

Time-Dependent Impact of Betulin and Its Derivatives on IL-8 Expression in Colorectal Cancer Cells with Molecular Docking Studies


Abstract
1. Introduction
2. Results
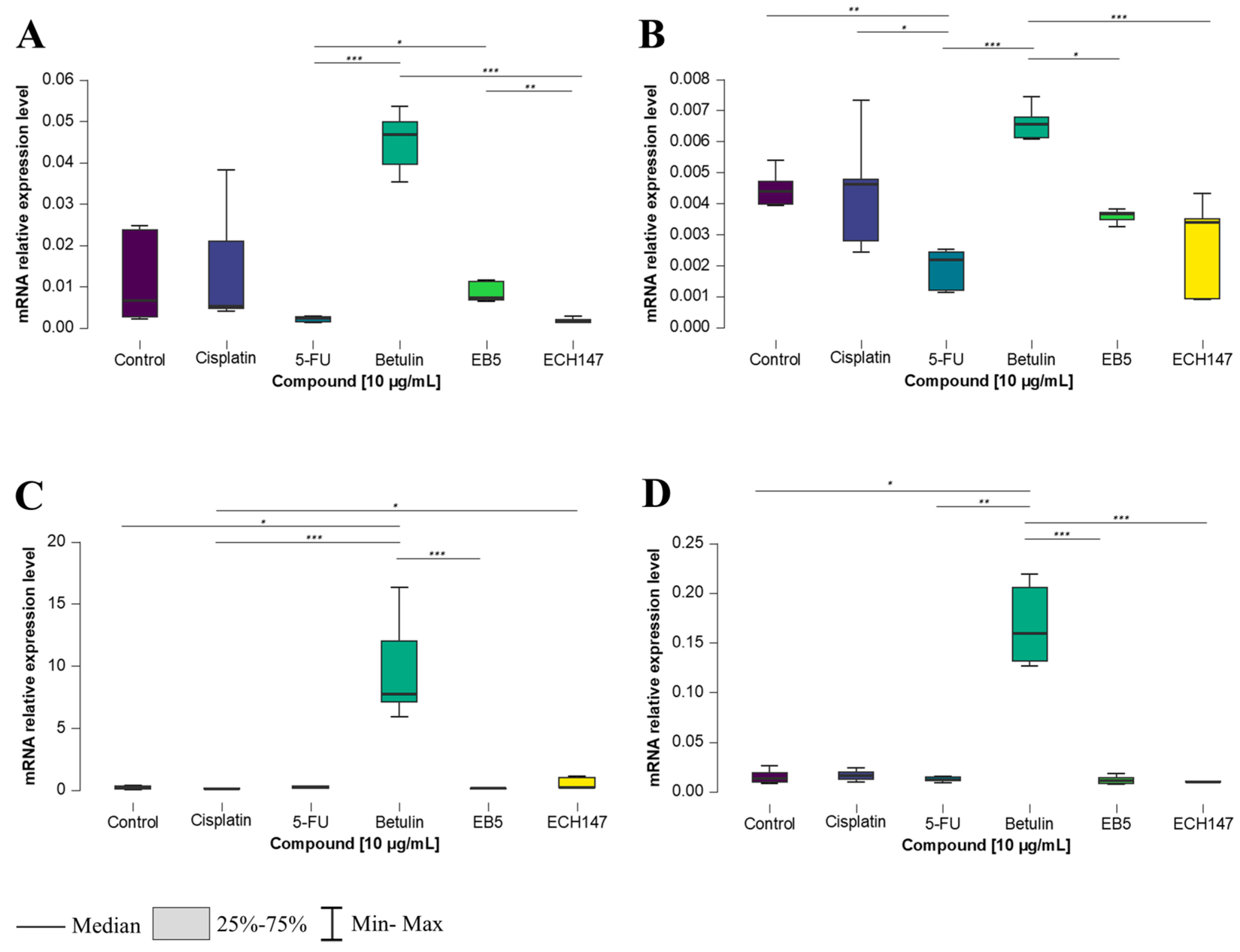
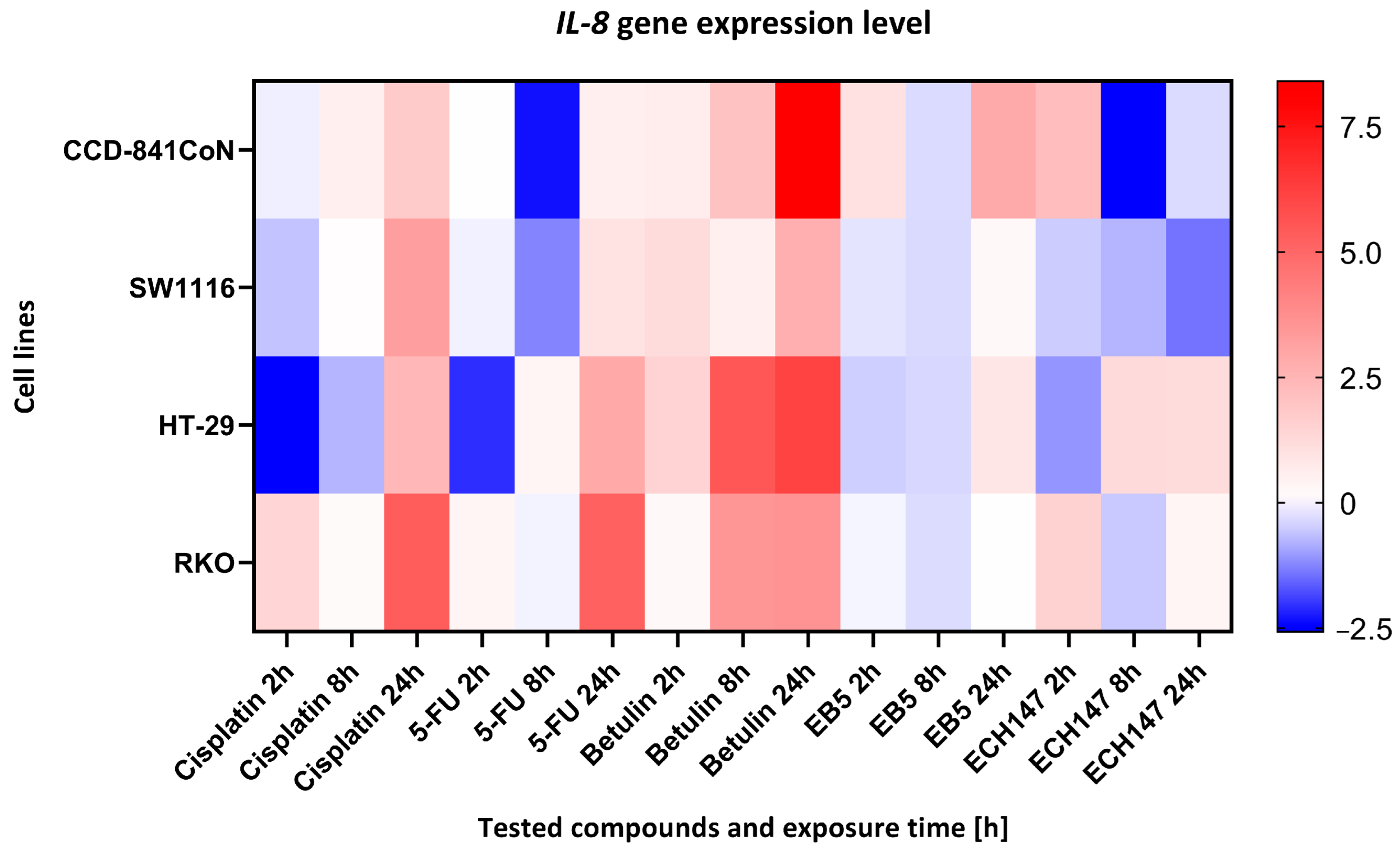
2.1. Temporal Profiling of IL-8 Expression in Colorectal Cancer Cell Lines Treated with Investigated Compounds
2.2. Impact of Exposure Time to Investigated Compounds on IL-8 Gene Expression
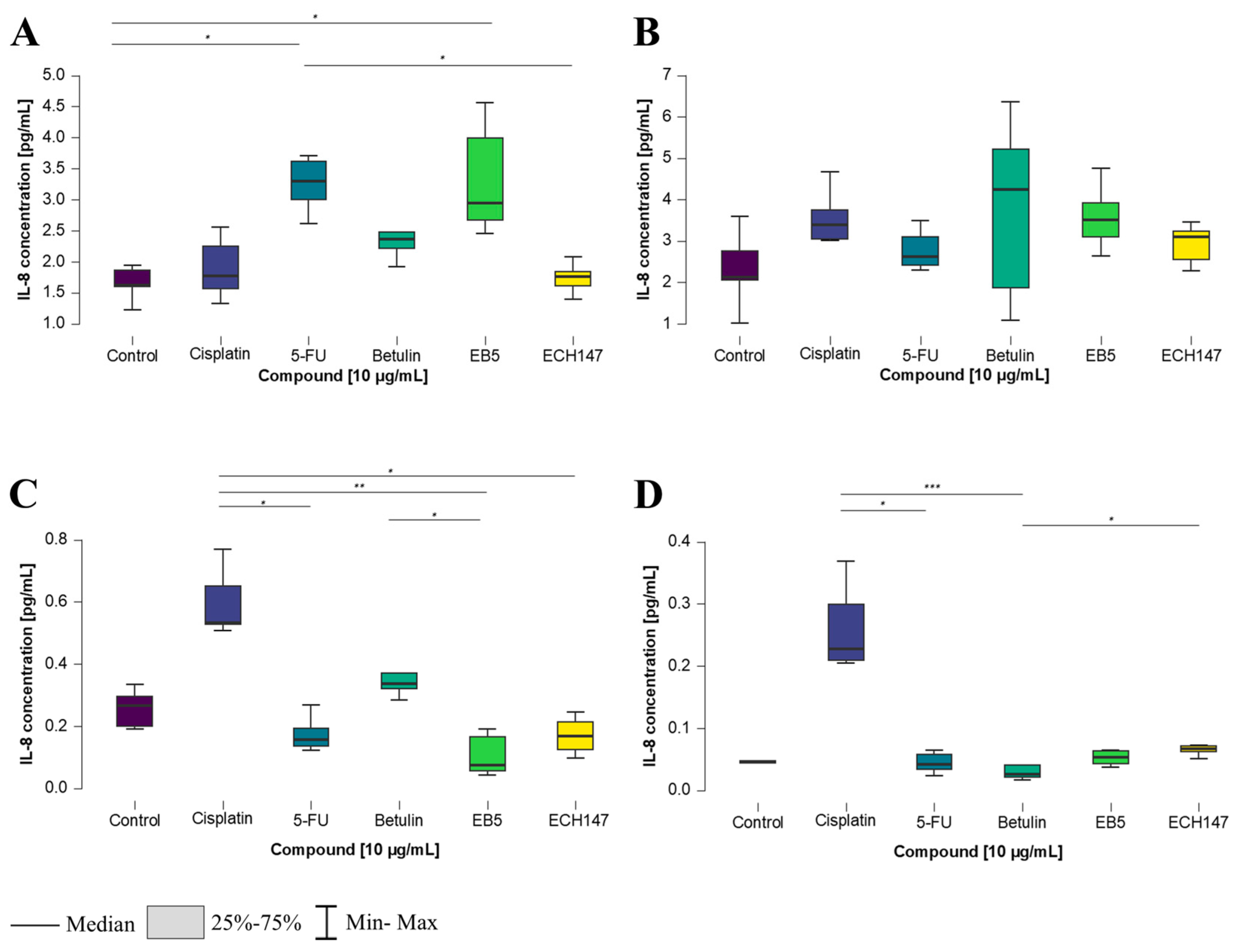
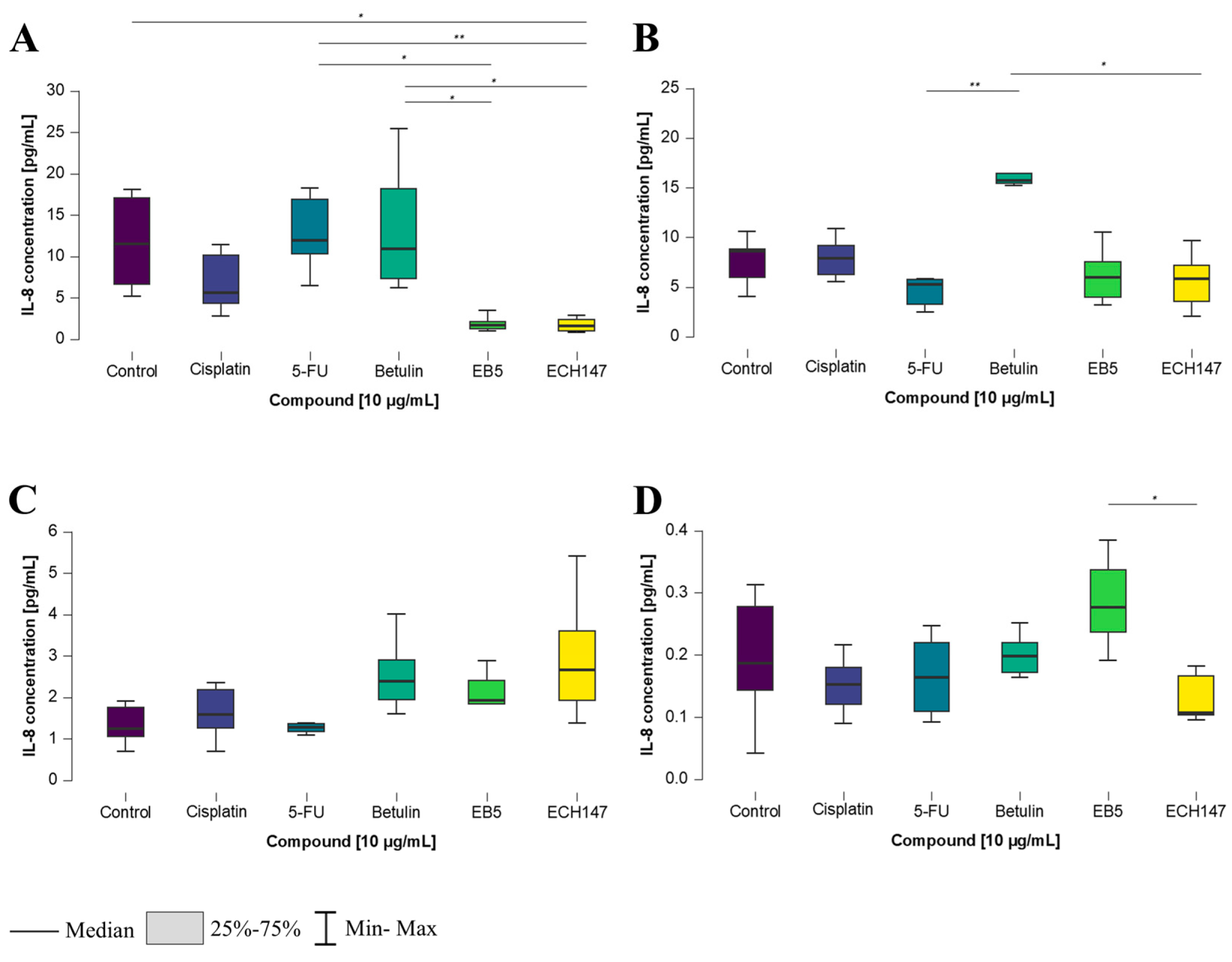
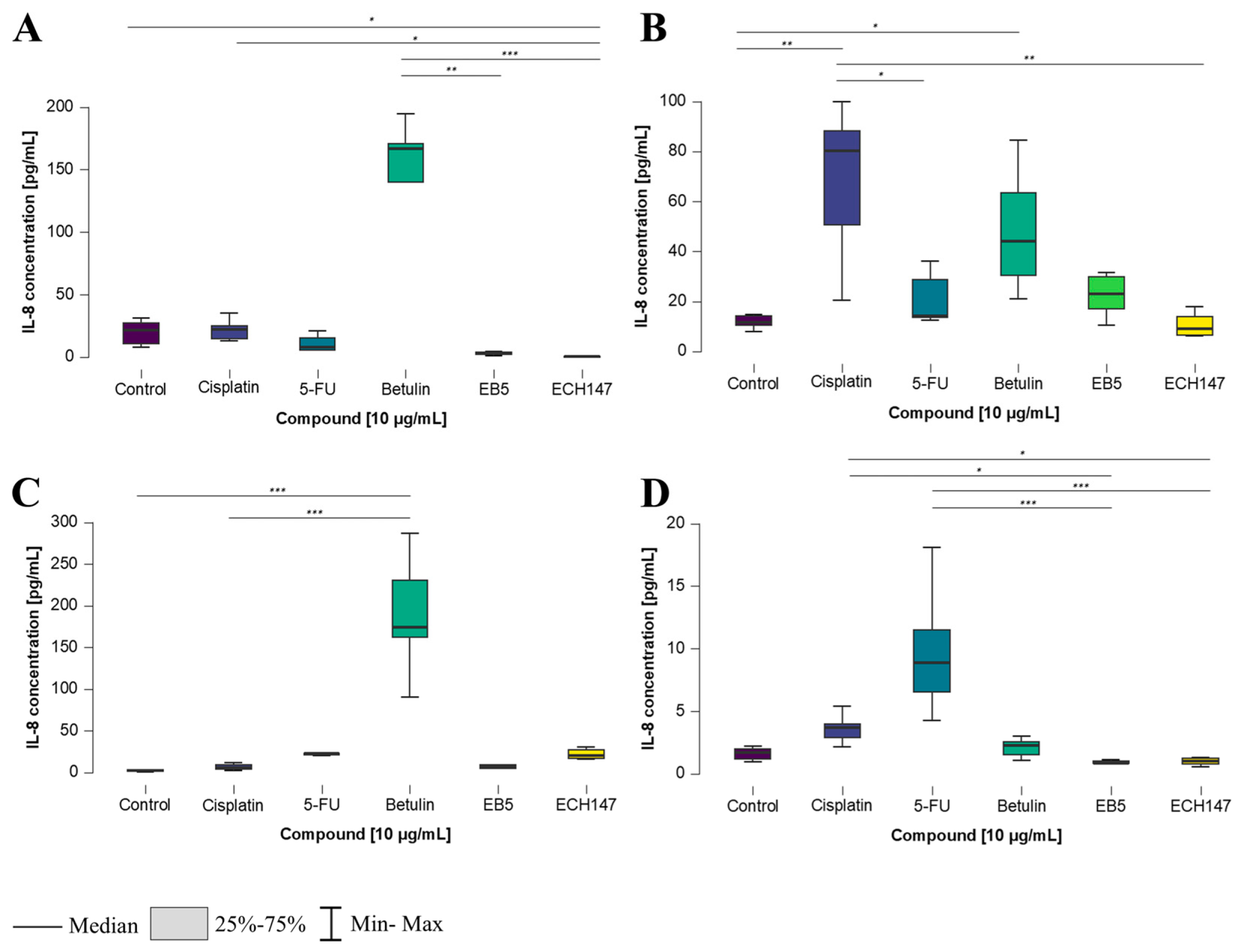
2.3. Time-Dependent Changes in IL-8 Protein Concentration in Cell Culture Medium in Colorectal Cancer Cell Lines
2.4. Time-Dependent Modulation of IL-8 Protein Levels by Investigated Compounds
2.5. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. In Vitro Culture of Colorectal Cancer Cell Lines and Normal Colonocytes
4.2. Cell Culture Treatment and Time Exposure
4.3. Ribonucleic Acid Extraction and the Qualitative and Quantitative Evaluation of Extracts
4.4. Gene-Level Alteration Assessments via RT-qPCR in Real Time
4.5. Evaluation of IL-8 Protein Change in the Cell Culture Medium Using Proximity Ligation Assay Technology (PLA)
4.6. Molecular Docking—In Silico Studies
4.7. Statistical Analysis
5. Limitations and Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-FU | 5-fluorouracil |
| ACTB | β-actin |
| BRAF | V-raf murine sarcoma viral oncogene homolog B |
| BRAF-wt | V-raf murine sarcoma viral oncogene homolog B wild-type |
| CAFs | Cancer-associated fibroblasts |
| CHOP | C/EBP homologous protein |
| CRC | Colorectal cancer |
| CXCL8 | C-X-C motif chemokine ligand 8 |
| E | Glutamic acid |
| EB5 | 28-propynoylbetulin |
