
Multi-Time Point Transcriptome Analysis and Functional Validation Revealed Bol4CL41 Negatively Regulates Black Rot Resistance in Cabbage

 ,
, 
 , , ,
, , ,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Comparison of the Phenotypes of MY and LY After Inoculation
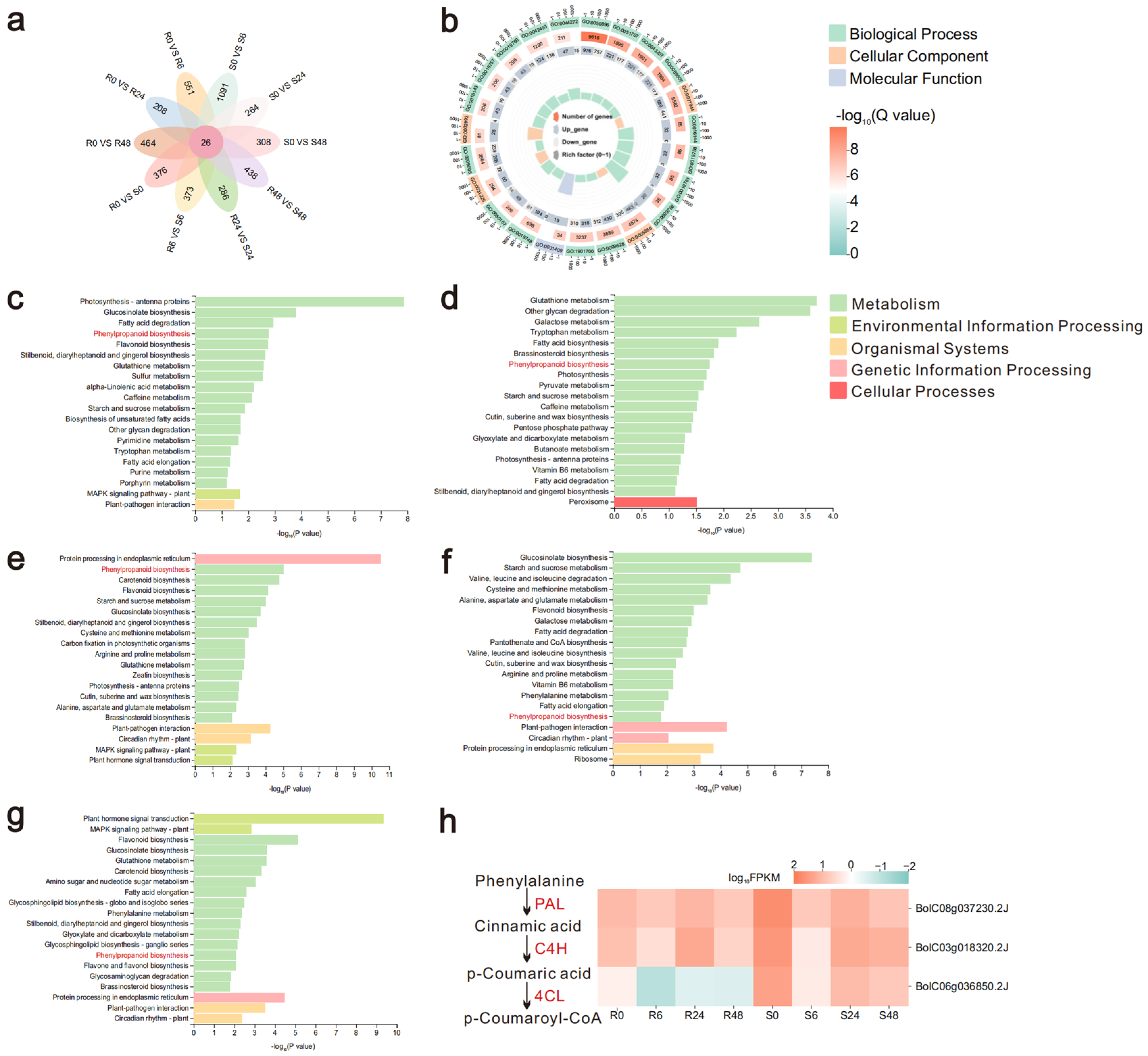
2.2. Functional Annotation of DEGs and Expression of Phenylpropanoid Biosynthesis Genes
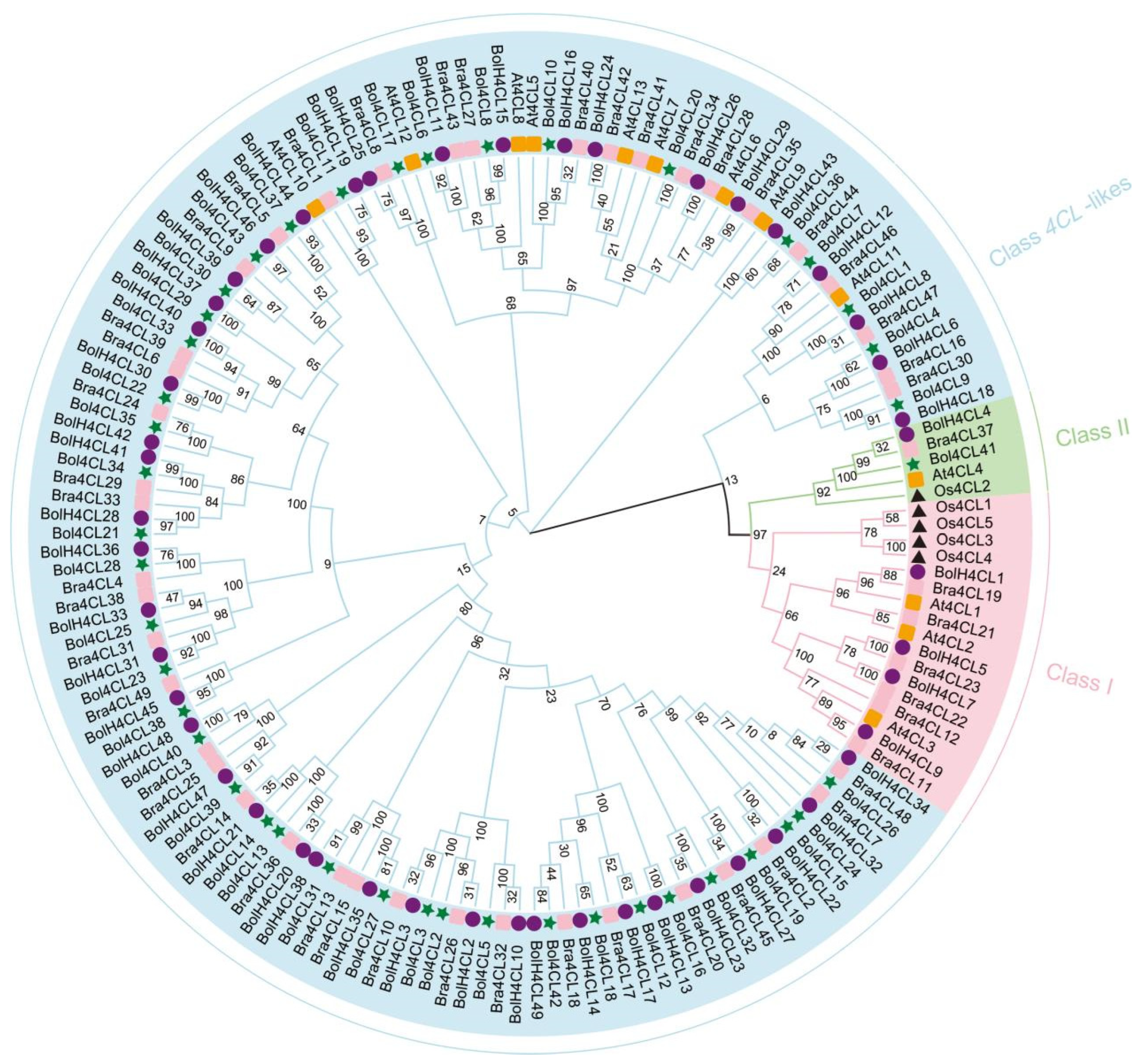
2.3. Systematic Identification and Phylogenetic Analysis Reveal Functional Divergence in the Cabbage 4CL Gene Family
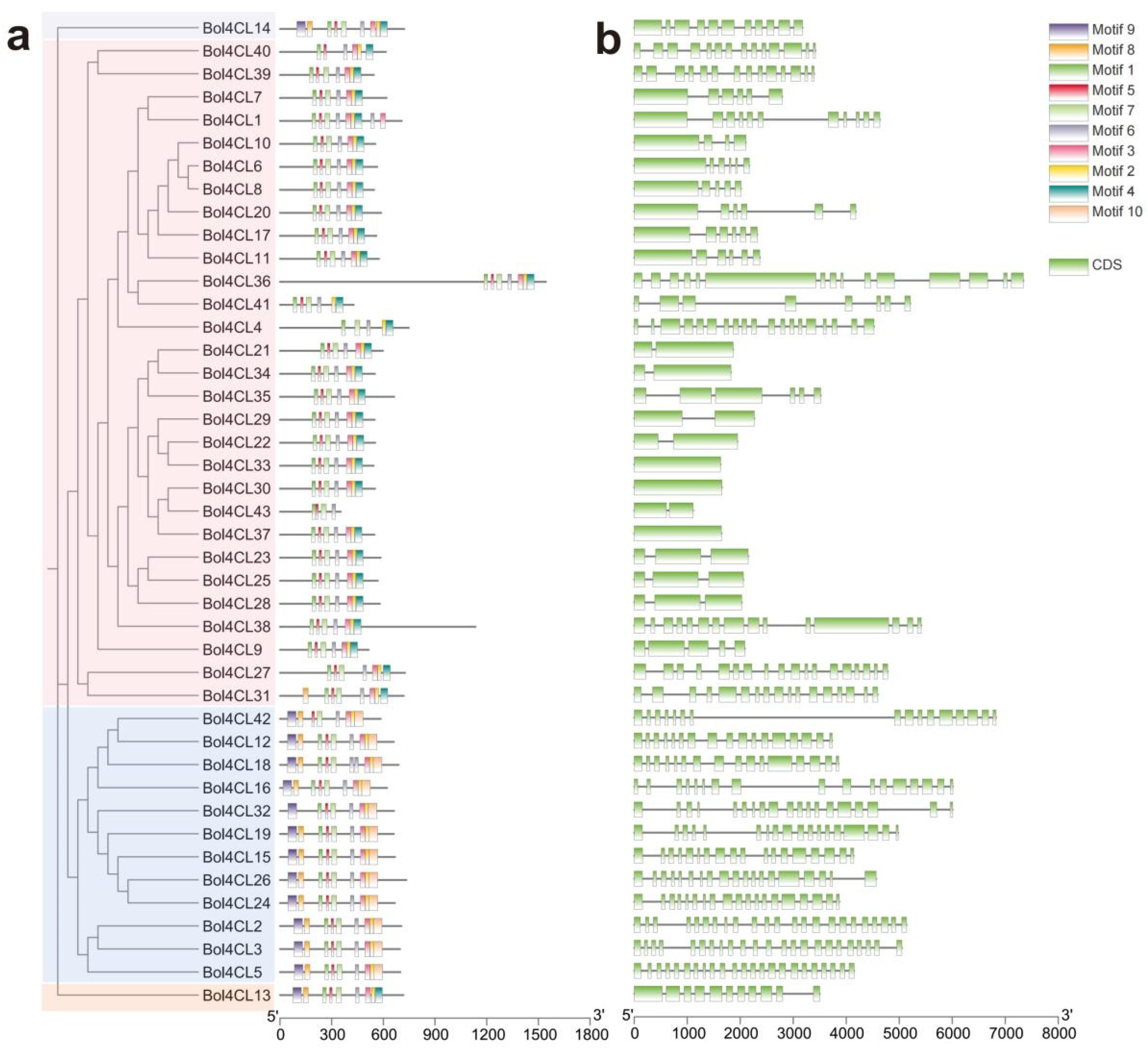
2.4. Gene Structure and Conserved Motif of Bol4CLs
2.5. Chromosomal Location and Synteny Analysis of Bol4CLs
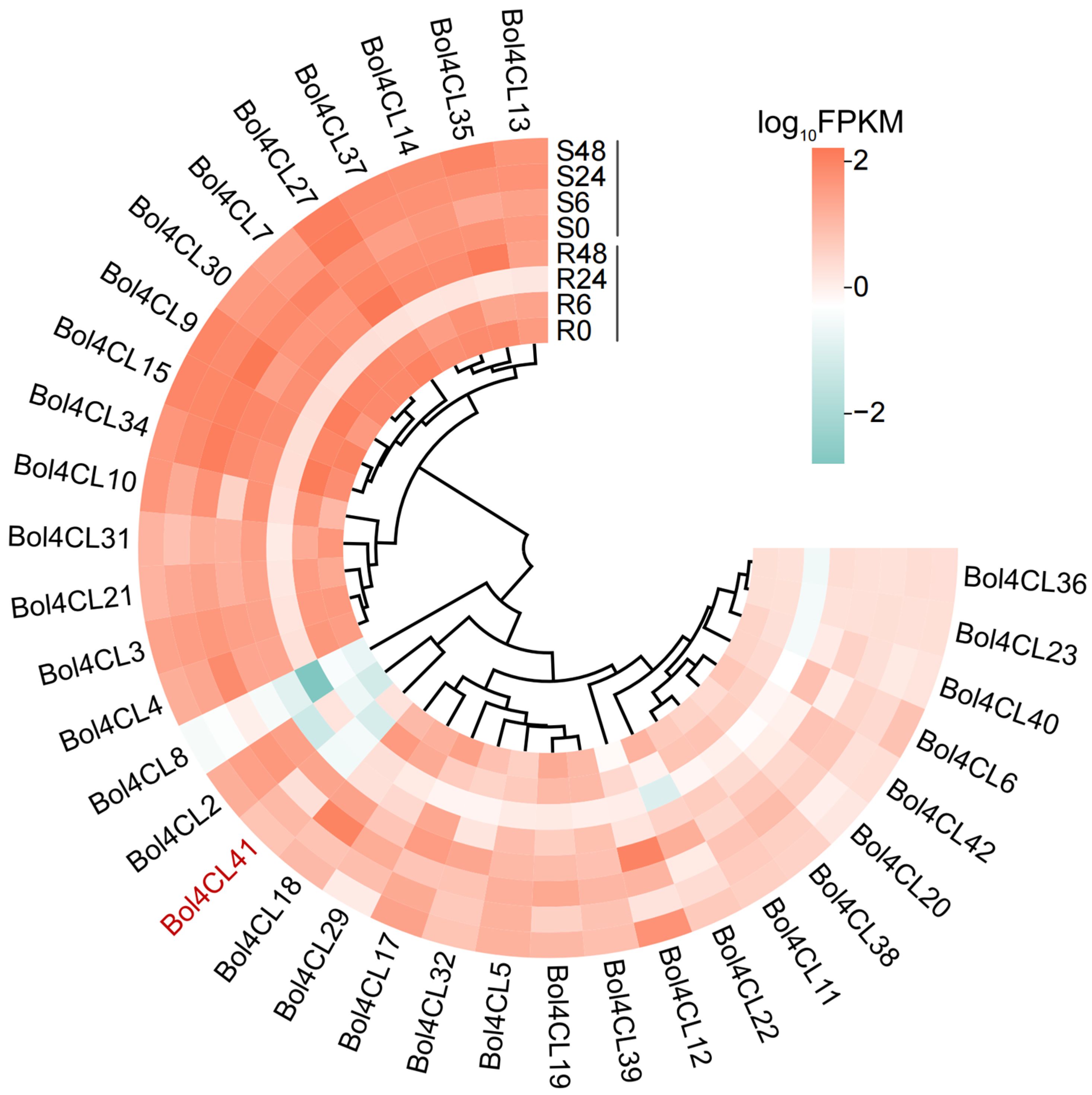
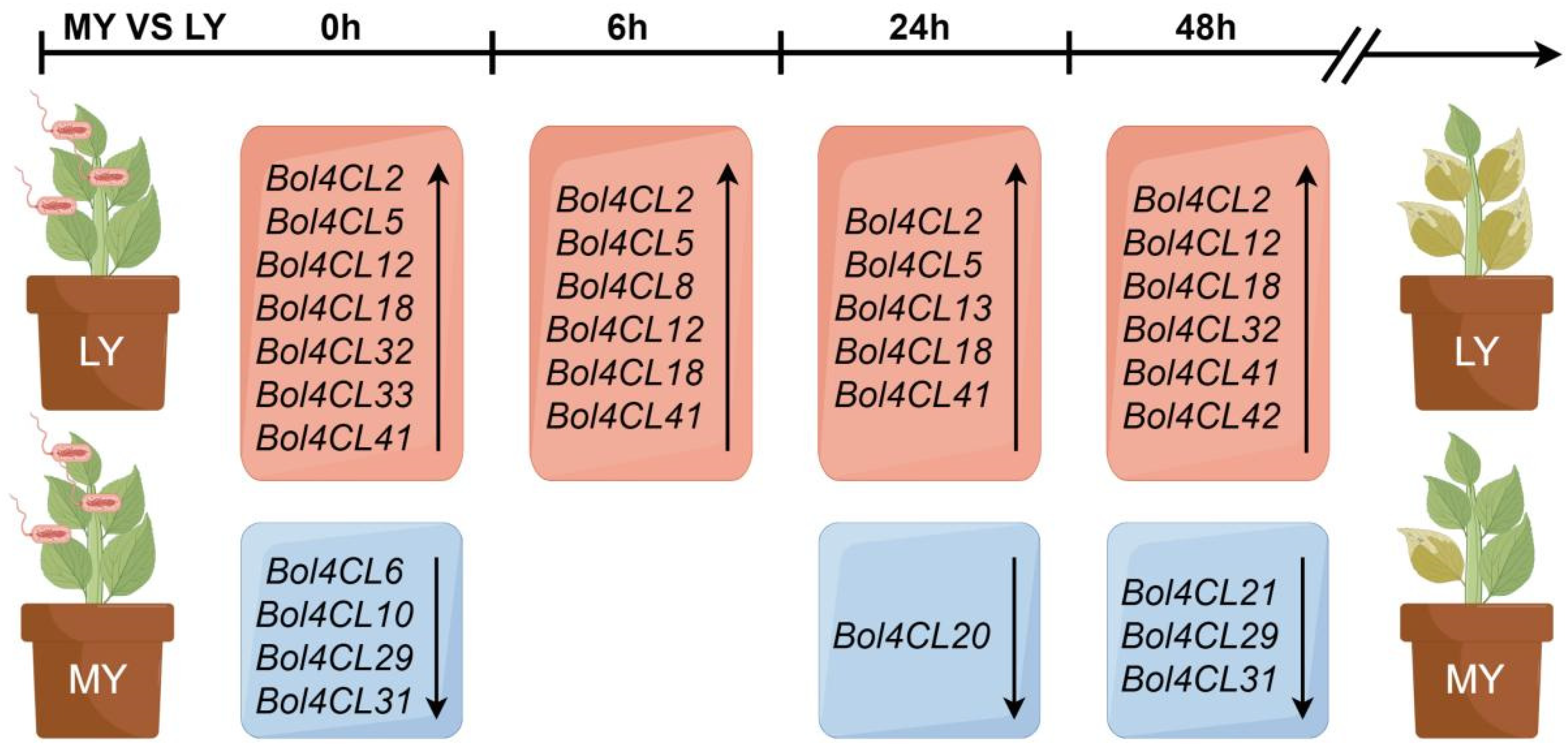
2.6. Gene Expression Profiles Heatmap of Bol4CL at Different Stages After Inoculation
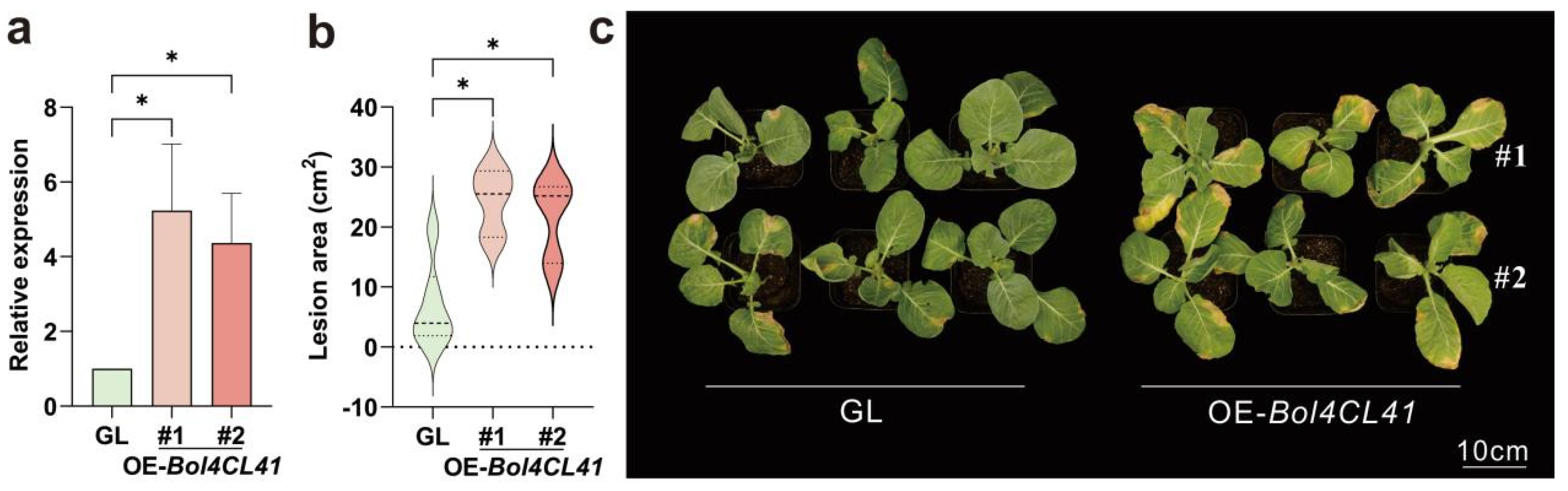
2.7. Bol4CL41 Negatively Regulates BR Resistance in Cabbage
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Inoculation Method
4.2. RNA-Seq and Transcriptome Analysis of Cabbage in Response to BR
4.3. Genome-Wide Identification of 4CL in Different Species
4.4. Construction of Phylogenetic Trees and Analysis of Protein Characteristics
4.5. Gene Structural Analysis of Bol4CL Genes
4.6. Chromosome Location and Synteny Analysis
4.7. Construction Gene Expression Profiles Heatmap
4.8. Vector Construction, Transgenic Plant Generation and Inoculation Treatments
4.9. Disease Mechanism Diagram Drawing, Resistance Evaluation and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vicente, J.; Holub, E. Anthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to brassica crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef]
