
Can Nature Overcome Invasive Gastrointestinal Infections?

 , , , , , and
, , , , , and
Abstract
1. Introduction
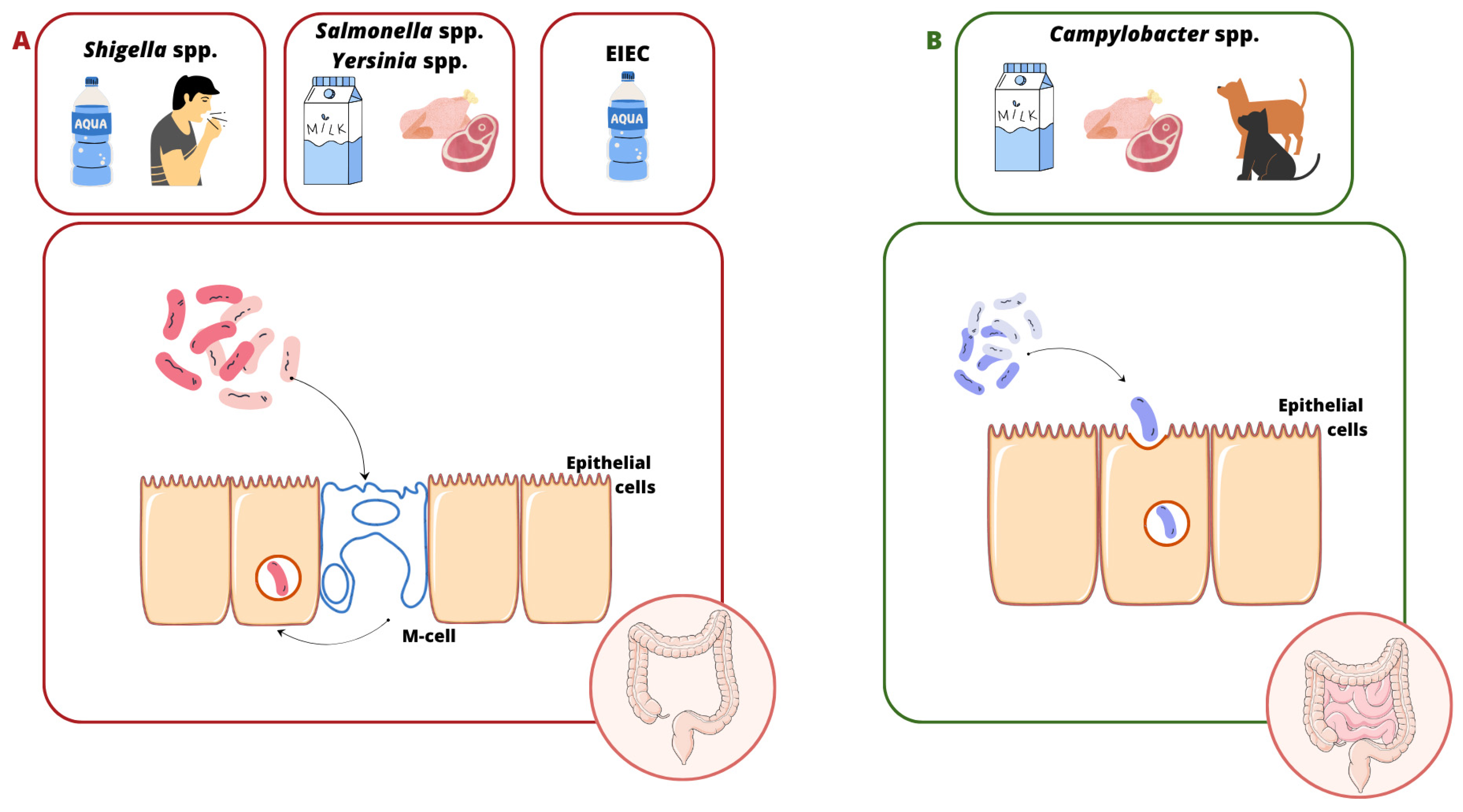
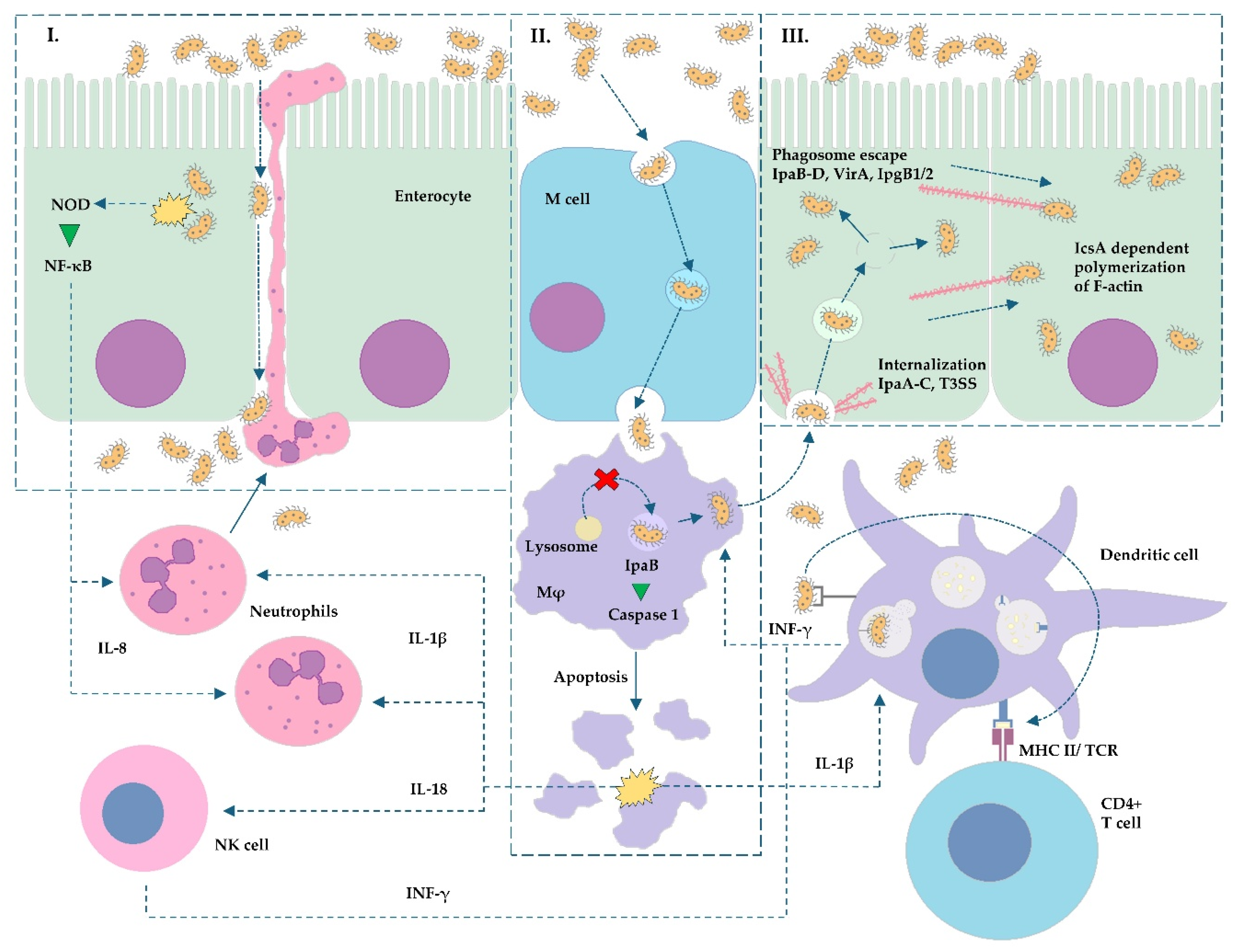
2. Molecular Basis of Enteroinvasive Pathogen Virulence
3. Berberine
3.1. Characteristics
3.2. Impact on Intestinal Mucosa
3.3. Impact on Gut Microbiome and Its Clinical Implications
3.4. Antimicrobial Properties Against Various Enteroinvasive Bacterial Pathogens
