
Prediabetes May Alter HPA Axis Activity and Regulation: A Study on Patients with Prediabetes
 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Population Demographics
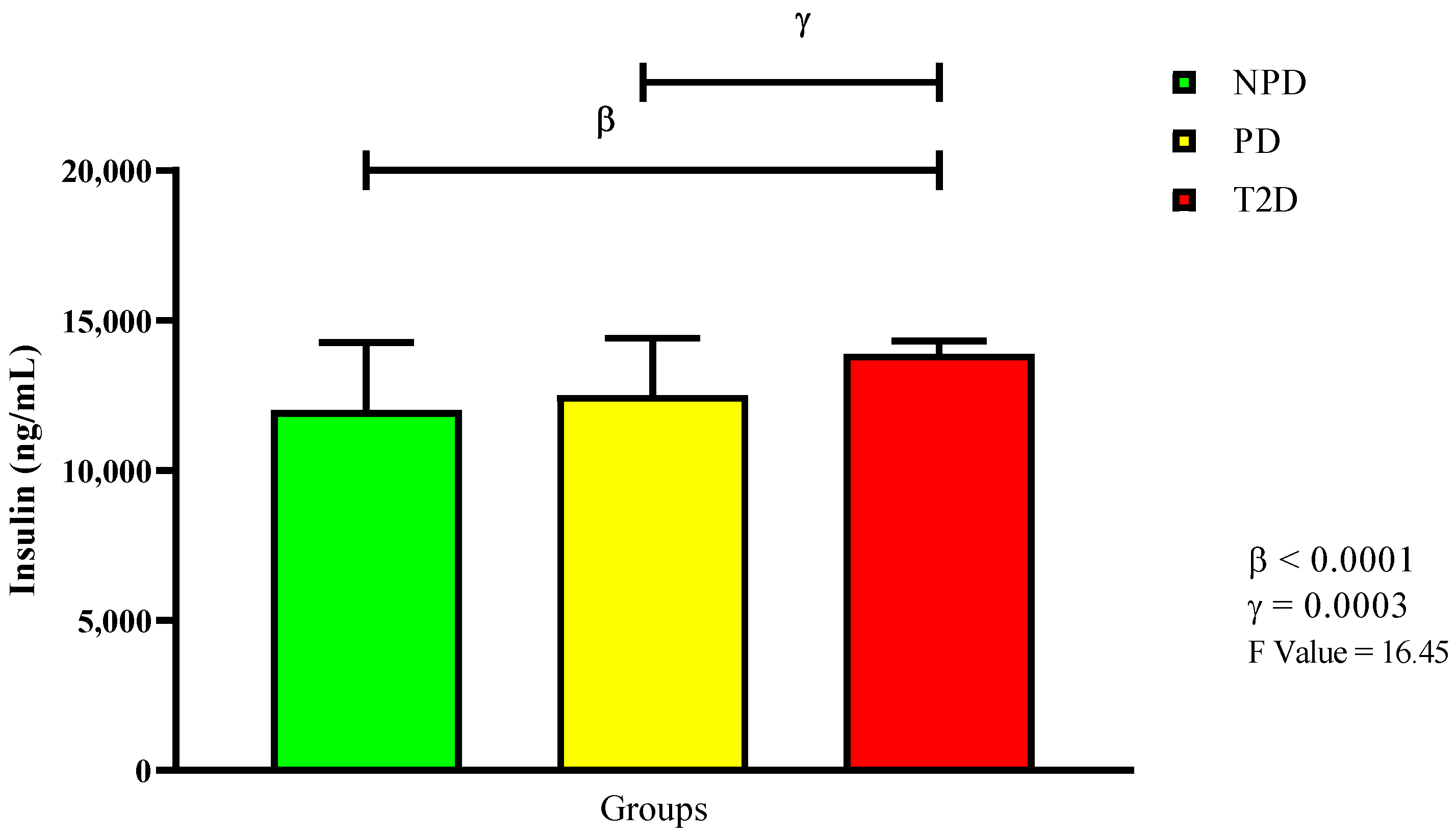
2.2. Glycated Hemoglobin, Plasma Glucose, and Plasma Insulin
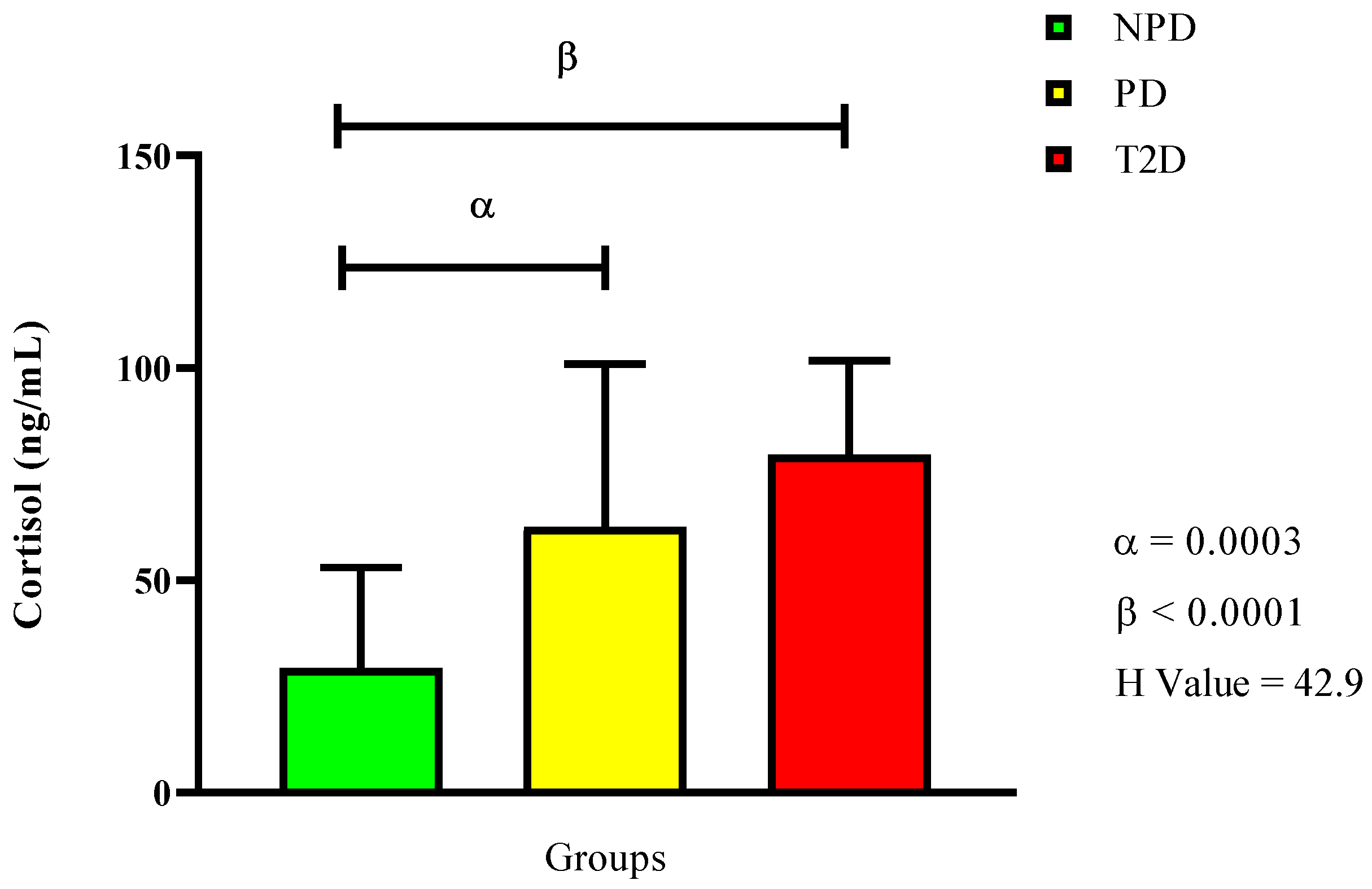
2.3. Plasma Cortisol
2.4. Plasma ACTH
2.5. Plasma Epinephrine and Norepinephrine
3. Discussion
4. Materials and Methods
4.1. Study Design and Setting
4.2. Sample Size Determination
4.3. Sampling
4.4. Biochemical Analysis
4.5. Statistical Analysis
5. Conclusions
5.1. Study Limitations
5.2. Future Studies Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, H.-G.; Cheon, E.-J.; Bai, D.-S.; Lee, Y.H.; Koo, B.-H. Stress and heart rate variability: A meta-analysis and review of the literature. Psychiatry Investig. 2018, 15, 235. [Google Scholar] [CrossRef] [PubMed]
- Janssen, J.A. New Insights into the Role of Insulin and Hypothalamic-Pituitary-Adrenal (HPA) Axis in the Metabolic Syndrome. Int. J. Mol. Sci. 2022, 23, 8178. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.L.; Deak, T. A users guide to HPA axis research. Physiol. Behav. 2017, 178, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603. [Google Scholar] [CrossRef]
- Rosmond, R. Stress induced disturbances of the HPA axis: A pathway to type 2 diabetes? Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2003, 9, RA35–RA39. [Google Scholar]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A comprehensive overview on stress neurobiology: Basic concepts and clinical implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef]
- Keller-Wood, M. Hypothalamic-pituitary-adrenal Axis—Feedback control. Compr. Physiol. 2011, 5, 1161–1182. [Google Scholar] [CrossRef]
- Sharma, V.K.; Singh, T.G. Chronic stress and diabetes mellitus: Interwoven pathologies. Curr. Diabetes Rev. 2020, 16, 546–556. [Google Scholar]
- Kim, S.; Park, E.S.; Chen, P.R.; Kim, E. Dysregulated hypothalamic–pituitary–adrenal axis is associated with increased inflammation and worse outcomes after ischemic stroke in diabetic mice. Front. Immunol. 2022, 13, 864858. [Google Scholar] [CrossRef]
- Prpić-Križevac, I.; Canecki-Varžić, S.; Bilić-Ćurčić, I. Hyperactivity of the hypothalamic-pituitary-adrenal axis in patients with type 2 diabetes and relations with insulin resistance and chronic complications. Wien. Klin. Wochenschr. 2012, 124, 403–411. [Google Scholar] [CrossRef]
