | [51] |
| luteolin 7-O-glucoside | C95 | 1268798 | S. edule, M. nana | [39,51] |
| isorhoifolin | C96 | 552-57-8 | S. edule | [51] |
| leucoside | C97 | S. edule | [46] | |
| myricitrin | C98 | 17912-87-7 | S. edule | [46] |
| vicenin 2 | C99 | 23666-13-9 | S. edule | [46] |
| all-trans-vaucheriaxanthin | C100 | M. nana | [69] | |
| neochrome | C101 | M. nana | [69] | |
| neoxanthin | C102 | 14660-91-4 | M. nana | [69] |
| 5,8:5′,8′-diepoxy-5,8,5′,8′-tetrahydro-beta,beta-carotene-3,3′-diol | C103 | M. nana | [69] | |
| violaxanthin | C104 | 126-29-4 | M. nana | [69] |
| lutein 5,6-epoxide | C105 | 28368-08-3 | M. nana | [69] |
| lutein | C106 | 127-40-2 | M. nana | [69] |
| fumaric acid | C107 | 110-17-8 | M. nana | [39] |
| cis-aconitic acid | C108 | 585-84-2 | M. nana | [39] |
| epigallocatechin | C109 | 970-74-1 | M. nana | [39] |
| 3,4-dihydroxybenzoic acid | C110 | 99-50-3 | M. nana, B. oleracea | [39,40] |
| protocatechualdehyde | C111 | 139-85-5 | M. nana | [39] |
| epigallocatechin gallate | C112 | 989-51-5 | M. nana | [39] |
| 1,5-dicaffeoylquinic acid | C113 | 30964-13-7 | M. nana | [39] |
| 4-carboxyphenylglycine | C114 | 7292-81-1 | M. nana | [39] |
| syringic acid | C115 | 530-57-4 | M. nana, B. oleracea, F. ananassa | [39,40,43] |
| vanillin | C116 | 121-33-5 | M. nana | [39] |
| syringaldehyde | C117 | 134-96-3 | M. nana | [39] |
| daidzin | C118 | 552-66-9 | M. nana | [39] |
| (-)-epicatechingallate | C119 | 25615-05-8 | M. nana | [39] |
| piceid | C120 | 27208-80-6 | M. nana | [39] |
| sinapinic acid | C121 | 7362-37-0 | M. nana, B. oleracea, F. ananassa | [39,40,43] |
| coumarin | C122 | 91-64-5 | M. nana | [39] |
| querciturone | C123 | 22688-79-5 | M. nana | [39] |
| isoquercetin | C124 | 482-35-9 | M. nana | [39] |
| hesperidin | C125 | 520-26-3 | M. nana | [39] |
| genistin | C126 | 529-59-9 | M. nana | [39] |
| rosmarinic acid | C127 | 20283-92-5 | M. nana, B. oleracea | [39,40] |
| apigetrin | C128 | 578-74-5 | M. nana | [39] |
| astragalin | C129 | 480-10-4 | M. nana | [39] |
| kaempferol-3-O-rutinoside | C130 | 17650-84-9 | M. nana, B. oleracea | [39,40] |
| fisetin | C131 | 528-48-3 | M. nana | [39] |
| daidzein | C132 | 486-66-8 | M. nana | [39] |
| quercetin 3′-isobutyrate | C133 | M. nana | [39] | |
| hesperetin | C134 | 520-33-2 | M. nana | [39] |
| genistein | C135 | 446-72-0 | M. nana | [39] |
| kaempferol | C136 | 520-18-3 | M. nana | [39] |
| amentoflavone | C137 | 1617-53-4 | M. nana | [39] |
| cinnamic acid | C138 | 140-10-3 | B. oleracea, F. ananassa | [40,43] |
| lactoyl isovanillic acid | C139 | B. oleracea | [40] | |
| isorhamnetin | C140 | 480-19-3 | B. oleracea | [40] |
| 3-hydroxyflavone | C141 | 577-85-5 | B. oleracea | [40] |
| pyrogallol | C142 | 87-66-1 | B. oleracea | [40] |
