
A Tourette Syndrome/ADHD-like Phenotype Results from Postnatal Disruption of CB1 and CB2 Receptor Signalling

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
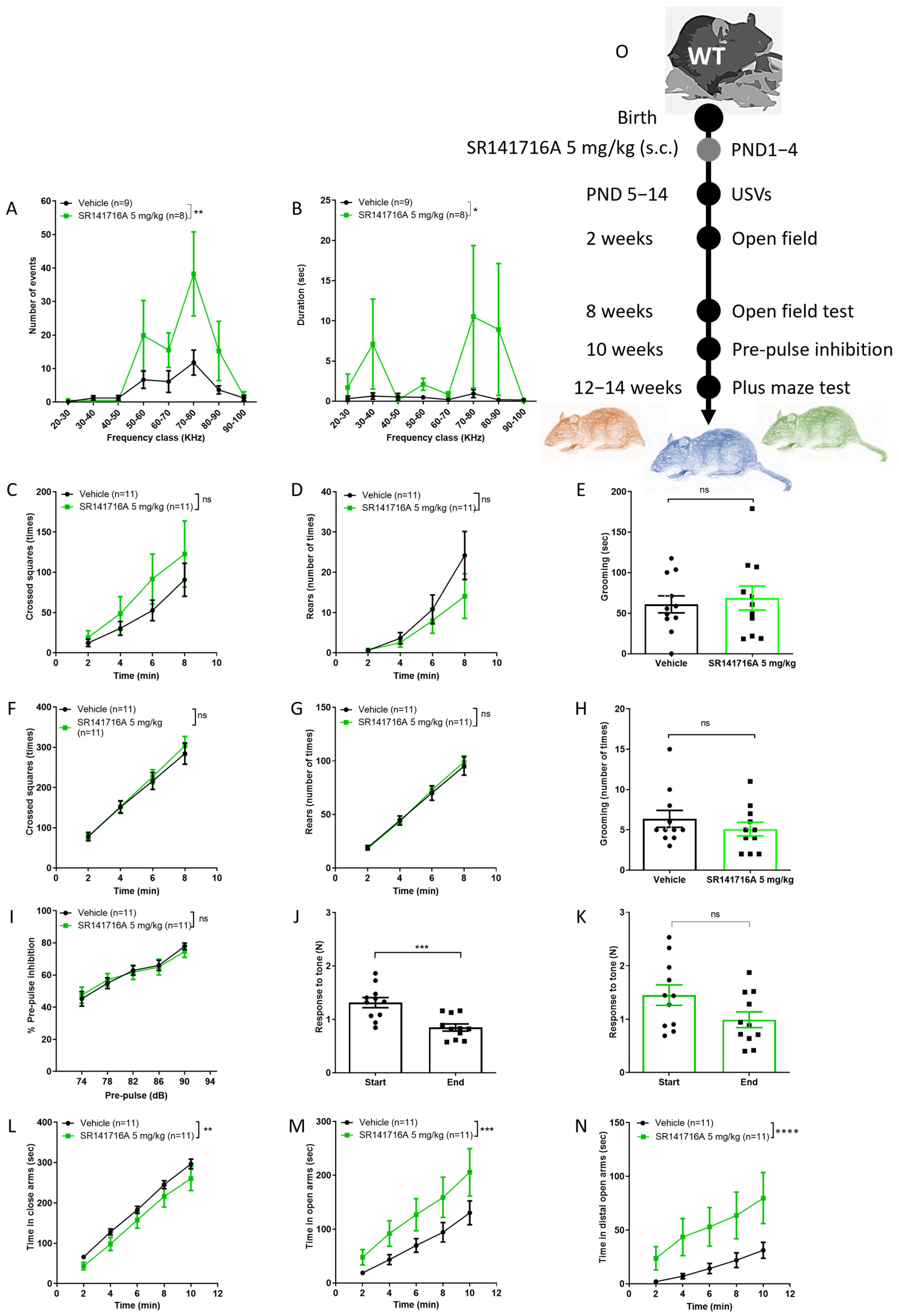
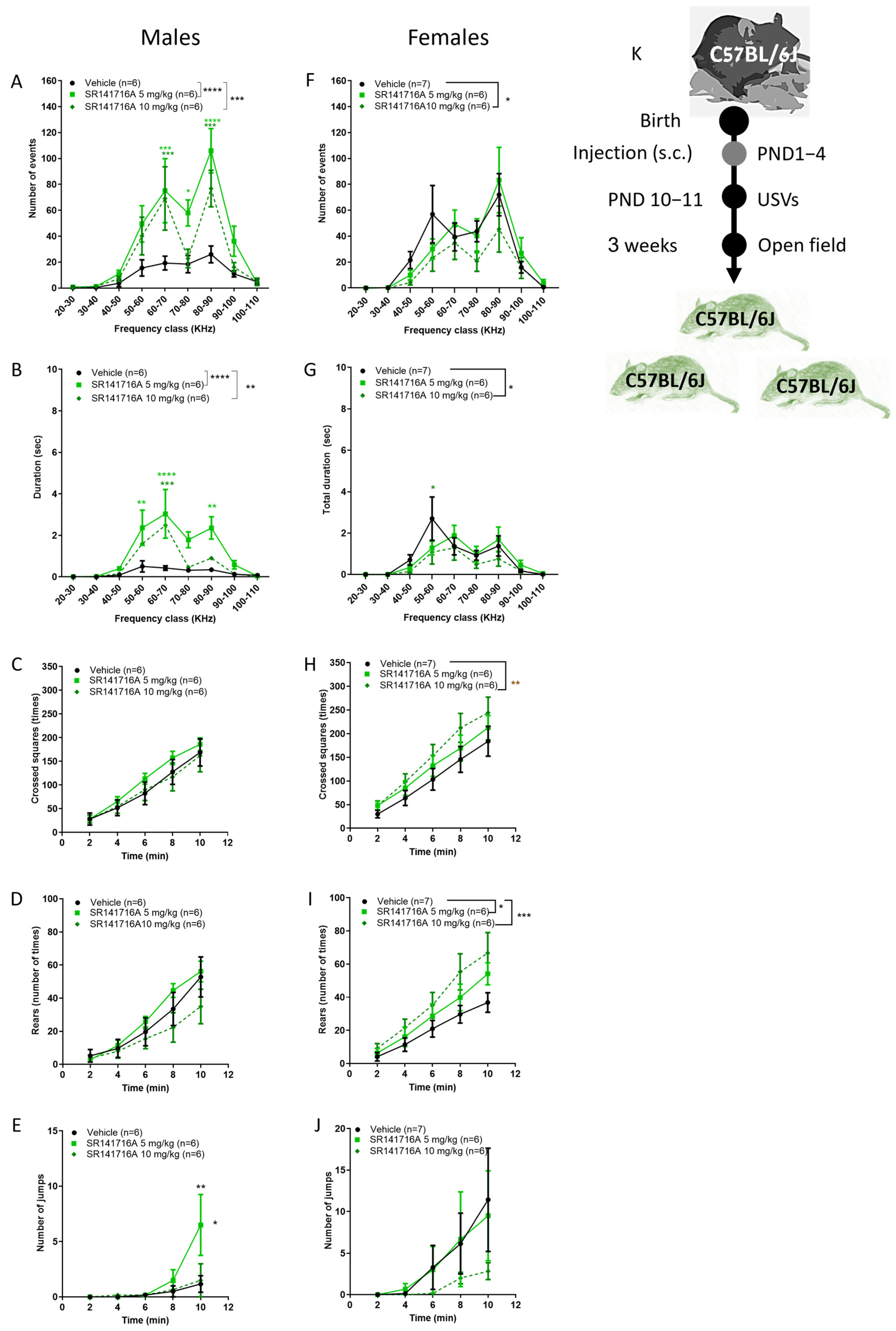
2.1. Effects of Postnatal Exposure to 5 mg/kg SR141716A on Male Mice
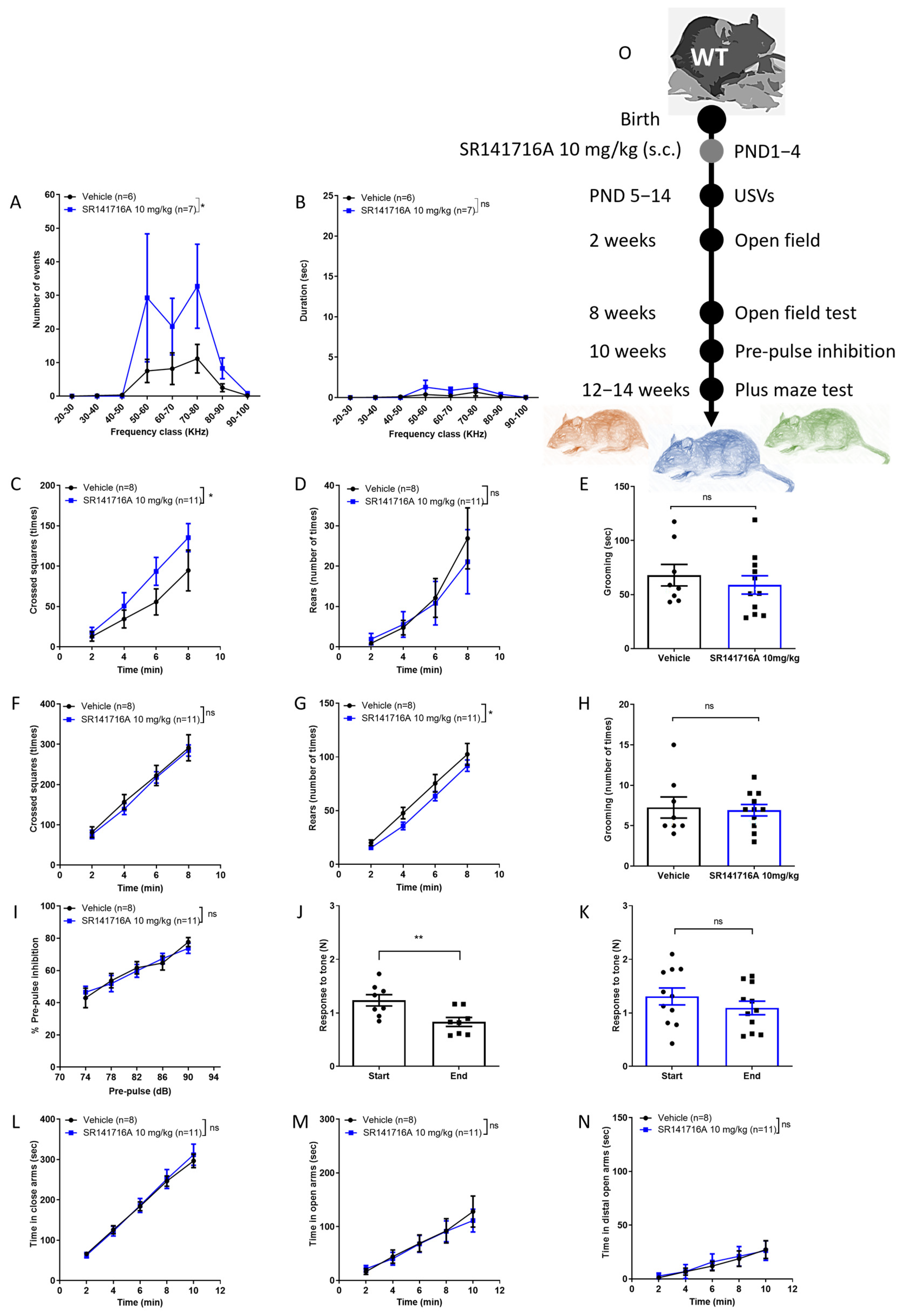
2.2. Effects of Postnatal Exposure to 10 mg/kg SR141716A on Male Mice
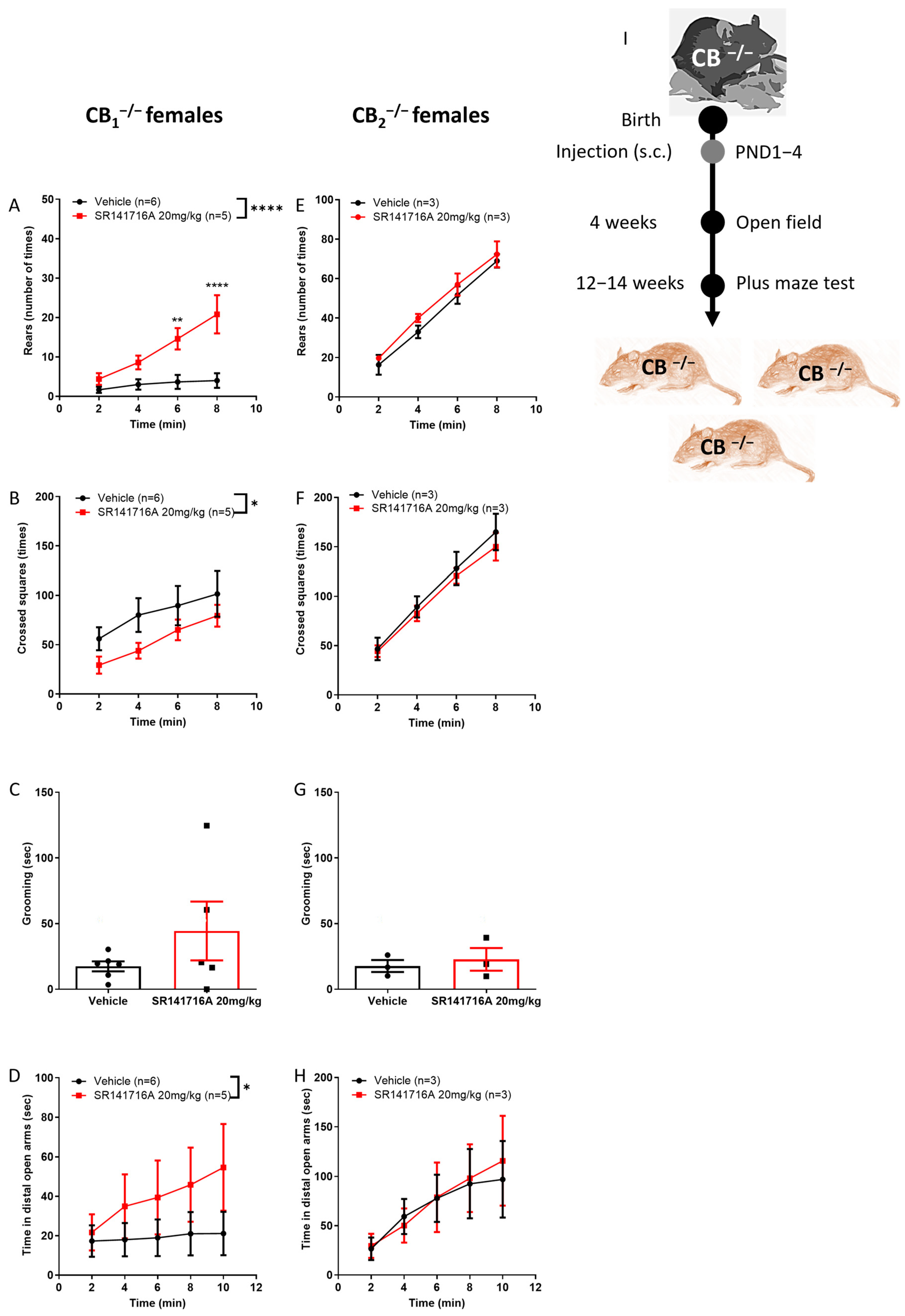
2.3. Effects of Postnatal Exposure to 20 mg/kg SR141716A on Male Mice
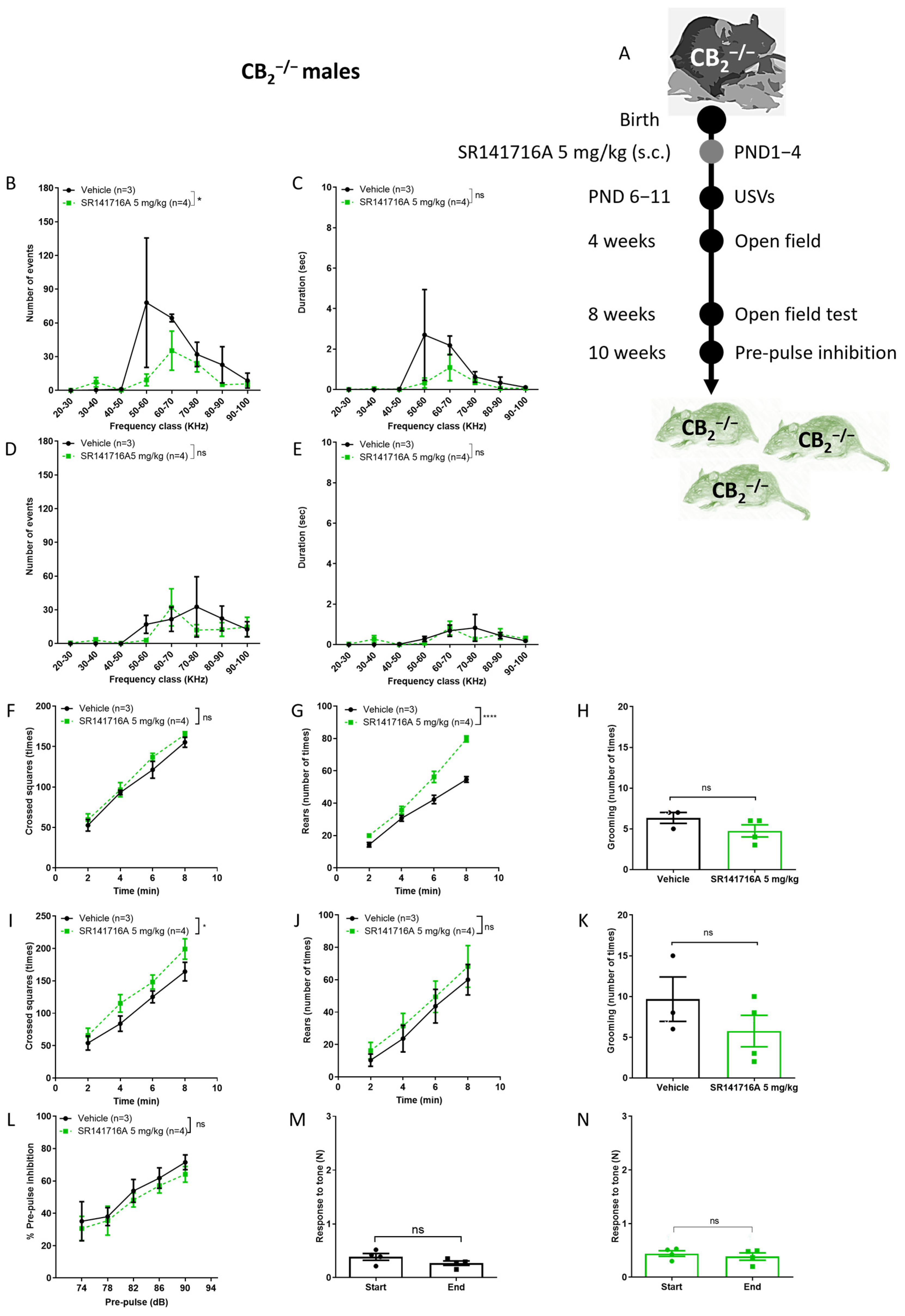
2.4. Effects of Postnatal Exposure to SR141716A on CB2 Receptor Knockout Male Mice
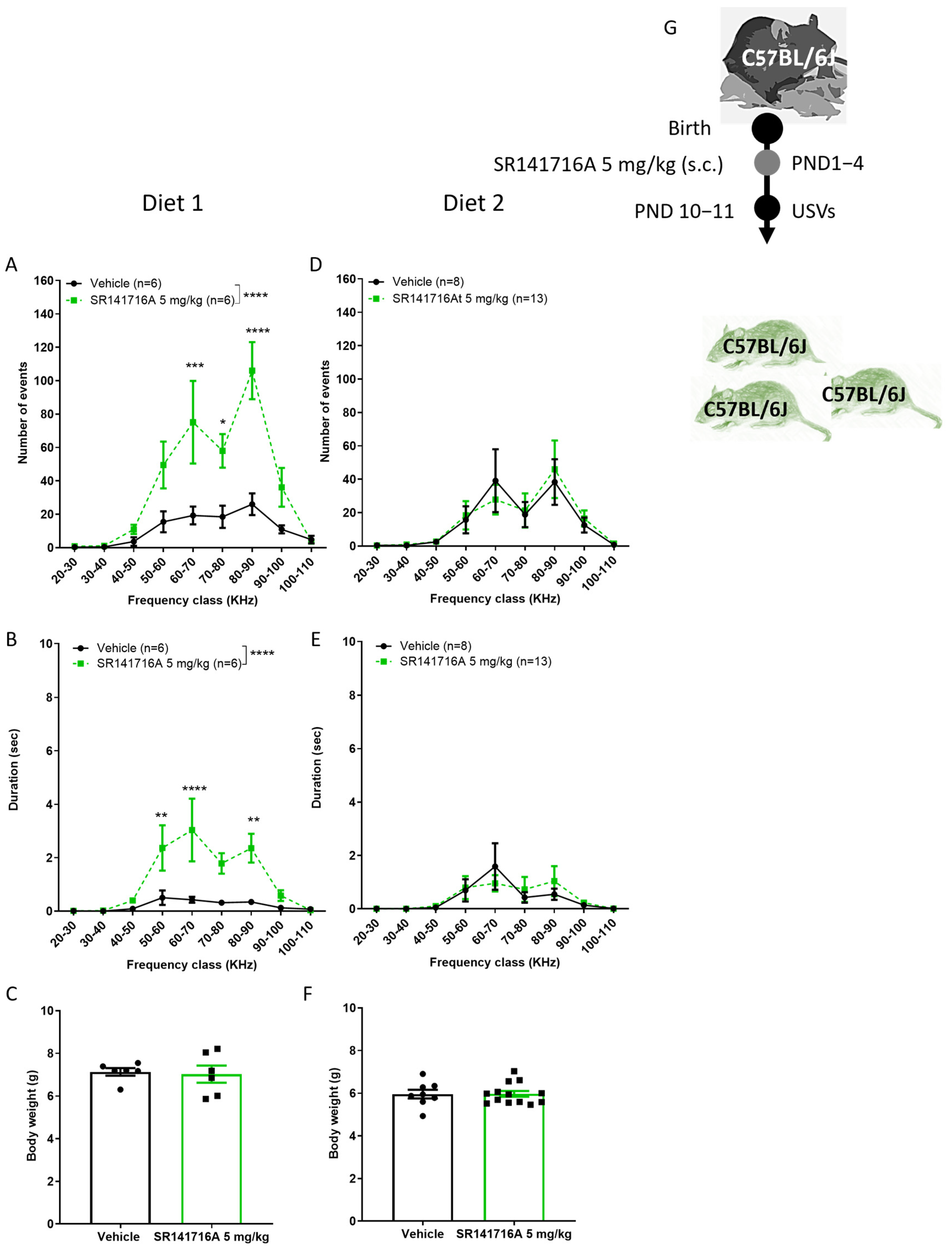
2.5. Effects of Diet on SR141716A-Induced Vocal-like Tics in Males
2.6. Sex Differences in the Effect of Postnatal SR141716A on Juveniles
2.7. Contribution of Cannabinoid Receptors to the Effect of SR141716A on Females
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Procedures
4.3. Drugs and Materials
4.4. USV Recording
4.5. Open Field Test
4.6. Pre-Pulse Inhibition (PPI) Test of the Startle Reflex
4.7. Elevated Plus-Maze (EPM) Test
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| OCD | Obsessive compulsive disorder |
| ESR | Ear scratch response |
| HTR | Head twitch response |
| ∆9-THC | ∆9-tetrahydrocannabinol |
| TS | Tourette syndrome |
| ADHD | Attention-deficit hyperactivity disorder |
