Abstract
The SNCA gene, encoding alpha-synuclein, is implicated in the pathogenesis of Parkinson’s disease (PD), with several single-nucleotide polymorphisms (SNPs) linked to increased risk. This study systematically evaluated the association between common SNCA polymorphisms and PD through a meta-analysis of cohort and case–control studies published before 20 November 2023. Eligible studies were identified via comprehensive searches of PubMed, Scopus, and Web of Science, and pooled odds ratios with 95% confidence intervals were calculated under allelic, dominant, and recessive models. Heterogeneity and publication bias were assessed, and subgroup and sensitivity analyses were performed. Twenty-seven studies were included. SNP rs11931074 showed consistent associations with PD across all models, with low heterogeneity and no evidence of publication bias. rs356219 and rs356165 were also significantly associated with PD, although regional differences contributed to heterogeneity. In contrast, rs2583988 showed marginal significance in the allelic model, which was lost after sensitivity analyses. No associations were found under dominant or recessive models for this SNP. These findings confirm rs11931074 as a robust PD risk variant and support the roles of rs356219 and rs356165 while suggesting weaker evidence for rs2583988. Large, multi-ethnic studies are warranted to elucidate underlying mechanisms and support precision medicine in PD.
1. Introduction
Parkinson’s Disease (PD) is a prevalent and progressive neurodegenerative disorder affecting millions worldwide, characterized by motor symptoms (e.g., bradykinesia, tremor, rigidity, postural instability) and non-motor symptoms (e.g., cognitive impairment, mood disorders, autonomic dysfunction) [1]. These manifestations lead to substantial morbidity and reduced survival. PD multifactorial etiology involves environmental and genetic factors, with genetic susceptibility increasingly recognized as a modulator of both risk and progression [2,3,4,5].
A key pathological hallmark of PD and Lewy body dementia is the aggregation of the alpha-synuclein protein, which is encoded by the SNCA gene, ultimately leading to the formation of Lewy bodies. Rare SNCA mutations identified through linkage studies cause familial PD via amino acid substitutions and altered protein configurations [1,6,7]. Beyond these, common SNCA variants, including single-nucleotide polymorphisms (SNPs) and promoter region polymorphisms such as the dinucleotide repeat sequence (REP1), have been associated with sporadic PD in various populations [8]. Notably, longer REP1 alleles correlate with increased SNCA expression and higher PD risk, reinforcing SNCA’s pivotal role in PD pathogenesis [9,10].
Emerging evidence suggests that SNCA variants may not only influence PD risk but also act as disease modifiers, affecting onset age, progression, and specific clinical manifestations [11]. For example, in a Han Chinese cohort, the SNCA SNPs rs11931074, rs7684318, and rs356219 showed strong linkage disequilibrium. The GG genotype of rs11931074 was associated with reduced PD risk but a significantly increased risk of developing dementia in PD [12]. Similarly, SNCA rs6826785 was associated with cognitive decline—non-carriers had a higher risk for PD-mild cognitive impairment (PD-MCI)—highlighting its potential as a therapeutic target for cognitive symptoms [13]. Beyond single variant effects, gene–gene and gene–environment interactions likely modulate PD risk and phenotype [9,14,15,16]. However, while genome-wide association studies (GWAS) have identified over 40 PD-associated loci, including SNCA, these explain only a modest portion of heritability. A genome-wide complex trait analysis (GCTA) suggests common variants account for up to 27% of PD risk, leaving much of the genetic contribution unidentified [17].
Given the substantial evidence linking SNCA polymorphisms with PD risk and progression, a systematic review and meta-analysis are essential to synthesize existing findings. This study aims to assess the impact of SNCA variants on PD susceptibility by integrating primary research studies. By systematically reviewing the literature and conducting meta-analyses on key SNCA polymorphisms, we aim to clarify their role in PD pathophysiology, identify inconsistencies, and inform future research. Results may enhance genetic risk prediction and inform therapeutic strategies in PD.
2. Methods
2.1. Study Selection
A comprehensive literature search using the electronic databases of PubMed, Scopus, and Web of Science for studies published before 20 November 2023, was conducted. Search terms included the following: (“Parkinson Disease” OR “Parkinson’s Disease”) AND (“Association” OR “Relationship” OR “Correlation” OR “Risk”) AND (“α-Synuclein” OR “SNCA” OR “Alpha-Synuclein” OR “aSyn”) AND (“Polymorphisms” OR “Variants” OR “Mutations” OR “Variation”). Four investigators (R.M, M.S, SF.M, and M.Z.Y.C.L)) independently screened studies applying the following inclusion criteria: (1) cohort or case–control studies on SNCA variants and PD, (2) English language, (3) human subjects, and (4) sufficient frequency data. Disagreements were resolved by consensus.
2.2. Data Extraction
Two researchers (SF.M and C.L.S) independently extracted the following from each study: first author, year of publication, region, ethnicity, sample size, age of case and control, PD type, SNP type, comparison type, odds ratios (ORs), and confidence intervals (CIs). Disagreements were resolved by consensus. Authors were not contacted for additional information.
2.3. Quality Assessment
Two investigators (R.M and S.E.S) independently assessed study quality using the Newcastle–Ottawa Scale (NOS) [18], which evaluates selection (4 points), comparability (2 points), and exposure/outcome (3 points), with a maximum of 9. Studies scoring ≥ 7 were rated high quality. Discrepancies were resolved by consensus.
2.4. Statistical Analysis
Meta-analyses were performed to evaluate the association between each SNP variant and PD risk, with ORs and 95% CIs calculated as effect sizes. Unadjusted ORs were used for most studies, except for two in which genotype frequencies were unavailable; in these cases, adjusted ORs were utilized. Heterogeneity across studies was assessed using Cochran’s Q test and the I2 statistic, where I2 values of 25%, 50%, and 75% were considered to indicate low, moderate, and high heterogeneity, respectively. Fixed-effects models were applied when heterogeneity was low (I2 < 25%), while random-effects models were used for moderate to high heterogeneity. To explore potential sources of heterogeneity, subgroup meta-analyses based on geographic region were conducted. Sensitivity analysis was performed using the leave-one-out method to examine the influence of individual studies on the pooled effect size. Publication bias was assessed using the trim-and-fill method to identify potential missing studies and evaluate funnel plot asymmetry. All statistical analyses were conducted using R software 4.4.2. (R Core Team (2024); R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org). A p-value < 0.05 was considered statistically significant.
3. Results
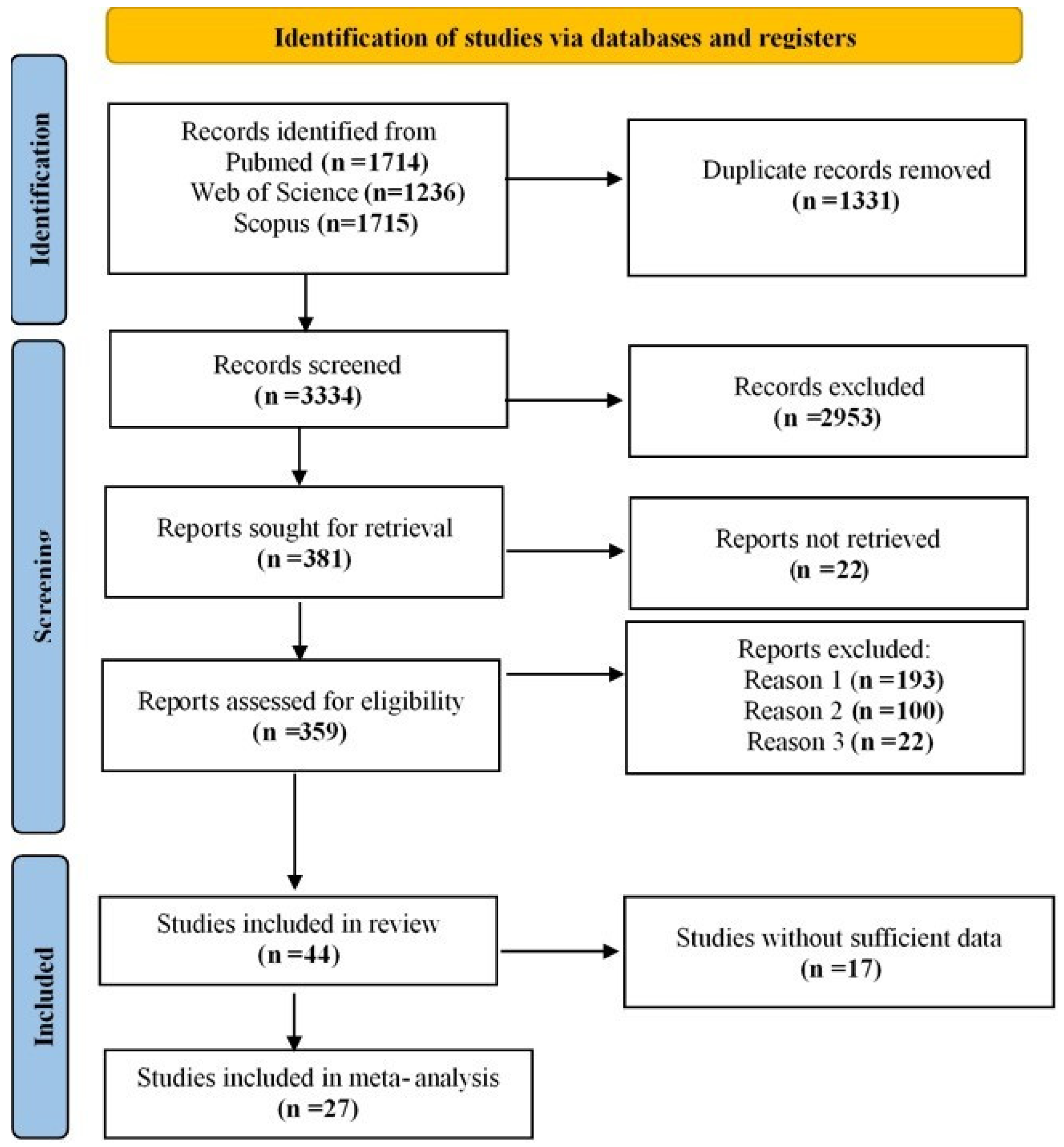
The systematic search identified 4665 studies, which were screened after removing duplicates, leaving 3334 studies for abstract screening. Based on the inclusion criteria, 381 studies were selected for full-text screening. However, 22 studies were unavailable, reducing the number to 359. Studies that lacked control groups, focused on Parkinson’s disease dementia (PDD) or early onset PD, or did not provide relevant data were excluded. Ultimately, 44 studies met the final inclusion criteria. Among them, 17 studies did not provide sufficient data on frequencies, while 27 contained sufficient data for meta-analysis. The included studies reported data on four SNCA polymorphisms: rs356165 (7 studies [2,19,20,21,22,23,24]; 8 allelic models: G vs. A, 5 dominant: AG + GG vs. AA, and 5 recessive: GG vs. AG + AA), rs2583988 (6 studies [9,19,20,25,26,27]; 7 allelic models: T vs. C, 3 dominant: TC + TT vs. CC, and 3 recessive: TT vs. TC + CC), rs356219 (16 studies [9,16,20,21,24,25,26,28,29,30,31,32,33,34,35,36]; 16 allelic models: G vs. A, 14 dominant: AG + GG vs. AA, and 14 recessive: GG vs. AG + AA), and rs11931074 (13 studies [2,5,19,21,23,24,25,26,37,38,39,40,41]; 14 allelic models: T vs. G, 9 dominant: TT + TG vs. GG, and 8 recessive: TT vs. TG + GG). All included studies were case–control in design, except for one study [28], which was longitudinal but used SNCA genotyping at baseline, making it comparable for meta-analysis. The study selection process is illustrated in Figure 1. The detailed characteristics and methodological quality of the included studies are summarized in the Supplementary Materials, Table S1. Seventeen studies scored 7 or above according to the NOS, indicating high quality and low risk of bias. Ten studies received scores of 5 or 6, suggesting moderate quality (Supplementary Materials, Table S2). The meta-analysis results for allele and genotype associations of the SNCA polymorphisms are presented in Table 1.