| ECH147 | 29-diethoxyphosphoryl-28-propynoylbetulin |
| EGFR | Epidermal growth factor receptor |
| ELISA | Enzyme-linked immunosorbent assay |
| EMEM | Eagle’s minimum essential medium |
| ERK | Extracellular signal-regulated kinase |
| GI | Gastrointestinal |
| IL-8 | Interleukin-8 |
| Log2FC | Log2 fold changes |
| MAPK | Mitogen-activated protein kinase |
| MMPs | Matrix metalloproteinases |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide |
| NGS | Next-generation sequencing |
| NHDF | Normal human dermal fibroblasts |
| PDB | Protein Data Bank |
| PLA | Proximity ligation assay technology |
| PROTAC | Proteolysis-targeting chimera |
| PTEN | Phosphatase and tensin homolog |
| qPCR | Quantitative PCR |
| TAMs | Tumor-associated macrophages |
| V | Valine |
References
- Puzzo, M.; De Santo, M.; Morelli, C.; Leggio, A.; Catalano, S.; Pasqua, L. Colorectal cancer: Current and future therapeutic approaches and related technologies addressing multidrug strategies against multiple level resistance mechanisms. Int. J. Mol. Sci. 2025, 26, 1313. [Google Scholar] [CrossRef] [PubMed]
- Tsukanov, V.V.; Vasyutin, A.V.; Tonkikh, J.L. Risk factors, prevention and screening of colorectal cancer: A rising problem. World J. Gastroenterol. 2025, 31, 98629. [Google Scholar] [CrossRef]
- Fadlallah, H.; El Masri, J.; Fakhereddine, H.; Youssef, J.; Chemaly, C.; Doughan, S.; Abou-Kheir, W. Colorectal cancer: Recent advances in management and treatment. World J. Clin. Oncol. 2024, 15, 1136–1156. [Google Scholar] [CrossRef] [PubMed]
- Swain, J.; Preeti; Mohanty, C.; Bajoria, A.A.; Patnaik, S.; Ward Gahlawat, A.; Nikhil, K.; Mohapatra, S.R. Deciphering the metabolic landscape of colorectal cancer through the lens of AhR-mediated intestinal inflammation. Discov. Oncol. 2025, 16, 275. [Google Scholar] [CrossRef] [PubMed]
