
Crystal Structure and Properties of Thallium(I) Salinomycinate

 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
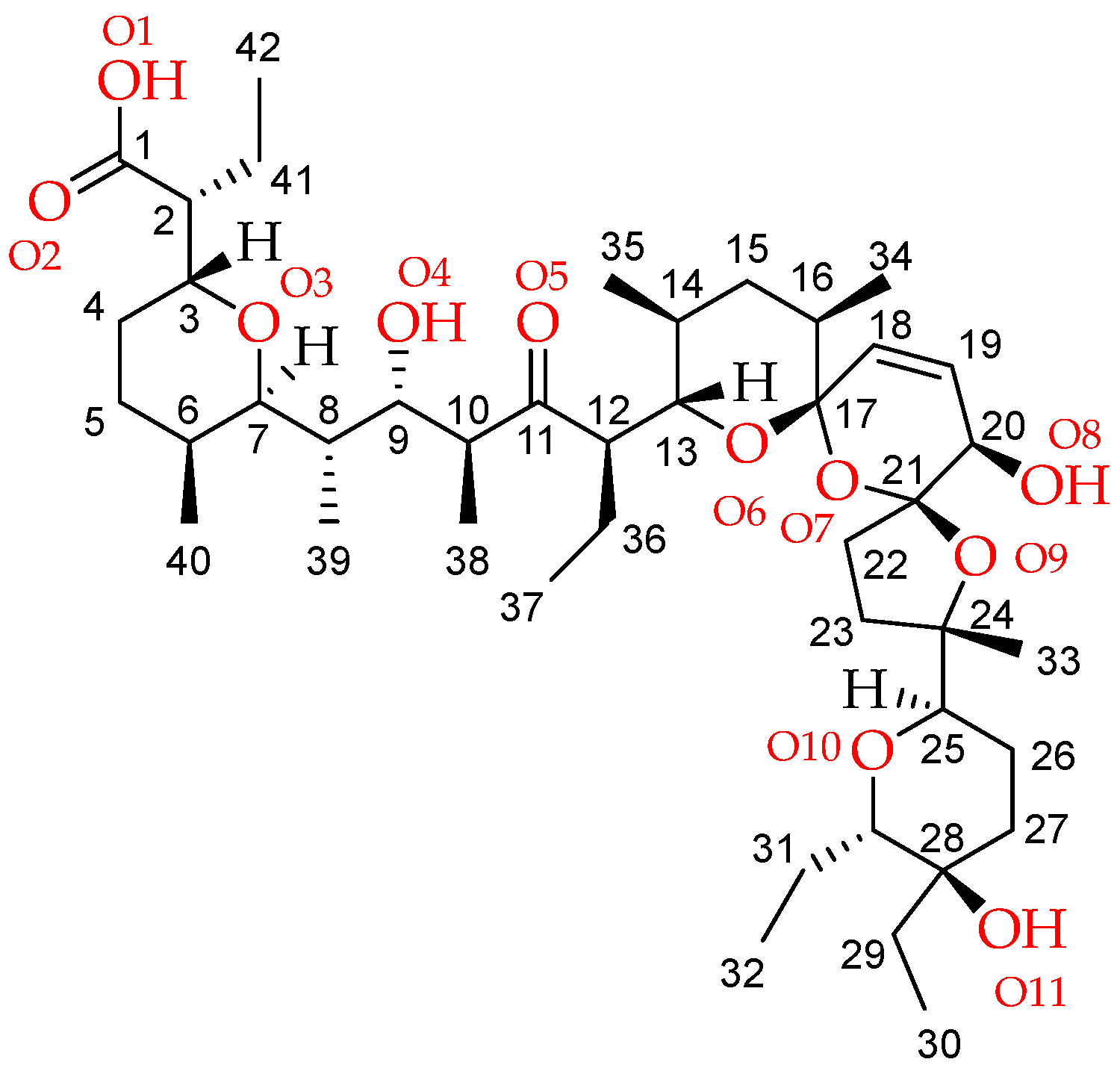
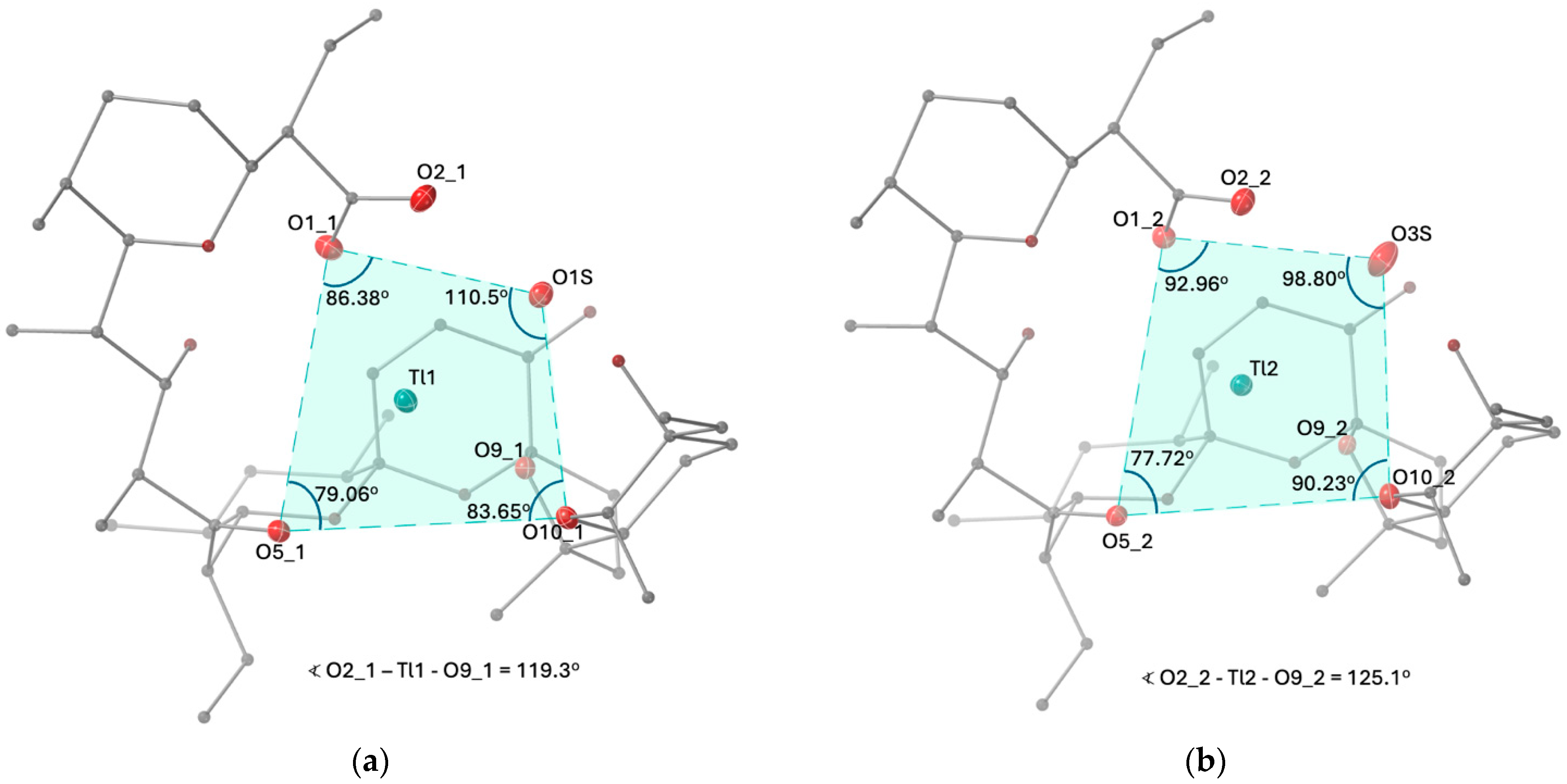
2.1. Description of the Crystal Structure of Thallium(I) Salinomycinate
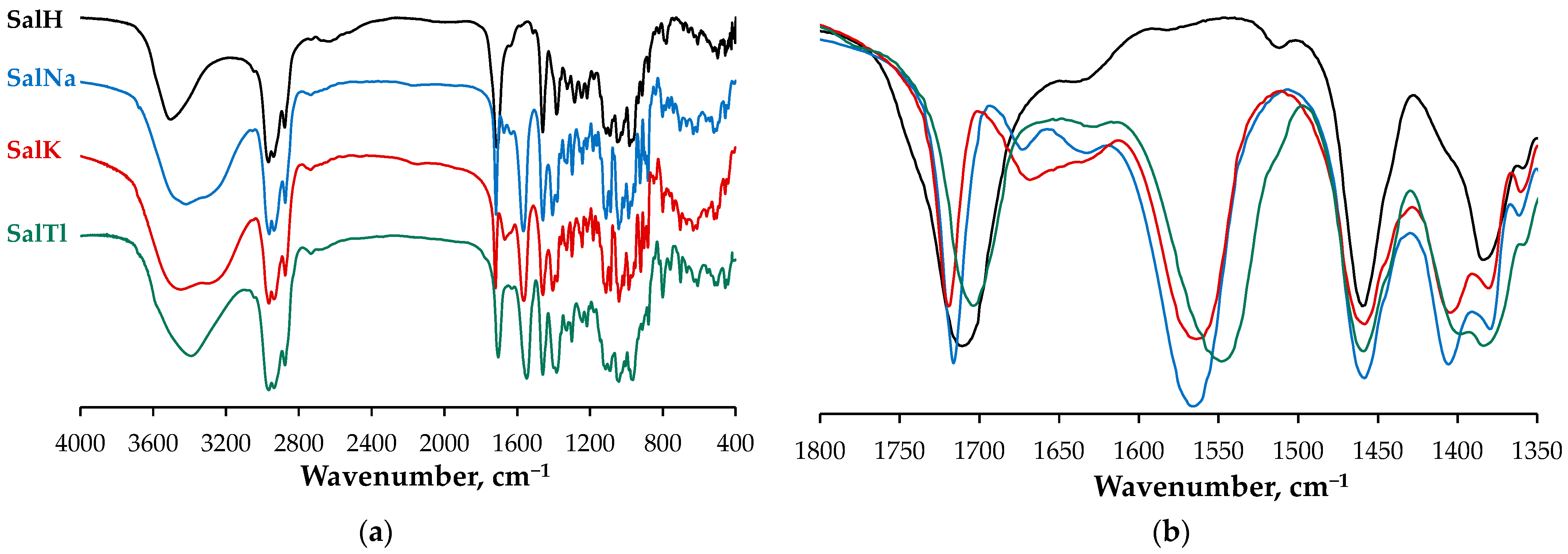
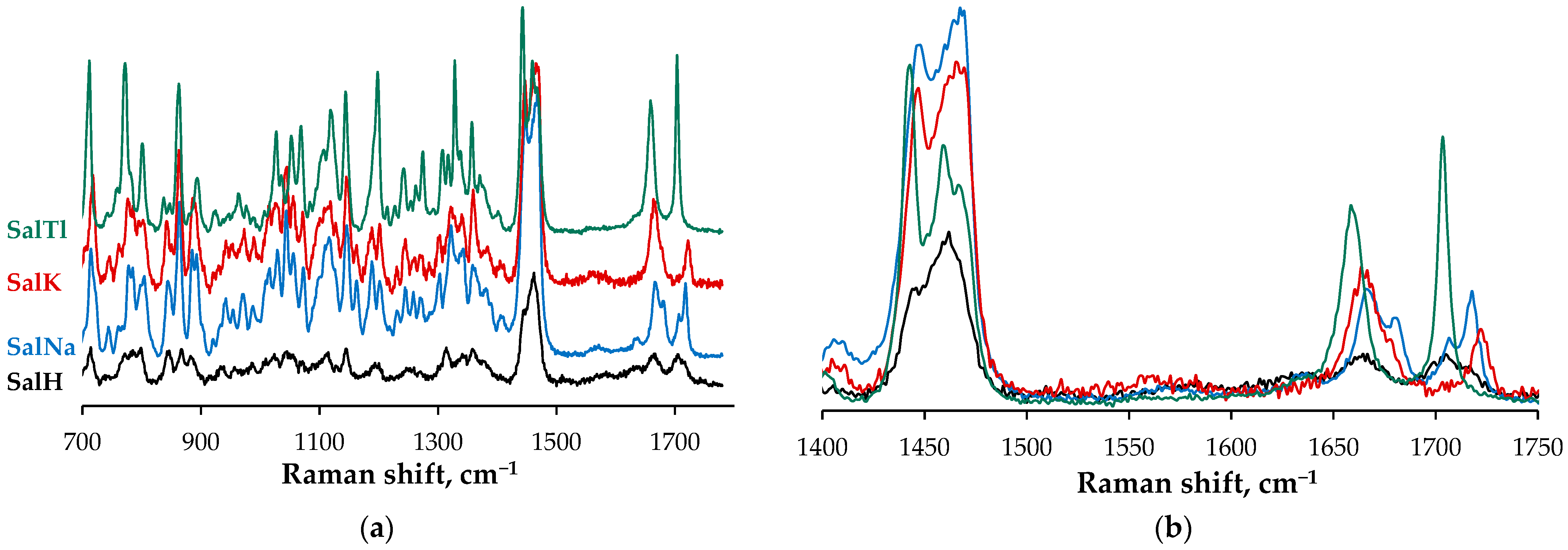
2.2. Spectral Characterization of SalTl
- -
- A symmetric band at 1703 cm−1, which is narrower (half-width of 28 cm−1) and less intense due to the carbonyl stretching of the isolated C=O group, engaged in Tl–O interaction according to the X-ray findings;
- -
- Two new bands at 1549 cm−1 and 1398 cm−1 (not observable in the spectrum of SalH), which are assigned to the asymmetric and symmetric stretches of the carboxylate function. These signals evidence the deprotonation of the carboxylic group in SalH under complexation with Tl+ cations. Furthermore, the magnitude of Δ (the difference between the two vibrations), calculated as 151 cm−1, is similar but still lower than that of the corresponding ionic values, and can be used as an indicator of an asymmetric bidentate coordination mode of the carboxylate function [37]. In contrast, this difference is 161 cm−1 for SalNa, where the carboxylate anion acts in a monodentate fashion [4], and is 159 cm−1 for SalK, supposed to be isostructural with SalNa;
- -
- An asymmetric broad band in the range of 3680–3120 cm−1 (centered at ca. 3390 cm−1, half-width of 312 cm−1) due to the OH-stretching vibrations of hydroxylic groups of salinomycinate ligand and water molecules in SalTl, engaged in the formation of Tl−O and H-bonds.