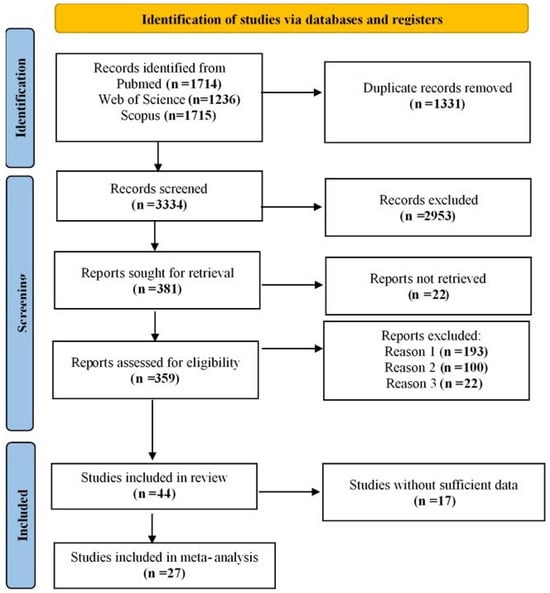
Figure 1.
PRISMA flow diagram illustrating the study selection process.

Table 1.
Summary of meta-analysis results for the association between SNPs and PD risk.
3.1. SNP rs356165
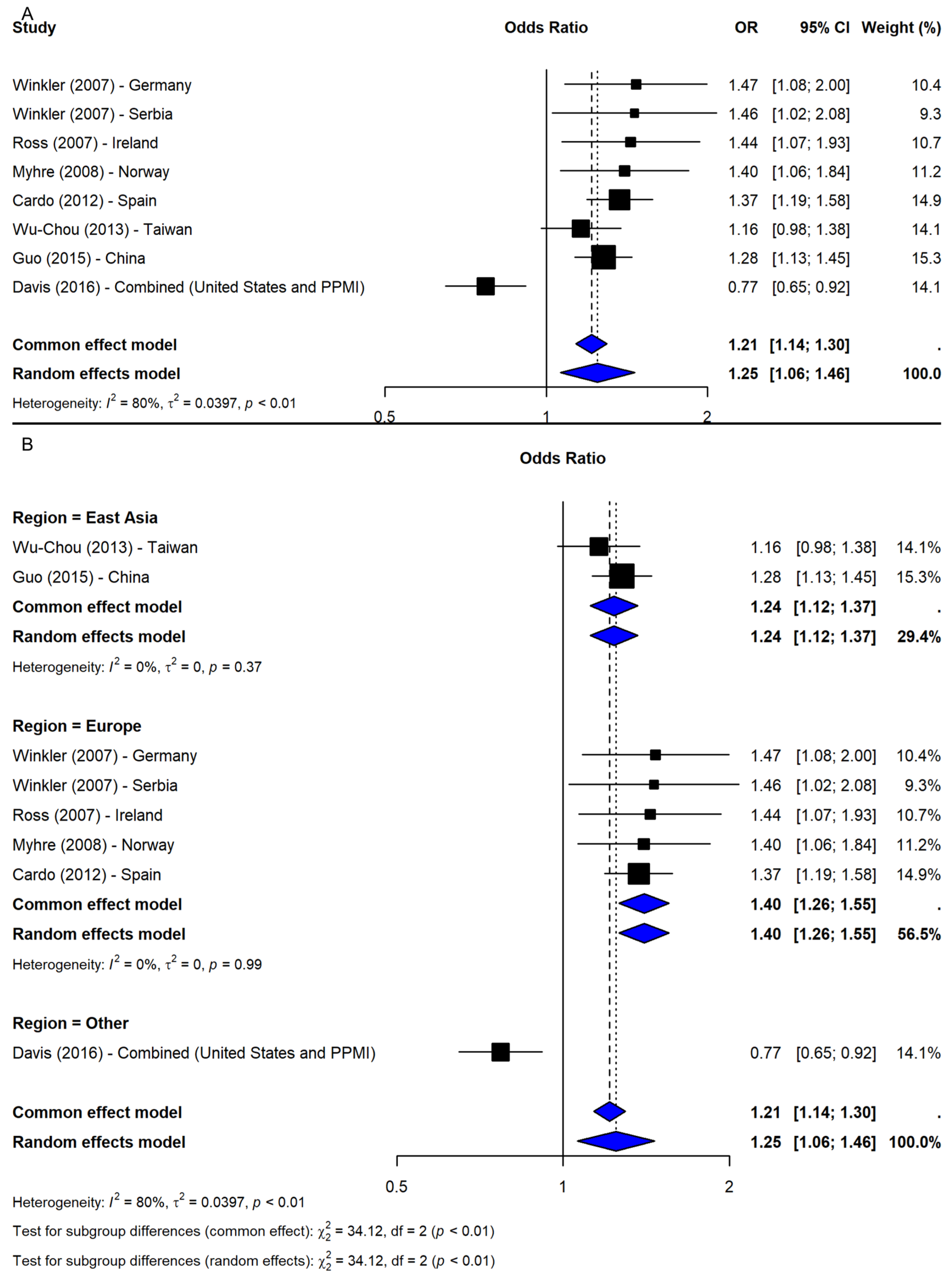
The meta-analysis of SNP rs356165 under the allelic model (G vs. A) included eight studies [2,19,20,21,22,23,24], revealing a significant association between the G allele and increased PD risk (OR = 1.25, 95% CI: 1.06–1.46, p-value = 0.0071; I2 = 80.1%; Figure 2A). Leave-one-out sensitivity analyses confirmed the robustness of the association (OR range: 1.22–1.31). Notably, exclusion of study [21], which combined two datasets (PPMI-European and Washington University), eliminated heterogeneity (I2 = 0%) and increased precision (OR = 1.31, 95% CI: 1.22–1.41), indicating dataset-specific influence. The trim-and-fill method imputed two missing studies; adjusted results remained significant (OR = 1.20, 95% CI: 1.04–1.38, p-value = 0.0136). Subgroup meta-analysis by geographic region showed significant associations in Europe (OR = 1.40, 95% CI: 1.26–1.55) and East Asia (OR = 1.24, 95% CI: 1.12–1.37), while the “Other” category (study [21]) showed an inverse association (OR = 0.77, 95% CI: 0.65–0.92). The subgroup difference was significant (Q = 34.12, df = 2, p-value < 0.0001; Figure 2B), suggesting regional variation contributed to heterogeneity.
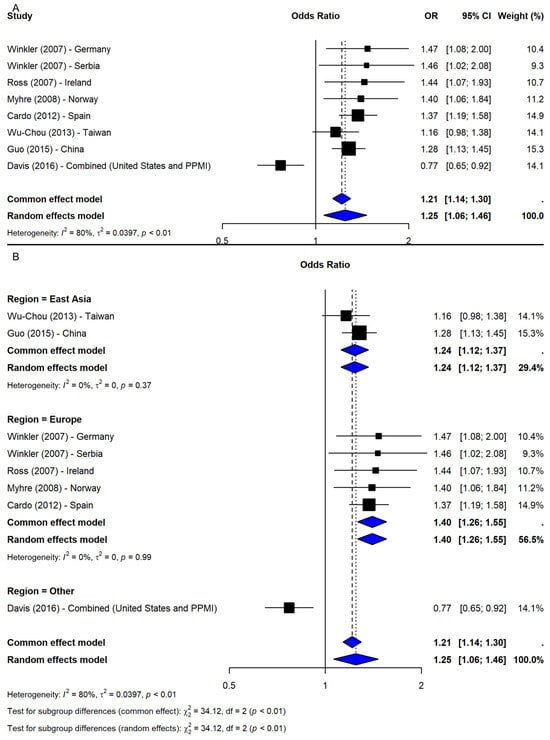
Figure 2.
Association between SNP rs356165 and PD risk under allelic model (G vs. A) [2,19,20,21,22,23,24]: (A) overall meta-analysis; (B) subgroup analyses by geographic region. Black squares represent odds ratios (ORs) from individual studies, with their size reflecting study weight; horizontal lines indicate 95% confidence intervals. Blue diamonds represent the pooled ORs.
Under the dominant model (AG + GG vs. AA), five studies [2,20,22,23,24] showed a significant association (OR = 1.37, 95% CI: 1.21–1.54, p-value < 0.0001; I2 = 7.8%; Supplementary Materials, Figure S1). The sensitivity analysis confirmed stability (OR range: 1.31–1.43), with no publication bias detected. Due to low heterogeneity and limited studies, no subgroup analysis was performed. Similarly, the recessive model (GG vs. AG + AA), using the same five studies [2,20,22,23,24], showed a significant association (OR = 1.50, 95% CI: 1.31–1.71, p-value < 0.0001; I2 = 0%; Supplementary Materials, Figure S2). The sensitivity analysis and publication bias assessments supported consistent findings.
3.2. SNP rs2583988
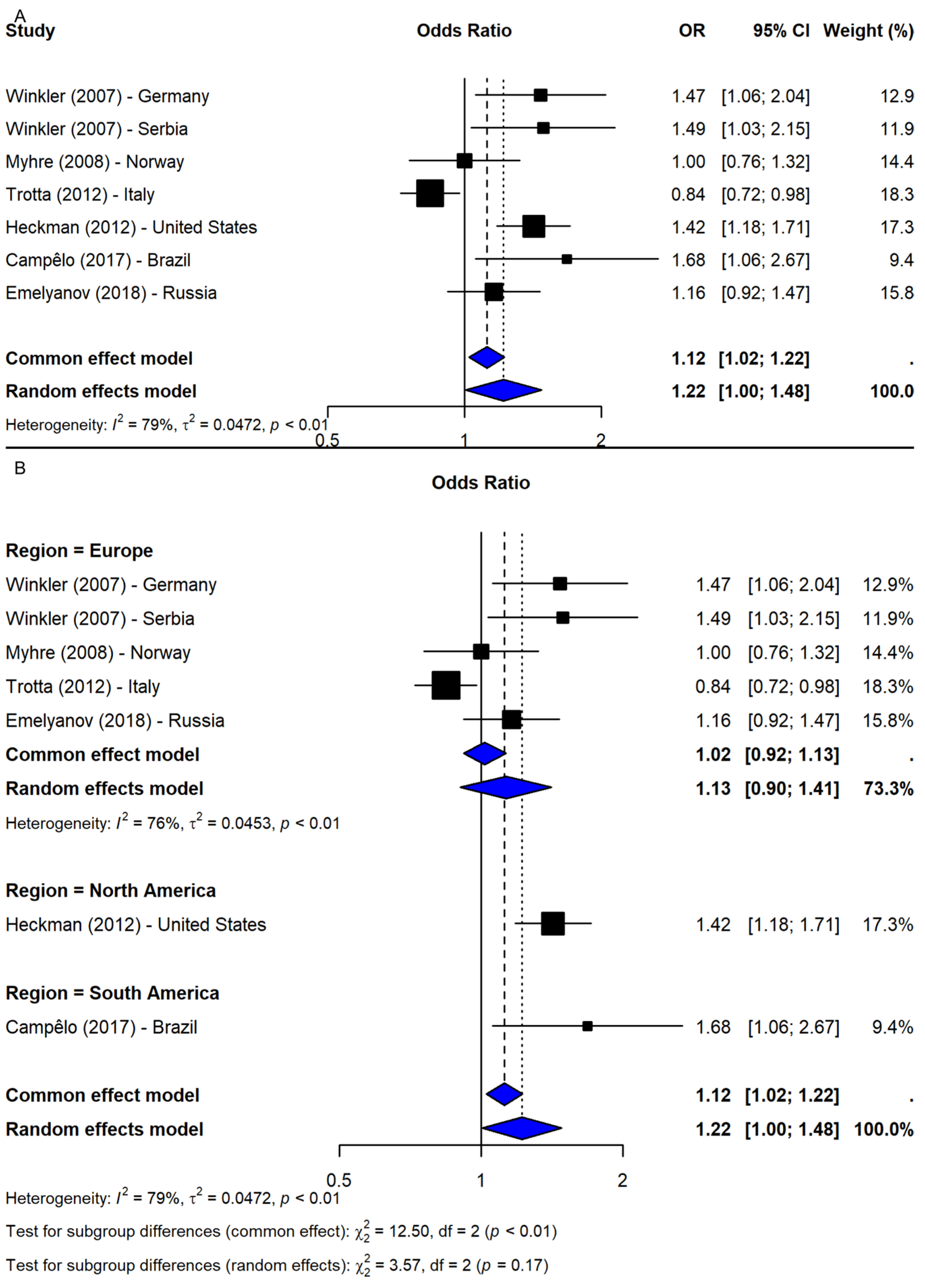
The meta-analysis of rs2583988 under the allelic model (T vs. C) included seven studies [9,19,20,25,26,27] and showed a borderline significant association with PD risk (OR = 1.22, 95% CI: 1.00–1.48, p-value = 0.0446; I2 = 79.2%; Figure 3A). However, the sensitivity analysis revealed that this association was not robust, with most leave-one-out iterations yielding non-significant results. Only when Myhre et al. [20] or Trotta et al. [9] were excluded did the association remain significant, suggesting stabilizing effects from these studies. After adjusting for potential publication bias using the trim-and-fill method, the association became non-significant (OR = 1.07, 95% CI: 0.88–1.30, p-value = 0.524), indicating possible small-study effects. Subgroup analysis by region showed non-significant results in Europe (OR = 1.13, 95% CI: 0.90–1.41; I2 = 75.6%), while significant associations were found in single studies from the U.S. (OR = 1.42, 95% CI: 1.18–1.71) and Brazil (OR = 1.68, 95% CI: 1.06–2.67). However, the test for subgroup differences was not statistically significant (Q = 3.57, df = 2, p-value = 0.1681; Figure 3B), suggesting that the geographic region does not fully account for heterogeneity.