- Listeria monocytogenes
- Salmonella spp.
- Shigella spp.
- Campylobacter spp.
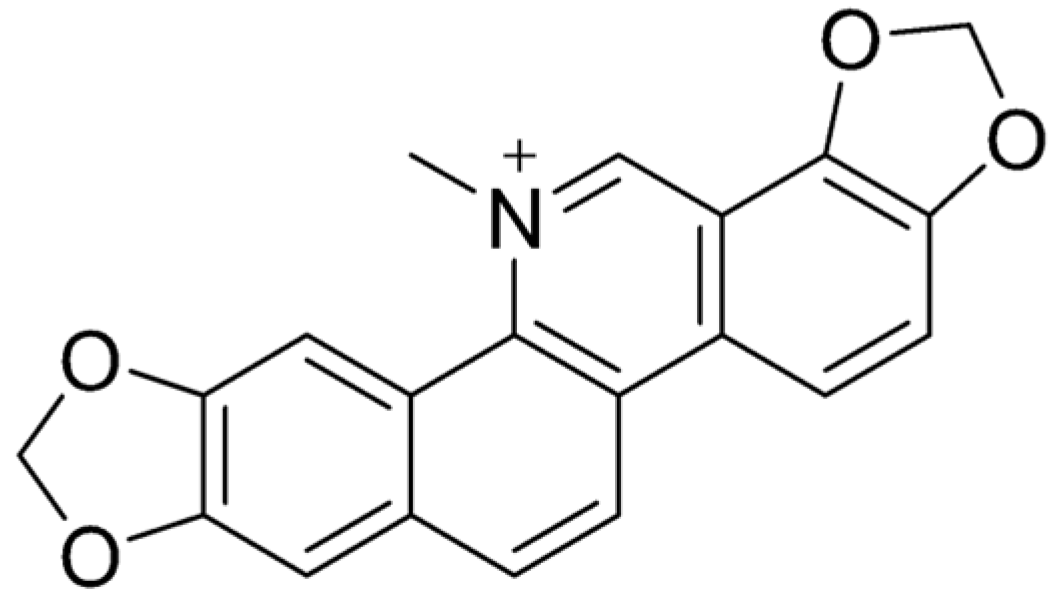
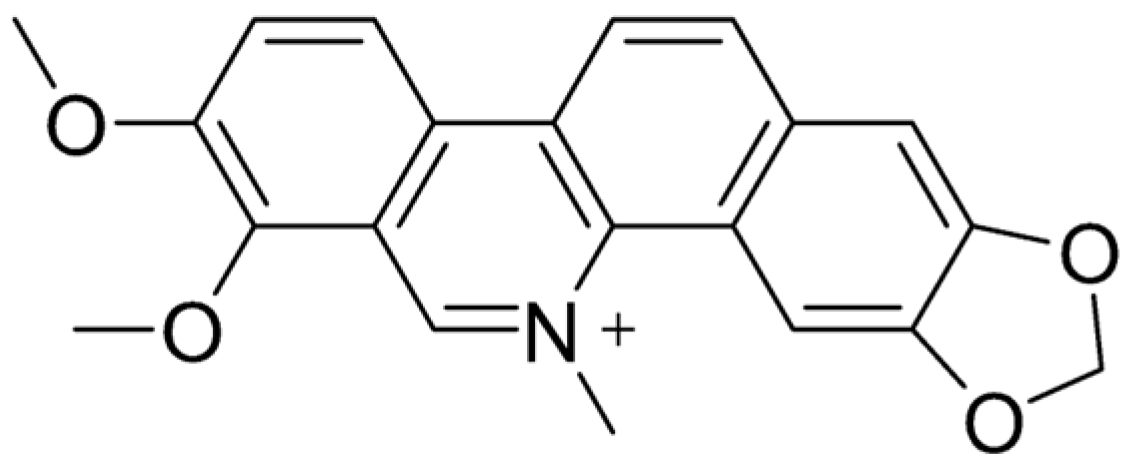
4. Sanguinarine
4.1. Characteristics
4.2. Impact on Intestinal Mucosa
4.3. Impact on Gut Microbiome and Its Clinical Implications
4.4. Antimicrobial Properties Against Various Enteroinvasive Bacterial Pathogens
- Listeria monocytogenes
- Salmonella spp.
- Shigella spp.