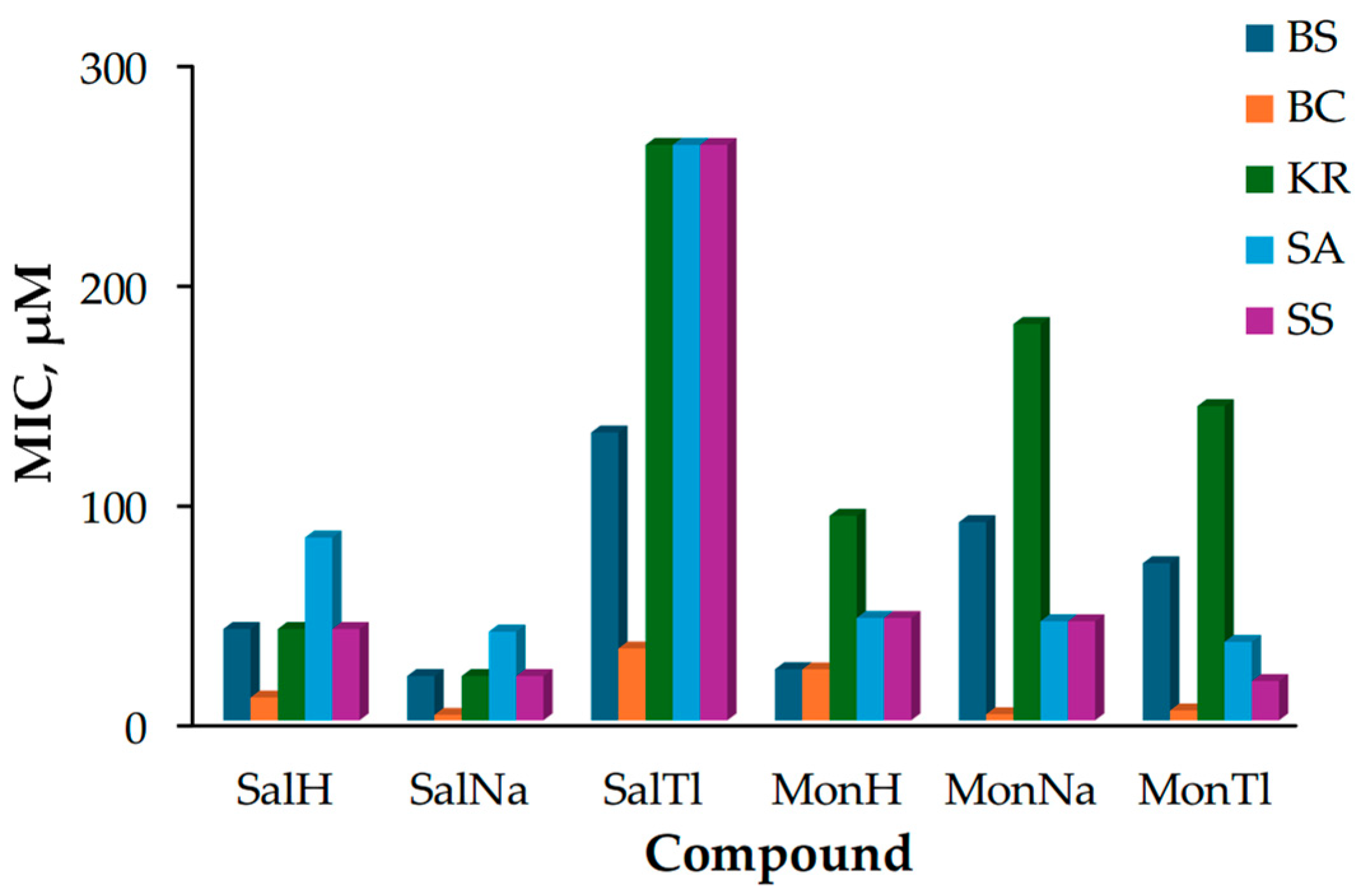
2.3. Biological Effect of SalTl and Related Compounds
- -
- -
- Bacillus cereus (BC) infections are associated with two types of food-borne gastroenteritis: emetic illness (due to the produced very stable toxin that is resistant to high temperatures and pH changes) and diarrhea (mediated by a less stable enterotoxin complex) [42];
- -
- -
- Staphylococcus aureus (SA) is an extraordinarily versatile pathogen responsible for staphylococcal food poisoning, hospital and community-acquired infections, as well as toxic shock syndrome [46];
- -
- Staphylococcus saprophyticus (SS) causes several rare infections, such as pyelonephritis, infective endocarditis, meningitis, as well as urinary tract infection, endophthalmitis, and uncomplicated cystitis [47].
- (1)
- The strain of B. cereus (orange boxes) is the most sensitive to the effects of the tested compounds, while K. rhizophila (green boxes) seems to be resistant except to SalNa and SalH;
- (2)
- Sodium salinomycinate (SalNa) appears to be the most toxic against the entire bacterial set, with its activity varying within a relatively narrow MIC range of 2.5–40.4 µM;
- (3)
- Thallium salinomycinate (SalTl) possesses the lowest toxicity and is less active than the corresponding monensinate counterpart (MonTl).
- BS: SalNa ≈ MonH > SalH > MonTl > MonNa > SalTl
- BC: SalNa ≈ MonNa > MonTl > SalH > MonH > SalTl
- KR: SalNa > SalH > MonH > MonTl > MonNa > SalTl
- SA: SalNa ≈ MonTl ≈ MonNa ≈ MonH > SalH > SalTl
- SS: SalNa ≈ MonTl > SalH ≈ MonNa ≈ MonH > SalTl.