| PPI | Pre-pulse inhibition |
| USV | Ultrasonic vocalization |
| CB1 | Cannabinoid receptor 1 |
| CB2 | Cannabinoid receptor 2 |
References
- Augustine, F.; Singer, H.S. Merging the Pathophysiology and Pharmacotherapy of Tics. Tremor Other Hyperkinet Mov. 2018, 8, 595. [Google Scholar] [CrossRef]
- Bloch, M.H.; Panza, K.E.; Landeros-Weisenberger, A.; Leckman, J.F. Meta-analysis: Treatment of attention-deficit/hyperactivity disorder in children with comorbid tic disorders. J. Am. Acad. Child. Adolesc. Psychiatry 2009, 48, 884–893. [Google Scholar] [CrossRef] [PubMed]
- McNaught, K.S.P.; Mink, J.W. Advances in understanding and treatment of Tourette syndrome. Nat. Rev. Neurol. 2011, 7, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Halmoy, A.; Klungsoyr, K.; Skjaerven, R.; Haavik, J. Pre- and perinatal risk factors in adults with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2012, 71, 474–481. [Google Scholar] [CrossRef]
- Fride, E. The endocannabinoid-CB(1) receptor system in pre- and postnatal life. Eur. J. Pharmacol. 2004, 500, 289–297. [Google Scholar] [CrossRef]
- Thuy, N.; Brian, F.T.; Yanan, Z. Overcoming the Psychiatric Side Effects of the Cannabinoid CB1 Receptor Antagonists: Current Approaches for Therapeutics Development. Curr. Top. Med. Chem. 2019, 19, 1418–1435. [Google Scholar] [CrossRef]
