

Serotonin N-acetyltransferase SlSNAT2 Positively Regulates Tomato Resistance Against Ralstonia solanacearum



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Bioinformatics Analysis of SlSNAT2
2.2. Transcriptional Analysis of SlSNAT2 in Tomatoes
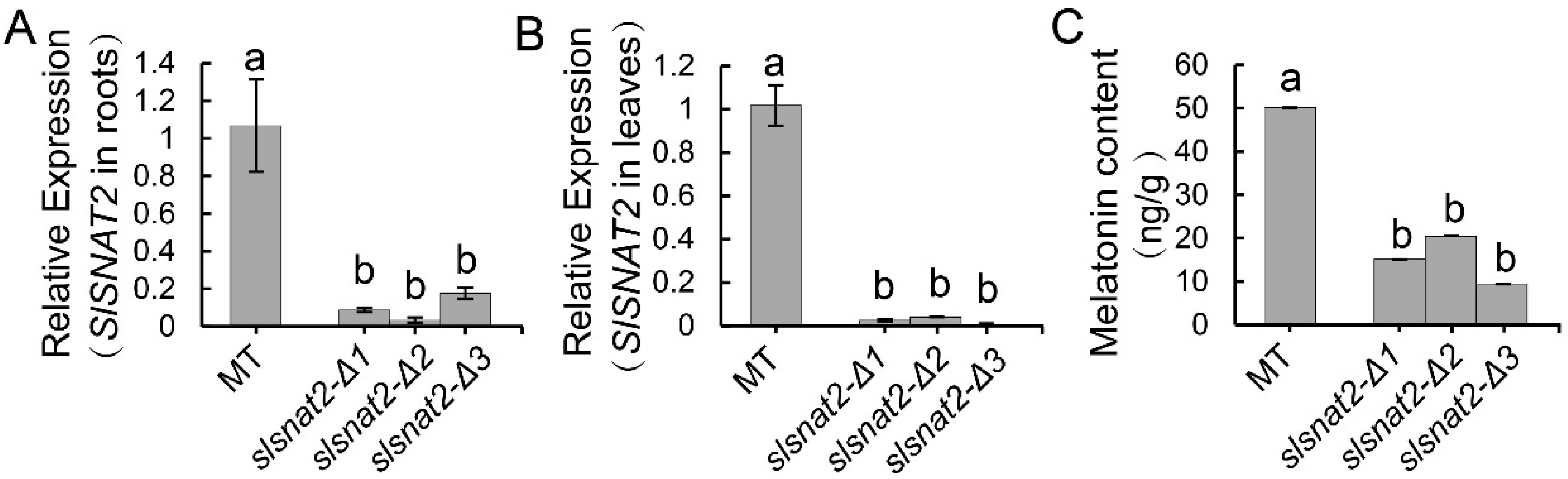
2.3. Knocking Out SlSNAT2 in Tomatoes Results in a Significant Decrease in Melatonin Levels
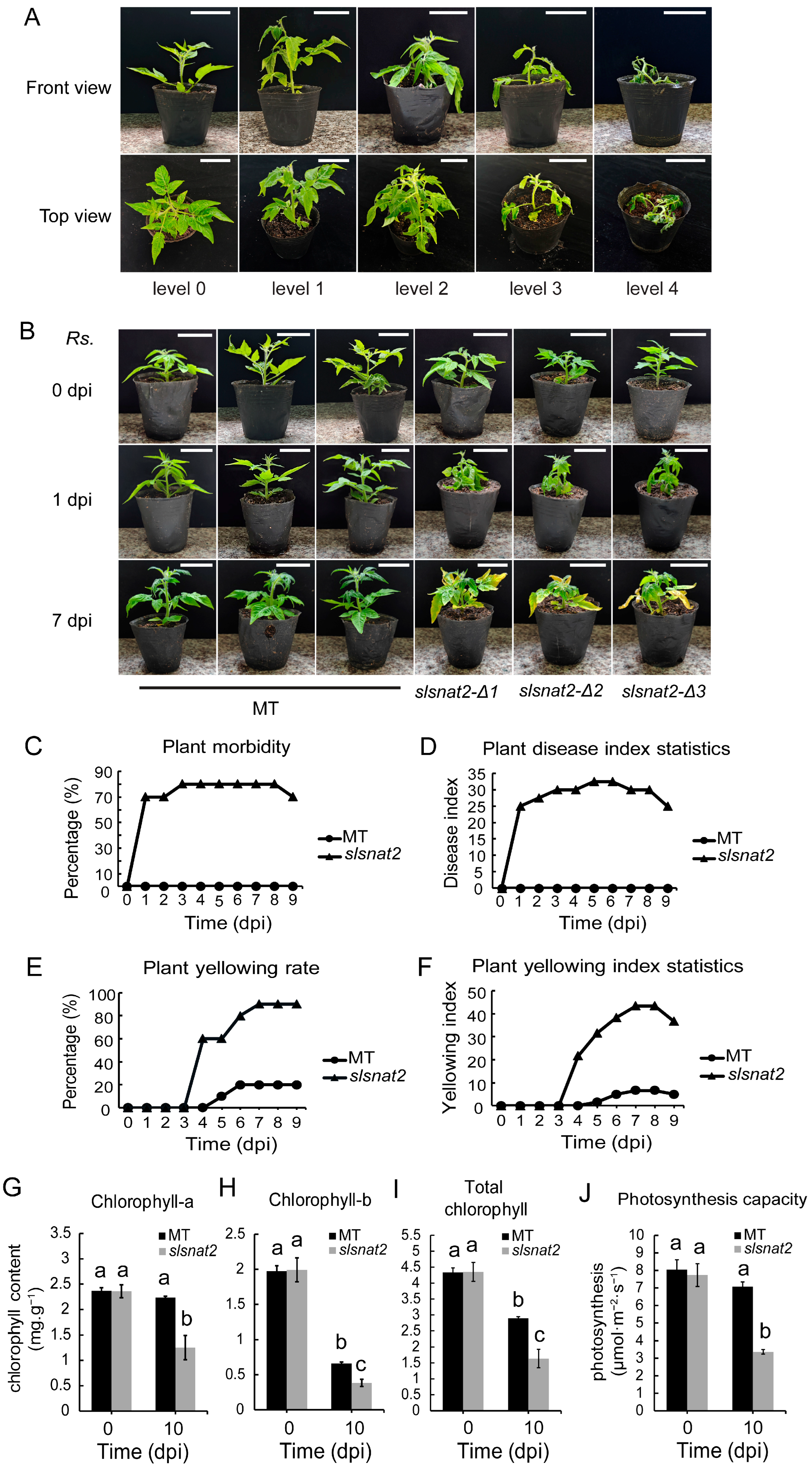
2.4. The slsnat2 Mutant Tomatoes Exhibited a Reduced Resistance to Bacterial Wilt
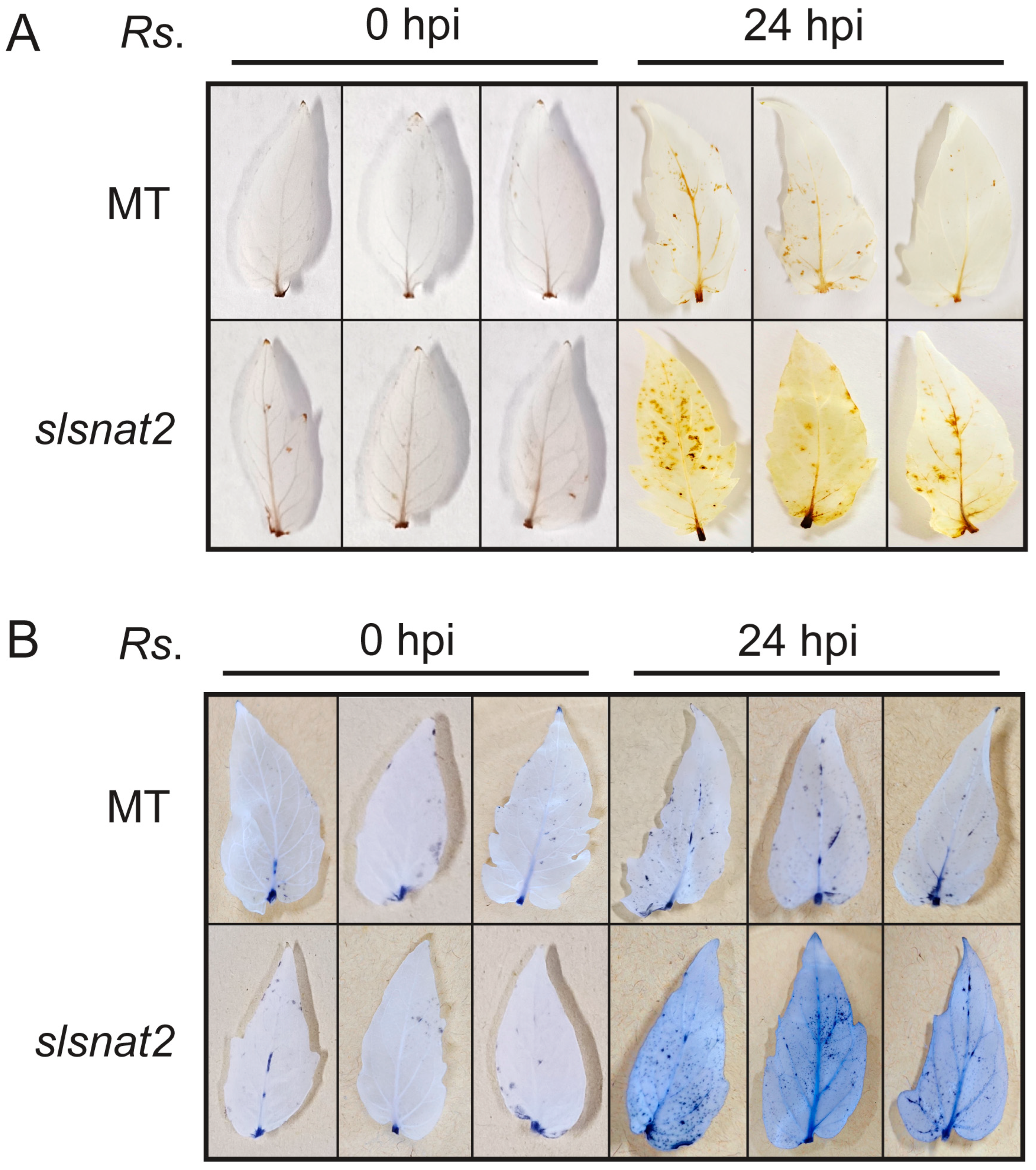
2.5. The slsnat2 Mutants Exhibit Enhanced R. solanacearum-Induced Reactive Oxygen Species Accumulation
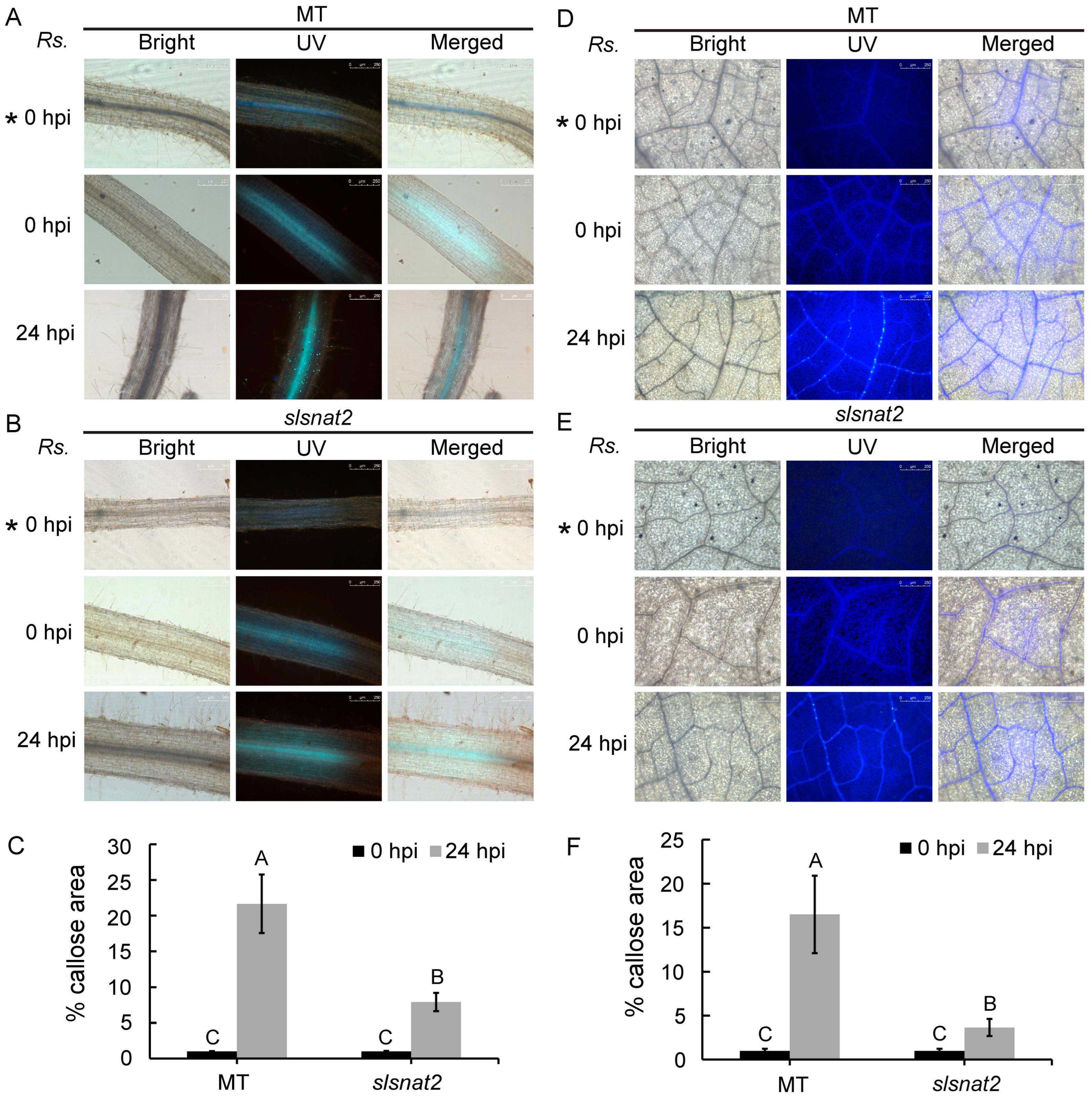
2.6. Suppression of Prominent Callose Accumulation in Root and Leaf Cells by Knocking Out SlSNAT2
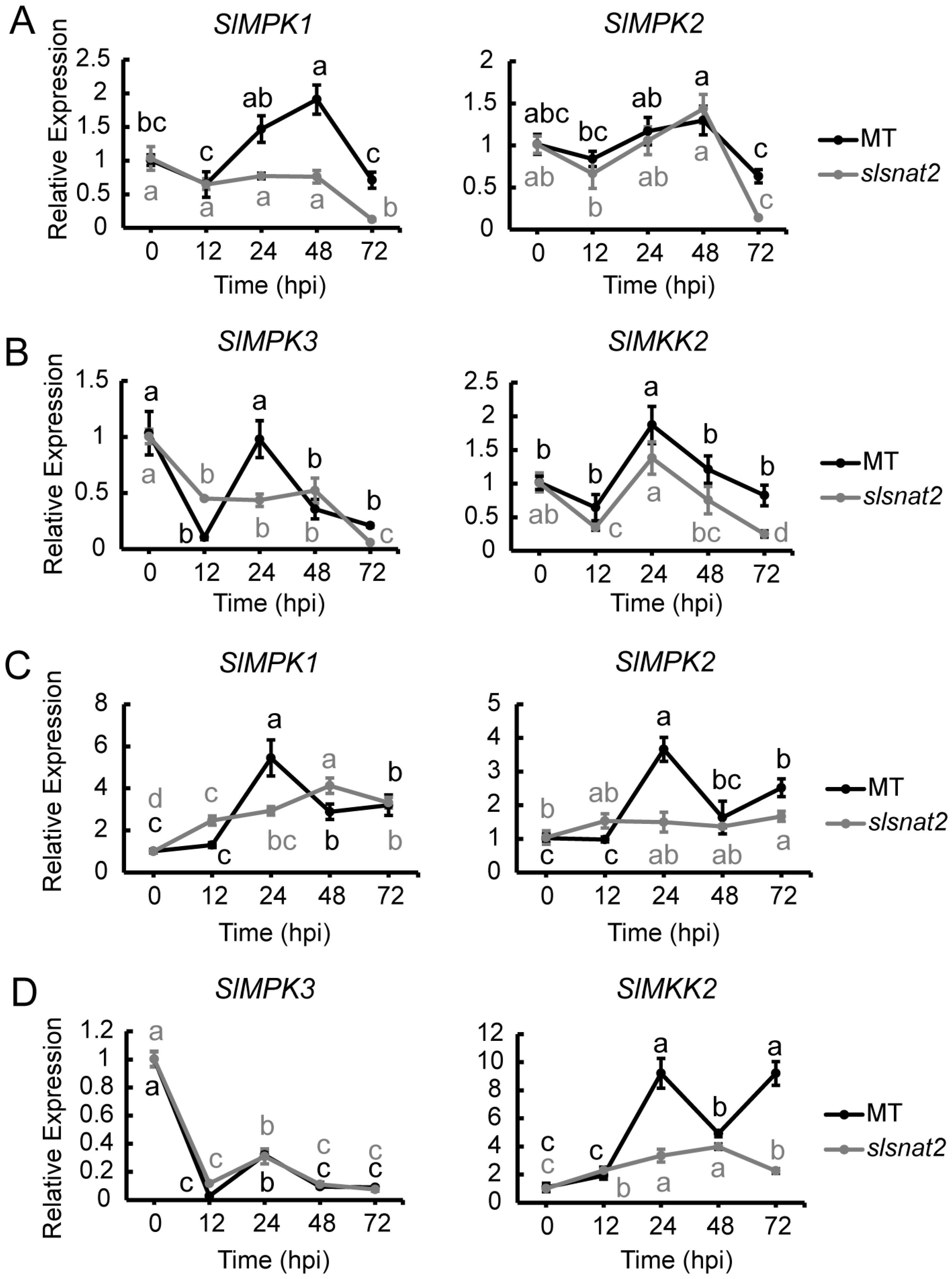
2.7. Expression Analysis of MAPK Pathway-Related Genes in Roots and Leaves of MT and slsnat2 Plants Within 72 h Post-Inoculation with R. solanacearum
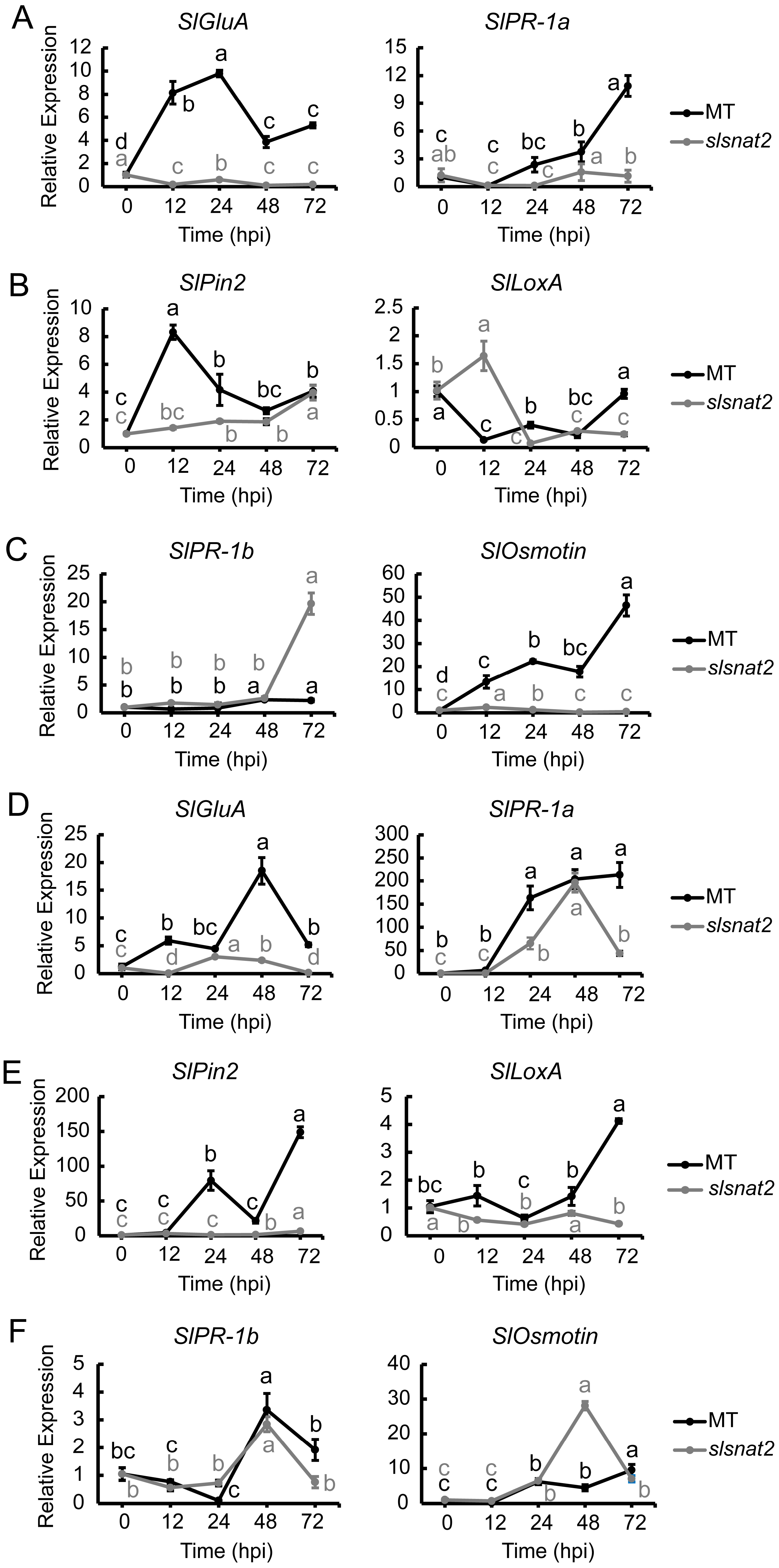
2.8. Expression Analysis of Salicylic Acid−, Jasmonic Acid, and Ethylene Pathway-Related Defense Genes in Roots and Leaves of MT and slsnat2 Plants Inoculated for 72 h
2.9. Expression Analysis of SlWRKY30 and SlWRKY81 in Roots and Leaves of MT and slsnat2 Plants Within 72 h Post-Inoculation with R. solanacearum
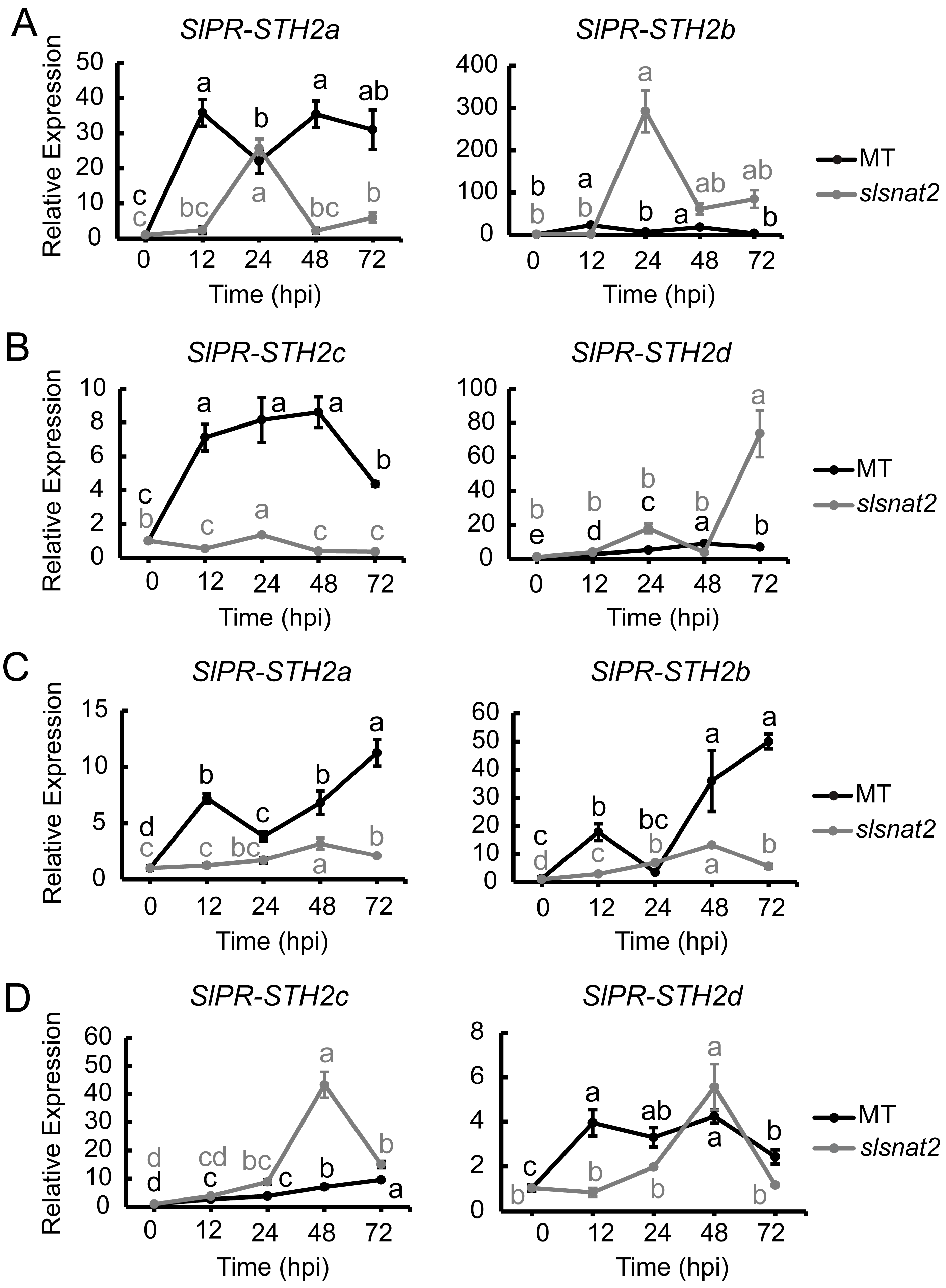
2.10. Expression Analysis of SlPR-STHs in Roots and Leaves of MT and slsnat2 Plants Within 72 h Post-Inoculation with R. solanacearum
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain, Plant Materials, and Growth Condition
4.2. Bioinformatics Analysis
4.3. Gene Expression Analysis by Quantitative Real-Time PCR (RT-qPCR)
4.4. Endogenous Melatonin Measurement
4.5. Ralstonia solanacearum Inoculation and Disease Index Calculation
4.6. Statistics of Yellowing Index
4.7. Determination of Chlorophyll Content and Net Photosynthetic Rate (Pn)
4.8. Histochemical Staining of Reactive Oxygen Species
4.9. Staining and Quantification of Callose Deposition
4.10. The Expression Levels of Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASMT | N-acetyl serotonin methyltransferase |
| aa. | amino acid |
| BW | Bacterial wilt |
| COMT | caffeic acid O-methyltransferase |
| DAB | 3,3′-diaminobenzidine |
| dpi | day post inoculation |