- Nolan, C.J.; Damm, P.; Prentki, M. Type 2 diabetes across generations: From pathophysiology to prevention and management. Lancet 2011, 378, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Ozougwu, J.; Obimba, K.; Belonwu, C.; Unakalamba, C. The pathogenesis and pathophysiology of type 1 and type 2 diabetes mellitus. J. Physiol. Pathophysiol. 2013, 4, 46–57. [Google Scholar] [CrossRef]
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2010, 33 (Suppl. S1), S62–S69, Erratum in Diabetes Care 2010, 33, e57. [Google Scholar]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Mkhize, B.C.; Mosili, P.; Ngubane, P.S.; Sibiya, N.H.; Khathi, A. The Relationship between Renin–Angiotensin–Aldosterone System (RAAS) Activity, Osteoporosis and Estrogen Deficiency in Type 2 Diabetes. Int. J. Mol. Sci. 2023, 24, 11963. [Google Scholar] [CrossRef]
- Mezuk, B.; Eaton, W.W.; Albrecht, S.; Golden, S.H. Depression and type 2 diabetes over the lifespan: A meta-analysis. Diabetes Care 2008, 31, 2383–2390. [Google Scholar] [CrossRef]
- Gemeay, E.M.; Moawed, S.A.; Mansour, E.A.; Ebrahiem, N.E.; Moussa, I.M.; Nadrah, W.O. The association between diabetes and depression. Saudi Med. J. 2015, 36, 1210. [Google Scholar] [CrossRef]
- Golden, S.H.; Lazo, M.; Carnethon, M.; Bertoni, A.G.; Schreiner, P.J.; Roux, A.V.D.; Lee, H.B.; Lyketsos, C. Examining a bidirectional association between depressive symptoms and diabetes. JAMA 2008, 299, 2751–2759. [Google Scholar] [CrossRef]
- Chiodini, I.; Adda, G.; Scillitani, A.; Coletti, F.; Morelli, V.; Di Lembo, S.; Epaminonda, P.; Masserini, B.; Beck-Peccoz, P.; Orsi, E. Cortisol secretion in patients with type 2 diabetes: Relationship with chronic complications. Diabetes Care 2007, 30, 83–88. [Google Scholar] [CrossRef]
- Lundqvist, M.H.; Pereira, M.J.; Wiklund, U.; Hetty, S.; Eriksson, J.W. Autonomic nervous system responses to hypo-and hyperglycemia in type 2 diabetes and prediabetes. Eur. J. Endocrinol. 2024, 191, 499–508. [Google Scholar] [CrossRef]
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimäki, M. Prediabetes: A high-risk state for developing diabetes. Lancet 2012, 379, 2279. [Google Scholar] [CrossRef] [PubMed]
- Sosibo, A.M.; Mzimela, N.C.; Ngubane, P.S.; Khathi, A. Prevalence and correlates of pre-diabetes in adults of mixed ethnicities in the South African population: A systematic review and meta-analysis. PLoS ONE 2022, 17, e0278347. [Google Scholar] [CrossRef] [PubMed]
- Ferrannini, E.; Gastaldelli, A.; Iozzo, P. Pathophysiology of prediabetes. Med. Clin. 2011, 95, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M. Pathophysiology of prediabetes and treatment implications for the prevention of type 2 diabetes mellitus. Endocrine 2013, 43, 504–513. [Google Scholar] [CrossRef]