- Fride, E.; Braun, H.; Matan, H.; Steinberg, S.; Reggio, P.H.; Seltzman, H.H. Inhibition of milk ingestion and growth after administration of a neutral cannabinoid CB1 receptor antagonist on the first postnatal day in the mouse. Pediatr. Res. 2007, 62, 533–536. [Google Scholar] [CrossRef]
- Fride, E.; Foox, A.; Rosenberg, E.; Faigenboim, M.; Cohen, V.; Barda, L.; Blau, H.; Mechoulam, R. Milk intake and survival in newborn cannabinoid CB1 receptor knockout mice: Evidence for a “CB3” receptor. Eur. J. Pharmacol. 2003, 461, 27–34. [Google Scholar] [CrossRef]
- Freestone, P.S.; Guatteo, E.; Piscitelli, F.; di Marzo, V.; Lipski, J.; Mercuri, N.B. Glutamate spillover drives endocannabinoid production and inhibits GABAergic transmission in the Substantia Nigra pars compacta. Neuropharmacology 2014, 79, 467–475. [Google Scholar] [CrossRef]
- Rubino, T.; Vigano, D.; Zagato, E.; Sala, M.; Parolaro, D. In vivo characterization of the specific cannabinoid receptor antagonist, SR141716A: Behavioral and cellular responses after acute and chronic treatments. Synapse 2000, 35, 8–14. [Google Scholar] [CrossRef]
- Galve-Roperh, I.; Chiurchiu, V.; Diaz-Alonso, J.; Bari, M.; Guzman, M.; Maccarrone, M. Cannabinoid receptor signaling in progenitor/stem cell proliferation and differentiation. Prog. Lipid Res. 2013, 52, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Mátyás, F.; Urbán, G.M.; Watanabe, M.; Mackie, K.; Zimmer, A.; Freund, T.F.; Katona, I. Identification of the sites of 