| Eth | ethylene |
| hpi | hour post inoculation |
| JA | jasmonic acid |
| MT | Micro-Tom |
| MAPK | mitogen-activated protein kinase |
| NBT | nitroblue tetrazolium |
| NO | nitric oxide |
| POD | peroxidase |
| PR-STH2 | pathogenesis-related salt tolerance homolog 2 |
| ROS | reactive oxygen species |
| SA | salicylic acid |
| SNAT | serotonin N-acetyltransferase |
| SOD | superoxide dismutase |
| T5H | tryptamine 5-hydroxylase |
| TDC | tryptophan decarboxylase |
References
- Kim, B.-S.; French, E.; Caldwell, D.; Harrington, E.J.; Iyer-Pascuzzi, A.S. Bacterial wilt disease: Host resistance and pathogen virulence mechanisms. Physiol. Mol. Plant Pathol. 2016, 95, 37–43. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- McGarvey, J.; Denny, T.; Schell, M. Spatial-temporal and quantitative analysis of growth and EPS I production by Ralstonia solanacearum in resistant and susceptible tomato cultivars. Phytopathology 1999, 89, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Rajan, A.; Kumar, S.; Sunil, C.K.; Radhakrishnan, M.; Rawson, A. Recent advances in the utilization of industrial byproducts and wastes generated at different stages of tomato processing: Status report. J. Food Process. Preserv. 2022, 46, e17063. [Google Scholar] [CrossRef]
- Shao, D.; Venkitasamy, C.; Li, X.; Pan, Z.; Shi, J.; Wang, B.; Teh, H.E.; McHugh, T. Thermal and storage characteristics of tomato seed oil. LWT-Food Sci. Technol. 2015, 63, 191–197. [Google Scholar] [CrossRef]
- Topal, U.; Sasaki, M.; Goto, M.; Hayakawa, K. Extraction of lycopene from tomato skin with supercritical carbon dioxide: Effect of operating conditions and solubility analysis. J. Agric. Food Chem. 2006, 54, 5604–5610. [Google Scholar] [CrossRef]
- Mohammed, A.F.; Oloyede, A.R.; Odeseye, A.O. Biological control of bacterial wilt of tomato caused by Ralstonia solanacearum using Pseudomonas species isolated from the rhizosphere of tomato plants. Arch. Phytopathol. Plant Prot. 2020, 53, 1–16. [Google Scholar] [CrossRef]
- Flores-Cruz, Z.; Allen, C. Ralstonia solanacearum encounters an oxidative environment during tomato infection. Mol. Plant-Microbe Interact. 2009, 22, 773–782. [Google Scholar] [CrossRef]
