
Discovery of Novel Genes Encoding Antimicrobial Peptides from the Pedobacter silvilitoris Genome with Broad-Spectrum Antimicrobial Activity


Abstract
1. Introduction
2. Results
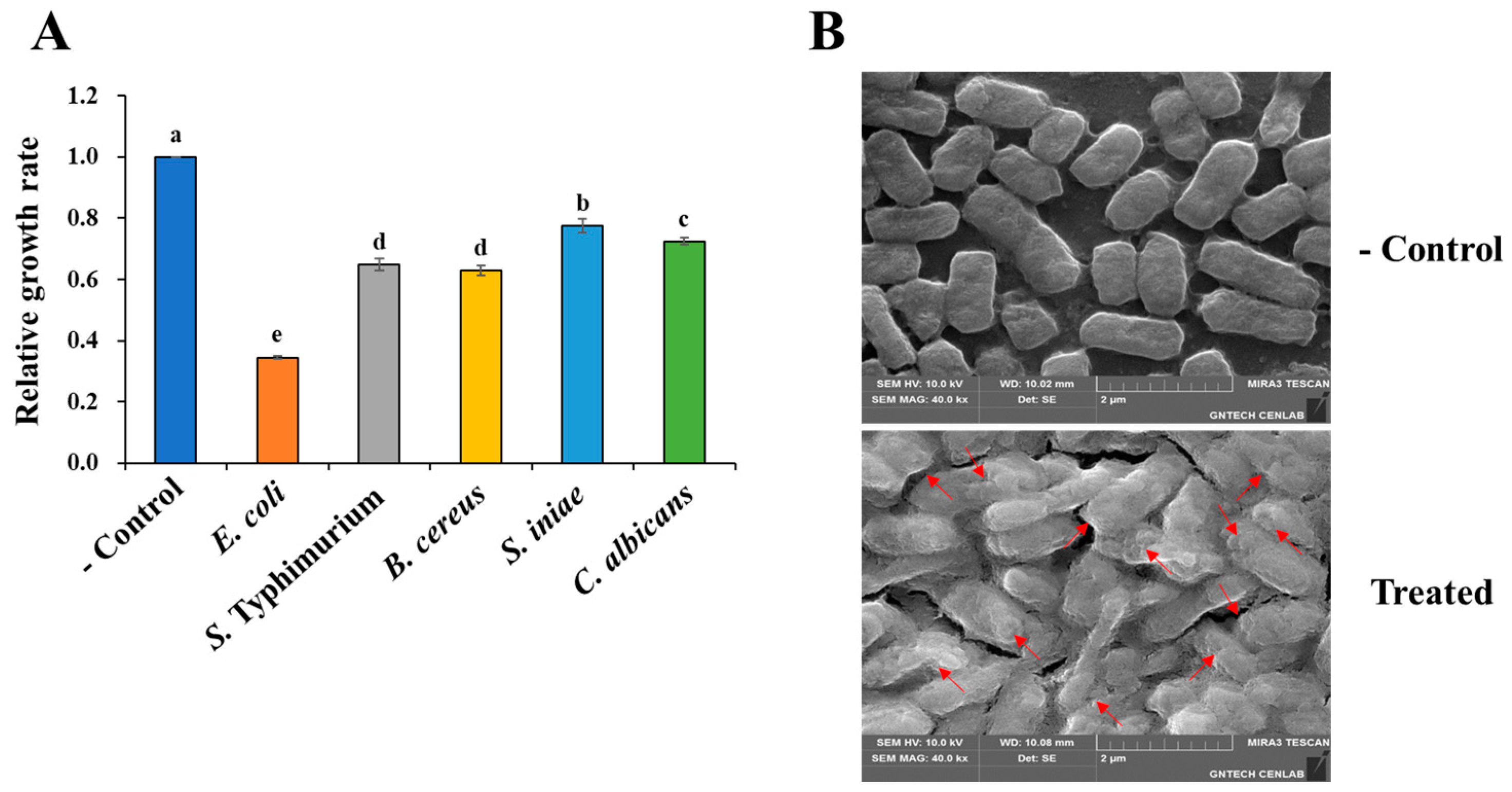
2.1. The Cell-Free Supernatant from P. silvilitoris Exhibits Antimicrobial Activity Against Various Pathogens
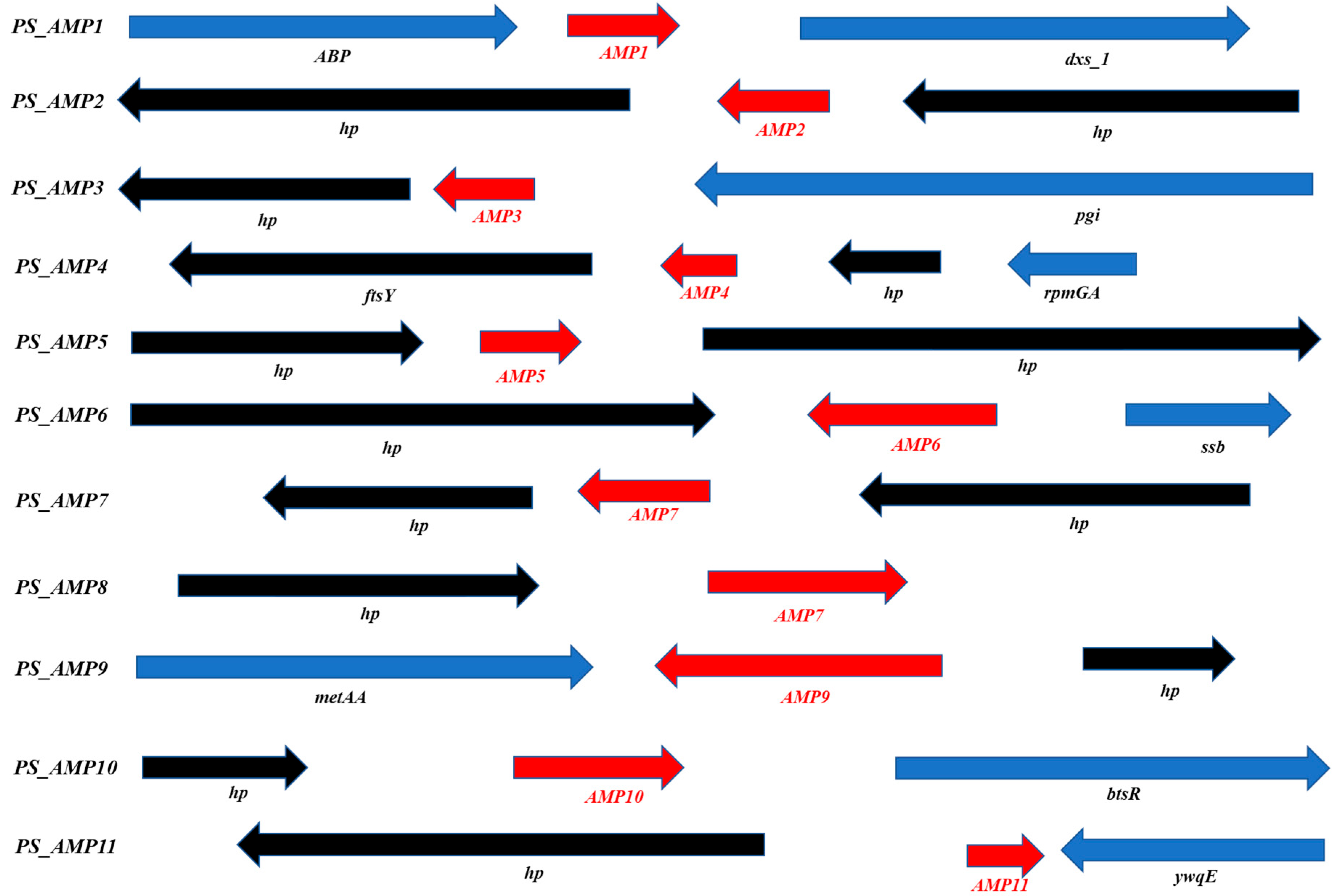
2.2. The P. silvilitoris Genome Includes Various AMP Genes
2.3. Transcriptomic Results of AMP Candidates from the P. silvilitoris Genome
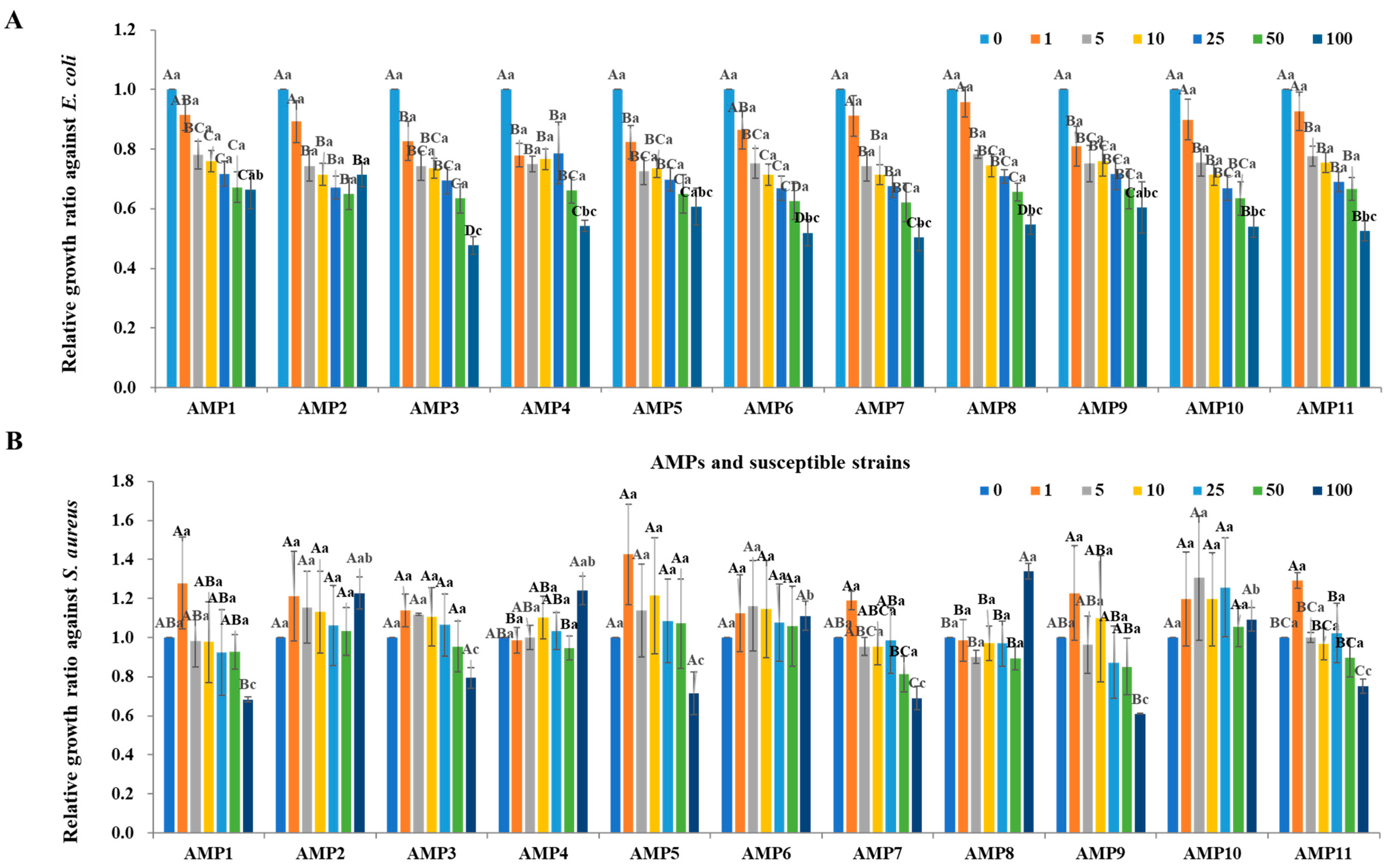
2.4. Antimicrobial Activity of Cloned AMPs from the P. silvilitoris Genome
2.5. Antimicrobial Activity of Synthetic AMPs Originated from the P. silvilitoris Genome
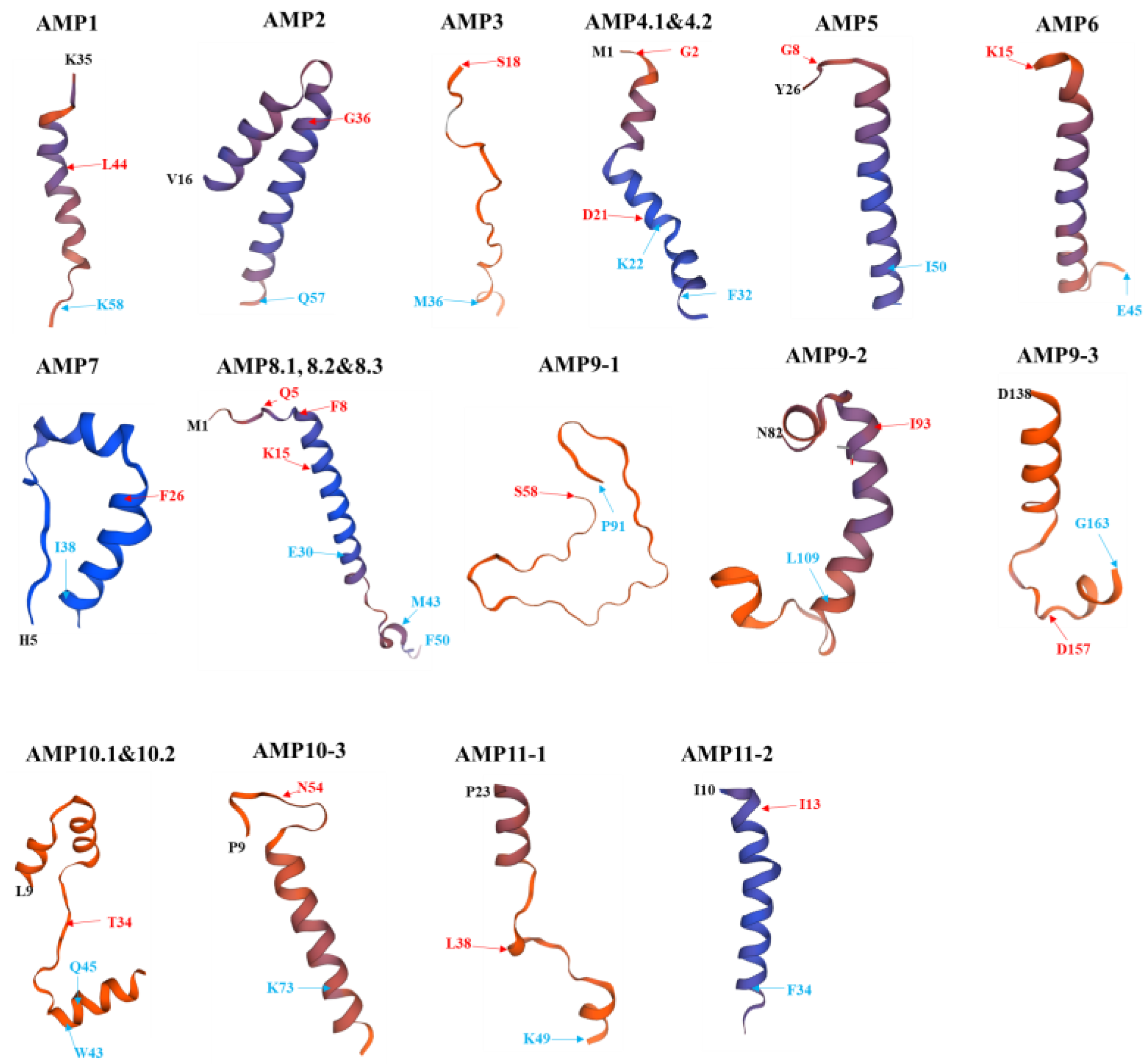
2.6. Structural Models of PS_AMPs
3. Discussion
4. Materials and Methods
4.1. Medium and Bacterial Strains Employed for This Study
4.2. P. silvilitoris Genome Sequencing
4.3. Bioinformatic Analysis for Identification of AMPs from the P. silvilitoris Genome
4.4. Exploration of AMPs Gene Expression Through Transcriptomic Analysis
4.5. P. silvilitoris AMPs Gene Cloning and Peptide Synthesis
4.6. Evaluation of Antimicrobial Activity of Cell-Free Supernatants and Synthetic Peptides
A: the treated sample A600 value
B: non-treated (control) A600 value
4.7. Scanning Electron Microscopy (SEM)
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMP | Antimicrobial peptide |
| SEM | Scanning electron microscopy |
References