- Saoudi González, N.; Ros, J.; Baraibar, I.; Salvà, F.; Rodríguez-Castells, M.; Alcaraz, A.; García, A.; Tabernero, J.; Élez, E. Cetuximab as a key partner in personalized targeted therapy for metastatic colorectal cancer. Cancers 2024, 16, 412. [Google Scholar] [CrossRef]
- Madej, M.; Gola, J.; Chrobak, E. Synthesis, pharmacological properties, and potential molecular mechanisms of antitumor activity of betulin and its derivatives in gastrointestinal cancers. Pharmaceutics 2023, 15, 2768. [Google Scholar] [CrossRef]
- Chrobak, E.; Świtalska, M.; Wietrzyk, J.; Bębenek, E. New difunctional derivatives of betulin: Preparation, characterization and antiproliferative potential. Molecules 2025, 30, 611. [Google Scholar] [CrossRef]
- Meier, C.; Brieger, A. The role of IL-8 in cancer development and its impact on immunotherapy resistance. Eur. J. Cancer. 2025, 218, 115267. [Google Scholar] [CrossRef]
- Bazzichetto, C.; Milella, M.; Zampiva, I.; Simionato, F.; Amoreo, C.A.; Buglioni, S.; Pacelli, C.; Le Pera, L.; Colombo, T.; Bria, E.; et al. Interleukin-8 in colorectal cancer: A systematic review and meta-analysis of its potential role as a prognostic biomarker. Biomedicines 2022, 10, 2631. [Google Scholar] [CrossRef]
- Fellhofer-Hofer, J.; Franz, C.; Vey, J.A.; Kahlert, C.; Kalkum, E.; Mehrabi, A.; Halama, N.; Probst, P.; Klupp, F. Chemokines as prognostic factor in colorectal cancer patients: A systematic review and meta-analysis. Int. J. Mol. Sci. 2024, 25, 5374. [Google Scholar] [CrossRef]
- Li, W.; Chen, F.; Gao, H.; Xu, Z.; Zhou, Y.; Wang, S.; Lv, Z.; Zhang, Y.; Xu, Z.; Huo, J.; et al. Cytokine concentration in peripheral blood of patients with colorectal cancer. Front. Immunol. 2023, 14, 1175513. [Google Scholar] [CrossRef] [PubMed]