2-arachidonoylglycerol synthesis and action imply retrograde endocannabinoid signaling at both GABAergic and glutamatergic synapses in the ventral tegmental area. Neuropharmacology 2008, 54, 95–107. [Google Scholar] [CrossRef]
- Molina-Holgado, F.; Rubio-Araiz, A.; García-Ovejero, D.; Williams, R.J.; Moore, J.D.; Arévalo-Martín, A.; Gómez-Torres, O.; Molina-Holgado, E. CB2 cannabinoid receptors promote mouse neural stem cell proliferation. Eur. J. Neurosci. 2007, 25, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Mulder, J.; Aguado, T.; Keimpema, E.; Barabás, K.; Ballester Rosado, C.J.; Nguyen, L.; Monory, K.; Marsicano, G.; Di Marzo, V.; Hurd, Y.L.; et al. Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning. Proc. Natl. Acad. Sci. USA 2008, 105, 8760–8765. [Google Scholar] [CrossRef] [PubMed]
- Palazuelos, J.; Aguado, T.; Egia, A.; Mechoulam, R.; Guzmán, M.; Galve-Roperh, I. Non-psychoactive CB2 cannabinoid agonists stimulate neural progenitor proliferation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 2405–2407. [Google Scholar] [CrossRef]
- Jarbe, T.U.; Ross, T.; DiPatrizio, N.V.; Pandarinathan, L.; Makriyannis, A. Effects of the CB1R agonist WIN-55,212-2 and the CB1R antagonists SR-141716 and AM-1387: Open-field examination in rats. Pharmacol. Biochem. Behav. 2006, 85, 243–252. [Google Scholar] [CrossRef]