- Hancock, R.E.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, D.; Zhang, H.; Wang, Z.; Liu, Y. Evolutionary trajectory of bacterial resistance to antibiotics and antimicrobial peptides in Escherichia coli. mSystems 2025, 10, e01700-24. [Google Scholar] [CrossRef]
- Antoshina, D.V.; Balandin, S.V.; Ovchinnikova, T.V. Structural features, mechanisms of action, and prospects for practical application of class II bacteriocins. Biochemistry 2022, 87, 1387–1403. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef]
- Baquero, F.; Lanza, V.F.; Baquero, M.-R.; Del Campo, R.; Bravo-Vázquez, D.A. Microcins in Enterobacteriaceae: Peptide antimicrobials in the eco-active intestinal chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef]
- Marković, K.G.; Grujović, M.Ž.; Koraćević, M.G.; Nikodijević, D.D.; Milutinović, M.G.; Semedo-Lemsaddek, T.; Djilas, M.D. Colicins and microcins produced by Enterobacteriaceae: Characterization, mode of action, and putative applications. Int. J. Environ. Res. Public Health 2022, 19, 11825. [Google Scholar] [CrossRef]
- Xu, D.; Lu, W. Defensins: A double-edged sword in host immunity. Front. Immunol. 2020, 11, 764. [Google Scholar] [CrossRef]
- Kumaresan, V.; Kamaraj, Y.; Subramaniyan, S.; Punamalai, G. Understanding the dynamics of human defensin antimicrobial peptides: Pathogen resistance and commensal induction. Appl. Biochem. Biotechnol. 2024, 196, 6993–7024. [Google Scholar] [CrossRef]
- Park, S.; Jung, Y.-T.; Park, J.-M.; Won, S.-M.; Yoon, J.-H. Pedobacter silvilitoris sp. nov., isolated from wood falls. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 4, 1284–1289. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, J.; Srivastava, P. Molecular basis of stationary phase survival and applications. Front. Microbiol. 2017, 8, 2000. [Google Scholar] [CrossRef]