- Yersinia enterocolitica
5. Chelerythrine
5.1. Characteristics
5.2. Impact on Intestinal Mucosa
5.3. Impact on Gut Microbiome and Its Clinical Implications
5.4. Antimicrobial Properties Against Various Enteroinvasive Bacterial Pathogens
6. Overcoming Toxicity
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Available online: https://www.who.int/data/gho/data/themes/who-estimates-of-the-global-burden-of-foodborne-diseases (accessed on 15 March 2025).
- Carayol, N.; Van Nhieu, G.T. The Inside Story of Shigella Invasion of Intestinal Epithelial Cells. CSH Perspect. 2013, 3, a016717. [Google Scholar] [CrossRef] [PubMed]
- Marchello, C.S.; Birkhold, M.; Crump, J.A.; Martin, L.B.; Ansah, M.O.; Breghi, G.; Canals, R.; Fiorino, F.; Gordon, M.A.; Kim, J.H.; et al. Complications and Mortality of Non-Typhoidal Salmonella Invasive Disease: A Global Systematic Review and Meta-Analysis. Lancet Infect. Dis. 2022, 22, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Berthenet, E.; Thépault, A.; Chemaly, M.; Rivoal, K.; Ducournau, A.; Buissonnière, A.; Bénéjat, L.; Bessède, E.; Mégraud, F.; Sheppard, S.K.; et al. Source Attribution of Campylobacter Jejuni Shows Variable Importance of Chicken and Ruminants Reservoirs in Non-Invasive and Invasive French Clinical Isolates. Sci. Rep. 2019, 9, 8098. [Google Scholar] [CrossRef]
- Uliczka, F.; Pisano, F.; Schaake, J.; Stolz, T.; Rohde, M.; Fruth, A.; Strauch, E.; Skurnik, M.; Batzilla, J.; Rakin, A.; et al. Unique Cell Adhesion and Invasion Properties of Yersinia enterocolitica O:3, the Most Frequent Cause of Human Yersiniosis. PLoS Pathog. 2011, 7, e1002117. [Google Scholar] [CrossRef]
- Croxen, M.A.; Finlay, B.B. Molecular Mechanisms of Escherichia coli Pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Mallon, C.A.; Van Elsas, J.D.; Salles, J.F. Microbial Invasions: The Process, Patterns, and Mechanisms. Trends Microbiol. 2015, 23, 719–729. [Google Scholar] [CrossRef]
- Dekker, J.; Frank, K. Salmonella, Shigella, and Yersinia. Clin. Lab. Med. 2015, 35, 225. [Google Scholar] [CrossRef]
- Wang, J.; Ma, S.; Li, W.; Wang, X.; Huang, D.; Jiang, L.; Feng, L. Salmonella enterica Serovar Typhi Induces Host Metabolic Reprogramming to Increase Glucose Availability for Intracellular Replication. Int. J. Mol. Sci. 2021, 22, 10003. [Google Scholar] [CrossRef]
- Cossart, P.; Toledo-Arana, A. Listeria Monocytogenes, a Unique Model in Infection Biology: An Overview. Microbes Infect. 2008, 10, 1041–1050. [Google Scholar] [CrossRef]
- Konkel, M.E.; Talukdar, P.K.; Negretti, N.M.; Klappenbach, C.M. Taking Control: Campylobacter jejuni Binding to Fibronectin Sets the Stage for Cellular Adherence and Invasion. Front. Microbiol. 2020, 11, 564. [Google Scholar] [CrossRef]
- Kemper, L.; Hensel, A. Campylobacter Jejuni: Targeting Host Cells, Adhesion, Invasion, and Survival. Appl. Microbiol. Biotechnol. 2023, 107, 2725–2754. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, C.A.; Balbo, P.B.; Pesci, E.C.; Cottle, D.L.; Mirabito, P.M.; Pickett, C.L. Campylobacter jejuni Cytolethal Distending Toxin Causes a G2-Phase Cell Cycle Block. Infect. Immun. 1998, 66, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Asakura, M.; Samosornsuk, W.; Taguchi, M.; Kobayashi, K.; Misawa, N.; Kusumoto, M.; Nishimura, K.; Matsuhisa, A.; Yamasaki, S. Comparative Analysis of Cytolethal Distending Toxin (cdt) Genes among Campylobacter jejuni, C. Coli and C. Fetus Strains. Microb. Pathog. 2007, 42, 174–183. [Google Scholar] [CrossRef]
- Bonazzi, M.; Lecuit, M.; Cossart, P. Listeria monocytogenes Internalin and E-Cadherin: From Structure to Pathogenesis. Cell. Microbiol. 2009, 11, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L. Arthritis, Guillain-Barré Syndrome, and Other Sequelae of Campylobacter jejuni Enteritis. J. Food Prot. 1995, 58, 1153–1170. [Google Scholar] [CrossRef]
- Hannu, T.; Mattila, L.; Rautelin, H.; Pelkonen, P.; Lahdenne, P.; Siitonen, A.; Leirisalo-Repo, M. Campylobacter-Triggered Reactive Arthritis: A Population-Based Study. Rheumatology 2002, 41, 312–318. [Google Scholar] [CrossRef]
- Sylte, M.J.; Sivasankaran, S.K.; Trachsel, J.; Sato, Y.; Wu, Z.; Johnson, T.A.; Chandra, L.C.; Zhang, Q.; Looft, T. The Acute Host-Response of Turkeys Colonized With Campylobacter Coli. Front. Vet. Sci. 2021, 8, 613203. [Google Scholar] [CrossRef]
- Finsterer, J. Triggers of Guillain–Barré Syndrome: Campylobacter jejuni Predominates. Int. J. Mol. Sci. 2022, 23, 14222. [Google Scholar] [CrossRef]
- Shafiee, D.; Salpynov, Z.; Gusmanov, A.; Khuanbai, Y.; Mukhatayev, Z.; Kunz, J. Enteric Infection-Associated Reactive Arthritis: A Systematic Review and Meta-Analysis. J. Clin. Med. 2024, 13, 3433. [Google Scholar] [CrossRef]
- Butler, T. Haemolytic Uraemic Syndrome during Shigellosis. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 395–399. [Google Scholar] [CrossRef]