- Mandal, S.; Das, R.K.; Mishra, S. Differential occurrence of oxidative burst and antioxidative mechanism in compatible and incompatible interactions of Solanum lycopersicum and Ralstonia solanacearum. Plant Physiol. Biochem. 2011, 49, 117–123. [Google Scholar] [CrossRef]
- Shi, H.; Liu, Y.; Ding, A.; Wang, W.; Sun, Y. Induced defense strategies of plants against Ralstonia solanacearum. Front. Microbiol. 2023, 14, 1059799. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Dubbels, R.; Reiter, R.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Xiao, S.; Liu, L.; Wang, H.; Li, D.; Bai, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Li, C. Exogenous melatonin accelerates seed germination in cotton (Gossypium hirsutum L.). PLoS ONE 2019, 14, e0216575. [Google Scholar] [CrossRef]
- Chen, Z.; Gu, Q.; Yu, X.; Huang, L.; Xu, S.; Wang, R.; Shen, W.; Shen, W. Hydrogen peroxide acts downstream of melatonin to induce lateral root formation. Ann. Bot. 2018, 121, 1127–1136. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Shi, K.; Shan, D.; Zhu, Y.; Wang, C.; Bai, Y.; Yan, T.; Zheng, X.; Kong, J. Apple tree flowering is mediated by low level of melatonin under the regulation of seasonal light signal. J. Pineal Res. 2019, 66, e12551. [Google Scholar] [CrossRef]
- Tijero, V.; Munoz, P.; Munne-Bosch, S. Melatonin as an inhibitor of sweet cherries ripening in orchard trees. Plant Physiol. Biochem. 2019, 140, 88–95. [Google Scholar] [CrossRef]
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin enhances root regeneration, photosynthetic pigments, biomass, total carbohydrates and proline content in the cherry rootstock PHL-C (Prunus avium × Prunus cerasus). Plant Physiol. Biochem. 2012, 61, 162–168. [Google Scholar] [CrossRef]
- Qiao, Y.; Yin, L.; Wang, B.; Ke, Q.; Deng, X.; Wang, S. Melatonin promotes plant growth by increasing nitrogen uptake and assimilation under nitrogen deficient condition in winter wheat. Plant Physiol. Biochem. 2019, 139, 342–349. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin against environmental plant stressors: A review. Curr. Protein Pept. Sci. 2021, 22, 413–429. [Google Scholar] [CrossRef]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.; Lu, X.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Wu, X.; Zhang, Q.; Li, Y.; Ye, Y.; Li, P.; Chen, S.; Peng, Y.; Hardeland, R.; Xia, Y. Bacteriostatic potential of melatonin: Therapeutic standing and mechanistic insights. Front. Immunol. 