- Hengge, R. Stationary-phase gene regulation in Escherichia coli. EcoSal Plus 2011, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Cheng, W. The mechanism of bacterial resistance and potential bacteriostatic strategies. Antibiotics 2022, 11, 1215. [Google Scholar] [CrossRef]
- Singh, G.; Rana, A.; Smriti. Decoding antimicrobial resistance: Unraveling molecular mechanisms and targeted strategies. Arch. Microbiol. 2024, 206, 280. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.A. Understanding and overcoming antibiotic resistance. PLoS Biol. 2017, 15, e2003775. [Google Scholar] [CrossRef]
- Tuerkova, A.; Kabelka, I.; Králová, T.; Sukeník, L.; Pokorná, Š.; Hof, M.; Vácha, R. Effect of helical kink in antimicrobial peptides on membrane pore formation. Elife 2020, 9, e47946. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.N.; Humblot, V.; Haney, E.F.; Ribeiro, S.M.; Hancock, R.E.; Ladram, A.; Franco, O.L. EcDBS1R6: A novel cationic antimicrobial peptide derived from a signal peptide sequence. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2020, 1864, 129633. [Google Scholar] [CrossRef]
- Chaudhari, A.A.; Jawale, C.V.; Kim, S.W.; Lee, J.H. Construction of a Salmonella Gallinarum ghost as a novel inactivated vaccine candidate and its protective efficacy against fowl typhoid in chickens. Vet. Res. 2012, 43, 44. [Google Scholar] [CrossRef]
- Assoni, L.; Milani, B.; Carvalho, M.R.; Nepomuceno, L.N.; Waz, N.T.; Guerra, M.E.S.; Converso, T.R.; Darrieux, M. Resistance mechanisms to antimicrobial peptides in gram-positive bacteria. Front. Microbiol. 2020, 11, 593215. [Google Scholar] [CrossRef]