- Rzeski, W.; Stepulak, A.; Szymański, M.; Juszczak, M.; Grabarska, A.; Sifringer, M.; Kaczor, J.; Kandefer-Szerszeń, M. Betulin elicits anti-cancer effects in tumour primary cultures and cell lines in vitro. Basic Clin. Pharmacol. Toxicol. 2009, 105, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Boryczka, S.; Bębenek, E.; Wietrzyk, J.; Kempińska, K.; Jastrzębska, M.; Kusz, J.; Nowak, M. Synthesis, structure and cytotoxic activity of new acetylenic derivatives of betulin. Molecules 2013, 18, 4526–4543. [Google Scholar] [CrossRef]
- Król, S.K.; Kiełbus, M.; Rivero-Müller, A.; Stepulak, A. Comprehensive review on betulin as a potent anticancer agent. Biomed. Res. Int. 2015, 2015, 584189. [Google Scholar] [CrossRef]
- Cabaj, J.; Bąk, W.; Wróblewska-Łuczka, P. Anti-cancer effect of betulin and its derivatives, with particular emphasis on the treatment of melanoma. J. Pre-Clin. Clin. Res. 2021, 15, 73–79. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Fang, L.; Tang, X. Angel and devil: The protective immunity and pathogenic inflammation of tissue resident memory T cells in ulcerative colitis. Front. Immunol. 2025, 16, 1518339. [Google Scholar] [CrossRef]
- Li, Q.; Geng, S.; Luo, H.; Wang, W.; Mo, Y.Q.; Luo, Q.; Wang, L.; Song, G.B.; Sheng, J.P.; Xu, B. Signaling pathways involved in colorectal cancer: Pathogenesis and targeted therapy. Signal Transduct. Target Ther. 2024, 9, 266. [Google Scholar] [CrossRef]
- Pecqueux, M.; Brückner, F.; Oehme, F.; Hempel, S.; Baenke, F.; Riediger, C.; Distler, M.; Weitz, J.; Kahlert, C. Preoperative IL-8 levels as prognostic indicators of overall survival: An extended follow-up in a prospective cohort with colorectal liver metastases. BMC Cancer 2024, 24, 90. [Google Scholar] [CrossRef]