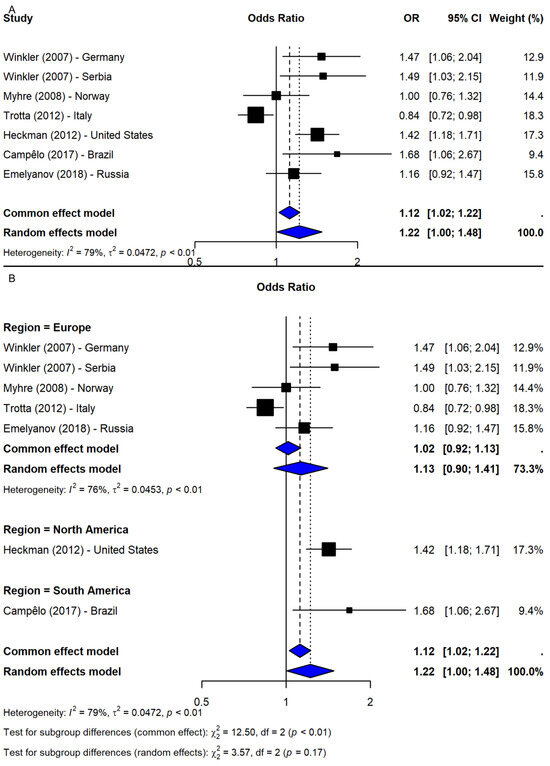
Figure 3.
Association between SNP rs2583988 and PD risk under allelic model (T vs. C) [9,19,20,25,26,27]: (A) overall meta-analysis; (B) subgroup analyses by geographic region. Black squares represent ORs from individual studies, with their size reflecting study weight; horizontal lines indicate 95% confidence intervals. Blue diamonds represent the pooled ORs.
Under the dominant model (TT + TC vs. CC), based on three studies [20,25,26], no significant association was observed (OR = 1.14, 95% CI: 0.93–1.40, p-value = 0.208; I2 = 0%; Supplementary Materials, Figure S3), and findings remained non-significant across the sensitivity and publication bias analyses. Similarly, the recessive model (TT vs. TC + CC) showed no significant association (OR = 1.31, 95% CI: 0.86–2.01, p-value = 0.206; I2 = 55.2%; Supplementary Materials, Figure S4), with instability noted in the sensitivity analysis and no evidence of a publication bias.
3.3. SNP rs356219
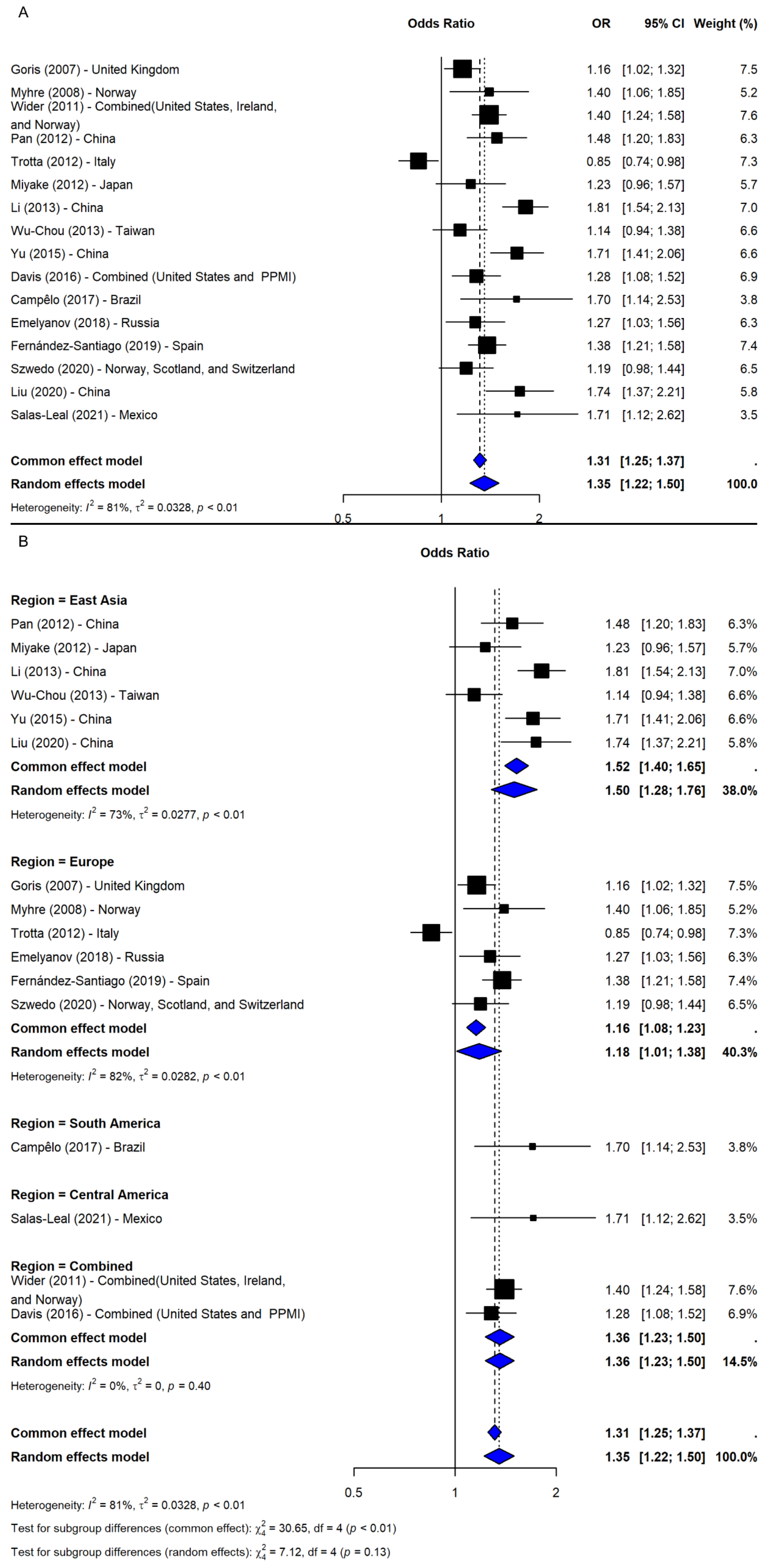
The meta-analysis of rs356219 under the allelic model (G vs. A), including 16 studies [9,16,20,21,24,25,26,28,29,30,31,32,33,34,35,36], showed a significant association with increased PD risk (OR = 1.35, 95% CI: 1.22–1.50, p-value < 0.0001; I2 = 80.7%; Figure 4A). The leave-one-out sensitivity analyses confirmed robustness (OR range: 1.32–1.40), and the association remained significant after adjusting for publication bias (OR = 1.23, 95% CI: 1.10–1.38, p-value = 0.0003), despite imputation of five potentially missing studies. A subgroup analysis by region revealed significant associations across all areas, with the strongest effects in South and Central America (ORs = 1.70 and 1.71, respectively), though based on single studies. Among regions with multiple studies, East Asia showed the most consistent association (OR = 1.50, 95% CI: 1.28–1.76), followed by Combined (OR = 1.36, 95% CI: 1.23–1.50) and Europe (OR = 1.18, 95% CI: 1.01–1.38). However, the test for subgroup differences was not significant (Q = 7.12, df = 4, p-value = 0.1296; Figure 4B).
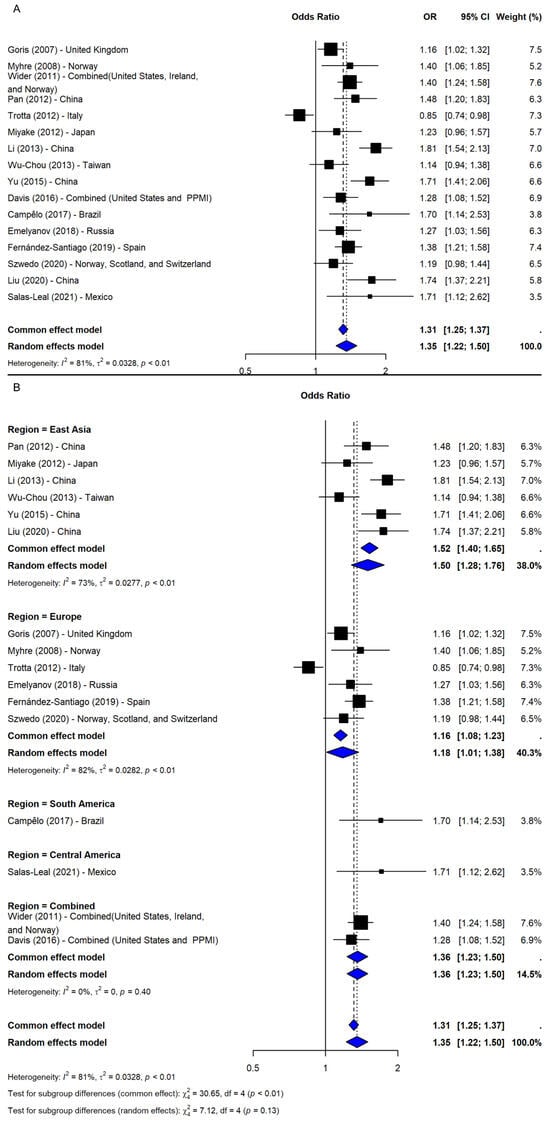
Figure 4.
The association between SNP rs356219 and PD risk under the allelic model (G vs. A) [9,16,20,21,24,25,26,28,29,30,31,32,33,34,35,36]: (A) the overall meta-analysis; (B) the subgroup analyses by geographic region. Black squares represent ORs from individual studies, with their size reflecting study weight; horizontal lines indicate 95% confidence intervals. Blue diamonds represent the pooled ORs.