- Kordi, M.; Talkhounche, P.G.; Vahedi, H.; Farrokhi, N.; Tabarzad, M. Heterologous production of antimicrobial peptides: Notes to consider. Protein J. 2024, 43, 129–158. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Bulaon, C.J.I.; Malla, A.; Phoolcharoen, W. Biotechnological insights on the expression and production of antimicrobial peptides in plants. Molecules 2021, 26, 4032. [Google Scholar] [CrossRef] [PubMed]
- Gagat, P.; Ostrówka, M.; Duda-Madej, A.; Mackiewicz, P. Enhancing antimicrobial peptide activity through modifications of charge, hydrophobicity, and structure. Int. J. Mol. Sci. 2024, 25, 10821. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, E.H.; Lohan, S.; Ghaffari, T.; Gupta, S.; Tiwari, R.K.; Parang, K. Membrane-active cyclic amphiphilic peptides: Broad-spectrum antibacterial activity alone and in combination with antibiotics. J. Med. Chem. 2022, 65, 15819–15839. [Google Scholar] [CrossRef] [PubMed]
- Nagib, M.; Sayed, A.M.; Korany, A.H.; Abdelkader, K.; Shari, F.H.; Mackay, W.G.; Rateb, M.E. Human Defensins: Structure, Function, and Potential as Therapeutic Antimicrobial Agents with Highlights Against SARS CoV-2. Probiotics Antimicrob. Proteins 2024, 17, 1563–1583. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, M.; Wang, Z.; Liu, Z.; Chen, S.; Li, X.; Shi, Y.; Hu, H. Discovery of novel antibacterial agent for the infected wound treatment: All-hydrocarbon stapling optimization of LL-37. Theranostics 2024, 14, 1181. [Google Scholar] [CrossRef]