2021, 12, 683879. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Byeon, Y.; Back, K. Melatonin as a signal molecule triggering defense responses against pathogen attack in Arabidopsis and tobacco. J. Pineal Res. 2014, 57, 262–268. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, L.; Su, T.; Jiang, Y.; Hu, L.; Ma, F. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC 3000 infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar]
- Lee, K.; Choi, G.H.; Back, K. Cadmium-induced melatonin synthesis in rice requires light, hydrogen peroxide, and nitric oxide: Key regulatory roles for tryptophan decarboxylase and caffeic acid O-methyltransferase. J. Pineal Res. 2017, 63, e12441. [Google Scholar] [CrossRef]
- Byeon, Y.; Back, K. Melatonin synthesis in rice seedlings in vivo is enhanced at high temperatures and under dark conditions due to increased serotonin N-acetyltransferase and N-acetylserotonin methyltransferase activities. J. Pineal Res. 2014, 56, 189–195. [Google Scholar] [CrossRef]
- Park, S.; Lee, K.; Kim, Y.S.; Back, K. Tryptamine 5-hydroxylase-deficient Sekiguchi rice induces synthesis of 5-hydroxytryptophan and N-acetyltryptamine but decreases melatonin biosynthesis during senescence process of detached leaves. J. Pineal Res. 2012, 52, 211–216. [Google Scholar] [CrossRef]
- Lee, H.Y.; Byeon, Y.; Tan, D.X.; Reiter, R.J.; Back, K. Arabidopsis serotonin N-acetyltransferase knockout mutant plants exhibit decreased melatonin and salicylic acid levels resulting in susceptibility to an avirulent pathogen. J. Pineal Res. 2015, 58, 291–299. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, M.-J.; Song, J.-B.; Zhang, S.-Y.; Guo, R.; Hou, D.-R.; Hao, C.-Y.; An, H.-L.; Huang, X. Roles of endogenous melatonin in resistance to Botrytis cinerea infection in an Arabidopsis model. Front. Plant Sci. 2021, 12, 683228. [Google Scholar] [CrossRef]
- Yu, Y.; Bian, L.; Jiao, Z.; Yu, K.; Wan, Y.; Zhang, G.; Guo, D. Molecular cloning and characterization of a grapevine (Vitis vinifera L.) serotonin N-acetyltransferase (VvSNAT2) gene involved in plant defense. BMC Genom. 2019, 20, 880. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Lin, Y.M.; Chao, T.C.; Wang, J.F.; Liu, A.C.; Ho, F.I.; Cheng, C.P. Virus-induced gene silencing reveals the involvement of ethylene-, salicylic acid-and mitogen-activated protein kinase-related defense pathways in the resistance of tomato to bacterial wilt. Physiol. Plant. 2009, 136, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Lin, J.; Li, Y.; Jiang, R.; Fang, Y.; Ding, F.; He, S.; Wang, Y. SlWRKY30 and SlWRKY81 synergistically modulate tomato immunity to Ralstonia solanacearum by directly regulating SlPR-STH2. Hortic. Res. 2023, 10, uhad050. [Google Scholar] [CrossRef]
- Atta, B.M.; Saleem, M.; Ali, H.; Arshad, H.M.I.; Ahmed, M. Chlorophyll as a biomarker for early disease diagnosis. Laser Phys. 2018, 28, 065607. [Google Scholar] [CrossRef]
- Sun, Y.; Gui, Z.; Yan, N.; Wang, Q.; Zhang, Z.; Zhang, H.; Sun, F.; Han, X.; Du, Y. Roles and preliminary mechanism of tobacco cis-abienol in inducing tomato resistance against bacterial wilt. Int. J. Mol. Sci. 2023, 24, 12226. [Google Scholar] [CrossRef]
- Yan, S.; Wang, Y.; Yu, B.; Gan, Y.; Lei, J.; Chen, C.; Zhu, Z.; Qiu, Z.; Cao, B. A putative E3 ubiquitin ligase substrate receptor degrades transcription factor SmNAC to enhance bacterial wilt resistance in eggplant. Hortic. Res. 2024, 11, uhad246. [Google Scholar] [CrossRef]
- Milling, A.; Babujee, L.; Allen, C. Ralstonia solanacearum extracellular polysaccharide is a specific elicitor of defense responses in wilt-resistant tomato plants. PLoS ONE 2011, 6, e15853. [Google Scholar] [CrossRef]
- Liu, C.; Chen, L.; Zhao, R.; Li, R.; Zhang, S.; Yu, W.; Sheng, J.; Shen, L. Melatonin induces disease resistance to Botrytis cinerea in tomato fruit by activating jasmonic acid signaling pathway. J. Agric. Food Chem. 2019, 67, 6116–6124. [Google Scholar] [CrossRef]
- Xia, Z.; Ye, Y.; Hu, C.; Wang, H.; Zheng, L.; Hu, Y.; Sheng, L.; Xing, J.; Jia, W.; Wang, Y. Exogenous melatonin orchestrates multiple defense responses against Botrytis cinerea in tomato leaves. Plant Stress 2024, 11, 100326. [Google Scholar] [CrossRef]
- Li, C.; He, Q.; Zhang, F.; Yu, J.; Li, C.; Zhao, T.; Zhang, Y.; Xie, Q.; Su, B.; Mei, L. Melatonin enhances cotton immunity to Verticillium wilt via manipulating lignin and gossypol biosynthesis. Plant J. 2019, 100, 784–800. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Almoneafy, A.; Mahmoud, A.; Elkelish, A.; Arnao, M.B.; Li, L.; Ai, S. Melatonin and its protective role against biotic stress impacts on plants. Biomolecules 2019, 10, 54. [Google Scholar] [CrossRef]