- Ettaro, R.; Laudermilk, L.; Clark, S.D.; Maitra, R. Behavioral assessment of rimonabant under acute and chronic conditions. Behav. Brain Res. 2020, 390, 112697. [Google Scholar] [CrossRef]
- Bosier, B.; Sarre, S.; Smolders, I.; Michotte, Y.; Hermans, E.; Lambert, D.M. Revisiting the complex influences of cannabinoids on motor functions unravels pharmacodynamic differences between cannabinoid agonists. Neuropharmacology 2010, 59, 503–510. [Google Scholar] [CrossRef]
- Gamble-George, J.C.; Conger, J.R.; Hartley, N.D.; Gupta, P.; Sumislawski, J.J.; Patel, S. Dissociable effects of CB1 receptor blockade on anxiety-like and consummatory behaviors in the novelty-induced hypophagia test in mice. Psychopharmacology 2013, 228, 401–409. [Google Scholar] [CrossRef]
- Gorberg, V.; Borisov, V.; Greig, I.R.; Pertwee, R.G.; McCaffery, P.; Anavi-Goffer, S. Motor-like Tics are Mediated by CB2 Cannabinoid Receptor-dependent and Independent Mechanisms Associated with Age and Sex. Mol. Neurobiol. 2022, 59, 5070–5083. [Google Scholar] [CrossRef]
- Rabichev, S.; Anavi-Goffer, S.; Ross, R.A.; Fride, E. Postnatal Inhibition of the Endocannabinoid System Is Associated with ADHD-like Symptoms in Adulthood. In Proceedings of the 20th Annual Symposium of the International Cannabinoid Research Society, Lund, Sweden, 23–27 July 2010; p. 26. [Google Scholar]
- Health and Care of People with Learning Disabilities, Experimental Statistics 2023 to 2024; NHS England Digital: Leeds, UK, 2023.