3. Materials and Methods
3.1. Reagents and Materials
3.2. Synthesis of SalTl
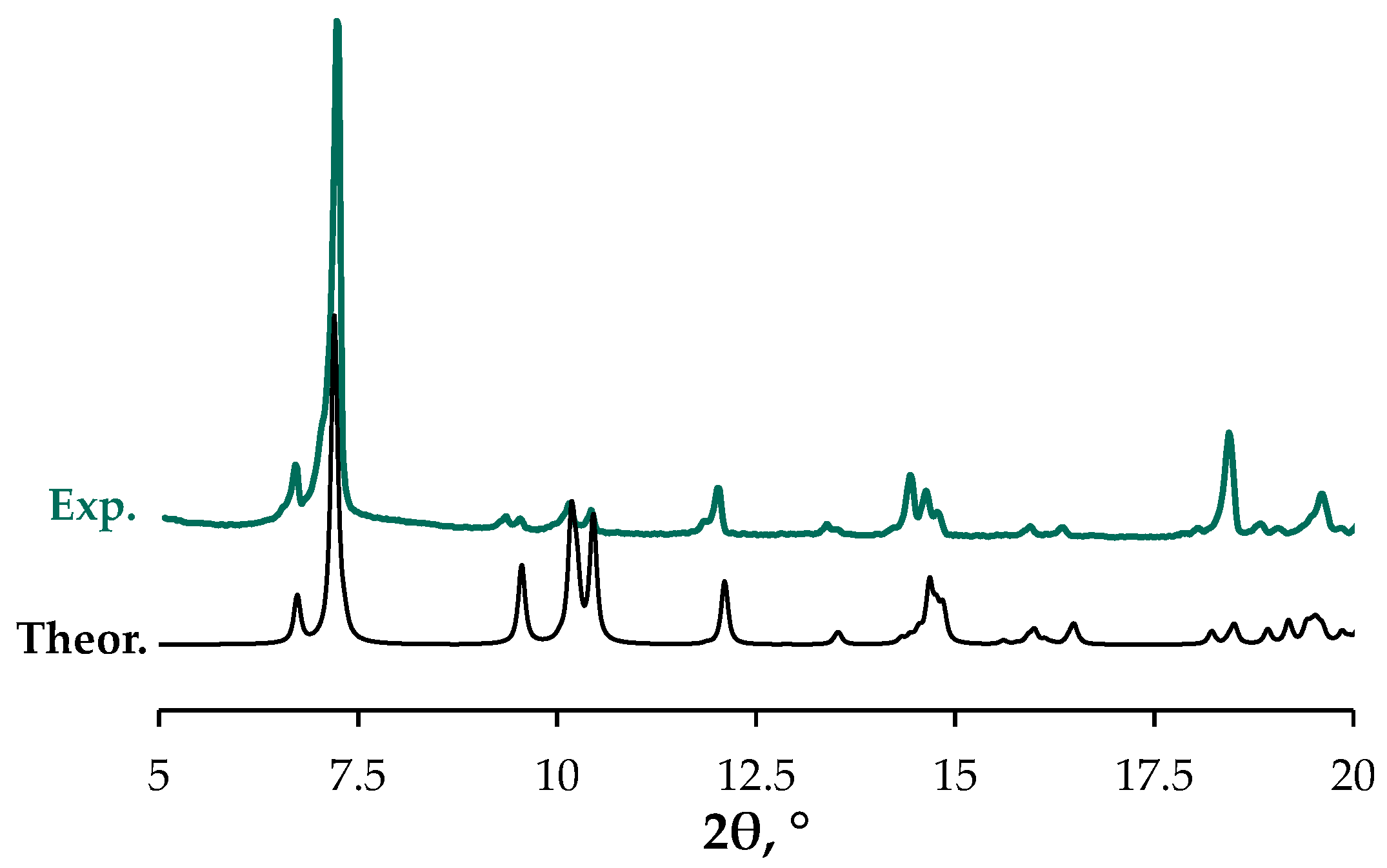
3.3. X-Ray Crystallography
3.4. Physical Measurements
3.5. Antibacterial Activity
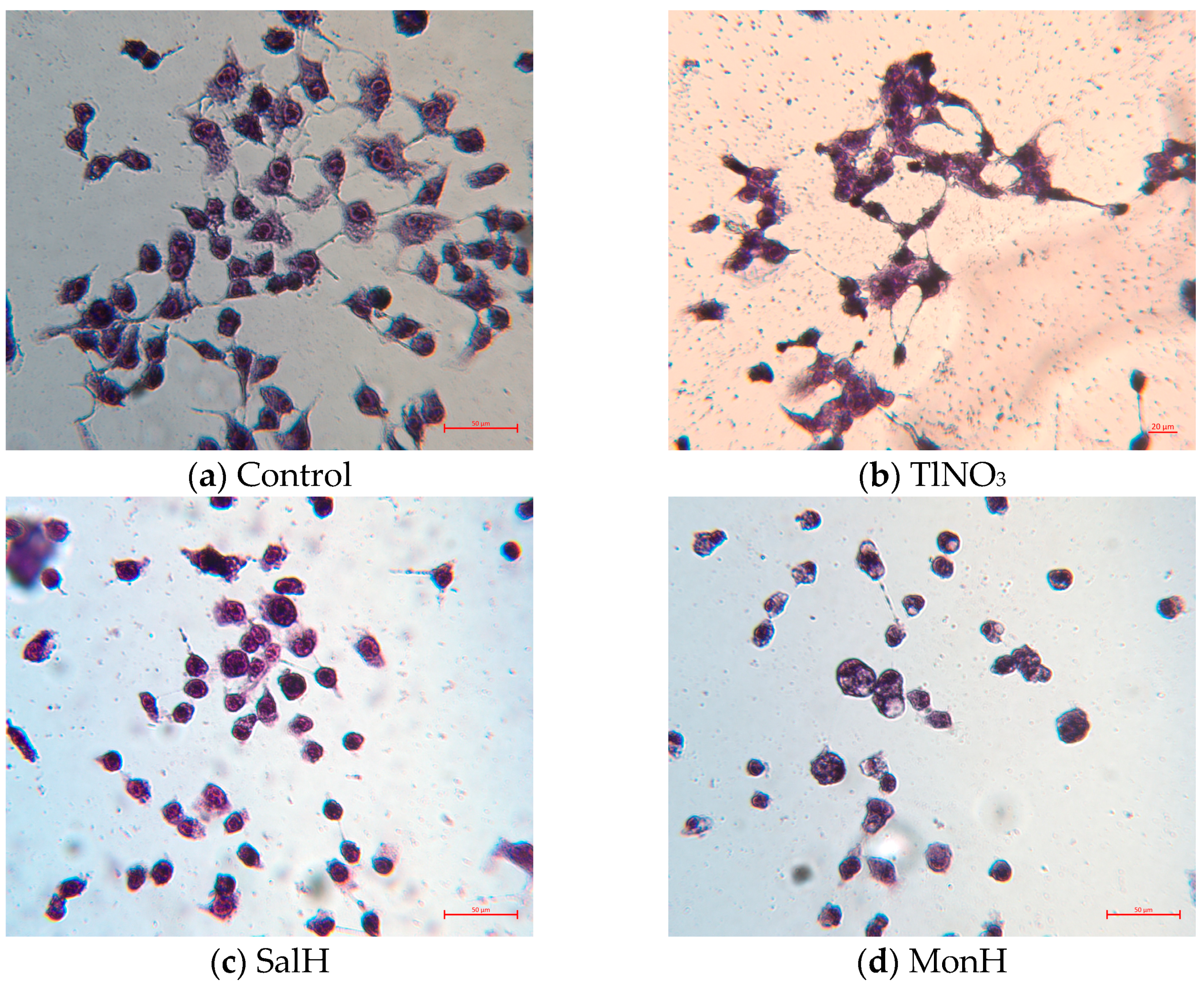
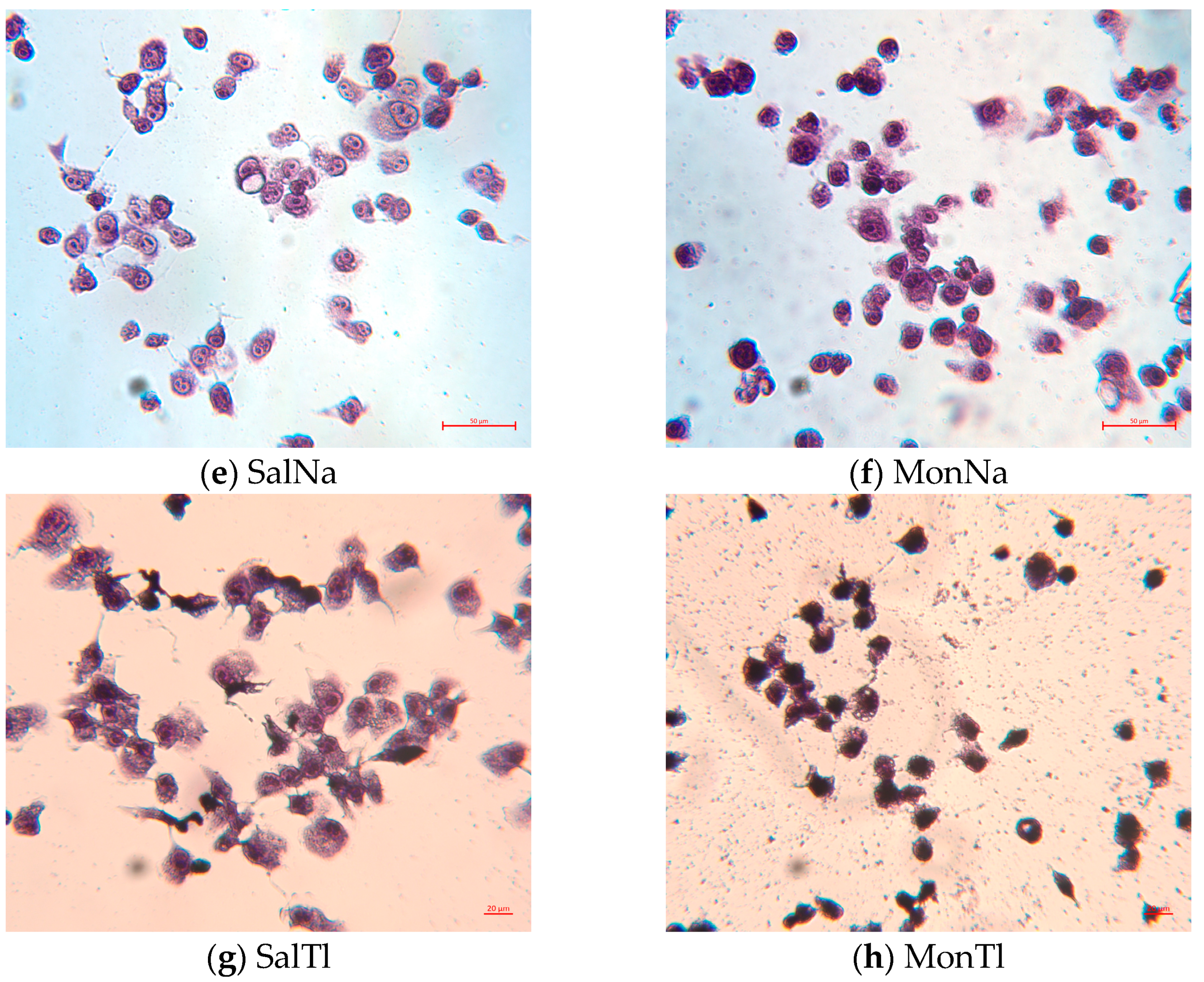
3.6. Cell Cultivation and Cytotoxicity Assay
3.7. May–Grünwald Giemsa (Pappenheim) Staining
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agtarap, A.; Chamberlin, J.W.; Pinkerton, M.; Steinrauf, L. The structure of monensic acid, a new biologically active compound. J. Am. Chem. Soc. 1967, 89, 5737–5739. [Google Scholar] [CrossRef] [PubMed]
- Agtarap, A.; Chamberlin, J.W. Monensin, a new biologically active compound. IV. Chemistry. Antimicrob. Agents Chemother. 1967, 7, 359–362. [Google Scholar] [PubMed]
- Miyazaki, Y.; Shibuya, M.; Sugawara, H.; Kawaguchi, O.; Hirsoe, C. Salinomycin, a new polyether antibiotic. J. Antibiotics 1974, 27, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Paulus, E.F.; Kurz, M.; Matter, H.; Vértesy, L. Solid-state and solution structure of the salinomycin−sodium complex: stabilization of different conformers for an ionophore in different environments. J. Am. Chem. Soc. 1998, 120, 8209–8221. [Google Scholar] [CrossRef]
- Patel, D.J.; Shen, C. Structural and kinetic studies of lasalocid A (X537A) and its silver, sodium, and barium salts in nonpolar solvents. Proc. Natl. Acad. Sci. USA 1976, 73, 1786–1790. [Google Scholar] [CrossRef]
- Erwin, G.S.; Heikkinen, J.; Pauliina Halima, P.; Haber, C.L. Streptomyces lasalocidi sp. nov. (formerly “Streptomyces lasaliensis”), an actinomycete isolated from soil which produces the polyether antibiotic lasalocid. Intern. J. System. Evolut. Microbiol. 2020, 70, 3076–3083. [Google Scholar] [CrossRef]
- Berg, D.H.; Hamil, R.L. The isolation and characterization of narasin, a new polyether antibiotic. J. Antibiot. 1978, 31, 1–6. [Google Scholar] [CrossRef]
- Jeffers, T.K.; Tonkinson, L.V.; Callender, M.E. Anticoccidial efficacy of narasin in battery cage trials. Poult. Sci. 1988, 67, 1043–1049. [Google Scholar] [CrossRef]
- Liu, C.M.; Hermann, T.E.; Downey, A.; Prosser, B.L.; Schildknecht, E.; Palleroni, N.J.; Westley, J.W.; Miller, P.A. Novel polyether antibiotics X-14868A, B, C, and D produced by a Nocardia. Discovery, fermentation, biological as well as ionophore properties and taxonomy of the producing culture. J. Antibiot. 1983, 36, 343–350. [Google Scholar] [CrossRef]
- McDougald, L.R.; Wang, G.T.; Kantor, S.; Schenkel, R.; Quarles, C. Efficacy of maduramicin against ionophore-tolerant field isolates of coccidia in broilers. Avian Dis. 1987, 31, 302–308. [Google Scholar] [CrossRef]
- Tynan, E.J., 3rd; Nelson, T.H.; Davies, R.A.; Wernau, W.C. The production of semduramicin by direct fermentation. J. Antibiot. 1992, 45, 813–815. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Rychlik, J.L.; Genovese, K.J.; Poole, T.L.; Jung, Y.S.; Bischoff, K.M.; Anderson, R.C.; Nisbet, D.J. Ionophores: Their use as ruminant growth promotants and impact on food safety. Curr. Issues Intest. Microbiol. 2003, 4, 43–51. [Google Scholar] [PubMed]