- Adams, C.; Vose, A.; Edmond, M.B.; Lyckholm, L. Shigella sonnei and Hemolytic Uremic Syndrome: A Case Report and Literature Review. IDCases 2017, 8, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.K.; Mehta, S.R.; Kumaravelu, S. Typhoid Fever Presenting as Acute Psychosis. Med. J. Armed Forces India 2003, 59, 252–253. [Google Scholar] [CrossRef] [PubMed]
- Chopra, P.; Bhatia, R.S.; Chopra, R. Mild Encephalopathy/Encephalitis with Reversible Splenial Lesion in a Patient with Salmonella typhi Infection: An Unusual Presentation with Excellent Prognosis. Indian J. Crit. Care Med. 2019, 23, 584–586. [Google Scholar] [CrossRef] [PubMed]
- Nwadike, V.U.; Fowotade, A.; Tuta, K.E.; Olusanya, O.O. A Rare Case Of Salmonella typhi Meningitis In An Eleven Month Old Infant: A Case Report. Ann. Ib. Postgrad. Med. 2012, 10, 38–39. [Google Scholar]
- Bottone, E.J. Yersinia enterocolitica: Overview and Epidemiologic Correlates. Microbes Infect. 1999, 1, 323–333. [Google Scholar] [CrossRef]
- Escoffier, P.; Gendrin, V.; Poloni, S.; Clerc, J.; Ducournau, A.; Klopfenstein, T.; Zayet, S. Disseminated Yersinia enterocolitica Infection Associated to Sweet’s Syndrome. BMC Infect. Dis. 2024, 24, 1036. [Google Scholar] [CrossRef]
- Pogreba-Brown, K.; Boyd, K.; Schaefer, K. Complications Associated with Foodborne Listeriosis: A Scoping Review. Foodborne Pathog. Dis. 2022, 19, 725–743. [Google Scholar] [CrossRef]
- Schnupf, P.; Sansonetti, P.J. Shigella Pathogenesis: New Insights through Advanced Methodologies. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Anderson, M.; Sansonetti, P.J.; Marteyn, B.S. Shigella Diversity and Changing Landscape: Insights for the Twenty-First Century. Front. Cell. Infect. Microbiol. 2016, 6, 45. [Google Scholar] [CrossRef]
- Nasser, A.; Mosadegh, M.; Azimi, T.; Shariati, A. Molecular Mechanisms of Shigella Effector Proteins: A Common Pathogen among Diarrheic Pediatric Population. Mol. Cell. Pediatr. 2022, 9, 12. [Google Scholar] [CrossRef]
- Parsot, C. Shigella Spp. and Enteroinvasive Escherichia coli Pathogenicity Factors. FEMS Microbiol. Lett. 2005, 252, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Muthuramalingam, M.; Whittier, S.K.; Picking, W.L.; Picking, W.D. The Shigella Type III Secretion System: An Overview from Top to Bottom. Microorganisms 2021, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Dillon, A.; Lo, D.D. M Cells: Intelligent Engineering of Mucosal Immune Surveillance. Front. Immunol. 2019, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Pilla, G.; McVicker, G.; Tang, C.M. Genetic Plasticity of the Shigella Virulence Plasmid Is Mediated by Intra-and Inter-Molecular Events between Insertion Sequences. PLoS Genet. 2017, 13, e1007014. [Google Scholar] [CrossRef]
- Agaisse, H. Molecular and Cellular Mechanisms of Shigella flexneri Dissemination. Front. Cell. Infect. Microbiol. 2016, 6, 29. [Google Scholar] [CrossRef]
- Ashida, H.; Suzuki, T.; Sasakawa, C. Shigella Infection and Host Cell Death: A Double-Edged Sword for the Host and Pathogen Survival. Curr. Opin. Microbiol. 2021, 59, 1–7. [Google Scholar] [CrossRef]
- Handa, Y.; Suzuki, M.; Ohya, K.; Iwai, H.; Ishijima, N.; Koleske, A.J.; Fukui, Y.; Sasakawa, C. Shigella IpgB1 Promotes Bacterial Entry through the ELMO–Dock180 Machinery. Nat. Cell Biol. 2007, 9, 121–128. [Google Scholar] [CrossRef]
- Alto, N.M.; Dixon, J.E. Analysis of Rho-GTPase Mimicry by a Family of Bacterial Type III Effector Proteins. Methods Enzymol. 2008, 439, 131–143. [Google Scholar]
- Weddle, E.A.; Köseoğlu, V.K.; DeVasure, B.A.; Agaisse, H.F. The Type Three Secretion System Effector Protein IpgB1 Promotes Shigella flexneri Cell-to-Cell Spread through Double-Membrane Vacuole Escape. PLoS Pathog. 2022, 18, e1010380. [Google Scholar] [CrossRef]
- Yang, S.-C.; Hung, C.-F.; Aljuffali, I.A.; Fang, J.-Y. The Roles of the Virulence Factor IpaB in Shigella Spp. in the Escape from Immune Cells and Invasion of Epithelial Cells. Microbiol. Res. 2015, 181, 43–51. [Google Scholar] [CrossRef]
- Wing, H.J.; Yan, A.W.; Goldman, S.R.; Goldberg, M.B. Regulation of IcsP, the Outer Membrane Protease of the Shigella Actin Tail Assembly Protein IcsA, by Virulence Plasmid Regulators VirF and VirB. J. Bacteriol. 2004, 186, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.B.; Theriot, J.A. Shigella flexneri Surface Protein IcsA Is Sufficient to Direct Actin-Based Motility. Proc. Natl. Acad. Sci. USA 1995, 92, 6572–6576. [Google Scholar] [CrossRef]
- Fukazawa, A.; Alonso, C.; Kurachi, K.; Gupta, S.; Lesser, C.F.; McCormick, B.A.; Reinecker, H.-C. GEF-H1 Mediated Control of NOD1 Dependent NF-KB Activation by Shigella Effectors. PLoS Pathog. 2008, 4, e1000228. [Google Scholar] [CrossRef]
- Singer, M.; Sansonetti, P.J. IL-8 Is a Key Chemokine Regulating Neutrophil Recruitment in a New Mouse Model of Shigella-Induced Colitis. J. Immunol. 2004, 173, 4197–4206. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Römer, W. Shiga Toxins—From Cell Biology to Biomedical Applications. Nat. Rev. Microbiol. 2010, 8, 105–116. [Google Scholar] [CrossRef]