- Williams, P.H. Black rot: Continuing threat to world crucifers. Plant Dis. 1980, 64, 736–742. [Google Scholar] [CrossRef]
- Jensen, B.; Vicente, J.; Manandhar, H.; Roberts, S. Occurrence and diversity of Xanthomonas campestris pv. campestris in vegetable Brassica fields in Nepal. Plant Dis. 2010, 94, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Fargier, E.; Saux, M.; Manceau, C. A multilocus sequence analysis of Xanthomonas campestris reveals a complex structure within crucifer-attacking pathovars of this species. Syst. Appl. Microbiol. 2011, 34, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Rathaur, P.; Vicente, J. Characterization, genetic diversity and distribution of Xanthomonas campestris pv. campestris races causing black rot disease in cruciferous crops of india. Plant Pathol. 2016, 65, 1411–1418. [Google Scholar] [CrossRef]
- Huang, D.; Li, C.; Si, J.; Ren, X.; Song, H. Research progress on physiological race division and disease resistance identification of cabbage black rot. Veg. China 2011, 1, 6–10. [Google Scholar]
- Alvarez, A.M.; Benedict, A.A.; Mizumoto, C.Y.; Hunter, J.E.; Gabriel, D.W. Serological, pathological and genetic diversity among strains of Xanthomonas campestris infecting crucifers. Phytopathology 1994, 84, 1449–1457. [Google Scholar] [CrossRef]
- Dong, N.; Lin, H. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, T.; Luo, W.; Xu, J.; Liu, J.; Wan, D. Identification of 4CL genes in desert poplars and their changes in expression in response to salt stress. Genes 2015, 6, 901–917. [Google Scholar] [CrossRef]