- Mosili, P.; Mkhize, B.C.; Ngubane, P.; Sibiya, N.; Khathi, A. The dysregulation of the hypothalamic–pituitary–adrenal axis in diet-induced prediabetic male Sprague Dawley rats. Nutr. Metab. 2020, 17, 104. [Google Scholar] [CrossRef]
- Sosibo, A.M.; Mzimela, N.C.; Ngubane, P.S.; Khathi, A. Prevalence of pre-diabetes in adults aged 25–45 years in a Durban-based clinical setting, South Africa: A retrospective study. Prim. Care Diabetes 2023, 17, 650–654. [Google Scholar] [CrossRef]
- Sosibo, A.M.; Mzimela, N.C.; Ngubane, P.S.; Khathi, A. Hormone imbalances detected in study participants with pre-diabetes in a Durban-based clinical setting, South Africa. Int. J. Diabetes Dev. Ctries. 2025, 45, 416–423. [Google Scholar] [CrossRef]
- Liu, H.; Bravata, D.M.; Cabaccan, J.; Raff, H.; Ryzen, E. Elevated late-night salivary cortisol levels in elderly male type 2 diabetic veterans. Clin. Endocrinol. 2005, 63, 642–649. [Google Scholar] [CrossRef]
- Malek, H.; Ebadzadeh, M.M.; Safabakhsh, R.; Razavi, A.; Zaringhalam, J. Dynamics of the HPA axis and inflammatory cytokines: Insights from mathematical modeling. Comput. Biol. Med. 2015, 67, 1–12. [Google Scholar] [CrossRef]
- Christiansen, J.J.; Djurhuus, C.B.; Gravholt, C.H.; Iversen, P.; Christiansen, J.S.; Schmitz, O.; Weeke, J.; Jørgensen, J.O.L.; Møller, N. Effects of cortisol on carbohydrate, lipid, and protein metabolism: Studies of acute cortisol withdrawal in adrenocortical failure. J. Clin. Endocrinol. Metab. 2007, 92, 3553–3559. [Google Scholar] [CrossRef]
- Hao, Y.; Tong, Y.; Guo, Y.; Lang, X.; Huang, X.; Xie, X.; Guan, Y.; Li, Z. Metformin attenuates the metabolic disturbance and depression-like behaviors induced by corticosterone and mediates the glucose metabolism pathway. Pharmacopsychiatry 2021, 54, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Caratti, G.; Matthews, L.; Poolman, T.; Kershaw, S.; Baxter, M.; Ray, D. Glucocorticoid receptor function in health and disease. Clin. Endocrinol. 2015, 83, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.; McKlveen, J.; Solomon, M.; Carvalho-Netto, E.; Myers, B. Neural regulation of the stress response: Glucocorticoid feedback mechanisms. Braz. J. Med. Biol. Res. 2012, 45, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Beaupere, C.; Liboz, A.; Fève, B.; Blondeau, B.; Guillemain, G. Molecular mechanisms of glucocorticoid-induced insulin resistance. Int. J. Mol. Sci. 2021, 22, 623. [Google Scholar] [CrossRef]