- Payasi, A.; Yadav, M.K.; Chaudhary, S.; Aggarwal, A. Evaluating nephrotoxicity reduction in a novel polymyxin B formulation: Insights from a 3D kidney-on-a-chip model. Antimicrob. Agents Chemother. 2024, 68, e00219-24. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.R.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Habor Laboratory: New York, NY, USA, 1989. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMPs Name | ORF Sequences * | Homologous AMPs | Synthesized AMPs | ||||
|---|---|---|---|---|---|---|---|
| Homolog Name | Identities/Positives (%) | Amino Acid Seq. | Amino Acid No. | pI | MW | ||
| PS_AMP1 | MSQFVLMGLFFFPLVVSVLTIKDVFENQGLDNGKKLMWIATIILLPLVGAIIYFFFGKSKQL | LAMP_Experimental_3178 | 47/73 | LLPLVGAIIYFFFGK | 15 | 8.59 | 1698.12 |
| PS_AMP2 | MEDKFDYQNYNWLYYVIGLVCGLLVGYAVEGHIITGAILGLLTGGLFQFLINKSREQEA | Melittin | 46/67 | GAILGLLTGGLFQFLINKSREQ | 22 | 8.75 | 2375.80 |
| PS_AMP3 | MMKKPIISLFVGGLLSLSLLNACDSNKNAEPEVTQMETLQDSLEQQETALNKDIENLQKSMQDLDQEFKKNN | Granulosusin-E1 | 40/66 | MKKPIISLFVGGLLSLSLLNACDSNKNAEPEVTQM | 35 | 5.93 | 3762.45 |
| PS_AMP4 | MGLFDFFKKKDKTTDEQEALDKGLEKTKDNFFPK | Pleurain-K1 | 58/83 | DKGLEKTKDNFF (PS_AMP4.1) | 12 | 6.12 | 1441.60 |
| Aurein-1.2 | 43/57 | GLFDFFKKKDKTTDEQEALDK (PS_AMP4.2) | 21 | 4.93 | 2503.79 | ||
| PS_AMP5 | MSQRKPYNHNTSYGRRKIREQSEKYYEGLPPEEKSSHNLLGILIMVIICIIMFLILGADGFLKWGTR | HsAp3 | 39/65 | GLPPEEKSSHNLLGILIMVIICI | 23 | 5.4 | 2490.06 |
| PS_AMP6 | MTFQDRKDKGKDWDKVKEKREELHERDEKIANEPKAVLKQDQETEQEKEE | AVP1829 | 27/71 | KDKGKDWDKVKEKREELHERDEKIANEPKAVLKQDQETEQE | 41 | 5.09 | 4978.46 |