- Ahmed, D.; Eide, P.W.; Eilertsen, I.A.; Danielsen, S.A.; Eknæs, A.; Hektoen, M.; Lind, G.E.; Lothe, R.A. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013, 2, e71. [Google Scholar] [CrossRef]
- Madej, M.; Kruszniewska-Rajs, C.; Kimsa-Dudek, M.; Synowiec-Wojtarowicz, A.; Chrobak, E.; Bębenek, E.; Boryczka, S.; Głuszek, S.; Adamska, J.; Kubica, S.; et al. The influence of betulin and its derivatives on selected colorectal cancer cell lines’ viability and their antioxidant systems. Cells 2024, 13, 1368. [Google Scholar] [CrossRef]
- Conciatori, F.; Bazzichetto, C.; Amoreo, C.A.; Sperduti, I.; Donzelli, S.; Diodoro, M.G.; Buglioni, S.; Falcone, I.; Shirasawa, S.; Blandino, G.; et al. BRAF status modulates Interelukin-8 expression through a CHOP-dependent mechanism in colorectal cancer. Commun. Biol. 2020, 3, 546. [Google Scholar] [CrossRef] [PubMed]
- Kaleta-Richter, M.; Aebisher, D.; Jaworska, D.; Czuba, Z.; Cieślar, G.; Kawczyk-Krupka, A. The influence of hypericin-mediated photodynamic therapy on interleukin-8 and -10 secretion in colon cancer cells. Integr. Cancer Ther. 2020, 19, 1534735420918931. [Google Scholar] [CrossRef] [PubMed]
- Lubczyńska, A.; Bębenek, E.; Garncarczyk, A.; Wcisło-Dziadecka, D. Evaluation of the effect of betulin and its alkynyl derivatives on the profile of changes in gene expression of the inflammatory process of colorectal adenocarcinoma cells (HT-29 cell line). Processes 2023, 11, 2676. [Google Scholar] [CrossRef]
- Malicki, S.; Winiarski, M.; Matlok, M.; Kostarczyk, W.; Guzdek, A.; Konturek, P.C. IL-6 and IL-8 responses of colorectal cancer in vivo and in vitro cancer cells subjected to simvastatin. J. Physiol. Pharmacol. 2009, 60, 141–146. [Google Scholar]