- Novilla, M.N. Ionophores. In Veterinary Toxicology; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2007; pp. 1021–1041. [Google Scholar] [CrossRef]
- Dutton, C.J.; Banks, B.J.; Cooper, C.B. Polyether ionophores. Nat. Prod. Rep. 1995, 12, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Riddell, F.G. Structure, conformation, and mechanism in the membrane transport of alkali metal ions by ionophoric antibiotics. Chirality 2002, 14, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Noack, S.; Chapman, H.D.; Selzer, P.M. Anticoccidial drugs of the livestock industry. Parasitol. Res. 2019, 118, 2009–2026. [Google Scholar] [CrossRef]
- Pinkerton, M.; Steinrauf, L.K. Molecular structure of monovalent metal cation complexes of monensin. J. Mol. Biol. 1970, 49, 533–546. [Google Scholar] [CrossRef]
- Duax, W.L.; Smith, G.D.; Strong, P.D. Complexation of metal ions by monensin. Crystal and molecular structure of hydrated and anhydrous crystal forms of sodium monensin. J. Am. Chem. Soc. 1980, 102, 6725–6729. [Google Scholar] [CrossRef]
- Pangborn, W.; Duax, W.L.; Langs, D. The hydrated potassium complex of the ionophore monensin A. J. Am. Chem. Soc. 1987, 109, 2163–2165. [Google Scholar] [CrossRef]
- Paz, F.A.A.; Gates, P.J.; Fowler, S.; Gallimore, A.; Harvey, B.; Lopes, N.P.; Stark, C.B.W.; Staunton, J.; Klinowski, J.; Spencer, J.B. Sodium monensin dihydrate. Acta Cryst. 2003, E59, m1050–m1052. [Google Scholar] [CrossRef]
- Paulus, E.F.; Vértesy, L. Crystal structure of the antibiotic SY-1 (20-deoxy-salinomycin): Sodium 2-(6-[2-(5-ethyl-5-hydroxy-6-methyl-tetrahydro-pyran-2-yl)-2,10,12-trimethyl-1,6,8-trioxa-dispiro [4.1.5.3]pentadec-13-en-9-yl]-2-hydroxy-1,3-dimethyl-4-oxo-heptyl-5-methyl-tetrahydro-pyran-2-yl)-butyrate—Methanolsolvate (1:0.69), C42H69NaO10 · 0.69CH3OH. Z. Kristallogr. NCS 2003, 218, 575–577. [Google Scholar] [CrossRef]
- Paulus, E.F.; Vértesy, L. Crystal structure of 2-(6-[2-(5-ethyl-5-hydroxy-6-methyl-tetrahydro-pyran-2-yl)-15-oxo-2,10,12-trimethyl-1,6,8-trioxa-dispiro[4.1.5.3]pentadec-13-en-9-yl]-2-hydroxy-1,3-dimethyl-4-oxo-heptyl-5-methyl-tetrahydro-pyran-2-yl)-butyrate sodium, Na(C42H67O11), SY-9—Antibiotic 20-oxo-salinomycin. Z. Kristallogr. NCS 2004, 219, 184–186. [Google Scholar] [CrossRef]
- Huczynski, A.; Ratajczak-Sitarz, M.; Katrusiak, A.; Brzezinski, B. Molecular structure of the 1:1 inclusion complex of monensin A sodium salt with acetonitrile. J. Mol. Struct. 2007, 832, 84–89. [Google Scholar] [CrossRef]
- Huczynski, A.; Ratajczak-Sitarz, M.; Katrusiak, A.; Brzezinski, B. Molecular structure of the 1:1 inclusion complex of monensin A lithium salt with acetonitrile. J. Mol. Struct. 2007, 871, 92–97. [Google Scholar] [CrossRef]
- Yildirim, S.O.; McKee, V.; Khardli, F.Z.; Mimouni, M.; Hadda, H.B. Rubidium(I) monensinate dihydrate. Acta Cryst. 2008, E64, m154–m155. [Google Scholar] [CrossRef]
- Huczynski, A.; Ratajczak-Sitarz, M.; Katrusiak, A.; Brzezinski, B. Molecular structure of rubidium six-coordinated dihydrate complex with monensin A. J. Mol. Struct. 2008, 888, 224–229. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Pooch, H.; Raether, W. The action of polyether ionophorous antibiotics (monensin, salinomycin, lasalocid) on developmental stages of Eimeria tenella (Coccidia, Sporozoa) in vivo and in vitro: Study by light and electron microscopy. Ztsch. Parasitenk. 1983, 69, 457–471. [Google Scholar] [CrossRef]