- Gathercole, L.L.; Lavery, G.G.; Morgan, S.A.; Cooper, M.S.; Sinclair, A.J.; Tomlinson, J.W.; Stewart, P.M. 11β-Hydroxysteroid dehydrogenase 1: Translational and therapeutic aspects. Endocr. Rev. 2013, 34, 525–555. [Google Scholar] [CrossRef]
- Stewart, P.M.; Krozowski, Z.S. 11β-Hydroxysteroid dehydrogenase. Vitam. Horm. 1997, 57, 249–324. [Google Scholar]
- Lavery, G.G.; Zielinska, A.E.; Gathercole, L.L.; Hughes, B.; Semjonous, N.; Guest, P.; Saqib, K.; Sherlock, M.; Reynolds, G.; Morgan, S.A. Lack of significant metabolic abnormalities in mice with liver-specific disruption of 11β-hydroxysteroid dehydrogenase type 1. Endocrinology 2012, 153, 3236–3248. [Google Scholar] [CrossRef]
- Lee, Y.B.; Hwang, H.J.; Kim, E.; Lim, S.H.; Chung, C.H.; Choi, E.H. Hyperglycemia-activated 11β-hydroxysteroid dehydrogenase type 1 increases endoplasmic reticulum stress and skin barrier dysfunction. Sci. Rep. 2023, 13, 9206. [Google Scholar] [CrossRef]
- Stulnig, T.; Waldhäusl, W. 11β-hydroxysteroid dehydrogenase type 1 in obesity and type 2 diabetes. Diabetologia 2004, 47, 1–11. [Google Scholar] [CrossRef]
- Spiga, F.; Lightman, S.L. Dynamics of adrenal glucocorticoid steroidogenesis in health and disease. Mol. Cell. Endocrinol. 2015, 408, 227–234. [Google Scholar] [CrossRef]
- Nezi, M.; Mastorakos, G.; Mouslech, Z. Corticotropin Releasing Hormone and the Immune/Inflammatory Response; MDText.com, Inc.: Dartmouth, MA, USA, 2015. [Google Scholar]
- Arnetz, L.; Ekberg, N.R.; Brismar, K.; Alvarsson, M. Gender difference in adrenal sensitivity to ACTH is abolished in type 2 diabetes. Endocr. Connect. 2015, 4, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Oliván, B.; Laferrère, B. Stress and obesity: The role of the hypothalamic–pituitary–adrenal axis in metabolic disease. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 340. [Google Scholar] [CrossRef] [PubMed]
- Bruehl, H.; Rueger, M.; Dziobek, I.; Sweat, V.; Tirsi, A.; Javier, E.; Arentoft, A.; Wolf, O.T.; Convit, A. Hypothalamic-pituitary-adrenal axis dysregulation and memory impairments in type 2 diabetes. J. Clin. Endocrinol. Metab. 2007, 92, 2439–2445. [Google Scholar] [CrossRef] [PubMed]
- Nouwen, A.; Winkley, K.; Twisk, J.; Lloyd, C.E.; Peyrot, M.; Ismail, K.; Pouwer, F.; Consortium, E.D.i.D.R. Type 2 diabetes mellitus as a risk factor for the onset of depression: A systematic review and meta-analysis. Diabetologia 2010, 53, 2480–2486. [Google Scholar] [CrossRef]
- Li, M.; Zhang, J.; Yang, G.; Zhang, J.; Han, M.; Zhang, Y.; Liu, Y. Effects of Anterior Pituitary Adenomas’ Hormones on Glucose Metabolism and Its Clinical Implications. Diabetes Metab. Syndr. Obes. 2023, 16, 409–424. [Google Scholar] [CrossRef]