- Chen, L.; Wang, M.R.; Li, J.W.; Feng, C.H.; Cui, Z.H.; Zhao, L.; Wang, Q.C. Exogenous application of melatonin improves eradication of apple stem grooving virus from the infected in vitro shoots by shoot tip culture. Plant Pathol. 2019, 68, 997–1006. [Google Scholar] [CrossRef]
- Lu, R.; Liu, Z.; Shao, Y.; Sun, F.; Zhang, Y.; Cui, J.; Zhou, Y.; Shen, W.; Zhou, T. Melatonin is responsible for rice resistance to rice stripe virus infection through a nitric oxide-dependent pathway. Virol. J. 2019, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Refaey, E.E.; Mohamed, H.I.; El-Dougdoug, N.K. Molecular characterization of the Alfalfa mosaic virus infecting Solanum melongena in Egypt and the control of its deleterious effects with melatonin and salicylic acid. Plants 2021, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Zhao, T.; Wang, Y.; Lei, N.; Liu, K.; Yu, H.; Zhang, Y.; Muhammad, U.; Ullah, H.; Yu, X. Melatonin and copper oxide nanoparticles synergistically mitigate clubroot disease and enhance growth dynamics in Brassica rapa. Plant Physiol. Biochem. 2024, 215, 109020. [Google Scholar] [CrossRef]
- Liu, Q.; Atta, U.R.; Wang, R.; Liu, K.; Ma, X.; Weng, Q. Defense-related hormone signaling coordinately controls the role of melatonin during Arabidopsis thaliana-Pseudomonas syringae interaction. Eur. J. Plant Pathol. 2021, 160, 707–716. [Google Scholar] [CrossRef]
- Arnao, M.B.; Giraldo-Acosta, M.; Castejón-Castillejo, A.; Losada-Lorán, M.; Sánchez-Herrerías, P.; El Mihyaoui, A.; Cano, A.; Hernández-Ruiz, J. Melatonin from microorganisms, algae, and plants as possible alternatives to synthetic melatonin. Metabolites 2023, 13, 72. [Google Scholar] [CrossRef]
- Lee, H.Y.; Lee, K.; Back, K. Knockout of Arabidopsis serotonin N-acetyltransferase-2 reduces melatonin levels and delays flowering. Biomolecules 2019, 9, 712. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin induction and its role in high light stress tolerance in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12504. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin plays a pivotal role in conferring tolerance against endoplasmic reticulum stress via mitogen-activated protein kinases and bZIP60 in Arabidopsis thaliana. Melatonin Res. 2018, 1, 94–108. [Google Scholar] [CrossRef]