- Ragg, H.; Kuhn, D.N.; Hahlbrock, K. Coordinated regulation of 4-coumarate:CoA ligase and phenylalanine ammonia-lyase mRNAs in cultured plant cells. J. Biol. Chem. 1981, 256, 10061–10065. [Google Scholar] [CrossRef]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef]
- Sun, H.; Li, Y.; Feng, S.; Zou, W.; Guo, K.; Fan, C.; Si, S.; Peng, L. Analysis of five rice 4-coumarate:coenzyme A ligase enzyme activity and stress response for potential roles in lignin and flavonoid biosynthesis in rice. Biochem. Biophys. Res. Commun. 2013, 430, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Lavhale, S.G.; Kalunke, R.M.; Giri, A.P. Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants. Planta 2018, 248, 1063–1078. [Google Scholar] [CrossRef]
- Kumar, A.; Ellis, B.E. 4-Coumarate:CoA ligase gene family in Rubus idaeus: cDNA structures, evolution, and expression. Plant Mol. Biol. 2003, 51, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zuo, D.; Zhang, X.; Li, H.; Ye, H.; Zhang, N.; Li, M.; Dang, M.; Geng, F.; Zhou, H.; et al. Genome-wide identification analysis of the 4-Coumarate: CoA ligase (4CL) gene family expression profiles in Juglans regia and its wild relatives J. Mandshurica resistance and salt stress. BMC Plant Biol. 2024, 24, 211. [Google Scholar] [CrossRef]
- Gális, I.; Šimek, P.; Narisawa, T.; Sasaki, M.; Horiguchi, T.; Fukuda, H.; Matsuoka, K. A novel R2R3 MYB transcription factor NtMYBJS1 is a methyl jasmonate-dependent regulator of phenylpropanoid-conjugate biosynthesis in tobacco. Plant J. 2006, 46, 573–592. [Google Scholar] [CrossRef] [PubMed]
- Ehlting, J.; Büttner, D.; Wang, Q.; Douglas, C.J.; Somssich, I.E.; Kombrink, E. Three 4-coumarate:coenzyme A ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [Google Scholar] [CrossRef]