| pyruvic acid | C143 | 127-17-3 | B. oleracea | [40] |
| lactic acid | C144 | 50-21-5 | B. oleracea | [40] |
| valine | C145 | 72-18-4 | B. oleracea | [40] |
| alanine | C146 | 56-41-7 | B. oleracea | [40] |
| glycolic acid | C147 | 79-14-1 | B. oleracea | [40] |
| linalool, (+/−)- | C148 | 78-70-6 | F. ananassa | [70] |
| beta-Farnesene | C149 | 18794-84-8 | F. ananassa | [70] |
| alpha-Terpineol | C150 | 98-55-5 | F. ananassa | [70] |
| damascenone | C151 | 23726-93-4 | F. ananassa | [70] |
| trans-Nerolidol | C152 | 35944-21-9 | F. ananassa | [70] |
| 2,5-dimethyl-4-methoxy-3(2H)-furanone | C153 | 4077-47-8 | F. ananassa | [70] |
| furaneol | C154 | 3658-77-3 | F. ananassa | [70] |
| (E)-5-(3-hexenyl)dihydrofuran-2(3H)-one | C155 | 97416-87-0 | F. ananassa | [70] |
| gamma-Octalactone | C156 | 104-50-7 | F. ananassa | [70] |
| gamma-Decalactone | C157 | 706-14-9 | F. ananassa | [70] |
| gamma-Dodecalactone | C158 | 148051 | F. ananassa | [70] |
| acetophenone | C159 | 202-708-7 | F. ananassa | [70] |
| 3,6-Octadienal,3,7-dimethyl- | C160 | 1754-00-3 | F. ananassa | [43] |
| (Z)-3,7-dimethylocta-3,6-dienal | C161 | 72203-97-5 | F. ananassa | [43] |
| citral | C162 | C10H16O | F. ananassa | [43] |
| nerylacetone | C163 | 3879-26-3 | F. ananassa | [43] |
| nizatidine | C164 | 76963-41-2 | F. ananassa | [43] |
| beta-caryophyllene oxide | C165 | 1139-30-6 | F. ananassa | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Chen, Y.; Wang, Y.; Fu, X.; Jin, S.; Yi, C.; Bai, X.; Lu, Y.; Miao, W.; Geng, X.; et al. The Discovery of Potential Repellent Compounds for Zeugodacus cucuribitae (Coquillett) from Six Non-Favored Hosts. Int. J. Mol. Sci. 2025, 26, 6556. https://doi.org/10.3390/ijms26146556
Fu Y, Chen Y, Wang Y, Fu X, Jin S, Yi C, Bai X, Lu Y, Miao W, Geng X, et al. The Discovery of Potential Repellent Compounds for Zeugodacus cucuribitae (Coquillett) from Six Non-Favored Hosts. International Journal of Molecular Sciences. 2025; 26(14):6556. https://doi.org/10.3390/ijms26146556
Chicago/Turabian StyleFu, Yu, Yupeng Chen, Yani Wang, Xinyi Fu, Shunda Jin, Chunyan Yi, Xue Bai, Youqing Lu, Wang Miao, Xingyu Geng, and et al. 2025. "The Discovery of Potential Repellent Compounds for Zeugodacus cucuribitae (Coquillett) from Six Non-Favored Hosts" International Journal of Molecular Sciences 26, no. 14: 6556. https://doi.org/10.3390/ijms26146556
APA StyleFu, Y., Chen, Y., Wang, Y., Fu, X., Jin, S., Yi, C., Bai, X., Lu, Y., Miao, W., Geng, X., Lu, X., Yan, R., Zhou, Z., & Cao, F. (2025). The Discovery of Potential Repellent Compounds for Zeugodacus cucuribitae (Coquillett) from Six Non-Favored Hosts. International Journal of Molecular Sciences, 26(14), 6556. https://doi.org/10.3390/ijms26146556