- van den Beld, M.; Reubsaet, F.A.G. Differentiation between Shigella, Enteroinvasive Escherichia coli (EIEC) and Noninvasive Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 899–904. [Google Scholar] [CrossRef]
- Lan, R.; Alles, M.C.; Donohoe, K.; Martinez, M.B.; Reeves, P.R. Molecular Evolutionary Relationships of Enteroinvasive Escherichia coli and Shigella Spp. Infect. Immun. 2004, 72, 5080–5088. [Google Scholar]
- Ud-Din, A.; Wahid, S. Relationship among Shigella Spp. and Enteroinvasive Escherichia coli (EIEC) and Their Differentiation. Braz. J. Microbiol. 2014, 45, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.C.R.; Gonçalves Ferreira, L.; Baquerizo Martinez, M. Enteroinvasive Escherichia coli vs. Shigella flexneri: How Different Patterns of Gene Expression Affect Virulence. FEMS Microbiol. Lett. 2009, 301, 156–163. [Google Scholar]
- Navarro-Garcia, F.; Serapio-Palacios, A.; Ugalde-Silva, P.; Tapia-Pastrana, G.; Chavez-Dueñas, L. Actin Cytoskeleton Manipulation by Effector Proteins Secreted by Diarrheagenic Escherichia coli Pathotypes. Biomed Res. Int. 2013, 2013, 374395. [Google Scholar] [CrossRef]
- Belotserkovsky, I.; Sansonetti, P.J. Shigella and Enteroinvasive Escherichia coli. Escherichia Coli A Versatile Pathog. 2018, 416, 1–26. [Google Scholar]
- Tsolis, R.M.; Kingsley, R.A.; Townsend, S.M.; Ficht, T.A.; Adams, L.G.; Bäumler, A.J. Of Mice, Calves, and Men: Comparison of the Mouse Typhoid Model with Other Salmonella Infections. Mech. Pathog. Enteric Dis. 2 1999, 473, 261–274. [Google Scholar]
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the Host and Disease: A Brief Review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.D.; Ghori, N.; Falkow, S. Salmonella typhimurium Initiates Murine Infection by Penetrating and Destroying the Specialized Epithelial M Cells of the Peyer’s Patches. J. Exp. Med. 1994, 180, 15–23. [Google Scholar] [CrossRef]
- Waterman, S.R.; Holden, D.W. Functions and Effectors of the Salmonella Pathogenicity Island 2 Type III Secretion System. Cell. Microbiol. 2003, 5, 501–511. [Google Scholar] [CrossRef]
- Martinez-Argudo, I.; Jepson, M.A. Salmonella Translocates across an in Vitro M Cell Model Independently of SPI-1 and SPI-2. Microbiology 2008, 154, 3887–3894. [Google Scholar] [CrossRef]
- Jepson, M.A.; Clark, M.A. The Role of M Cells in Salmonella Infection. Microbes Infect. 2001, 3, 1183–1190. [Google Scholar] [CrossRef]
- Zhang, K.; Riba, A.; Nietschke, M.; Torow, N.; Repnik, U.; Pütz, A.; Fulde, M.; Dupont, A.; Hensel, M.; Hornef, M. Minimal SPI1-T3SS Effector Requirement for Salmonella Enterocyte Invasion and Intracellular Proliferation in Vivo. PLoS Pathog. 2018, 14, e1006925. [Google Scholar] [CrossRef]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive Virulence Factors and Pathogenesis. Cell. Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef]
- Ibarra, J.A.; Steele-Mortimer, O. Salmonella–the Ultimate Insider. Salmonella Virulence Factors That Modulate Intracellular Survival. Cell. Microbiol. 2009, 11, 1579–1586. [Google Scholar]
- Hegazy, W.A.H.; Abbas, H.A. Evaluation of the Role of SsaV Salmonella Pathogenicity Island-2 Dependent Type III Secretion System Components on the Virulence Behavior of Salmonella enterica Serovar Typhimurium. Afr. J. Biotechnol. 2017, 16, 718–726. [Google Scholar]
- Beuzón, C.R.; Banks, G.; Deiwick, J.; Hensel, M.; Holden, D.W. PH-dependent Secretion of SseB, a Product of the SPI-2 Type III Secretion System of Salmonella typhimurium. Mol. Microbiol. 1999, 33, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ruiz-Albert, J.; Unsworth, K.E.; Garvis, S.; Liu, M.; Holden, D.W. SpiC Is Required for Secretion of Salmonella Pathogenicity Island 2 Type III Secretion System Proteins. Cell. Microbiol. 2002, 4, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.L.; Cookson, B.T. Pyroptosis and Host Cell Death Responses during Salmonella Infection. Cell. Microbiol. 2007, 9, 2562–2570. [Google Scholar] [CrossRef]
- Monack, D.M.; Raupach, B.; Hromockyj, A.E.; Falkow, S. Salmonella typhimurium Invasion Induces Apoptosis in Infected Macrophages. Proc. Natl. Acad. Sci. USA 1996, 93, 9833–9838. [Google Scholar] [CrossRef]
- Cornelis, G.R. Yersinia Pathogenicity Factors. In Bacterial Pathogenesis of Plants and Animals: Molecular and Cellular Mechanisms; Dangl, J.L., Ed.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 243–263. ISBN 978-3-642-78624-2. [Google Scholar]
- Hammerl, J.A.; Klein, I.; Lanka, E.; Appel, B.; Hertwig, S. Genetic and Functional Properties of the Self-Transmissible Yersinia enterocolitica Plasmid PYE854, Which Mobilizes the Virulence Plasmid PYV. J. Bacteriol. 2008, 190, 991–1010. [Google Scholar] [CrossRef]