- Reuben, C. Attention-Deficit/Hyperactivity Disorder in Children Ages 5–17 Years: United States, 2020–2022. NCHS Data Brief 2024, 499. [Google Scholar] [CrossRef]
- Feifel, D.; Minassian, A.; Perry, W. Prepulse inhibition of startle in adults with ADHD. J. Psychiatr. Res. 2009, 43, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Singer, P.; Hauser, J.; Llano Lopez, L.H.; Peleg-Raibstein, D.; Feldon, J.; Gargiulo, P.A.; Yee, B.K. Prepulse inhibition predicts working memory performance whilst startle habituation predicts spatial reference memory retention in C57BL/6 mice. Behav. Brain Res. 2013, 242, 166–177. [Google Scholar] [CrossRef]
- Geyer, M.A. Are cross-species measures of sensorimotor gating useful for the discovery of procognitive cotreatments for schizophrenia? Dialogues Clin. Neurosci. 2006, 8, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Alvaro, A.; Navarrete, F.; Aracil-Fernandez, A.; Navarro, D.; Berbel, P.; Manzanares, J. Differential Pharmacological Regulation of Sensorimotor Gating Deficit in CB1 Knockout Mice and Associated Neurochemical and Histological Alterations. Neuropsychopharmacology 2015, 40, 2639–2647. [Google Scholar] [CrossRef]
- Isaacs, D.; Riordan, H. Sensory hypersensitivity in Tourette syndrome: A review. Brain Dev. 2020, 42, 627–638. [Google Scholar] [CrossRef]
- Malone, D.T.; Taylor, D.A. The effect of Delta9-tetrahydrocannabinol on sensorimotor gating in socially isolated rats. Behav. Brain Res. 2006, 166, 101–109. [Google Scholar] [CrossRef]
- Malone, D.T.; Long, L.E.; Taylor, D.A. The effect of SR 141716 and apomorphine on sensorimotor gating in Swiss mice. Pharmacol. Biochem. Behav. 2004, 77, 839–845. [Google Scholar] [CrossRef]
- Marongiu, M.F.; Poddie, D.; Porcu, S.; Manchinu, M.F.; Castelli, M.P.; Sogos, V.; Bini, V.; Frau, R.; Caredda, E.; Collu, M.; et al. Reversible disruption of pre-pulse inhibition in hypomorphic-inducible and reversible CB1−/− mice. PLoS ONE 2012, 7, e35013. [Google Scholar] [CrossRef]
- Gorberg, V.; McCaffery, P.; Anavi-Goffer, S. Different responses of repetitive behaviours in juvenile and young adult mice to Delta(9)-tetrahydrocannabinol and cannabidiol may affect decision making for Tourette syndrome. Br. J. Pharmacol. 2021, 178, 614–625. [Google Scholar] [CrossRef]
- McGregor, I.S.; Dastur, F.N.; McLellan, R.A.; Brown, R.E. Cannabinoid modulation of rat pup ultrasonic vocalizations. Eur. J. Pharmacol. 1996, 313, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Darmani, N.A.; Pandya, D.K. Involvement of other neurotransmitters in behaviors induced by the cannabinoid CB1 receptor antagonist SR 141716A in naive mice. J. Neural Transm. 2000, 107, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Patrini, G.; Massi, P.; Fuzio, D.; Vigano, D.; Giagnoni, G.; Parolaro, D. Cannabinoid-precipitated withdrawal: A time-course study of the behavioral aspect and its correlation with cannabinoid receptors and G protein expression. J. Pharmacol. Exp. Ther. 1998, 285, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Hofer, M.A.; Shair, H.N.; Brunelli, S.A. Ultrasonic vocalizations in rat and mouse pups. Curr. Protoc. Neurosci. 2002, 17, 8–14. [Google Scholar] [CrossRef]
- Buonsenso, D.; De Rose, C.; Mariotti, P. Children experienced new or worsening tic issues when they were separated from their parents during the Italian COVID-19 lockdown. Acta Paediatr. 2021, 110, 394–396. [Google Scholar] [CrossRef]
- Rizzo, F.; Nespoli, E.; Abaei, A.; Bar-Gad, I.; Deelchand, D.K.; Fegert, J.; Rasche, V.; Hengerer, B.; Boeckers, T.M. Aripiprazole Selectively Reduces Motor Tics in a Young Animal Model for Tourette’s Syndrome and Comorbid Attention Deficit and Hyperactivity Disorder. Front. Neurol. 2018, 9, 59. [Google Scholar] [CrossRef]
- Anavi-Goffer, S.; Baillie, G.; Irving, A.J.; Gertsch, J.; Greig, I.R.; Pertwee, R.G.; Ross, R.A. Modulation of L-alpha-lysophosphatidylinositol/GPR55 mitogen-activated protein kinase (MAPK) signaling by cannabinoids. J. Biol. Chem. 2012, 287, 91–104. [Google Scholar] [CrossRef]
- Anavi-Goffer, S.; Mulder, J. The polarised life of the endocannabinoid system in CNS development. Chembiochem Eur. J. Chem. Biol. 2009, 10, 1591–1598. [Google Scholar] [CrossRef]
- Xi, Z.X.; Peng, X.Q.; Li, X.; Song, R.; Zhang, H.Y.; Liu, Q.R.; Yang, H.J.; Bi, G.H.; Li, J.; Gardner, E.L. Brain cannabinoid CB(2) receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1166. [Google Scholar] [CrossRef]
- Ortega-Alvaro, A.; Aracil-Fernandez, A.; Garcia-Gutierrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 2011, 36, 1489–1504. [Google Scholar] [CrossRef]