- Monzer, N.; Hartmann, M.; Buckert, M.; Wolff, K.; Nawroth, P.; Kopf, S.; Kender, Z.; Friederich, H.-C.; Wild, B. Associations of childhood neglect with the ACTH and plasma cortisol stress response in patients with Type 2 diabetes. Front. Psychiatry 2021, 12, 679693. [Google Scholar] [CrossRef]
- Adam, T.C.; Hasson, R.E.; Ventura, E.E.; Toledo-Corral, C.; Le, K.-A.; Mahurkar, S.; Lane, C.J.; Weigensberg, M.J.; Goran, M.I. Cortisol is negatively associated with insulin sensitivity in overweight Latino youth. J. Clin. Endocrinol. Metab. 2010, 95, 4729–4735. [Google Scholar] [CrossRef]
- Blondeau, B.; Sahly, I.; Massourides, E.; Singh-Estivalet, A.; Valtat, B.; Dorchene, D.; Jaisser, F.; Breant, B.; Tronche, F. Novel transgenic mice for inducible gene overexpression in pancreatic cells define glucocorticoid receptor-mediated regulations of beta cells. PLoS ONE 2012, 7, e30210. [Google Scholar] [CrossRef]
- Lee, Z.; Chan, J.; Yeung, V.; Chow, C.-C.; Lau, M.; Ko, G.; Li, J.; Cockram, C.S.; Critchley, J. Plasma insulin, growth hormone, cortisol, and central obesity among young Chinese type 2 diabetic patients. Diabetes Care 1999, 22, 1450–1457. [Google Scholar] [CrossRef]
- Piroli, G.G.; Grillo, C.A.; Reznikov, L.R.; Adams, S.; McEwen, B.S.; Charron, M.J.; Reagan, L.P. Corticosterone impairs insulin-stimulated translocation of GLUT4 in the rat hippocampus. Neuroendocrinology 2007, 85, 71–80. [Google Scholar] [CrossRef]
- Lambillotte, C.; Gilon, P.; Henquin, J.-C. Direct glucocorticoid inhibition of insulin secretion. An in vitro study of dexamethasone effects in mouse islets. J. Clin. Investig. 1997, 99, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Hermans, E.J.; Henckens, M.J.; Joëls, M.; Fernández, G. Dynamic adaptation of large-scale brain networks in response to acute stressors. Trends Neurosci. 2014, 37, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Wadsworth, M.E.; Broderick, A.V.; Loughlin-Presnal, J.E.; Bendezu, J.J.; Joos, C.M.; Ahlkvist, J.A.; Perzow, S.E.; McDonald, A. Co-activation of SAM and HPA responses to acute stress: A review of the literature and test of differential associations with preadolescents’ internalizing and externalizing. Dev. Psychobiol. 2019, 61, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Palkovits, M. Catecholamines and stress. IDEGGYÓGYÁSZATI Szle./Clin. Neurosci. 2014, 67, 89–93. [Google Scholar]
- Onyango, A.N. Cellular stresses and stress responses in the pathogenesis of insulin resistance. Oxid. Med. Cell. Longev. 2018, 2018, 4321714. [Google Scholar] [CrossRef]