- Clark, M.A.; Hirst, B.H.; Jepson, M.A. M-Cell Surface Β1 Integrin Expression and Invasin-Mediated Targeting of Yersinia Pseudotuberculosis to Mouse Peyer’s Patch M Cells. Infect. Immun. 1998, 66, 1237–1243. [Google Scholar] [CrossRef]
- Isberg, R.R.; Hamburger, Z.; Dersch, P. Signaling and Invasin-Promoted Uptake via Integrin Receptors. Microbes Infect. 2000, 2, 793–801. [Google Scholar] [CrossRef]
- Trosky, J.E.; Liverman, A.D.B.; Orth, K. Yersinia Outer Proteins: Yops. Cell. Microbiol. 2008, 10, 557–565. [Google Scholar] [CrossRef]
- Dersch, P. A Region of the Yersinia Pseudotuberculosis Invasin Protein Enhances Integrin-Mediated Uptake into Mammalian Cells and Promotes Self-Association. EMBO J. 1999, 18, 1199–1213. [Google Scholar] [CrossRef]
- Grassl, G.A.; Bohn, E.; Müller, Y.; Bühler, O.T.; Autenrieth, I.B. Interaction of Yersinia enterocolitica with Epithelial Cells: Invasin beyond Invasion. Int. J. Med. Microbiol. 2003, 293, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Tertti, R.; Skurnik, M.; Vartio, T.; Kuusela, P. Adhesion Protein YadA of Yersinia Species Mediates Binding of Bacteria to Fibronectin. Infect. Immun. 1992, 60, 3021–3024. [Google Scholar] [CrossRef]
- Flügel, A.; Schulze-Koops, H.; Heesemann, J.; Kühn, K.; Sorokin, L.; Burkhardt, H.; von der Mark, K.; Emmrich, F. Interaction of Enteropathogenic Yersinia enterocolitica with Complex Basement Membranes and the Extracellular Matrix Proteins Collagen Type IV, Laminin-1 and-2, and Nidogen/Entactin. J. Biol. Chem. 1994, 269, 29732–29738. [Google Scholar] [CrossRef]
- Eitel, J.; Dersch, P. The YadA Protein of Yersinia Pseudotuberculosis Mediates High-Efficiency Uptake into Human Cells under Environmental Conditions in Which Invasin Is Repressed. Infect. Immun. 2002, 70, 4880–4891. [Google Scholar] [CrossRef]
- China, B.; N’guyen, B.T.; De Bruyère, M.; Cornelis, G.R. Role of YadA in Resistance of Yersinia enterocolitica to Phagocytosis by Human Polymorphonuclear Leukocytes. Infect. Immun. 1994, 62, 1275–1281. [Google Scholar] [CrossRef]
- Ruckdeschel, K.; Roggenkamp, A.; Schubert, S.; Heesemann, J. Differential Contribution of Yersinia enterocolitica Virulence Factors to Evasion of Microbicidal Action of Neutrophils. Infect. Immun. 1996, 64, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, P.K.; Negretti, N.M.; Turner, K.L.; Konkel, M.E. Molecular Dissection of the Campylobacter jejuni CadF and FlpA Virulence Proteins in Binding to Host Cell Fibronectin. Microorganisms 2020, 8, 389. [Google Scholar] [CrossRef]
- Monteville, M.R. Maximal Adherence and Invasion of INT 407 Cells by Campylobacter jejuni Requires the CadF Outer-Membrane Protein and Microfilament Reorganization. Microbiology 2003, 149, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Song, Y.C.; Emili, A.; Sherman, P.M.; Chan, V.L. JlpA of Campylobacter jejuni Interacts with Surface-Exposed Heat Shock Protein 90alpha and Triggers Signalling Pathways Leading to the Activation of NF-KappaB and P38 MAP Kinase in Epithelial Cells. Cell. Microbiol. 2003, 5, 165–174. [Google Scholar] [CrossRef]
- Bæk, K.T.; Vegge, C.S.; Brøndsted, L. HtrA Chaperone Activity Contributes to Host Cell Binding in Campylobacter jejuni. Gut Pathog. 2011, 3, 13. [Google Scholar] [CrossRef]
- Monteville, M.R.; Konkel, M.E. Fibronectin-Facilitated Invasion of T84 Eukaryotic Cells by Campylobacter jejuni Occurs Preferentially at the Basolateral Cell Surface. Infect. Immun. 2002, 70, 6665–6671. [Google Scholar] [CrossRef]
- Harrer, A.; Bücker, R.; Boehm, M.; Zarzecka, U.; Tegtmeyer, N.; Sticht, H.; Schulzke, J.D.; Backert, S. Campylobacter jejuni Enters Gut Epithelial Cells and Impairs Intestinal Barrier Function through Cleavage of Occludin by Serine Protease HtrA. Gut Pathog. 2019, 11, 4. [Google Scholar] [CrossRef]
- Klappenbach, C.M.; Negretti, N.M.; Aaron, J.; Chew, T.-L.; Konkel, M.E. Campylobacter jejuni Triggers Signaling through Host Cell Focal Adhesions To Inhibit Cell Motility. MBio 2021, 12, e0149421. [Google Scholar] [CrossRef] [PubMed]
- Larson, C.L.; Samuelson, D.R.; Eucker, T.P.; O’Loughlin, J.L.; Konkel, M.E. The Fibronectin-Binding Motif within FlpA Facilitates Campylobacter jejuni Adherence to Host Cell and Activation of Host Cell Signaling. Emerg. Microbes Infect. 2013, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eucker, T.P.; Konkel, M.E. The Cooperative Action of Bacterial Fibronectin-binding Proteins and Secreted Proteins Promote Maximal Campylobacter jejuni Invasion of Host Cells by Stimulating Membrane Ruffling. Cell. Microbiol. 2012, 14, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Buelow, D.R.; Christensen, J.E.; Neal-McKinney, J.M.; Konkel, M.E. Campylobacter jejuni Survival within Human Epithelial Cells Is Enhanced by the Secreted Protein CiaI. Mol. Microbiol. 2011, 80, 1296–1312. [Google Scholar] [CrossRef]
- Ó Cróinín, T.; Backert, S. Host Epithelial Cell Invasion by Campylobacter jejuni: Trigger or Zipper Mechanism? Front. Cell. Infect. Microbiol. 2012, 2, 25. [Google Scholar] [CrossRef]