- Wang, Z.; Suo, X.; Xia, X.; Shen, J. Influence of monensin on cation influx and Na+-K+-ATPase activity of Eimeria tenella sporozoites in vitro. J. Parasitol. 2006, 92, 1092–1096. [Google Scholar] [CrossRef]
- Gough, L.P.; Shacklette, H.T.; Case, A.A. Element concentrations toxic to plants, animals, and man. Geol. Survey. Bull. 1979, 1466, 1–78. [Google Scholar] [CrossRef]
- Karbowska, B. Presence of thallium in the environment: Sources of contaminations, distribution and monitoring methods. Environ. Monit. Assess 2016, 188, art. 640. [Google Scholar] [CrossRef]
- Gagne, O.C.; Hawthorne, F.C. Bond-length distributions for ions bonded to oxygen: Metalloids and post-transition metals. Acta Cryst. 2018, B74, 63–78. [Google Scholar] [CrossRef]
- Aoki, K.; Suh, H.; Nagashima, H.; Uzawa, J.; Yamazaki, H. Crystal structures of two polymorphic thallium(I) salts of the antibiotic lasalocid A: A polymeric form involving metal-phenyl π-bonding and a monomeric form involving the “half-naked” metal ion. J. Am. Chem. Soc. 1992, 114, 5722–5729. [Google Scholar] [CrossRef]
- Akkurt, M.; Yıldırım, S.Ö.; Khardli, F.-Z.; Mimouni, M.; McKee, V.; Haddab, T.B. Crystal structure of a new polymeric thallium-lasalocid complex: Lasalocide anion-thallium(I) containing aryl-Tl interactions. Arkivoc 2008, xv, 121–132. [Google Scholar] [CrossRef]
- Alleaume, M.; Hickel, D. Crystal structure of the thallium salt of the antibiotic grisorixin. Chem. Comm. 1972, 175–176. [Google Scholar] [CrossRef]
- Sakurai, T.; Kobayashi, K.; Nakamura, G.; Isono, K. Structure of the thallium salt of cationomycin. Acta Cryst. 1982, B38, 2471–2473. [Google Scholar] [CrossRef]
- Riche, C.; Pascard-Billy, C. Emericid-thallium(I). In Proceedings of the 3rd European Crystallographic Meeting, Zurich, Switzerland, 6–10 September 1976. [Google Scholar]
- Deacon, G.B.; Phillips, R.J. Relationships between the carbon-oxygen stretching frequencies of carboxylato complexes and the type of carboxylate coordination. Coord. Chem. Rev. 1980, 33, 227–250. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Firsov, A.M.; Khailova, L.S.; Kotova, E.A.; Antonenko, Y.N. Selectivity of cation transport across lipid membranes by the antibiotic salinomycin. Biochim Biophys Acta Biomembr. 2023, 1865, 184182. [Google Scholar] [CrossRef]
- Gerasimova, T.P.; Katsyuba, S.A. Infrared and Raman bands of cyclopentadienyl ligands as indicators of electronic configuration of metal centers in metallocenes. J. Organom. Chem. 2015, 776, 30e34. [Google Scholar] [CrossRef]
- Saleh, F.; Kheirandish, F.; Azizi, H.; Azizi, M. Molecular diagnosis and characterization of Bacillus subtilis isolated from burn wound in Iran. Res. Mol. Med. 2014, 2, 40–44. [Google Scholar] [CrossRef]
- Loggenberg, S.R.; Twilley, D.; de Cahna, M.N.; Lall, N. Medicinal plants used in South Africa as antibacterial agents for wound healing. In Medicinal Plants as Anti-Infectives; Chassagne, F., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 139–182. [Google Scholar] [CrossRef]