- Ran, F.; Xiang, C.; Wang, C.; Zang, Y.; Liu, L.; Wu, S.; Wang, C.; Cai, J.; Wang, D.; Min, Y. Identification of the 4CL family in cassava (Manihot esculenta Crantz) and expression pattern analysis of the Me4CL32 gene. Biochem. Biophys. Res. Commun. 2024, 735, 150731. [Google Scholar] [CrossRef]
- Shinde, B.A.; Dholakia, B.B.; Hussain, K.; Panda, S.; Meir, S.; Rogachev, I.; Aharoni, A.; Giri, A.P.; Kamble, A.C. Dynamic metabolic reprogramming of steroidal glycol-alkaloid and phenylpropanoid biosynthesis may impart early blight resistance in wild tomato (Solanum arcanum Peralta). Plant Mol. Biol. 2017, 95, 411–423. [Google Scholar] [CrossRef]
- Shaw, R.K.; Shen, Y.; Wang, J.; Sheng, X.; Zhao, Z.; Yu, H.; Gu, H. Advances in Multi-Omics Approaches for Molecular Breeding of Black Rot Resistance in Brassica oleracea L. Front. Plant Sci. 2021, 2, 742553. [Google Scholar] [CrossRef]
- Ma, H.; Kong, C.; Deng, S.; Zhao, T.; Ji, J.; Wang, Y.; Zhang, Y.; Zhuang, M.; Yang, L.; Marina, L.; et al. Resistance screening of cabbage to black rot and inheritance pattern analysis. Sci. Hortic. 2025, 345, 114129. [Google Scholar] [CrossRef]
- Kong, C.; Passo, V.H.D.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Vicente, J.G.; Lv, H. Complete Genome Sequence of Strain WHRI 3811 Race 1 of Xanthomonas campestris pv. campestris, the causal agent of black rot of cruciferous Vegetables. Mol. Plant Microbe Interact. 2019, 32, 1571–1573. [Google Scholar] [CrossRef]
- Ohta, H.; Hattori, T. Bacteria sensitive to nutrient broth medium in terrestrial environments. Soil Sci. Plant Nutr. 1980, 26, 99–107. [Google Scholar] [CrossRef]