- Marie, L.S.; Palmiter, R.D. Norepinephrine and epinephrine-deficient mice are hyperinsulinemic and have lower blood glucose. Endocrinology 2003, 144, 4427–4432. [Google Scholar] [CrossRef]
- Hunt, D.G.; Ivy, J.L. Epinephrine inhibits insulin-stimulated muscle glucose transport. J. Appl. Physiol. 2002, 93, 1638–1643. [Google Scholar] [CrossRef]
- Granados, G.; Garay-Sevilla, M.; Malacara, J.; Wróbel-Zasada, K.; Rivera-Cisneros, A. Plasma epinephrine and norepinephrine response to stimuli in autonomic neuropathy of type 2 diabetes mellitus. Acta Diabetol. 2000, 37, 55–60. [Google Scholar] [CrossRef]
- Phadke, D.; Beller, J.P.; Tribble, C. The disparate effects of epinephrine and norepinephrine on hyperglycemia in cardiovascular surgery. Heart Surg. Forum 2018, 21, E522–E526. [Google Scholar] [CrossRef]
- Sherwin, R.S.; Sacca, L. Effect of epinephrine on glucose metabolism in humans: Contribution of the liver. Am. J. Physiol.-Endocrinol. Metab. 1984, 247, E157–E165. [Google Scholar] [CrossRef]
- Rizza, R.; Cryer, P.; Haymond, M.; Gerich, J. Adrenergic mechanisms of catecholamine action on glucose homeostasis in man. Metabolism 1980, 29, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Di Dalmazi, G.; Pagotto, U.; Pasquali, R.; Vicennati, V. Glucocorticoids and type 2 diabetes: From physiology to pathology. J. Nutr. Metab. 2012, 2012, 525093. [Google Scholar] [CrossRef] [PubMed]
- Dragoş, D.; Tănăsescu, M.D. The effect of stress on the defense systems. J. Med. Life 2010, 3, 10. [Google Scholar] [PubMed]
- Fernandez, A.M.; Torres-Alemán, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef]
- Carbone, E.; Borges, R.; Eiden, L.E.; García, A.G.; Hernández-Cruz, A. Chromaffin cells of the adrenal medulla: Physiology, pharmacology, and disease. Compr. Physiol. 2019, 9, 1443–1502. [Google Scholar] [CrossRef]
- Ziegler, M.G.; Elayan, H.; Milic, M.; Sun, P.; Gharaibeh, M. Epinephrine and the metabolic syndrome. Curr. Hypertens. Rep. 2012, 14, 1–7. [Google Scholar] [CrossRef]
- Chernogubova, E.; Cannon, B.; Bengtsson, T. Norepinephrine increases glucose transport in brown adipocytes via β3-adrenoceptors through a cAMP, PKA, and PI3-kinase-dependent pathway stimulating conventional and novel PKCs. Endocrinology 2004, 145, 269–280. [Google Scholar] [CrossRef]
- Nonogaki, K. New insights into sympathetic regulation of glucose and fat metabolism. Diabetologia 2000, 43, 533–549. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383. [Google Scholar] [CrossRef]