- Riekstina, U.; Vitolina, S.; Goluba, K.; Jekabsons, K.; Muceniece, R.; Berzins, R.; Rizhikovs, J.; Godina, D.; Teresko, A.; Paze, A. Effect of betulin colloidal particles on proliferation and cytokine secretion of human skin fibroblasts. Plants 2023, 12, 3088. [Google Scholar] [CrossRef]
- Alshahrani, M.Y.; Alkhathami, A.G.; Almoyad, M.A.A.; Ahmad, M.Z.; Mohanto, S.; Ahmad, W.; Wahab, S. Phytochemicals as potential inhibitors of interleukin-8 for anticancer therapy: In silico evaluation and molecular dynamics analysis. J. Biomol. Struct. Dyn. 2025, 43, 2917–2928. [Google Scholar] [CrossRef]
- Olakowska, E.; Wlaszczuk, A.; Turek, A.; Borecka, A.; Liskiewicz, A.; Wawro, D.; Kasperczyk, J.; Jedrzejowska-Szypulka, H. Effects of 17-β-estradiol released from shape-memory terpolymer rods on sciatic nerve regeneration after injury and repair with chitosan nerve conduit in female rats. J. Appl. Biomed. 2022, 20, 87–97. [Google Scholar] [CrossRef]
- Rech, J.; Wilińska, J.; Turek, A. Application of fibrin in drug technology: Achievements and perspectives. Postępy Hig. Med. Dośw. 2020, 74, 322–330. [Google Scholar] [CrossRef]
- Uner, B.; Akyildiz, E.O.; Kolci, K.; Reis, R. Nanoparticle formulations for intracellular delivery in colorectal cancer therapy. AAPS PharmSciTech 2025, 26, 81. [Google Scholar] [CrossRef]
- Chrobak, E.; Bębenek, E.; Kadela-Tomanek, M.; Latocha, M.; Jelsch, C.; Wenger, E.; Boryczka, S. Betulin phosphonates; synthesis, structure, and cytotoxic activity. Molecules 2016, 21, 1123. [Google Scholar] [CrossRef]