- Cossart, P.; Cerda, J.P. Bacterial Adhesion and Entry into Host Cells. Cell 2006, 124, 715–727. [Google Scholar]
- Shen, Y.; Naujokas, M.; Park, M.; Ireton, K. InlB-Dependent Internalization of Listeria Is Mediated by the Met Receptor Tyrosine Kinase. Cell 2000, 103, 501–510. [Google Scholar] [CrossRef]
- Lecuit, M.; Vandormael-Pournin, S.; Lefort, J.; Huerre, M.; Gounon, P.; Dupuy, C.; Babinet, C.; Cossart, P. A Transgenic Model for Listeriosis: Role of Internalin in Crossing the Intestinal Barrier. Science 2001, 292, 1722–1725. [Google Scholar] [CrossRef]
- Bergmann, B.; Raffelsbauer, D.; Kuhn, M.; Goetz, M.; Hom, S.; Goebel, W. InlA- but Not InlB-Mediated Internalization of Listeria monocytogenes by Non-Phagocytic Mammalian Cells Needs the Support of Other Internalins. Mol. Microbiol. 2002, 43, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Mengaud, J.; Ohayon, H.; Gounon, P.; Mège, R.-M.; Cossart, P. E-Cadherin Is the Receptor for Internalin, a Surface Protein Required for Entry of L. Monocytogenes into Epithelial Cells. Cell 1996, 84, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Bonazzi, M.; Veiga, E.; Pizarro-Cerdá, J.; Cossart, P. Successive Post-translational Modifications of E-cadherin Are Required for InlA-mediated Internalization of Listeria Monocytogenes. Cell. Microbiol. 2008, 10, 2208–2222. [Google Scholar] [CrossRef]
- Veiga, E.; Cossart, P. Listeria Hijacks the Clathrin-Dependent Endocytic Machinery to Invade Mammalian Cells. Nat. Cell Biol. 2005, 7, 894–900. [Google Scholar] [CrossRef]
- Veiga, E.; Cossart, P. The Role of Clathrin-Dependent Endocytosis in Bacterial Internalization. Trends Cell Biol. 2006, 16, 499–504. [Google Scholar] [CrossRef]
- Ruan, Y.; Rezelj, S.; Bedina Zavec, A.; Anderluh, G.; Scheuring, S. Listeriolysin O Membrane Damaging Activity Involves Arc Formation and Lineaction—Implication for Listeria monocytogenes Escape from Phagocytic Vacuole. PLoS Pathog. 2016, 12, e1005597. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.N.; Peterson, B.N.; Portnoy, D.A. Listeriolysin O: A Phagosome-Specific Cytolysin Revisited. Cell. Microbiol. 2019, 21, e12988. [Google Scholar] [CrossRef]
- Alvarez-Dominguez, C.; Stahl, P.D. Increased Expression of Rab5a Correlates Directly with Accelerated Maturation of Listeria monocytogenes Phagosomes. J. Biol. Chem. 1999, 274, 11459–11462. [Google Scholar] [CrossRef]
- Xayarath, B.; Alonzo, F.; Freitag, N.E. Identification of a Peptide-Pheromone That Enhances Listeria monocytogenes Escape from Host Cell Vacuoles. PLoS Pathog. 2015, 11, e1004707. [Google Scholar] [CrossRef]
- Singh, R.; Jamieson, A.; Cresswell, P. GILT Is a Critical Host Factor for Listeria monocytogenes Infection. Nature 2008, 455, 1244–1247. [Google Scholar] [CrossRef]
- Travier, L.; Guadagnini, S.; Gouin, E.; Dufour, A.; Chenal-Francisque, V.; Cossart, P.; Olivo-Marin, J.C.; Ghigo, J.M.; Disson, O.; Lecuit, M. ActA Promotes Listeria monocytogenes Aggregation, Intestinal Colonization and Carriage. PLoS Pathog. 2013, 9, e1003131. [Google Scholar] [CrossRef] [PubMed]
- Kocks, C.; Gouin, E.; Tabouret, M.; Berche, P.; Ohayon, H.; Cossart, P.L. Monocytogenes-Induced Actin Assembly Requires the actA Gene Product, a Surface Protein. Cell 1992, 68, 521–531. [Google Scholar] [CrossRef]
- Lecuit, M. Human Listeriosis and Animal Models. Microbes Infect. 2007, 9, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, B.; Guilhabert, M.R.; Miller, W.G.; Mandrell, R.E.; Lastovica, A.J.; Parker, C.T. Comparative Genomic Analysis of Clinical Strains of Campylobacter jejuni from South Africa. PLoS ONE 2008, 3, e2015. [Google Scholar] [CrossRef]
- Yue, S.J.; Liu, J.; Wang, W.X.; Wang, A.T.; Yang, X.Y.; Guan, H.S.; Wang, C.Y.; Yan, D. Berberine Treatment-Emergent Mild Diarrhea Associated with Gut Microbiota Dysbiosis. Biomed. Pharmacother. 2019, 116, 109002. [Google Scholar] [CrossRef] [PubMed]
- Jael Teresa de Jesús, Q.V.; Gálvez-Ruíz, J.C.; Márquez Ibarra, A.A.; Leyva-Peralta, M.A. Perspectives on Berberine and the Regulation of Gut Microbiota: As an Anti-Inflammatory Agent. Pharmaceuticals 2025, 18, 193. [Google Scholar] [CrossRef]
- Li, G.X.; Wang, X.M.; Jiang, T.; Gong, J.F.; Niu, L.Y.; Li, N. Berberine Prevents Intestinal Mucosal Barrier Damage during Early Phase of Sepsis in Rat through the Toll-like Receptors Signaling Pathway. Korean J. Physiol. Pharmacol. 2015, 19, 1–7. [Google Scholar] [CrossRef]
- Yang, F.; Gao, R.; Luo, X.; Liu, R.; Xiong, D. Berberine Influences Multiple Diseases by Modifying Gut Microbiota. Front. Nutr. 2023, 10, 1187718. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, X.; Yang, R.; Chen, F.; Liao, Y.; Zhu, Z.; Wu, Z.; Sun, X.; Wang, L. Effects of Berberine on the Gastrointestinal Microbiota. Front. Cell. Infect. Microbiol. 2021, 10, 588517. [Google Scholar] [CrossRef]