Under the dominant model (AG + GG vs. AA), 14 studies [16,20,24,25,26,28,29,30,31,32,33,34,35,36] showed a significant association (OR = 1.46, 95% CI: 1.29–1.65, p-value < 0.0001; I2 = 51.8%; Supplementary Materials, Figure S5). The sensitivity analysis confirmed consistency, and the association remained significant after adjusting for three potentially missing studies (OR = 1.36, 95% CI: 1.20–1.55).
The recessive model (GG vs. AG + AA), using the same 14 studies, revealed a stronger association (OR = 1.69, 95% CI: 1.49–1.92, p-value < 0.0001; I2 = 50.2%; Supplementary Materials, Figure S6). The result was stable in the sensitivity analysis and remained significant after adjusting for five potentially missing studies (OR = 1.49, 95% CI: 1.29–1.72). The subgroup analyses for both models showed consistent associations across all regions. In the dominant model, East Asia showed the strongest effect (OR = 1.65, 95% CI: 1.34–2.03), followed by Combined (OR = 1.55, 95% CI: 1.28–1.88) and Europe (OR = 1.26, 95% CI: 1.10–1.43); subgroup differences were not significant (Q = 7.17, df = 4, p-value = 0.1273; Supplementary Materials, Figure S7). Similar trends were observed under the recessive model, with East Asia again showing the highest effect (OR = 1.75, 95% CI: 1.39–2.22), followed by Europe (OR = 1.54, 95% CI: 1.34–1.78) and Combined (OR = 1.53, 95% CI: 1.26–1.86); subgroup differences were also non-significant (Q = 3.44, df = 4, p-value = 0.4863; Supplementary Materials, Figure S8).
3.4. SNP rs11931074
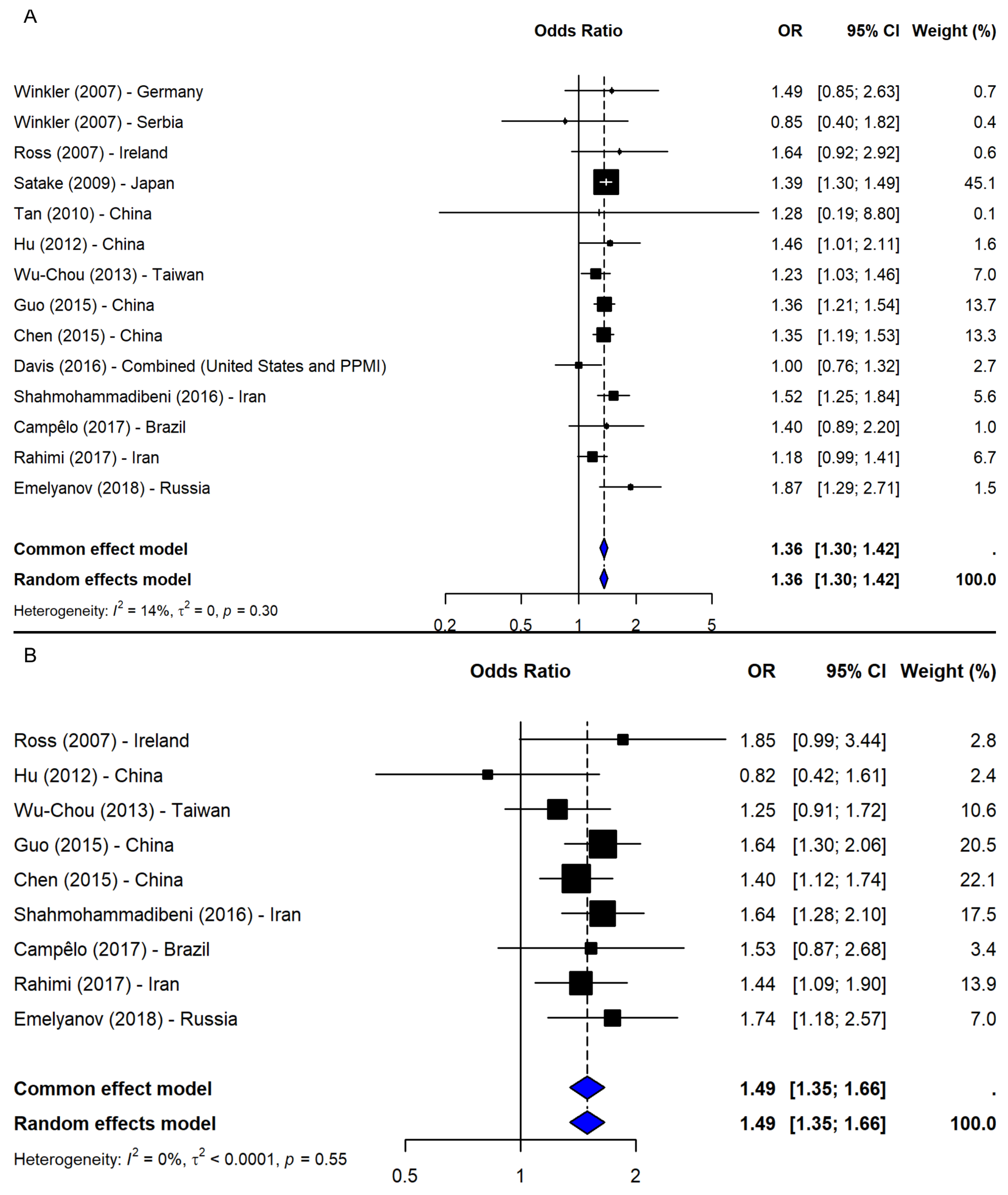
A meta-analysis of 14 studies [2,5,19,21,23,24,25,26,37,38,39,40,41] under the allelic model (T vs. G) showed a significant association between the T allele and increased PD risk (OR = 1.36, 95% CI: 1.30–1.42, p-value < 0.0001; I2 = 13.7%; Figure 5A), with low heterogeneity and no evidence of publication bias. The leave-one-out sensitivity analyses confirmed the stability of the results (OR range: 1.33–1.37). Given the consistency, no subgroup analysis was conducted.
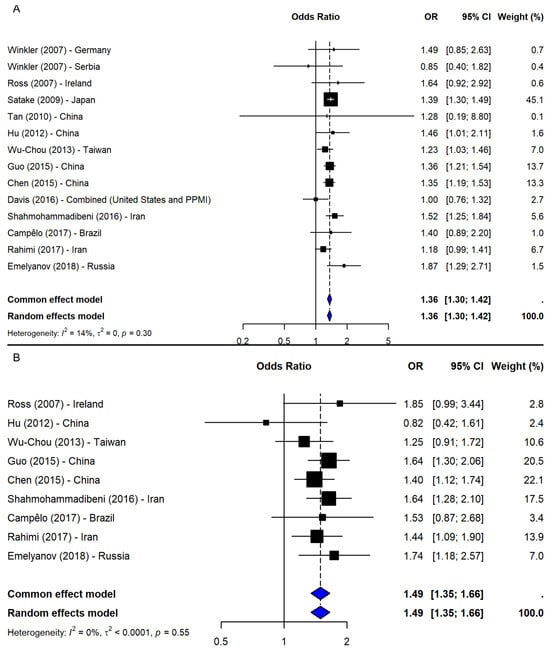
Figure 5.
Association between SNP rs11931074 and PD risk under the (A) allelic model (T vs. G) [2,5,19,21,23,24,25,26,37,38,39,40,41] and the (B) dominant model (TT + TG vs. GG) [2,23,24,25,26,37,38,40,41]. Black squares represent ORs from individual studies, with their size reflecting study weight; horizontal lines indicate 95% confidence intervals. Blue diamonds represent the pooled ORs.
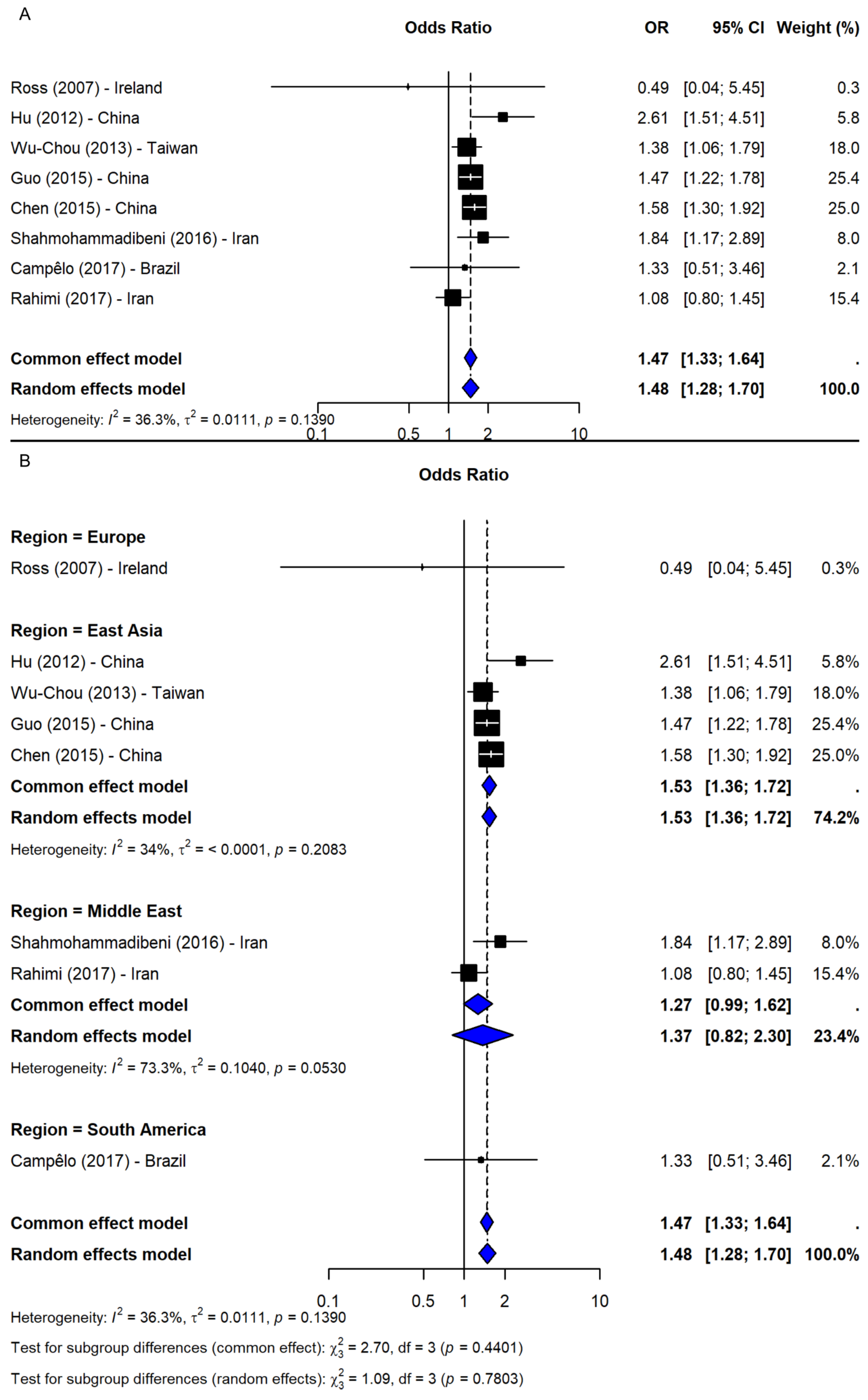
Under the dominant model (TT + TG vs. GG), based on nine studies [2,23,24,25,26,37,38,40,41], a significant association was observed (OR = 1.49, 95% CI: 1.35–1.66, p-value < 0.0001; I2 = 0%), with robust sensitivity results (OR range: 1.46–1.53) and no publication bias (Figure 5B). The recessive model (TT vs. TG + GG), including eight studies [2,23,24,25,37,38,40,41], also showed a significant association (OR = 1.48, 95% CI: 1.28–1.70, p-value < 0.0001; I2 = 36.3%; Figure 6A). The results remained consistent in the sensitivity analysis (OR range: 1.43–1.54), and no missing studies were identified via trim-and-fill. The subgroup analysis for the recessive model showed the strongest association in East Asia (4 studies, OR = 1.53, 95% CI: 1.36–1.72). Associations were weaker and non-significant in the Middle East (2 studies), South America (1 study), and Europe (1 study), though interpretation was limited by small sample sizes. No significant subgroup differences were detected (Q = 1.09, df = 3, p-value = 0.7803; Figure 6B).