- Kruszniewska-Rajs, C.; Strzałka-Mrozik, B.; Kimsa-Dudek, M.; Synowiec-Wojtarowicz, A.; Chrobak, E.; Bębenek, E.; Boryczka, S.; Głuszek, S.; Gola, J.M. The influence of betulin and its derivatives EB5 and ECH147 on the antioxidant status of human renal proximal tubule epithelial cells. Int. J. Mol. Sci. 2022, 23, 2524. [Google Scholar] [CrossRef] [PubMed]
- Martis, E.A.F.; Téletchéa, S. Ten quick tips to perform meaningful and reproducible molecular docking calculations. PLoS Comput. Biol. 2025, 21, e1013030. [Google Scholar] [CrossRef] [PubMed]
- Serrafi, A.; Wasilewski, A. Synthesis and antimicrobial activity of new betulin derivatives. Sci. Rep. 2025, 15, 17719. [Google Scholar] [CrossRef] [PubMed]
- Tâlvan, C.D.; Budișan, L.; Tâlvan, E.T.; Grecu, V.; Zănoagă, O.; Mihalache, C.; Cristea, V.; Berindan-Neagoe, I.; Mohor, C.I. Serum interleukins 8, 17, and 33 as potential biomarkers of colon cancer. Cancers 2024, 16, 745. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, G.; Luo, W.; Xu, M.; Peng, R.; Du, Z.; Liu, Y.; Bai, Z.; Xiao, X.; Qin, S. PROTAC technology: From drug development to probe technology for target deconvolution. Eur. J. Med. Chem. 2024, 276, 116725. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, X. Key advances and application prospects of PROTAC technologies in the next 5 years. Future Med. Chem. 2025, 17, 987–989. [Google Scholar] [CrossRef]
- Wang, H.; Huang, J.; Fang, X.; Liu, M.; Fan, X.; Li, Y. Advances in next-generation sequencing (NGS) applications in drug discovery and development. Expert Opin. Drug Discov. 2025, 20, 537–550. [Google Scholar] [CrossRef]
- Niazi, S.K.; Mariam, Z. Artificial intelligence in drug development: Reshaping the therapeutic landscape. Ther. Adv. Drug Saf. 2025, 16, 20420986251321704. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ΔG [kcal/mol] |