- Schwarzenberg, S.J.; Georgieff, M.K.; Committee On, N. Advocacy for Improving Nutrition in the First 1000 Days to Support Childhood Development and Adult Health. Pediatrics 2018, 141, e20173716. [Google Scholar] [CrossRef] [PubMed]
- Wachs, T.D. Models linking nutritional deficiencies to maternal and child mental health. Am. J. Clin. Nutr. 2009, 89, 935S–939S. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, A.K.; Rogers, S.L. Understanding the impact of diet and nutrition on symptoms of Tourette syndrome: A scoping review. J. Child Health Care Prof. Work. Child Hosp. Community 2018, 22, 68–83. [Google Scholar] [CrossRef]
- Biagioni, A.F.; dos Anjos-Garcia, T.; Ullah, F.; Fisher, I.R.; Falconi-Sobrinho, L.L.; de Freitas, R.L.; Felippotti, T.T.; Coimbra, N.C. Neuroethological validation of an experimental apparatus to evaluate oriented and non-oriented escape behaviours: Comparison between the polygonal arena with a burrow and the circular enclosure of an open-field test. Behav. Brain Res. 2016, 298, 65–77. [Google Scholar] [CrossRef]
- Robinson, S.; Hedderly, T. Novel Psychological Formulation and Treatment of “Tic Attacks” in Tourette Syndrome. Front. Pediatr. 2016, 4, 46. [Google Scholar] [CrossRef]
- Lees, A.J.; Robertson, M.; Trimble, M.R.; Murray, N.M. A clinical study of Gilles de la Tourette syndrome in the United Kingdom. J. Neurol. Neurosurg. Psychiatry 1984, 47, 1–8. [Google Scholar] [CrossRef]
- Davis, M.I.; Crittenden, J.R.; Feng, A.Y.; Kupferschmidt, D.A.; Naydenov, A.; Stella, N.; Graybiel, A.M.; Lovinger, D.M. The cannabinoid-1 receptor is abundantly expressed in striatal striosomes and striosome-dendron bouquets of the substantia nigra. PLoS ONE 2018, 13, e0191436. [Google Scholar] [CrossRef]
- Crittenden, J.R.; Yoshida, T.; Venu, S.; Mahar, A.; Graybiel, A.M. Cannabinoid Receptor 1 Is Required for Neurodevelopment of Striosome-Dendron Bouquets. eNeuro 2022, 9, ENEURO.0318-21.2022. [Google Scholar] [CrossRef]
- Sallee, F.; Kohegyi, E.; Zhao, J.; McQuade, R.; Cox, K.; Sanchez, R.; Van Beek, A.; Nyilas, M.; Carson, W.; Kurlan, R. Randomized, Double-Blind, Placebo-Controlled Trial Demonstrates the Efficacy and Safety of Oral Aripiprazole for the Treatment of Tourette’s Disorder in Children and Adolescents. J. Child Adolesc. Psychopharmacol. 2017, 27, 771–781. [Google Scholar] [CrossRef]
- Israelashvili, M.; Yael, D.; Vinner, E.; Belelovsky, K.; Bar-Gad, I. Common neuronal mechanisms underlying tics and hyperactivity. Cortex 2020, 127, 231–247. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, H.; Lee, P.H.; Tsetsos, F.; Davis, L.K.; Yu, D.; Lee, S.H.; Dalsgaard, S.; Haavik, J.; Barta, C.; et al. Investigating Shared Genetic Basis Across Tourette Syndrome and Comorbid Neurodevelopmental Disorders Along the Impulsivity-Compulsivity Spectrum. Biol. Psychiatry 2021, 90, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Müller-Vahl, K.R. Cannabinoids in the Treatment of Selected Mental Illnesses: Practical Approach and Overview of the Literature. Pharmacopsychiatry 2024, 57, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Darmani, N.A.; Janoyan, J.J.; Kumar, N.; Crim, J.L. Behaviorally active doses of the CB1 receptor antagonist SR 141716A increase brain serotonin and dopamine levels and turnover. Pharmacol. Biochem. Behav. 2003, 75, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Canseco-Alba, A.; Sanabria, B.; Hammouda, M.; Bernadin, R.; Mina, M.; Liu, Q.R.; Onaivi, E.S. Cell-Type Specific Deletion of CB2 Cannabinoid Receptors in Dopamine Neurons Induced Hyperactivity Phenotype: Possible Relevance to Attention-Deficit Hyperactivity Disorder. Front. Psychiatry 2021, 12, 803394. [Google Scholar] [CrossRef]
- Bortolato, M.; Pittenger, C. Modeling tics in rodents: Conceptual challenges and paths forward. J. Neurosci. Methods 2017, 292, 12–19. [Google Scholar] [CrossRef]
- Jordan, C.J.; Xi, Z.X. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019, 98, 208–220. [Google Scholar] [CrossRef]
- Gorman, D.A.; Zhu, H.; Anderson, G.M.; Davies, M.; Peterson, B.S. Ferritin levels and their association with regional brain volumes in Tourette’s syndrome. Am. J. Psychiatry 2006, 163, 1264–1272. [Google Scholar] [CrossRef]
- Nosratmirshekarlou, E.; Shafiq, S.; Goodarzi, Z.S.; Martino, D.; Pringsheim, T. Effect of diet, exercise and sleep on tic severity: A scoping review protocol. BMJ Open 2019, 9, e024653. [Google Scholar] [CrossRef]
- Smith, B.L.; Ludlow, A.K. Patterns of Nutritional Supplement Use in Children with Tourette Syndrome. J. Diet. Suppl. 2023, 20, 28–43. [Google Scholar] [CrossRef]