- Kong, C.; Chen, G.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Ji, J.; Fang, Z.; Lv, H. Germplasm screening and inheritance analysis of resistance to cabbage black rot in a worldwide collection of cabbage (Brassica oleracea var. capitata) resources. Sci. Hortic. 2021, 288, 110234. [Google Scholar] [CrossRef]
- Putri, G.H.; Anders, S.; Pyl, P.T.; Pimanda, J.E.; Zanini, F. Analysing high-throughput sequencing data in Python with HTSeq 2.0. Bioinformatics 2022, 38, 2943–2945. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 5, 550. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenführer, J. Gene set enrichment analysis with topGO. Bioconduct. Improv. 2009, 27, 776. [Google Scholar]
- Alexa, A.; Rahnenführer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.; Han, Y.; He, Q. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Wulf, A.; Manthey, K.; Doll, J.; Perlick, A.M.; Linke, B.; Bekel, T.; Meyer, F.; Franken, P.; Küster, H.; Krajinski, F. Transcriptional Changes in Response to Arbuscular Mycorrhiza Development in the Model Plant Medicago truncatula. Mol. Plant-Microbe Interact. 2003, 16, 306–314. [Google Scholar] [CrossRef]
- Singh, A.; Shukla, N.; Kabadwal, B.C.; Tewari, A.K.; Kumar, J. Review on Plant-Trichoderma-Pathogen Interaction. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2382–2397. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, E.; Liu, Y.; Xu, Z.; Hui, M.; Zhang, X.; Cai, M. Transcriptome analysis of two lines of Brassica oleracea in response to early infection with Xanthomonas campestris pv. Campestris. Can. J. Plant Pathol. 2021, 43, 127–139. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, C.; Yang, L.; Zhang, Y.; Wang, Y.; Fang, Z.; Lv, H. Genome-Wide Identification, Expression Profile of the TIFY Gene Family in Brassica oleracea var. capitata, and Their Divergent Response to Various Pathogen Infections and Phytohormone Treatments. Genes 2020, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, M.; Cartea, M.E.; Rodríguez, V.M.; Velasco, P. ‘Omic’ profiling of B. oleracea challenged with Xanthomonas campestris pv. Campestris. Acta Hortic. 2018, 1202, 63–68. [Google Scholar] [CrossRef]
- Tortosa, M.; Cartea, M.E.; Velasco, P.; Soengas, P.; Rodriguez, V.M. Calcium-signaling proteins mediate the plant transcriptomic response during a well-established Xanthomonas campestris pv. campestris infection. Hortic. Res. 2019, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Nan, X.; Li, W.; Mao, J.; Chen, B. Comprehensive genomic identification and expression analysis 4CL gene family in apple. Gene 2023, 858, 147197. [Google Scholar] [CrossRef]
- Rao, G.; Pan, X.; Xu, F.; Zhang, Y.; Cao, S.; Jiang, X.; Lu, H. Divergent and Overlapping Function of Five 4-Coumarate/Coenzyme A Ligases from Populus tomentosa. Plant Mol. Biol. Rep. 2015, 33, 841–854. [Google Scholar] [CrossRef]
- Uhlmann, A.; Ebel, J. Molecular cloning and expression of 4-coumarate:coenzyme A ligase, an enzyme involved in the resistance response of soybean (Glycine max L.) against pathogen attack. Plant Physiol. 1993, 102, 1147–1156. [Google Scholar] [CrossRef]
- Voo, K.S.; Whetten, R.W.; O’Malley, D.M.; Sederoff, R.R. 4-coumarate:coenzyme a ligase from loblolly pine xylem. Isolation, characterization, and complementary DNA cloning. Plant Physiol. 1995, 108, 85–97. [Google Scholar] [CrossRef]