|---|---|
| EB5 | −8.7 |
| Betulin | −8.0 |
| ECH147 | −7.9 |
| 5-FU | −4.4 |
| Parameter | EB5 | Betulin | ECH147 | 5-FU |
|---|---|---|---|---|
| Binding Energy [kcal/mol] | −8.7 | −8.0 | −7.9 | −4.4 |
| Hydrogen Bonds | 1 | 1 | 1 | 2 |
| Residues H-bonded | Val27 | Leu25 | Arg68 | Glu38; Asn36 |
| Hydrophobic Interactions | 8 | 5 | 7 | 4 |
| Carbon–Hydrogen Bonds | 0 | 0 | 1 | 1 |
| Van der Waals Interactions | 4 | 2 | 3 | 7 |
| Alkyl Interactions | 4 | 3 | 3 | 0 |
| Π–Sulfur Interactions | 0 | 0 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madej, M.; Halama, A.; Chrobak, E.; Gola, J.M. Time-Dependent Impact of Betulin and Its Derivatives on IL-8 Expression in Colorectal Cancer Cells with Molecular Docking Studies. Int. J. Mol. Sci. 2025, 26, 6186. https://doi.org/10.3390/ijms26136186
Madej M, Halama A, Chrobak E, Gola JM. Time-Dependent Impact of Betulin and Its Derivatives on IL-8 Expression in Colorectal Cancer Cells with Molecular Docking Studies. International Journal of Molecular Sciences. 2025; 26(13):6186. https://doi.org/10.3390/ijms26136186
Chicago/Turabian StyleMadej, Marcel, Adrianna Halama, Elwira Chrobak, and Joanna Magdalena Gola. 2025. "Time-Dependent Impact of Betulin and Its Derivatives on IL-8 Expression in Colorectal Cancer Cells with Molecular Docking Studies" International Journal of Molecular Sciences 26, no. 13: 6186. https://doi.org/10.3390/ijms26136186
APA StyleMadej, M., Halama, A., Chrobak, E., & Gola, J. M. (2025). Time-Dependent Impact of Betulin and Its Derivatives on IL-8 Expression in Colorectal Cancer Cells with Molecular Docking Studies. International Journal of Molecular Sciences, 26(13), 6186. https://doi.org/10.3390/ijms26136186