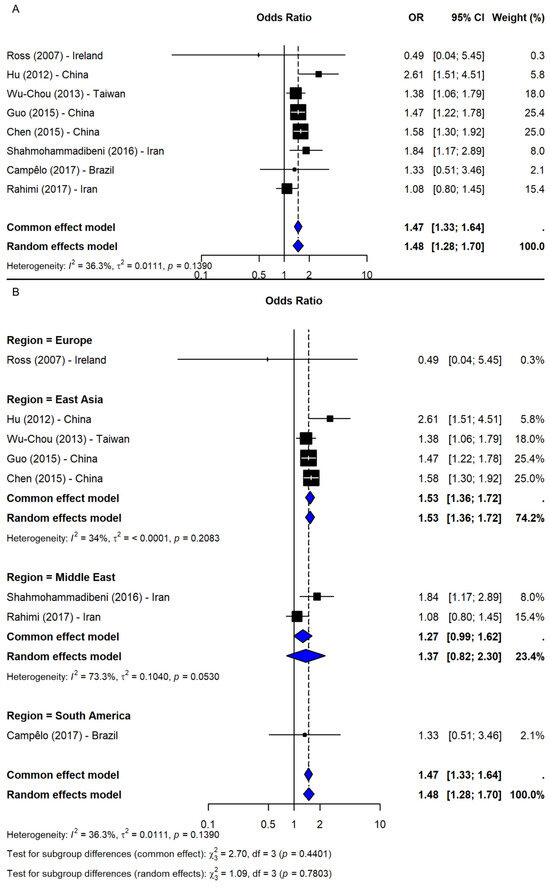
Figure 6.
Association between SNP rs11931074 and PD risk under the recessive model (TT vs. TG + GG) [2,23,24,25,37,38,40,41]: (A) the overall meta-analysis; (B) the subgroup analyses by geographic region. Black squares represent ORs from individual studies, with their size reflecting study weight; horizontal lines indicate 95% confidence intervals. Blue diamonds represent the pooled ORs.
4. Discussion
PD is a complex neurodegenerative disorder, with growing evidence implicating genetic variability in its susceptibility and progression [42]. A hallmark feature of PD is the presence of Lewy bodies, which are intracellular inclusions primarily composed of aggregated alpha-synuclein protein. The alpha-synuclein protein is encoded by the SNCA gene located on chromosome 4q22 [43,44]. While SNCA mutations are rare and typically associated with familial PD, common variants have been shown to influence the risk of sporadic PD. Additionally, extracellular alpha-synuclein levels have been linked to PD diagnosis and severity, supporting its role as a potential biomarker.
Over the past two decades, GWASs have identified over 40 loci significantly associated with PD, highlighting the role of genetic factors in disease pathogenesis [45]. Among these, SNCA has emerged as a key locus due to its central involvement in Lewy body formation and regulation of alpha-synuclein expression. Evidence suggests that SNCA variants can influence both protein levels and transcript diversity, thereby modulating PD risk [13,38,46,47,48,49]. In this context, the present study aimed to systematically evaluate and synthesize existing evidence on the association between SNCA polymorphisms and PD susceptibility through a comprehensive meta-analysis.
In this comprehensive meta-analysis, we evaluated the association between four SNCA single-nucleotide polymorphisms—rs356165, rs2583988, rs356219, and rs11931074—and PD risk. Among the 27 studies included, our results confirmed that rs356165, rs356219, and rs11931074 are significantly associated with increased PD risk across allelic, dominant, and recessive genetic models. Notably, rs356219 demonstrated the strongest association, particularly under the recessive model (OR = 1.69, 95% CI: 1.49–1.92). In contrast, rs2583988 showed a weaker and less consistent association, reaching marginal significance only under the allelic model and not under dominant or recessive models. Subgroup analyses by geographic region revealed generally consistent effects, with particularly strong associations observed in East Asian populations. Our findings align with those of previous large-scale meta-analyses investigating SNCA polymorphisms. Specifically, a comprehensive meta-analysis by Zhang et al. (2018) identified eight SNCA SNPs associated with PD risk, including rs356165, rs356219, and rs11931074—three of the four SNPs evaluated in our study [50]. These variants were also among those with the strongest associations in Zhang et al.’s analysis, with rs11931074 and rs356219 demonstrating increased risk across multiple ethnic groups, including East Asian and European populations. The consistency between our findings and those of Zhang et al. strengthens the evidence supporting these SNCA variants as robust genetic risk factors for PD and highlights their potential relevance in genetic screening strategies.
In the following sections, we discuss the specific findings and implications of each SNCA variant in detail.
4.1. SNP rs356165
Our meta-analysis consistently demonstrates that SNP rs356165 is associated with increased PD risk across allelic, dominant, and recessive models. Heterogeneity in the allelic model was primarily driven by one outlier study; its exclusion improved consistency. Subgroup analyses confirmed significant associations in both European and East Asian populations, though the combined dataset showed an inverse effect, likely due to population-specific differences. The dominant and recessive models yielded stable, significant associations with minimal heterogeneity and no publication bias.
Located in the 3′ untranslated region (3′ UTR) of SNCA, rs356165 plays a regulatory role in post-transcriptional expression. Ghanbari et al. [51] proposed that the G allele disrupts microRNA binding sites, potentially increasing SNCA expression. This aligns with findings of elevated SNCA mRNA and α-synuclein protein levels in G allele carriers versus AA genotypes [42]. These effects may be especially pronounced in brain tissue, where longer 3′ UTRs amplify post-transcriptional regulation. Additional studies further support rs356165’s biological relevance. A Spanish study reported significant association under the dominant model and with earlier age at onset (AAO) [52], while a Chinese study found no significant association [53], indicating possible ethnic differences. Rajput et al. [54] validated the association in a Canadian cohort.
Regarding related phenotypes, a Croatian study found no direct association between rs356165 and PD but reported intermediate linkage disequilibrium with other SNCA 3′ UTR variants (rs1045722 and rs3857053), suggesting shared regulatory mechanisms [42]. A population-based study suggested an additive interaction between head injury and rs356165, with a 3–4.5-fold increased PD risk in carriers post-trauma, though no multiplicative interaction was found [55]. Moreover, rs356165 was more frequent in PD than in idiopathic REM sleep behaviour disorder (iRBD), implying a potential role in prodromal disease progression [56]. A longitudinal study also showed that while rs356165 alone did not predict motor progression, it contributed additively when combined with the REP1 promoter variant [57].
In sum, our meta-analysis and supporting literature provide strong evidence for rs356165’s role in PD susceptibility. Its consistent associations across genetic models, functional implications in SNCA regulation, and links to phenotypic variability and progression underscore its biological significance. Future studies involving functional assays and diverse populations are warranted to further clarify its role in PD pathogenesis.
4.2. SNP rs2583988
Our meta-analysis indicates a possible but inconclusive association between rs2583988 and PD risk. A marginally significant effect was observed under the allelic model; however, this association lacked robustness due to substantial heterogeneity and inconsistent sensitivity analysis results. After adjusting for publication bias, the effect was no longer significant, suggesting small-study influences. Subgroup analysis by region showed no association in European cohorts, while stronger effects were reported in single studies from the United States and Brazil, limiting generalizability. Neither the dominant nor recessive models showed significant associations, indicating that any link between rs2583988 and PD is likely weak and uncertain.
Beyond our analysis, one study found that individuals homozygous for the C allele had a higher risk of rapid motor progression, though this association did not remain significant after correction for multiple comparisons [54]. The overall heterogeneity and methodological differences across studies underscore the need for larger, well-powered investigations to clarify the potential role of rs2583988 in PD susceptibility and progression.
4.3. SNP rs356219
Our meta-analysis provided strong and consistent evidence for an association between SNP rs356219 and increased PD risk across all genetic models. The G allele was significantly linked to higher PD risk, with results remaining stable in sensitivity and publication bias analyses. While heterogeneity was notable under the allelic model, subgroup meta-analyses confirmed the association across all regions, particularly in East Asian populations. The dominant and recessive models also supported this association, with consistent findings across studies and minimal heterogeneity. These findings position the rs356219 G allele, especially in homozygous form, as a robust genetic risk factor for PD across diverse populations. These findings are in line with the previous systematic review and meta-analysis by Hou et al. (2019), which also reported significant associations between rs356219 and PD risk across all genetic models [58].
Subgroup comparisons showed regional variability in effect size, though not statistically significant. The strongest associations were observed in East Asia, followed by combined and European datasets. These findings align with studies from Chinese populations that highlight SNCA as a key contributor to PD susceptibility [9,14,16,30,33,59]. For instance, a large Han Chinese case–control study reported significant associations between the G allele and increased PD risk and earlier AAO [33]. In contrast, Refenes et al. found no association in Greek and Italian cohorts, suggesting ethnic variability [60]. In Japan, rs356219 was not directly associated with PD risk, but a significant additive interaction with smoking was observed [34].
Functional studies support the biological relevance of rs356219. The G allele has been associated with elevated α-synuclein mRNA and protein levels, and its combination with plasma α-synuclein improved early PD diagnosis [31]. Szwedo et al. [28] reported faster cognitive decline in GG carriers, and Goris et al. [32] observed a synergistic effect between rs356219 and MAPT variants in increasing dementia risk. Emelyanov et al. [26] further demonstrated increased α-synuclein in CD45+ blood cells of G allele carriers, indicating regulatory effects on SNCA expression.
Additional studies highlight rs356219 as a disease-modifying variant. In a Spanish LRRK2-PD cohort, the GG genotype was associated with PD onset up to 11 years earlier than the AA genotype [61]. This was supported by regression models showing consistent associations between the risk allele and earlier PD onset [62] and by studies reporting gene–gene interactions between rs356219 and the LRRK2 G2019S mutation [63]. A 30-month longitudinal study also found the G allele linked to reduced motor and cognitive decline risk, especially among rapid eye movement behaviour disorder (RBD) patients, suggesting a potential prognostic role [64].
Complementary evidence has emerged from studies examining rs356219 under the C vs. T model [65,66,67]. Mata et al. (2010) found the C allele to be a stronger predictor of PD risk than REP1 in U.S. cohorts and associated it with elevated plasma α-synuclein under an additive model [65]. Lucchini et al. (2020) reported significantly higher PD risk among CC genotype carriers in an Italian cohort, with additive—but not multiplicative—effects in individuals exposed to metals [68].
Collectively, these findings confirm rs356219 as a strong susceptibility and disease-modifying marker in PD. Its impact on SNCA expression, cognitive decline, and interactions with environmental and genetic factors underscores its multifaceted role in PD pathogenesis, particularly in East Asian populations. Future research should prioritize integrative approaches to clarify its clinical and biological implications across diverse populations.
4.4. SNP rs11931074
Our meta-analysis demonstrated a consistent and significant association between SNP rs11931074 and increased PD risk across allelic, dominant, and recessive models. These associations remained robust in sensitivity analyses, with low heterogeneity and no evidence of publication bias. Subgroup analyses confirmed the association across regions, particularly in East Asian and European populations, supporting rs11931074 as a reliable genetic marker of PD susceptibility.