- Hu, W.; Kawaoka, A.; Tsai, C.J.; Lung, J.; Osakabe, K.; Ebinuma, H.; Chiang, V.L. Compartmentalized expression of two structurally and functionally distinct 4-coumarate:CoA ligase genes in aspen (Populus tremuloides). Proc. Natl. Acad. Sci. USA 1998, 95, 5407–5412. [Google Scholar] [CrossRef]
- Harding, S.A.; Leshkevich, J.; Chiang, V.L.; Tsai, C.J. Differential substrate inhibition couples kinetically distinct 4-coumarate: Coenzyme a ligases with spatially distinct metabolic roles in quaking aspen. Plant Physiol. 2002, 128, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, Z.; Ding, Y.; Guo, M. Characterization of HSP70 family in watermelon (Citrullus lanatus): Identification, structure, evolution, and potential function in response to ABA, cold and drought stress. Front. Genet. 2023, 14, 1201535. [Google Scholar] [CrossRef]
- Douglas, C.; Hoffmann, H.; Schulz, W.; Hahlbrock, K. Structure and elicitor or u.v.-light-stimulated expression of two 4-coumarate: CoA ligase genes in parsley. EMBO J. 1987, 6, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Trezzini, G.F.; Horrichs, A.; Somssich, I.E. Isolation of putative defense-related genes from Arabidopsis thaliana and expression in fungal elicitor-treated cells. Plant Mol. Biol. 1993, 21, 385–389. [Google Scholar] [CrossRef]
- Becker-André, M.; Schulze-Lefert, P.; Hahlbrock, K. Structural comparison, modes of expression, and putative cis-acting elements of the two 4-coumarate: CoA ligase genes in potato. J. Biol. Chem. 1991, 266, 8551–8559. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, E.; Kruger-Lebus, S.; Hahlbrock, K. Temporal and Spatial Patterns of Gene Expression around Sites of Attempted Fungal Infection in Parsley Leaves. Plant Cell 1989, 1, 993–1001. [Google Scholar] [CrossRef]