| PS_AMP7 | MNIYHFFPLPHPSPRNIIWQAKNRWFVKEVLPQLKEIISVHI | Kasstasin | 62/77 | FVKEVLPQLKEII | 13 | 6.14 | 1555.92 |
| PS_AMP8 | MARSQETFSKKENEKKRLKKRQEKQQKKEERRESSTGGGLENMMAYVDEFGNITDTPPDPIRKKKEIDASTIEIGIPKQEAEDLTATKKGKIEFFNDSKGFGFIKEDNTQEKFFVHVNGLTEEVREGDKVSFELEKGLKGMNAVNVKRI | Magainin-2 | 28/64 | KKRLKKRQEKQQKKEERRESSTGGGLENMMAYVDEF (PS_AMP8.1) | 36 | 9.77 | 4343.95 |
| AVP1828 | 26/56 | QETFSKKENEKKRLKKRQEKQQKKEERRESSTGGGLENM (PS_AMP8.2) | 39 | 9.9 | 4709.28 | ||
| Alpha-benincasin | 30/83 | FSKKENEKKRLKKRQEKQQKKEE (PS_AMP8.3) | 23 | 10.09 | 2975.44 | ||
| PS_AMP9 | MLQNLINLVKENAGESIINNPSVPNEKNDKAIEVTGNSIIETLKNAVAGGDISQLKSLFNQEPEAAKTSTLANNAQNNVVANLIAKIGLSPEIATKIASTVVPLVMSKLVSKTNDPNNSSFNIQDILGNLAGGNGGFDVGSILSGINSANDKNKKGDVLGKLGGLFGK | Opistoporin-1 | 44/54 | SQLKSLFNQEPEAAKTSTLANNAQNNVVANLIAKIGLSP (PS_AMP9.1) | 39 | 8.22 | 4095.62 |
| D83 | 59/76 | IATKIASTVVPLVMSKL (PS_AMP9.2) | 13 | 10 | 1771.23 | ||
| Ponericin-G7 | 75/83 | DVLGKLGGLFGK (PS_AMP9.3) | 12 | 8.59 | 1203.45 | ||
| PS_AMP10 | MFFVTYNSLNKAIRYFFVDPYIEQYEKENGNFATWWKTQGTPWQQFINPIAFSNAIKGANMVVWIAVTIKLFKLWYQKRKAALYAELNFLKSQVHPHFLFNTLNNLYALTLNQSPKAPQVVMGLSEILRYMLYECDKETISLKKN | Pep-1-K | 67/67 | TWWKTQGTPWQQ (PS_AMP10.1) | 12 | 8.41 | 1546.70 |
| PK-12 | 70/70 | TWWKTQGTPW (PS_AMP10.2) | 10 | 8.41 | 1290.44 | ||
| Brevinin-2-RA17 peptide precursor | 55/70 | NAIKGANMVVWIAVTIKLFK (PS_AMP10.3) | 20 | 10.3 | 2216.76 | ||
| PS_AMP11 | MEFILPGIGIPEIITILIALIIPIAIITMIVLYFRKNLKLKKEQIELLKKIANK | E17KGG | 53/76 | LKLKKEQIELLKKIANK (PS_AMP11.1) | 17 | 10 | 2037.56 |
| CAMPST453 | 38/58 | ITILIALIIPIAIITMIVLYFRKNLK (PS_AMP11.2) | 26 | 10.29 | 2997.89 | ||