- Lee, J.; Lee, A.; Kim, J.-H.; Shin, Y.M.; Kim, S.-J.; Cho, W.D.; Lee, S.I. Effect of Omega-3 and Korean Red Ginseng on Children with Attention Deficit Hyperactivity Disorder: An Open-Label Pilot Study. Clin. Psychopharmacol. Neurosci. 2020, 18, 75–80. [Google Scholar] [CrossRef]
- Ghanbari, M.-M.; Loron, A.G.; Sayyah, M. The ω-3 endocannabinoid docosahexaenoyl ethanolamide reduces seizure susceptibility in mice by activating cannabinoid type 1 receptors. Brain Res. Bull. 2021, 170, 74–80. [Google Scholar] [CrossRef]
- Rizzo, R.; Prato, A.; Scerbo, M.; Saia, F.; Barone, R.; Curatolo, P. Use of Nutritional Supplements Based on L-Theanine and Vitamin B6 in Children with Tourette Syndrome, with Anxiety Disorders: A Pilot Study. Nutrients 2022, 14, 852. [Google Scholar] [CrossRef]
- Mantel, B.J.; Meyers, A.; Tran, Q.Y.; Rogers, S.; Jacobson, J.S. Nutritional supplements and complementary/alternative medicine in Tourette syndrome. J. Child. Adolesc. Psychopharmacol. 2004, 14, 582–589. [Google Scholar] [CrossRef]
- Das, J.K.; Salam, R.A.; Hadi, Y.B.; Sadiq Sheikh, S.; Bhutta, A.Z.; Weise Prinzo, Z.; Bhutta, Z.A. Preventive lipid-based nutrient supplements given with complementary foods to infants and young children 6 to 23 months of age for health, nutrition, and developmental outcomes. Cochrane Database Syst. Rev. 2019, 5, Cd012611. [Google Scholar] [CrossRef]
- Zimmer, A.; Zimmer, A.M.; Hohmann, A.G.; Herkenham, M.; Bonner, T.I. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc. Natl. Acad. Sci. USA 1999, 96, 5780–5785. [Google Scholar] [CrossRef]
- Hanus, L.; Breuer, A.; Tchilibon, S.; Shiloah, S.; Goldenberg, D.; Horowitz, M.; Pertwee, R.G.; Ross, R.A.; Mechoulam, R.; Fride, E. HU-308: A specific agonist for CB(2), a peripheral cannabinoid receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 14228–14233. [Google Scholar] [CrossRef]
- Skodzik, T.; Holling, H.; Pedersen, A. Long-Term Memory Performance in Adult ADHD. J. Atten. Disord. 2017, 21, 267–283. [Google Scholar] [CrossRef]
- Lafenetre, P.; Chaouloff, F.; Marsicano, G. Bidirectional regulation of novelty-induced behavioral inhibition by the endocannabinoid system. Neuropharmacology 2009, 57, 715–721. [Google Scholar] [CrossRef]
- Bagheri, F.; Goudarzi, I.; Lashkarbolouki, T.; Elahdadi Salmani, M.; Goudarzi, A.; Morley-Fletcher, S. The Combined Effects of Perinatal Ethanol and Early-Life Stress on Cognition and Risk-Taking Behavior through Oxidative Stress in Rats. Neurotox. Res. 2022, 40, 925–940. [Google Scholar] [CrossRef]
- Cao, Y.; Sun, C.; Huang, J.; Sun, P.; Wang, L.; He, S.; Liao, J.; Lu, Z.; Lu, Y.; Zhong, C. Dysfunction of the Hippocampal-Lateral Septal Circuit Impairs Risk Assessment in Epileptic Mice. Front. Mol. Neurosci. 2022, 15, 828891. [Google Scholar] [CrossRef]
- Viola, T.W.; Wearick-Silva, L.E.; Creutzberg, K.C.; Kestering-Ferreira, É.; Orso, R.; Centeno-Silva, A.; Albrechet-Souza, L.; Marshall, P.R.; Li, X.; Bredy, T.W.; et al. Postnatal impoverished housing impairs adolescent risk-assessment and increases risk-taking: A sex-specific effect associated with histone epigenetic regulation of Crfr1 in the medial prefrontal cortex. Psychoneuroendocrinology 2019, 99, 8–19. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorberg, V.; Harpaz, T.; Shamir, E.N.; Karminsky, O.D.; Fride, E.; Pertwee, R.G.; Greig, I.R.; McCaffery, P.; Anavi-Goffer, S. A Tourette Syndrome/ADHD-like Phenotype Results from Postnatal Disruption of CB1 and CB2 Receptor Signalling. Int. J. Mol. Sci. 2025, 26, 6052. https://doi.org/10.3390/ijms26136052
Gorberg V, Harpaz T, Shamir EN, Karminsky OD, Fride E, Pertwee RG, Greig IR, McCaffery P, Anavi-Goffer S. A Tourette Syndrome/ADHD-like Phenotype Results from Postnatal Disruption of CB1 and CB2 Receptor Signalling. International Journal of Molecular Sciences. 2025; 26(13):6052. https://doi.org/10.3390/ijms26136052
Chicago/Turabian StyleGorberg, Victoria, Tamar Harpaz, Emilya Natali Shamir, Orit Diana Karminsky, Ester Fride, Roger G. Pertwee, Iain R. Greig, Peter McCaffery, and Sharon Anavi-Goffer. 2025. "A Tourette Syndrome/ADHD-like Phenotype Results from Postnatal Disruption of CB1 and CB2 Receptor Signalling" International Journal of Molecular Sciences 26, no. 13: 6052. https://doi.org/10.3390/ijms26136052
APA StyleGorberg, V., Harpaz, T., Shamir, E. N., Karminsky, O. D., Fride, E., Pertwee, R. G., Greig, I. R., McCaffery, P., & Anavi-Goffer, S. (2025). A Tourette Syndrome/ADHD-like Phenotype Results from Postnatal Disruption of CB1 and CB2 Receptor Signalling. International Journal of Molecular Sciences, 26(13), 6052. https://doi.org/10.3390/ijms26136052