Located 7.2 kb downstream of the 3′ end of the SNCA gene, rs11931074 has been repeatedly implicated in PD risk [69]. Despite notable differences in allele frequency between populations, where the T allele is more common in Asians (~58%) and less frequent in Caucasians (~7%), an overrepresentation of the T allele among PD patients has been consistently observed across diverse ethnic groups [40,70]. Our findings align with prior GWAS and case–control studies, including one in European populations identifying rs11931074 as a top PD risk variant [4], and others in Chinese and Korean cohorts reporting associations with both disease susceptibility and milder clinical phenotypes, such as hyposmia and less severe motor symptoms [71,72].
Beyond risk association, rs11931074 has been implicated in PD heterogeneity. The T allele is linked to lower serum α-synuclein levels and non-motor symptoms like constipation and REM sleep behaviour disorder [40,72]. TT genotype carriers often exhibit milder motor symptoms, slower progression, and lower comorbidity burden [73,74]. Additional studies report reduced salivary α-synuclein and altered brain activity in TT carriers, specifically increased right angular gyrus activity inversely correlated with motor severity [75,76].
Although some studies report conflicting findings, such as a possible protective role of the G allele in early-onset or familial PD [77] or no association with peripheral α-synuclein levels [26], the broader evidence supports a multifactorial role for rs11931074 in PD risk and phenotype.
Functionally, rs11931074 affects SNCA transcript processing, increasing the expression of shorter isoforms like SNCA-98 and SNCA-112, which may enhance protein aggregation [22]. Our findings are further supported by a recent large-scale meta-analysis of 13 studies involving over 13,000 PD cases and 28,000 controls, which confirmed a robust association between rs11931074 and PD risk across five genetic models, with the strongest effects observed in Asian populations [78]. Furthermore, emerging evidence suggests that rs11931074 may also influence peripheral α-synuclein pathology. Moreover, exploratory research in the enteric nervous system (ENS) found the G allele associated with increased α-synuclein immunostaining (OR = 5.96, p = 0.01) and a significant interaction with PD status, suggesting potential involvement in early or extracerebral PD pathology [79].
Taken together, these findings reinforce rs11931074 as a key susceptibility variant that may also influence the PD phenotype and progression, potentially through mechanisms involving SNCA regulation and α-synuclein expression.
5. Strengths and Limitations
This meta-analysis synthesizes data from 44 studies, with 27 providing sufficient quantitative data for analysis, leveraging robust statistical methods like leave-one-out sensitivity analysis and trim-and-fill to ensure finding stability. It systematically evaluates four SNCA polymorphisms across genetic models, yielding consistent associations for rs356165, rs356219, and rs11931074.
A key strength of our study is its timeliness and comprehensiveness. Unlike previous systematic reviews, which were published before 2019, our review includes studies up to 20 November 2023, allowing us to incorporate a broader and more current evidence base. We also explicitly acknowledged the existence of previous reviews and outlined clear differences in our search timeline, number of included studies, and variants analyzed. These distinctions enhance the relevance and clinical utility of our findings in the current research landscape. However, limitations include the exclusion of 17 studies due to inadequate genotype or OR data, potentially reducing generalizability. Notable heterogeneity, especially in allelic models of rs356165 (I2 = 80.1%) and rs356219 (I2 = 80.7%), suggests variability from study design, population differences, or unmeasured factors (e.g., genotyping accuracy, environmental exposures). Additionally, the geographic distribution of studies is skewed toward European and East Asian populations, with limited representation from other ethnic groups, which may restrict the global generalizability of the findings.
6. Implications and Future Directions
This meta-analysis highlights the contribution of SNCA polymorphisms—particularly rs356165, rs11931074, and rs356219—to PD risk across genetic models and populations. These variants may aid in polygenic risk stratification and selection for α-synuclein-targeted therapies. Their regulatory effects also suggest potential as progression biomarkers. Heterogeneity observed in rs356165, rs2583988, and rs356219 underscores the need for larger, multi-ethnic studies, especially in underrepresented African and South American populations. Integrating genetic, environmental, and clinical data is essential to understanding PD’s etiology. As common variants explain only ~27% of PD heritability [17], polygenic models, functional studies, and longitudinal cohorts are needed to clarify variant effects on disease onset and progression.
7. Conclusions
This meta-analysis provides strong evidence that SNCA variants—particularly rs11931074—are robustly associated with increased PD risk across diverse populations and genetic models, with consistent findings, low heterogeneity, and no publication bias. Rs356165 and rs356219 also showed significant associations across all models, though high heterogeneity limits interpretability. In contrast, rs2583988 demonstrated weaker, context-dependent associations with greater variability across studies. Despite these limitations, our findings reinforce SNCA’s central role in PD pathogenesis and underscore the importance of future large-scale, ethnically diverse studies to explore gene–environment and gene–gene interactions and support the development of personalized therapeutic strategies.
Supplementary Materials
The supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms26136001/s1.
Author Contributions
Conceptualization, R.M., M.S., S.F.S.-M., M.Z.Y.C.L., C.L.S., S.M.H.F., J.Y.T., X.D. and S.Y.E.N.; methodology, R.M. and S.E.S.; Software, R.M. and S.E.S.; validation, R.M., M.S., S.F.S.-M., M.Z.Y.C.L., C.L.S., S.M.H.F., J.Y.T., X.D., S.Y.E.N., A.S.L.N., L.C.S.T. and S.E.S.; formal analysis, R.M. and S.E.S.; investigation, R.M., M.S., S.F.S.-M., M.Z.Y.C.L., C.L.S., S.M.H.F., J.Y.T. and S.Y.E.N.; resources, R.M. and S.E.S.; data curation, R.M., M.S., S.F.S.-M., M.Z.Y.C.L., C.L.S., S.M.H.F., J.Y.T., X.D. and S.Y.E.N.; writing—original draft preparation, R.M.; writing—review and editing, R.M., S.E.S., A.S.L.N., L.C.S.T. and S.E.S.; visualization, R.M. and S.E.S.; supervision, A.S.L.N., L.C.S.T. and S.E.S.; project administration, R.M.; funding acquisition, L.C.S.T. and S.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Singapore Ministry of Health’s National Medical Research Council (MOH-OFLCG18May-0002, MOH-CSAINV21-0005, CNIG22jul-0004).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study (i.e., extracted study-level information) are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| PD | Parkinson’s Disease |
| SNP | Single-nucleotide polymorphisms |
| PD-MCI | Parkinson’s disease-mild cognitive impairment |
| GCTA | Genome-wide complex trait analysis |
| GWAS | Genome-wide association studies |
| OR | Odds ratios |
| CI | Confidence interval |
| NOS | Newcastle-Ottawa Scale |
| PDD | Parkinson’s disease dementia |
| AAO | Earlier age at onset |
| 3′ UTR | 3′ untranslated region |
| iRBD | Idiopathic REM sleep behaviour disorder |
References
- Maraganore, D.M.; de Andrade, M.; Elbaz, A.; Farrer, M.J.; Ioannidis, J.P.; Krüger, R.; Rocca, W.A.; Schneider, N.K.; Lesnick, T.G.; Lincoln, S.J.; et al. Collaborative analysis of alpha-synuclein gene promoter variability and Parkinson disease. JAMA 2006, 296, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.F.; Li, K.; Yu, R.L.; Sun, Q.Y.; Wang, L.; Yao, L.Y.; Hu, Y.C.; Lv, Z.Y.; Luo, L.Z.; Shen, L.; et al. Polygenic determinants of Parkinson’s disease in a Chinese population. Neurobiol. Aging 2015, 36, 1765.e1–1765.e6. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.C.; Fuchs, J.; Hofer, A.; Zimprich, A.; Lichtner, P.; Illig, T.; Berg, D.; Wüllner, U.; Meitinger, T.; Gasser, T. Multiple regions of α-synuclein are associated with Parkinson’s disease. Ann. Neurol. 2005, 57, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Simón-Sánchez, J.; Schulte, C.; Bras, J.M.; Sharma, M.; Gibbs, J.R.; Berg, D.; Paisan-Ruiz, C.; Lichtner, P.; Scholz, S.W.; Hernandez, D.G.; et al. Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat. Genet. 2009, 41, 1308–1312. [Google Scholar] [CrossRef]
- Satake, W.; Nakabayashi, Y.; Mizuta, I.; Hirota, Y.; Ito, C.; Kubo, M.; Kawaguchi, T.; Tsunoda, T.; Watanabe, M.; Takeda, A.; et al. Genome-wide association study identifies common variants at four loci as genetic risk factors for Parkinson’s disease. Nat. Genet. 2009, 41, 1303–1307. [Google Scholar] [CrossRef]
- Kruger, R. Ala 30 Pro mutation in the gene encoding α synuclein in Parkinson disease. Nat. Genet. 1998, 18, 106–108. [Google Scholar] [CrossRef]
- Zarranz, J.J.; Alegre, J.; Gómez-Esteban, J.C.; Lezcano, E.; Ros, R.; Ampuero, I.; Vidal, L.; Hoenicka, J.; Rodriguez, O.; Atarés, B. The new mutation, E46K, of α-synuclein causes parkinson and Lewy body dementia. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 2004, 55, 164–173. [Google Scholar] [CrossRef]
- Hadjigeorgiou, G.M.; Xiromerisiou, G.; Gourbali, V.; Aggelakis, K.; Scarmeas, N.; Papadimitriou, A.; Singleton, A. Association of alpha-synuclein Rep1 polymorphism and Parkinson’s disease: Influence of Rep1 on age at onset. Mov. Disord. 2006, 21, 534–539. [Google Scholar] [CrossRef]
- Trotta, L.; Guella, I.; Soldà, G.; Sironi, F.; Tesei, S.; Canesi, M.; Pezzoli, G.; Goldwurm, S.; Duga, S.; Asselta, R. SNCA and MAPT genes: Independent and joint effects in Parkinson disease in the Italian population. Park. Relat. Disord. 2012, 18, 257–262. [Google Scholar] [CrossRef]
- Krüger, R.; Vieira-Saecker, A.M.; Kuhn, W.; Berg, D.; Müller, T.; Kühnl, N.; Fuchs, G.A.; Storch, A.; Hungs, M.; Woitalla, D.; et al. Increased susceptibility to sporadic Parkinson’s disease by a certain combined alpha-synuclein/apolipoprotein E genotype. Ann. Neurol. 1999, 45, 611–617. [Google Scholar] [CrossRef]
- Lesage, S.; Brice, A. Parkinson’s disease: From monogenic forms to genetic susceptibility factors. Hum. Mol. Genet. 2009, 18, R48–R59. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zang, Q.; Hu, F.; Wei, H.; Ma, J.; Xu, Y. Alpha-synuclein gene polymorphism affects risk of dementia in Han Chinese with Parkinson’s disease. Neurosci. Lett. 2019, 706, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Ning, Y.; Saffari, S.E.; Xiao, B.; Niu, C.; Ng, S.Y.E.; Chia, N.; Choi, X.; Heng, D.L.; Tan, Y.J.; et al. Identifying clinical features and blood biomarkers associated with mild cognitive impairment in Parkinson disease using machine learning. Eur. J. Neurol. 2023, 30, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, A.; Ross, O.A.; Ioannidis, J.P.A.; Soto-Ortolaza, A.I.; Moisan, F.; Aasly, J.; Annesi, G.; Bozi, M.; Brighina, L.; Chartier-Harlin, M.C. Independent and joint effects of the MAPT and SNCA genes in Parkinson disease. Ann. Neurol. 2011, 69, 778–792. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Scott, W.K.; Wang, G.; Mayhew, G.; Li, Y.J.; Vance, J.M.; Martin, E.R. Gene-gene interaction between FGF20 and MAOB in Parkinson disease. Ann. Hum. Genet. 2008, 72, 157–162. [Google Scholar] [CrossRef]
- Wider, C.; Vilariño-Güell, C.; Heckman, M.G.; Jasinska-Myga, B.; Ortolaza-Soto, A.I.; Diehl, N.N.; Crook, J.E.; Cobb, S.A.; Bacon, J.A.; Aasly, J.O.; et al. SNCA, MAPT, and GSK3B in Parkinson disease: A gene-gene interaction study. Eur. J. Neurol. 2011, 18, 876–881. [Google Scholar] [CrossRef]
- Keller, M.F.; Saad, M.; Bras, J.; Bettella, F.; Nicolaou, N.; Simón-Sánchez, J.; Mittag, F.; Büchel, F.; Sharma, M.; Gibbs, J.R. Using genome-wide complex trait analysis to quantify ‘missing heritability’in Parkinson’s disease. Hum. Mol. Genet. 2012, 21, 4996–5009. [Google Scholar] [CrossRef]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses. Available online: http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 9 April 2025).