- Gędek, A.; Modrzejewski, S.; Materna, M.; Szular, Z.; Wichniak, A.; Mierzejewski, P.; Dominiak, M. Efficacy and Safety of Agomelatine in Depressed Patients with Diabetes: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2024, 25, 12631. [Google Scholar] [CrossRef]
- Bahar, A.; Kashi, Z.; Daneshpour, E.; Akha, O.; Ala, S. Effects of cabergoline on blood glucose levels in type 2 diabetic patients: A double-blind controlled clinical trial. Medicine 2016, 95, e4818. [Google Scholar] [CrossRef]
- Chan, O.; Chan, S.; Inouye, K.; Shum, K.; Matthews, S.G.; Vranic, M. Diabetes impairs hypothalamo-pituitary-adrenal (HPA) responses to hypoglycemia, and insulin treatment normalizes HPA but not epinephrine responses. Diabetes 2002, 51, 1681–1689. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Prediabetes (n = 40) | Prediabetes (n = 40) | Type 2 Diabetes (n = 40) | X2 (df) | p | |
|---|---|---|---|---|---|
| Age (N = 120) | 6.242 (2) | 0.0441 | |||
| 25–35 | 11 | 13 | 4 | ||
| 36–45 | 29 | 27 | 36 | ||
| Gender (N = 120) | 6.07 (2) | 0.0481 | |||
| Male | 15 | 20 | 26 | ||
| Female | 25 | 20 | 14 | ||
| Ethnicity (N = 120) | 2.98 (6) | 0.8115 | |||
| African | 25 | 24 | 28 | ||
| White | 7 | 4 | 3 | ||
| Indian | 8 | 10 | 8 | ||
| Coloured | 0 | 2 | 1 |
| Groups | HbA1c (mmol/mol) | HbA1c % | FBG (mmol/L) |
|---|---|---|---|
| NPD | 36 (4 #) β | 5.4 (0.3 #) β | 5.7 ± 0.24 β |
| PD | 43 (3 #) γ | 6.1 (0.3 #) γ | 6.6 ± 0.38 γ |
| T2D | 75.5 (33.25 #) | 9.3 (4.2 #) | 11.95 ± 3.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosili, P.; Mkhize, B.C.; Ngubane, P.S.; Sibiya, N.H.; Khathi, A. Prediabetes May Alter HPA Axis Activity and Regulation: A Study on Patients with Prediabetes. Int. J. Mol. Sci. 2025, 26, 6231. https://doi.org/10.3390/ijms26136231
Mosili P, Mkhize BC, Ngubane PS, Sibiya NH, Khathi A. Prediabetes May Alter HPA Axis Activity and Regulation: A Study on Patients with Prediabetes. International Journal of Molecular Sciences. 2025; 26(13):6231. https://doi.org/10.3390/ijms26136231
Chicago/Turabian StyleMosili, Palesa, Bongeka Cassandra Mkhize, Phikelelani Sethu Ngubane, Ntethelelo Hopewell Sibiya, and Andile Khathi. 2025. "Prediabetes May Alter HPA Axis Activity and Regulation: A Study on Patients with Prediabetes" International Journal of Molecular Sciences 26, no. 13: 6231. https://doi.org/10.3390/ijms26136231
APA StyleMosili, P., Mkhize, B. C., Ngubane, P. S., Sibiya, N. H., & Khathi, A. (2025). Prediabetes May Alter HPA Axis Activity and Regulation: A Study on Patients with Prediabetes. International Journal of Molecular Sciences, 26(13), 6231. https://doi.org/10.3390/ijms26136231