- Li, D.; Guo, Y.; Zhang, D.; He, S.; Gong, J.; Ma, H.; Gao, X.; Wang, Z.; Jiang, L.; Dun, X. Melatonin represses oil and anthocyanin accumulation in seeds. Plant Physiol. 2020, 183, 898–914. [Google Scholar] [CrossRef]
- Wang, L.; Li, T.; Zhang, Y.; Guo, J.; Lu, K.; Liu, W. CAND2/PMTR1 is required for melatonin-conferred osmotic stress tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 4014. [Google Scholar] [CrossRef] [PubMed]
- Yildiztugay, E.; Abdulveli, B.A.; Ozfidan-Konakci, C.; Turkan, I. Melatonin mediated tolerance to benzalkonium chloride phytotoxicity through improved growth, photochemical reactions, and antioxidant system in wild-type and snat2 mutant Arabidopsis lines. Plant Physiol. Biochem. 2024, 212, 108779. [Google Scholar] [CrossRef] [PubMed]
- BA, C.A.; Clarke, A. Chemistry and biology of (1,3)-b-Glucans. J. Sci. Food Agric. 1992, 15, 33–36. [Google Scholar]
- Wang, Y.; Li, X.; Fan, B.; Zhu, C.; Chen, Z. Regulation and function of defense-related callose deposition in plants. Int. J. Mol. Sci. 2021, 22, 2393. [Google Scholar] [CrossRef]
- Khasin, M.; Bernhardson, L.F.; O’Neill, P.M.; Palmer, N.A.; Scully, E.D.; Sattler, S.E.; Funnell-Harris, D.L. Pathogen and drought stress affect cell wall and phytohormone signaling to shape host responses in a sorghum COMT bmr12 mutant. BMC Plant Biol. 2021, 21, 391. [Google Scholar] [CrossRef]
- Kohler, A.; Schwindling, S.; Conrath, U. Benzothiadiazole-induced priming for potentiated responses to pathogen infection, wounding, and infiltration of water into leaves requires the NPR1/NIM1 gene in Arabidopsis. Plant Physiol. 2002, 128, 1046–1056. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant-Microbe Interact. 2011, 24, 183–193. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Zhang, Y.; Lai, X.; Yang, S.; Ren, H.; Yuan, J.; Jin, H.; Shi, C.; Lai, Z.; Xia, G. Functional analysis of tomato CHIP ubiquitin E3 ligase in heat tolerance. Sci. Rep. 2021, 11, 1713. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Qiu, Z.; Yan, S.; Xia, B.; Jiang, J.; Yu, B.; Lei, J.; Chen, C.; Chen, L.; Yang, Y.; Wang, Y. The eggplant transcription factor MYB44 enhances resistance to bacterial wilt by activating the expression of spermidine synthase. J. Exp. Bot. 2019, 70, 5343–5354. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, Y.; Liu, B.; Zhou, Y.; Che, J.; Lin, N. Relationship between Ralstonia solanacearum diversity and severity of bacterial wilt disease in tomato fields in China. J. Phytopathol. 2014, 162, 607–616. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Ding, T.; Chu, D. Synergistic Effects of a Tomato chlorosis virus and Tomato yellow leaf curl virus Mixed Infection on Host Tomato Plants and the Whitefly Vector. Front. Plant Sci. 2021, 12, 672400. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, J.; Li, R.; Tian, Q.; Wu, Y.; Yu, J. Alleviative effect of exogenous ALA on salt stress in tomato seedlings. Chin. Melons Veg. 2023, 36, 56–64. [Google Scholar]
- Mason, K.N.; Ekanayake, G.; Heese, A. Staining and automated image quantification of callose in Arabidopsis cotyledons and leaves. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 160, pp. 181–199. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xia, G.; Xie, X.; Wang, H.; Zheng, L.; He, Z.; Ye, J.; Xu, K.; Shi, Q.; Yang, H.; et al. Serotonin N-acetyltransferase SlSNAT2 Positively Regulates Tomato Resistance Against Ralstonia solanacearum. Int. J. Mol. Sci. 2025, 26, 6530. https://doi.org/10.3390/ijms26136530
Wang Y, Xia G, Xie X, Wang H, Zheng L, He Z, Ye J, Xu K, Shi Q, Yang H, et al. Serotonin N-acetyltransferase SlSNAT2 Positively Regulates Tomato Resistance Against Ralstonia solanacearum. International Journal of Molecular Sciences. 2025; 26(13):6530. https://doi.org/10.3390/ijms26136530
Chicago/Turabian StyleWang, Yixi, Gengshou Xia, Xinyi Xie, Hao Wang, Lingyun Zheng, Zhijie He, Junxian Ye, Kangtong Xu, Qi Shi, Hui Yang, and et al. 2025. "Serotonin N-acetyltransferase SlSNAT2 Positively Regulates Tomato Resistance Against Ralstonia solanacearum" International Journal of Molecular Sciences 26, no. 13: 6530. https://doi.org/10.3390/ijms26136530
APA StyleWang, Y., Xia, G., Xie, X., Wang, H., Zheng, L., He, Z., Ye, J., Xu, K., Shi, Q., Yang, H., & Zhang, Y. (2025). Serotonin N-acetyltransferase SlSNAT2 Positively Regulates Tomato Resistance Against Ralstonia solanacearum. International Journal of Molecular Sciences, 26(13), 6530. https://doi.org/10.3390/ijms26136530