- Anany, H.; Brovko, L.Y.; El Arabi, T.; Griffiths, M.W. Bacteriophages as antimicrobials in food products: Applications against particular pathogens. In Handbook of Natural Antimicrobials for Food Safety and Quality; Taylor, T.M., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 89–116. [Google Scholar] [CrossRef]
- Zaheen, Z.; War, A.F.; Ali, F.; Yatoo, A.M.; Ali, M.N.; Ahmad, S.B.; Rehman, M.U.; Paray, B.A. Common bacterial infections affecting freshwater fish fauna and impact of pollution and water quality characteristics on bacterial pathogenicity. Bact. Fish Dis. 2022, 133–154. [Google Scholar] [CrossRef]
- Hajam, Y.A.; Kumar, R.; Rani, R.; Sharma, P.; Diksha. Efficacy of different treatments available against bacterial pathogens in fish. Bact. Fish Dis. 2022, 379–398. [Google Scholar] [CrossRef]
- Becker, K.; Rutsch, F.; Uekötter, A.; Kipp, F.; König, J.; Marquardt, T.; Peters, G.; von Eiff, C. Kocuria rhizophila adds to the emerging spectrum of micrococcal species involved in human infections. J. Clin. Microbiol. 2008, 46, 3537–3539. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Silva, J.; Teixeira, P. Staphylococcus aureus, a food pathogen: Virulence factors and antibiotic resistance. In Handbook of Food Bioengineering, Foodborne Diseases; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 213–238. [Google Scholar] [CrossRef]
- Azimi, T.; Mirzadeh, M.; Sabour, A.; Nasser, A.; Fallah, F.; Pourmand, M.R. Coagulase-negative staphylococci (CoNS) meningitis: A narrative review of the literature from 2000 to 2020. New Microbes New Infect. 2020, 37, 100755. [Google Scholar] [CrossRef]
- Toxicological Review of Thallium and Compounds. U.S. Environmental Protection Agency, Washington, DC. EPA/635/R-08/001F. Available online: www.epa.gov/iris (accessed on 26 April 2025).
- Cervical cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cervical-cancer (accessed on 24 April 2025).
- Yahya, M.A.; Sharon, S.M.; Hantisteanu, S.; Hallak, M.; Bruchim, I. The role of the insulin-like growth factor 1 pathway in immune tumor microenvironment and its clinical ramifications in gynecologic malignancies. Front. Endocrinol. 2018, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal violet assay for determining viability of cultured cells. Cold Spring Harb Protoc. 2016, 2016, pdb.prot087379. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Zhu, Y.; Wu, Z.; Cui, C.; Cai, F. Anticancer mechanisms of salinomycin in breast cancer and its clinical applications. Front. Oncol. 2021, 11, 654428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Deng, Y.; Wang, J.; Yan, Z.; Wei, Q.; Ye, J.; Zhang, J.; He, T.-C.; Qiao, M. Effect of antibiotic monensin on cell proliferation and IGF1R signaling pathway in human colorectal cancer cells. Annals Med. 2023, 55, 954–964. [Google Scholar] [CrossRef]
- SADABS, v 2016/2; Bruker AXS Inc.: Madison, WI, USA, 2016.
- SAINTV8.38A, Bruker AXS Inc.: Madison, WI, USA, 2018.
- SHELXTL 2018, Bruker AXS Inc.: Madison, WI, USA, 2018.