- Lee, D.; Ellard, M.; Wanner, L.A.; Davis, K.R.; Douglas, C.J. The Arabidopsis thaliana 4-coumarate:CoA ligase (4CL) gene: Stress and developmentally regulated expression and nucleotide sequence of its cDNA. Plant Mol. Biol. 1995, 28, 871–884. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.; Deng, S.; Kong, C.; Zhang, Y.; Zhao, T.; Ji, J.; Wang, Y.; Zhang, Y.; Zhuang, M.; Yang, L.; et al. Multi-Time Point Transcriptome Analysis and Functional Validation Revealed Bol4CL41 Negatively Regulates Black Rot Resistance in Cabbage. Int. J. Mol. Sci. 2025, 26, 6179. https://doi.org/10.3390/ijms26136179
Ma H, Deng S, Kong C, Zhang Y, Zhao T, Ji J, Wang Y, Zhang Y, Zhuang M, Yang L, et al. Multi-Time Point Transcriptome Analysis and Functional Validation Revealed Bol4CL41 Negatively Regulates Black Rot Resistance in Cabbage. International Journal of Molecular Sciences. 2025; 26(13):6179. https://doi.org/10.3390/ijms26136179
Chicago/Turabian StyleMa, Hongxue, Siping Deng, Congcong Kong, Yulun Zhang, Tong Zhao, Jialei Ji, Yong Wang, Yangyong Zhang, Mu Zhuang, Limei Yang, and et al. 2025. "Multi-Time Point Transcriptome Analysis and Functional Validation Revealed Bol4CL41 Negatively Regulates Black Rot Resistance in Cabbage" International Journal of Molecular Sciences 26, no. 13: 6179. https://doi.org/10.3390/ijms26136179
APA StyleMa, H., Deng, S., Kong, C., Zhang, Y., Zhao, T., Ji, J., Wang, Y., Zhang, Y., Zhuang, M., Yang, L., Lebedeva, M., Taranov, V., Artemyeva, A. M., Fang, Z., Yu, J., Hu, Z., & Lv, H. (2025). Multi-Time Point Transcriptome Analysis and Functional Validation Revealed Bol4CL41 Negatively Regulates Black Rot Resistance in Cabbage. International Journal of Molecular Sciences, 26(13), 6179. https://doi.org/10.3390/ijms26136179