- Winkler, S.; Hagenah, J.; Lincoln, S.; Heckman, M.; Haugarvoll, K.; Lohmann-Hedrich, K.; Kostic, V.; Farrer, M.; Klein, C. alpha-Synuclein and Parkinson disease susceptibility. Neurology 2007, 69, 1745–1750. [Google Scholar] [CrossRef]
- Myhre, R.; Toft, M.; Kachergus, J.; Hulihan, M.M.; Aasly, J.O.; Klungland, H.; Farrer, M.J. Multiple alpha-synuclein gene polymorphisms are associated with Parkinson’s disease in a Norwegian population. Acta Neurol. Scand. 2008, 118, 320–327. [Google Scholar] [CrossRef]
- Davis, A.A.; Andruska, K.M.; Benitez, B.A.; Racette, B.A.; Perlmutter, J.S.; Cruchaga, C. Variants in GBA, SNCA, and MAPT influence Parkinson disease risk, age at onset, and progression. Neurobiol. Aging 2016, 37, 209.e1–209.e7. [Google Scholar] [CrossRef]
- Cardo, L.F.; Coto, E.; de Mena, L.; Ribacoba, R.; Mata, I.F.; Menéndez, M.; Moris, G.; Alvarez, V. Alpha-synuclein transcript isoforms in three different brain regions from Parkinson’s disease and healthy subjects in relation to the SNCA rs356165/rs11931074 polymorphisms. Neurosci. Lett. 2014, 562, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Ross, O.A.; Gosal, D.; Stone, J.T.; Lincoln, S.J.; Heckman, M.G.; Irvine, G.B.; Johnston, J.A.; Gibson, J.M.; Farrer, M.J.; Lynch, T. Familial genes in sporadic disease: Common variants of alpha-synuclein gene associate with Parkinson’s disease. Mech. Ageing Dev. 2007, 128, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Wu-Chou, Y.H.; Chen, Y.T.; Yeh, T.H.; Chang, H.C.; Weng, Y.H.; Lai, S.C.; Huang, C.L.; Chen, R.S.; Huang, Y.Z.; Chen, C.C.; et al. Genetic variants of SNCA and LRRK2 genes are associated with sporadic PD susceptibility: A replication study in a Taiwanese cohort. Park. Relat. Disord. 2013, 19, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Campêlo, C.L.C.; Cagni, F.C.; de Siqueira Figueredo, D.; Oliveira, L.G., Jr.; Silva-Neto, A.B.; Macêdo, P.T.; Santos, J.R.; Izídio, G.S.; Ribeiro, A.M.; de Andrade, T.G.; et al. Variants in SNCA Gene Are Associated with Parkinson’s Disease Risk and Cognitive Symptoms in a Brazilian Sample. Front. Aging Neurosci. 2017, 9, 198. [Google Scholar] [CrossRef]
- Emelyanov, A.; Kulabukhova, D.; Garaeva, L.; Senkevich, K.; Verbitskaya, E.; Nikolaev, M.; Andoskin, P.; Kopytova, A.; Milyukhina, I.; Yakimovskii, A.; et al. SNCA variants and alpha-synuclein level in CD45+ blood cells in Parkinson’s disease. J. Neurol. Sci. 2018, 395, 135–140. [Google Scholar] [CrossRef]
- Heckman, M.G.; Soto-Ortolaza, A.I.; Diehl, N.N.; Carrasquillo, M.M.; Uitti, R.J.; Wszolek, Z.K.; Graff-Radford, N.R.; Ross, O.A. Evaluation of the role of SNCA variants in survival without neurological disease. PLoS ONE 2012, 7, e42877. [Google Scholar] [CrossRef]
- Szwedo, A.A.; Pedersen, C.C.; Ushakova, A.; Forsgren, L.; Tysnes, O.B.; Counsell, C.E.; Alves, G.; Lange, J.; Macleod, A.D.; Maple-Grødem, J. Association of SNCA Parkinson’s Disease Risk Polymorphisms With Disease Progression in Newly Diagnosed Patients. Front. Neurol. 2020, 11, 620585. [Google Scholar] [CrossRef]
- Fernández-Santiago, R.; Martín-Flores, N.; Antonelli, F.; Cerquera, C.; Moreno, V.; Bandres-Ciga, S.; Manduchi, E.; Tolosa, E.; Singleton, A.B.; Moore, J.H.; et al. SNCA and mTOR Pathway Single Nucleotide Polymorphisms Interact to Modulate the Age at Onset of Parkinson’s Disease. Mov. Disord. 2019, 34, 1333–1344. [Google Scholar] [CrossRef]
- Pan, F.; Dong, H.; Ding, H.; Ye, M.; Liu, W.; Wu, Y.; Zhang, X.; Chen, Z.; Luo, Y.; Ding, X. SNP rs356219 of the α-synuclein (SNCA) gene is associated with Parkinson’s disease in a Chinese Han population. Park. Relat. Disord. 2012, 18, 632–634. [Google Scholar] [CrossRef]
- Salas-Leal, A.C.; Salas-Pacheco, S.M.; Gavilán-Ceniceros, J.A.P.; Castellanos-Juárez, F.X.; Méndez-Hernández, E.M.; La Llave-León, O.; Camacho-Luis, A.; Quiñones-Canales, G.; Romero-Gutiérrez, E.; Arias-Carrión, O.; et al. α-syn and SNP rs356219 as a potential biomarker in blood for Parkinson’s disease in Mexican Mestizos. Neurosci. Lett. 2021, 754, 135901. [Google Scholar] [CrossRef]
- Goris, A.; Williams-Gray, C.H.; Clark, G.R.; Foltynie, T.; Lewis, S.J.; Brown, J.; Ban, M.; Spillantini, M.G.; Compston, A.; Burn, D.J.; et al. Tau and alpha-synuclein in susceptibility to, and dementia in, Parkinson’s disease. Ann. Neurol. 2007, 62, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Li, N.N.; Mao, X.Y.; Chang, X.L.; Zhao, D.M.; Zhang, J.H.; Liao, Q.; Yu, W.J.; Tan, E.K.; Peng, R. SNCA rs356219 variant increases risk of sporadic Parkinson’s disease in ethnic Chinese. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2013, 162b, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Sasaki, S.; Tsuboi, Y.; Yamada, T.; Oeda, T.; Shimada, H.; Kawamura, N.; et al. SNCA polymorphisms, smoking, and sporadic Parkinson’s disease in Japanese. Park. Relat. Disord. 2012, 18, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.J.; Cheng, L.; Li, N.N.; Wang, L.; Tan, E.K.; Peng, R. Interaction between SNCA, LRRK2 and GAK increases susceptibility to Parkinson’s disease in a Chinese population. eNeurologicalSci 2015, 1, 3–6. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, H.; Gen, S.; Zhang, H.; Wang, S.; Hua, L.; Yang, X.; Zhang, S.; Li, J.; Wang, Y. Interaction between SNCA gene polymorphisms and T2DM with Parkinson’s disease. Acta Neurol. Scand. 2020, 142, 443–448. [Google Scholar] [CrossRef]
- Shahmohammadibeni, N.; Rahimi-Aliabadi, S.; Jamshidi, J.; Emamalizadeh, B.; Shahmohammadibeni, H.A.; Zare Bidoki, A.; Akhavan-Niaki, H.; Eftekhari, H.; Abdollahi, S.; Shekari Khaniani, M.; et al. The analysis of association between SNCA, HUSEYO and CSMD1 gene variants and Parkinson’s disease in Iranian population. Neurol. Sci. 2016, 37, 731–736. [Google Scholar] [CrossRef]
- Chen, Y.; Wei, Q.Q.; Ou, R.; Cao, B.; Chen, X.; Zhao, B.; Guo, X.; Yang, Y.; Chen, K.; Wu, Y.; et al. Genetic Variants of SNCA Are Associated with Susceptibility to Parkinson’s Disease but Not Amyotrophic Lateral Sclerosis or Multiple System Atrophy in a Chinese Population. PLoS ONE 2015, 10, e0133776. [Google Scholar] [CrossRef]
- Tan, E.K.; Kwok, H.H.; Tan, L.C.; Zhao, W.T.; Prakash, K.M.; Au, W.L.; Pavanni, R.; Ng, Y.Y.; Satake, W.; Zhao, Y.; et al. Analysis of GWAS-linked loci in Parkinson disease reaffirms PARK16 as a susceptibility locus. Neurology 2010, 75, 508–512. [Google Scholar] [CrossRef]
- Hu, Y.; Tang, B.; Guo, J.; Wu, X.; Sun, Q.; Shi, C.; Hu, L.; Wang, C.; Wang, L.; Tan, L.; et al. Variant in the 3’ region of SNCA associated with Parkinson’s disease and serum α-synuclein levels. J. Neurol. 2012, 259, 497–504. [Google Scholar] [CrossRef]
- Rahimi, M.; Akbari, M.; Jamshidi, J.; Tafakhori, A.; Emamalizadeh, B.; Darvish, H. Genetic analysis of SNCA gene polymorphisms in Parkinson’s disease in an Iranian population. Basal Ganglia 2017, 10, 4–7. [Google Scholar] [CrossRef]
- Blažeković, A.; Jerčić, K.G.; Borovečki, F. SNCA 3’ UTR Genetic Variants in Patients with Parkinson’s Disease. Biomolecules 2021, 11, 1799. [Google Scholar] [CrossRef] [PubMed]
- Mollenhauer, B.; Locascio, J.J.; Schulz-Schaeffer, W.; Sixel-Döring, F.; Trenkwalder, C.; Schlossmacher, M.G. α-Synuclein and tau concentrations in cerebrospinal fluid of patients presenting with parkinsonism: A cohort study. Lancet Neurol. 2011, 10, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Tichopad, A.; Golub, Y.; Munz, M.; Schweitzer, K.J.; Wolf, B.; Berg, D.; Mueller, J.C.; Gasser, T. Genetic variability in the SNCA gene influences alpha-synuclein levels in the blood and brain. FASEB J. 2008, 22, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Blauwendraat, C.; Heilbron, K.; Vallerga, C.L.; Bandres-Ciga, S.; von Coelln, R.; Pihlstrøm, L.; Simón-Sánchez, J.; Schulte, C.; Sharma, M.; Krohn, L.; et al. Parkinson’s disease age at onset genome-wide association study: Defining heritability, genetic loci, and α-synuclein mechanisms. Mov. Disord. 2019, 34, 866–875. [Google Scholar] [CrossRef]
- Ng, A.S.L.; Tan, Y.J.; Zhao, Y.; Saffari, S.E.; Lu, Z.; Ng, E.Y.L.; Ng, S.Y.E.; Chia, N.S.Y.; Setiawan, F.; Xu, Z. SNCA Rep1 promoter variability influences cognition in Parkinson’s disease. Mov. Disord. 2019, 34, 1232–1236. [Google Scholar] [CrossRef]
- Ng, A.S.L.; Tan, Y.J.; Lu, Z.; Ng, E.Y.L.; Ng, S.Y.E.; Chia, N.S.Y.; Setiawan, F.; Xu, Z.; Tay, K.Y.; Prakash, K.M. Plasma alpha-synuclein detected by single molecule array is increased in PD. Ann. Clin. Transl. Neurol. 2019, 6, 615–619. [Google Scholar] [CrossRef]