- Dolomanov, O.V.; Bourhis, L.J.; Gildea, R.J.; Howard, J.A.; Puschmann, H.J. OLEX2: A complete structure solution, refinement and analysis program. J. Appl. Crystallogr. 2009, 42, 339–341. [Google Scholar] [CrossRef]
- Sheldrick, G.M. SHELXT—Integrated space-group and crystal-structure determination. Acta Cryst. A 2015, 71, 3–8. [Google Scholar] [CrossRef]
- Parsons, S.; Flack, H.D.; Wagner, T. Use of intensity quotients and differences in absolute structure refinement. Acta Cryst. B 2013, 69, 249–259. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bond Lenght | SalTl1 | SalTl2 |
|---|---|---|
| Tl—O1_n | 2.742(7) | 2.741(6) |
| Tl—O2_n | 3.330(6) | 3.138(6) |
| Tl—O5_n | 2.866(5) | 2.897(5) |
| Tl—O9_n | 2.788(5) | 2.724(5) |
| Tl—O10_n | 3.266(5) | 3.041(6) |
| Tl—O1S | 2.699(6) | - |
| Tl—O3S | - | 3.029(9) |
| Bond angle | SalTl1 | SalTl2 |
| O1_n—Tl—O5_n | 109.5(2) | 107.3(2) |
| O1_n—Tl—O10_n | 152.6(2) | 155.3(2) |
| O1_n—Tl—O1S | 78.5(2) | - |
| O1_n—Tl—O3S | - | 76.1(2) |
| O1_n—Tl—O2_n | 41.9(2) | 43.6(2) |
| O2_n—Tl—O1S | 59.8(2) | - |
| O2_n—Tl—O3S | - | 51.4(2) |
| O9_n—Tl—O5_n | 103.6(1) | 96.9(1) |
| O9_n—Tl—O10_n | 56.4(1) | 58.3(2) |
| Bond | SalTl1 | SalTl2 |
|---|---|---|
| O1_n—O4_n | 2.680(9) | 2.720(8) |
| O2_n—O1S | 2.602(8) | - |
| O2_n—O3S | - | 2.675(9) |
| O8_n—O1S | 2.718(8) | - |
| O8_n—O3S | - | 2.684(9) |
| O11_n—O1S | 2.672(8) | - |
| O11_n—O3S | - | 2.651(10) |
| O8_n—O2S | 2.714(9) | - |
| O2_n—O2S | - | 2.848(9) |
| O11_n—O2S | - | 2.738(9) |
| Compound | Vibration | IR, cm−1 | Raman, cm−1 |
|---|---|---|---|
| SalH | νC=O (isolated) | 1704 | 1705 |
| νC=O (COOH) | 1716 | - | |
| νC=C | “hidden” | 1664 | |
| SalNa | νC=O (isolated) | 1716 | 1718, 1706 |
| νC=O (COO−, asym) | 1567 | 1570 | |
| νC=O (COO−, sym) | 1406 | - | |
| νC=C | “hidden” | 1681/1667 | |
| SalK | νC=O (isolated) | 1719 | 1722 |
| νC=O (COO−, asym) | 1564 | 1560 | |
| νC=O (COO−, sym) | 1405 | - | |
| νC=C | “hidden” | 1665 | |
| SalTl | νC=O (isolated) | 1703 | 1703 |
| νC=O (COO−, asym) | 1549 | 1555 | |
| νC=O (COO−, sym) | 1398 | - | |
| νC=C | “hidden” | 1659 |
| Band | C17−C18 | C18=C19 | C19−C20 | C20−C21 | C21−O7 | O7−C17 |
|---|---|---|---|---|---|---|
| SalNa1 | 1.485 | 1.344 | 1.492 | 1.556 | 1.429 | 1.433 |
| SalNa2 | 1.472 | 1.311 | 1.420 | 1.566 | 1.435 | 1.429 |
| average | 1.479 | 1.328 | 1.456 | 1.561 | 1.432 | 1.431 |
| SalTl1 | 1.507 | 1.318 | 1.497 | 1.522 | 1.419 | 1.431 |
| SalTl2 | 1.484 | 1.328 | 1.494 | 1.527 | 1.422 | 1.425 |
| average | 1.500 | 1.323 | 1.500 | 1.525 | 1.421 | 1.428 |
| Angle | C17−C18−C19 | C18=C19−C20 | C19−C20−C21 | C20−C21−O7 | C21−O7−C17 | O7−C17−C18 |
| SalNa1 | 122.1 | 121.0 | 112.4 | 105.7 | 122.4 | 113.0 |
| SalNa2 | 123. 0 | 124.7 | 111.1 | 109.1 | 120.7 | 112.2 |
| average | 122.6 | 122.9 | 111.8 | 107.4 | 121.6 | 112.6 |
| SalTl1 | 122.7 | 121.2 | 108.9 | 110.5 | 121.4 | 112.7 |
| SalTl2 | 122.7 | 120.2 | 109.0 | 109.0 | 120.3 | 113.6 |
| average | 122.7 | 120.7 | 109.0 | 109.8 | 120.9 | 113.2 |
| Crystal Data | SalTl |
|---|---|
| Chemical formula | C42H69O11Tl × 1.5H2O |
| Mr | 981.36 |
| Crystal system, space group | Monoclinic, P1 |
| Temperature (K) | 107 |
| a (Å) | 12.2005(7) |
| b (Å) | 13.7304(8) |
| c (Å) | 14.7041(8) |
| α (°) | 116.623(2) |
| β (°) | 93.382(2) |
| γ (°) | 90.454(2) |
| V (Å3) | 2196.6(2) |
| Z | 2 |
| Radiation type, λ [Å] | Cu Kα, 1.54178 |
| µ (mm−1) | 7.559 |
| Crystal size (mm3) | 0.09 × 0.15 × 0.19 |
| Data collection | |
| Diffractometer | Bruker APEX-II CCD |
| Absorption correction | Multi-scan |
| Tmin, Tmax | 0.482, 0.753 |
| No. of measured, independent, and observed [I > 2σ(I)] reflections | 40202, 14134, 14042 |
| Rint | 0.0527 |
| Refinement | |
| R[F2 > 2σ(F2)], wR(F2), S | 0.0360, 0.0918, 1.061 |
| No. of parameters | 1033 |
| No. of restraints | 3 |
| Δρmax, Δρmin (e Å−3) | 2.8 (around the very heavy Tl atom)/−0.8 |
| Absolute structure | Flack x determined using quotients [(I+)-(I–)]/[(I+)+(I–)] [59] |
| 0.086(7) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petkov, N.; Dorkov, P.; Ugrinov, A.; Encheva, E.; Abrashev, M.; Zasheva, D.; Daneva, T.; Pantcheva, I.N. Crystal Structure and Properties of Thallium(I) Salinomycinate. Int. J. Mol. Sci. 2025, 26, 6504. https://doi.org/10.3390/ijms26136504
Petkov N, Dorkov P, Ugrinov A, Encheva E, Abrashev M, Zasheva D, Daneva T, Pantcheva IN. Crystal Structure and Properties of Thallium(I) Salinomycinate. International Journal of Molecular Sciences. 2025; 26(13):6504. https://doi.org/10.3390/ijms26136504
Chicago/Turabian StylePetkov, Nikolay, Petar Dorkov, Angel Ugrinov, Elzhana Encheva, Miroslav Abrashev, Diana Zasheva, Teodora Daneva, and Ivayla N. Pantcheva. 2025. "Crystal Structure and Properties of Thallium(I) Salinomycinate" International Journal of Molecular Sciences 26, no. 13: 6504. https://doi.org/10.3390/ijms26136504
APA StylePetkov, N., Dorkov, P., Ugrinov, A., Encheva, E., Abrashev, M., Zasheva, D., Daneva, T., & Pantcheva, I. N. (2025). Crystal Structure and Properties of Thallium(I) Salinomycinate. International Journal of Molecular Sciences, 26(13), 6504. https://doi.org/10.3390/ijms26136504