| Sample | Total Reads | Mapped Reads | Mapping Rate % | Count (>0) | Exp. (>1) |
|---|---|---|---|---|---|
| Pedobacter_1 | 51,645,840 | 16,626,807 | 32.19 | 3736 | 3736 |
| Pedobacter_2 | 57,465,126 | 17,422,082 | 30.32 | 3737 | 3737 |
| AMPs Name | Pedobacter_1c | Pedobacter_2c | Pedobacter_1e | Pedobacter_2e | Fold Change (PS1/PS2) | p Values |
|---|---|---|---|---|---|---|
| AMP1 | 3698.7 | 5874.8 | 344.072 | 218.014 | 1.578211 | 0.7023 |
| AMP2 | 5050.79 | 5853.11 | 469.85 | 217.209 | 2.163124 | 0.1024 |
| AMP3 | 4591.43 | 3140.14 | 427.118 | 116.531 | 3.665274 | 0.0015 |
| AMP4 | 4352.97 | 9476.85 | 404.935 | 351.686 | 1.151411 | 0.18465 |
| AMP5 | 1007.97 | 3463.72 | 93.766 | 128.539 | 0.729475 | 0.93455 |
| AMP6 | 4059.62 | 3269.95 | 377.646 | 121.348 | 3.112091 | 0.50425 |
| AMP7 | 2923.71 | 9205.92 | 271.978 | 341.632 | 0.796114 | 0.18795 |
| AMP8 | 18734.59 | 38260.64 | 1742.786 | 1419.853 | 1.227441 | 0.8038 |
| AMP9 | 1869.67 | 1455.25 | 173.926 | 54.004 | 3.220613 | 0.016 |
| AMP10 | 400.45 | 1727.74 | 37.252 | 64.117 | 0.581 | 0.42205 |
| AMP11 | 808.69 | 4104.77 | 75.228 | 152.328 | 0.493855 | 0.87705 |
| Forward Primers | Reverse Primers | ||
|---|---|---|---|
| Name | Nucleotide Seq. (5′ → 3′) | Name | Nucleotide Seq. (5′ → 3′) |
| PS_AMP1-F | ATTGCCCGTTCCCTGTTAAG | PS_AMP1-R | TTGCACCAAAATGACCTCCG |
| PS_AMP2-F | CCAAGGCAATGATCTGTCAC | PS_AMP2-R | GAGCGAGCCTGTTAATTTAGG |
| PS_AMP3-F | TCCACCTATCGCTTACCTTC | PS_AMP3-R | ATCGCTCCAATCCTTAACGC |
| PS_AMP4-F | TAGCGCTTGCACCTCTTCTC | PS_AMP4-R | TCACCTAAAACAGGTGCTTACT |
| PS_AMP5-F | TGGTTATATGGCTCGGGAAC | PS_AMP5-R | TGGATTGTCAGCATTGTCGG |