- Edwards, T.L.; Scott, W.K.; Almonte, C.; Burt, A.; Powell, E.H.; Beecham, G.W.; Wang, L.; Züchner, S.; Konidari, I.; Wang, G.; et al. Genome-wide association study confirms SNPs in SNCA and the MAPT region as common risk factors for Parkinson disease. Ann. Hum. Genet. 2010, 74, 97–109. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Makarious, M.B.; Leonard, H.L.; Bandres-Ciga, S.; Iwaki, H.; Nalls, M.A.; Noyce, A.J.; Singleton, A.B. A population scale analysis of rare SNCA variation in the UK Biobank. Neurobiol. Dis. 2021, 148, 105182. [Google Scholar] [CrossRef]
- Zhang, Y.; Shu, L.; Sun, Q.; Pan, H.; Guo, J.; Tang, B. A comprehensive analysis of the association between SNCA polymorphisms and the risk of Parkinson’s disease. Front. Mol. Neurosci. 2018, 11, 391. [Google Scholar] [CrossRef]
- Ghanbari, M.; Darweesh, S.K.; de Looper, H.W.; van Luijn, M.M.; Hofman, A.; Ikram, M.A.; Franco, O.H.; Erkeland, S.J.; Dehghan, A. Genetic Variants in MicroRNAs and Their Binding Sites Are Associated with the Risk of Parkinson Disease. Hum. Mutat. 2016, 37, 292–300. [Google Scholar] [CrossRef]
- Cardo, L.F.; Coto, E.; De Mena, L.; Ribacoba, R.; Lorenzo-Betancor, O.; Pastor, P.; Samaranch, L.; Mata, I.F.; Díaz, M.; Moris, G.; et al. A search for SNCA 3′ UTR variants identified SNP rs356165 as a determinant of disease risk and onset age in Parkinson’s disease. J. Mol. Neurosci. 2012, 47, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.Y.; Hu, W.B.; Liu, L.; Yu, L.H.; Xi, J.; He, X.H.; Zhu, M.R.; Liu, Z.L.; Xu, Y.M. Lack of replication of a previously reported association between polymorphism in the 3’UTR of the alpha-synuclein gene and Parkinson’s disease in Chinese subjects. Neurosci. Lett. 2010, 479, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.; Vilariño-Güell, C.; Rajput, M.L.; Ross, O.A.; Soto-Ortolaza, A.I.; Lincoln, S.J.; Cobb, S.A.; Heckman, M.G.; Farrer, M.J.; Rajput, A. Alpha-synuclein polymorphisms are associated with Parkinson’s disease in a Saskatchewan population. Mov. Disord. 2009, 24, 2411–2414. [Google Scholar] [CrossRef]
- Lee, P.C.; Bordelon, Y.; Bronstein, J.; Sinsheimer, J.S.; Farrer, M.; Ritz, B. Head injury, α-synuclein genetic variability and Parkinson’s disease. Eur. J. Neurol. 2015, 22, 874–878. [Google Scholar] [CrossRef]
- Toffoli, M.; Dreussi, E.; Cecchin, E.; Valente, M.; Sanvilli, N.; Montico, M.; Gagno, S.; Garziera, M.; Polano, M.; Savarese, M.; et al. SNCA 3′UTR genetic variants in patients with Parkinson’s disease and REM sleep behavior disorder. Neurol. Sci. 2017, 38, 1233–1240. [Google Scholar] [CrossRef]
- Ritz, B.; Rhodes, S.L.; Bordelon, Y.; Bronstein, J. α-Synuclein Genetic Variants Predict Faster Motor Symptom Progression in Idiopathic Parkinson Disease. PLoS ONE 2012, 7, e36199. [Google Scholar] [CrossRef]
- Hou, B.; Zhang, X.; Liu, Z.; Wang, J.; Xie, A. Association of rs356219 and rs3822086 polymorphisms with the risk of Parkinson’s disease: A meta-analysis. Neurosci. Lett. 2019, 709, 134380. [Google Scholar] [CrossRef]
- Tan, E.K.; Chai, A.; Teo, Y.Y.; Zhao, Y.; Tan, C.; Shen, H.; Chandran, V.R.; Teoh, M.L.; Yih, Y.; Pavanni, R.; et al. Alpha-synuclein haplotypes implicated in risk of Parkinson’s disease. Neurology 2004, 62, 128–131. [Google Scholar] [CrossRef]
- Refenes, N.; Kreutz, R.; Bolbrinker, J.; Tagaris, G.; Orlacchio, A.; Drakoulis, N. Non-replication of Association between MAPT-SNCA Synergistical Interaction and Susceptibility to Parkinson’s Disease in a Southern European population. Rev. Clin. Pharmacol. Pharmacokinet. Int. 2010, 24, 205–207. [Google Scholar]
- Fernández-Santiago, R.; Garrido, A.; Infante, J.; González-Aramburu, I.; Sierra, M.; Fernández, M.; Valldeoriola, F.; Muñoz, E.; Compta, Y.; Martí, M.J.; et al. α-synuclein (SNCA) but not dynamin 3 (DNM3) influences age at onset of leucine-rich repeat kinase 2 (LRRK2) Parkinson’s disease in Spain. Mov. Disord. 2018, 33, 637–641. [Google Scholar] [CrossRef]
- Brockmann, K.; Schulte, C.; Hauser, A.K.; Lichtner, P.; Huber, H.; Maetzler, W.; Berg, D.; Gasser, T. SNCA: Major genetic modifier of age at onset of Parkinson’s disease. Mov. Disord. 2013, 28, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Botta-Orfila, T.; Ezquerra, M.; Pastor, P.; Fernández-Santiago, R.; Pont-Sunyer, C.; Compta, Y.; Lorenzo-Betancor, O.; Samaranch, L.; Martí, M.J.; Valldeoriola, F. Age at onset in LRRK2-associated PD is modified by SNCA variants. J. Mol. Neurosci. 2012, 48, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Li, Y.; Niu, M.; Zhou, L.; Yao, M.; Zhu, L.; Ye, G.; Kang, W.; Liu, J. Variants in the SNCA Locus Are Associated with the Progression of Parkinson’s Disease. Front. Aging Neurosci. 2019, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Mata, I.F.; Shi, M.; Agarwal, P.; Chung, K.A.; Edwards, K.L.; Factor, S.A.; Galasko, D.R.; Ginghina, C.; Griffith, A.; Higgins, D.S.; et al. SNCA variant associated with Parkinson disease and plasma alpha-synuclein level. Arch. Neurol. 2010, 67, 1350–1356. [Google Scholar] [CrossRef]
- Mata, I.F.; Yearout, D.; Alvarez, V.; Coto, E.; de Mena, L.; Ribacoba, R.; Lorenzo-Betancor, O.; Samaranch, L.; Pastor, P.; Cervantes, S.; et al. Replication of MAPT and SNCA, but not PARK16-18, as susceptibility genes for Parkinson’s disease. Mov. Disord. 2011, 26, 819–823. [Google Scholar] [CrossRef]
- Westerlund, M.; Belin, A.C.; Anvret, A.; Håkansson, A.; Nissbrandt, H.; Lind, C.; Sydow, O.; Olson, L.; Galter, D. Cerebellar alpha-synuclein levels are decreased in Parkinson’s disease and do not correlate with SNCA polymorphisms associated with disease in a Swedish material. FASEB J. 2008, 22, 3509–3514. [Google Scholar] [CrossRef]
- Lucchini, R.G.; Guazzetti, S.; Renzetti, S.; Broberg, K.; Caci, M.; Covolo, L.; Crippa, P.; Gelatti, U.; Hashim, D.; Oppini, M.; et al. Metal Exposure and SNCA rs356219 Polymorphism Associated with Parkinson Disease and Parkinsonism. Front. Neurol. 2020, 11, 556337. [Google Scholar] [CrossRef]
- Al-Chalabi, A.; Dürr, A.; Wood, N.W.; Parkinson, M.H.; Camuzat, A.; Hulot, J.S.; Morrison, K.E.; Renton, A.; Sussmuth, S.D.; Landwehrmeyer, B.G.; et al. Genetic variants of the alpha-synuclein gene SNCA are associated with multiple system atrophy. PLoS ONE 2009, 4, e7114. [Google Scholar] [CrossRef]
- Jo, S.; Park, K.W.; Hwang, Y.S.; Lee, S.H.; Ryu, H.S.; Chung, S.J. Microarray Genotyping Identifies New Loci Associated with Dementia in Parkinson’s Disease. Genes 2021, 12, 1975. [Google Scholar] [CrossRef]
- Chung, S.J.; Jung, Y.; Hong, M.; Kim, M.J.; You, S.; Kim, Y.J.; Kim, J.; Song, K. Alzheimer’s disease and Parkinson’s disease genome-wide association study top hits and risk of Parkinson’s disease in Korean population. Neurobiol. Aging 2013, 34, 2695.e1–2695.e7. [Google Scholar] [CrossRef]
- Chen, W.; Kang, W.Y.; Chen, S.; Wang, Y.; Xiao, Q.; Wang, G.; Liu, J.; Chen, S.D. Hyposmia correlates with SNCA variant and non-motor symptoms in Chinese patients with Parkinson’s disease. Park. Relat. Disord. 2015, 21, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, Y.; Chen, W.; Chen, S.; Wang, Y.; Xiao, Q.; Liu, J.; Fung, V.S.; Halliday, G.; Chen, S. Variants in the SNCA gene associate with motor progression while variants in the MAPT gene associate with the severity of Parkinson’s disease. Park. Relat. Disord. 2016, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Liang, D.; Pan, H.; Xu, Q.; Guo, J.; Sun, Q.; Tang, B.; Yan, X. Genetic Impact on Clinical Features in Parkinson’s Disease: A Study on SNCA -rs11931074. Park. Dis. 2018, 2018, 2754541. [Google Scholar] [CrossRef]
- Kang, W.; Chen, W.; Yang, Q.; Zhang, L.; Zhang, L.; Wang, X.; Dong, F.; Zhao, Y.; Chen, S.; Quinn, T.J.; et al. Salivary total α-synuclein, oligomeric α-synuclein and SNCA variants in Parkinson’s disease patients. Sci. Rep. 2016, 6, 28143. [Google Scholar] [CrossRef]
- Si, Q.Q.; Yuan, Y.S.; Zhi, Y.; Wang, M.; Wang, J.W.; Shen, Y.T.; Wang, L.N.; Li, J.Y.; Wang, X.X.; Zhang, K.Z. SNCA rs11931074 polymorphism correlates with spontaneous brain activity and motor symptoms in Chinese patients with Parkinson’s disease. J. Neural Transm. 2019, 126, 1037–1045. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, Q.; Wang, Y.; Xu, Z.M.; Wang, Y.; Yang, Q.; Wang, G.; Tan, Y.Y.; Ma, J.F.; Zhang, J.; et al. Analysis of genome-wide association study-linked loci in Parkinson’s disease of Mainland China. Mov. Disord. 2013, 28, 1892–1895. [Google Scholar] [CrossRef]
- Du, B.; Xue, Q.; Liang, C.; Fan, C.; Liang, M.; Zhang, Y.; Bi, X.; Hou, L. Association between alpha-synuclein (SNCA) rs11931074 variability and susceptibility to Parkinson’s disease: An updated meta-analysis of 41,811 patients. Neurol. Sci. 2020, 41, 271–280. [Google Scholar] [CrossRef]
- Chung, S.J.; König, I.R.; Lohmann, K.; Hinrichs, F.; Kim, J.; Ryu, H.S.; Lee, H.J.; Kim, K.; Lee, J.H.; Jung, K.W.; et al. Association of SNCA variants with α-synuclein of gastric and colonic mucosa in Parkinson’s disease. Park. Relat. Disord. 2019, 61, 151–155. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).