| PS_AMP6-F | AACAGTGCTACTCGTTTCGC | PS_AMP6-R | GTAAGTGCTACTCCGCTACA |
| PS_AMP7-F | TTAACCTGCTGTGTGCTAGG | PS_AMP7-R | TTTGCTAGCCGCCTAAGGTA |
| PS_AMP8-F | GCTGGAGTAGCTTGGTTAATG | PS_AMP8-R | TTATGAAGAGAGCGTCTGTACA |
| PS_AMP9-F | ATAAGGGACTGCCTATTGCC | PS_AMP9-R | CATATTCTGACTGCATACCACC |
| PS_AMP10-F | AGTTACCTTACTTGGGTTAACC | PS_AMP10-R | CGATACTGTTGGCAACGGTA |
| PS_AMP11-F | GAGCGTTTTAACACGCCATAC | AMP11-R | TACGGCTGCTACTTTCAGTTAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bang, W.Y.; Hur, J.; Kim, S.W. Discovery of Novel Genes Encoding Antimicrobial Peptides from the Pedobacter silvilitoris Genome with Broad-Spectrum Antimicrobial Activity. Int. J. Mol. Sci. 2025, 26, 6176. https://doi.org/10.3390/ijms26136176
Bang WY, Hur J, Kim SW. Discovery of Novel Genes Encoding Antimicrobial Peptides from the Pedobacter silvilitoris Genome with Broad-Spectrum Antimicrobial Activity. International Journal of Molecular Sciences. 2025; 26(13):6176. https://doi.org/10.3390/ijms26136176
Chicago/Turabian StyleBang, Woo Young, Jin Hur, and Sam Woong Kim. 2025. "Discovery of Novel Genes Encoding Antimicrobial Peptides from the Pedobacter silvilitoris Genome with Broad-Spectrum Antimicrobial Activity" International Journal of Molecular Sciences 26, no. 13: 6176. https://doi.org/10.3390/ijms26136176
APA StyleBang, W. Y., Hur, J., & Kim, S. W. (2025). Discovery of Novel Genes Encoding Antimicrobial Peptides from the Pedobacter silvilitoris Genome with Broad-Spectrum Antimicrobial Activity. International Journal of Molecular Sciences, 26(13), 6176. https://doi.org/10.3390/ijms26136176