- Wolf, P.G.; Devendran, S.; Doden, H.L.; Ly, L.K.; Moore, T.; Takei, H.; Nittono, H.; Murai, T.; Kurosawa, T.; Chlipala, G.E.; et al. Berberine Alters Gut Microbial Function through Modulation of Bile Acids. BMC Microbiol. 2021, 21, 24. [Google Scholar] [CrossRef]
- He, Y.; Yuan, X.; Zuo, H.; Sun, Y.; Feng, A. Berberine Exerts a Protective Effect on Gut-Vascular Barrier via the Modulation of the Wnt/Beta-Catenin Signaling Pathway during Sepsis. Cell. Physiol. Biochem. 2018, 49, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, X.; Zhang, M.; Zou, Z.; Yang, S.; Yi, T.; Wang, J.; Liu, D.; Shen, Y.; Dai, C.; et al. Dual Effects of Feed-Additive-Derived Chelerythrine in Combating Mobile Colistin Resistance. Engineering 2024, 32, 163–173. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Hu, D.; Zhang, Y.L.; Zou, J.; Chen, G.L.; Guo, M.Q. Inhibitors Targeting Multiple Janus Kinases From Zanthoxylum simulans Mediate Inhibition and Apoptosis Against Gastric Cancer Cells via the Estrogen Pathway. Front. Chem. 2022, 10, 922110. [Google Scholar] [CrossRef]
- Xin, J.; Pu, Q.; Wang, R.; Gu, Y.; He, L.; Du, X.; Tang, G.; Han, D. Antibacterial Activity and Mechanism of Chelerythrine against Streptococcus agalactiae. Front. Vet. Sci. 2024, 11, 1408376. [Google Scholar] [CrossRef] [PubMed]
- Sukhotnik, I.; Bitterman, S.; Shahar, Y.B.; Pollak, Y.; Bitterman, N.; Halabi, S.; Coran, A.G.; Bitterman, A. Effect of Chelerythrine on Intestinal Cell Turnover Following Intestinal Ischemia-Reperfusion Injury in a Rat Model. Eur. J. Pediatr. Surg. 2017, 27, 36–43. [Google Scholar] [CrossRef]
- Tavares, L.d.C.; Zanon, G.; Weber, A.D.; Neto, A.T.; Mostardeiro, C.P.; Da Cruz, I.B.M.; Oliveira, R.M.; Ilha, V.; Dalcol, I.I.; Morel, A.F. Structure-Activity Relationship of Benzophenanthridine Alkaloids from Zanthoxylum rhoifolium Having Antimicrobial Activity. PLoS ONE 2014, 9, e97000. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Wang, T.; Deng, X.; Chu, X. Natural Compound Sanguinarine Chloride Targets the Type III Secretion System of Salmonella enterica Serovar Typhimurium. Biochem. Biophys. Rep. 2018, 14, 149–154. [Google Scholar] [CrossRef]
- Song, D.; Hao, J.; Fan, D. Biological Properties and Clinical Applications of Berberine. Front. Med. 2020, 14, 564–582. [Google Scholar] [CrossRef]
- Utami, A.R.; Maksum, I.P.; Deawati, Y. Berberine and Its Study as an Antidiabetic Compound. Biology 2023, 12, 973. [Google Scholar] [CrossRef]
- Xu, F.; Liu, M.; Liao, Y.; Zhou, Y.; Zhang, P.; Zeng, Y.; Liu, Z. Improvement of Anticancer Effect of Berberine by Salt Formation Modifications. Phytomedicine 2022, 104, 154314. [Google Scholar] [CrossRef]
- Kuo, C.L.; Chi, C.W.; Liu, T.Y. The Anti-Inflammatory Potential of Berberine in Vitro and in Vivo. Cancer Lett. 2004, 203, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Čerňáková, M.; Košťálová, D. Antimicrobial Activity of Berberine—A Constituent of Mahonia Aquifolium. Folia Microbiol. 2002, 47, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Nechepurenko, I.V.; Boyarskikh, U.A.; Khvostov, M.V.; Baev, D.S.; Komarova, N.I.; Filipenko, M.L.; Tolstikova, T.G.; Salakhutdinov, N.F. Hypolipidemic Berberine Derivatives with a Reduced Aromatic Ring C. Chem. Nat. Compd. 2015, 51, 916–922. [Google Scholar] [CrossRef]
- Sunhe, Y.-X.; Zhang, Y.-H.; Fu, R.-J.; Xu, D.-Q.; Tang, Y.-P. Neuroprotective Effect and Preparation Methods of Berberine. Front. Pharmacol. 2024, 15, 1429050. [Google Scholar] [CrossRef]
- Khoshandam, A.; Imenshahidi, M.; Hosseinzadeh, H. Pharmacokinetic of Berberine, the Main Constituent of Berberis Vulgaris L.: A Comprehensive Review. Phyther. Res. 2022, 36, 4063–4079. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, Y.; Ren, H.; Wang, S.; Zhong, H.; Zhao, X.; Ma, J.; Gu, X.; Xue, Y.; Huang, S.; et al. Gut Microbiome-Related Effects of Berberine and Probiotics on Type 2 Diabetes (the PREMOTE Study). Nat. Commun. 2020, 11, 5015. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Lu, M.; Pan, Q.; Fichna, J.; Zheng, L.; Wang, K.; Yu, Z.; Li, Y.; Li, K.; Song, A.; et al. Berberine Improves Intestinal Motility and Visceral Pain in the Mouse Models Mimicking Diarrhea-Predominant Irritable Bowel Syndrome (IBS-D) Symptoms in an Opioid-Receptor Dependent Manner. PLoS ONE 2015, 10, e0145556. [Google Scholar] [CrossRef]
- Liao, Z.; Xie, Y.; Zhou, B.; Zou, B.; Xiao, D.; Liu, W.; Cai, Y.; Liu, D.; Liao, Q.; Xie, Z. Berberine Ameliorates Colonic Damage Accompanied with the Modulation of Dysfunctional Bacteria and Functions in Ulcerative Colitis Rats. Appl. Microbiol. Biotechnol. 2020, 104, 1737–1749. [Google Scholar] [CrossRef]
- Dong, Y.; Fan, H.; Zhang, Z.; Jiang, F.; Li, M.; Zhou, H.; Guo, W.; Zhang, Z.; Kang, Z.; Gui, Y.; et al. Berberine Ameliorates DSS-Induced Intestinal Mucosal Barrier Dysfunction through Microbiota-Dependence and Wnt/β-Catenin Pathway. Int. J. Biol. Sci. 2022, 18, 1381. [Google Scholar] [CrossRef]
- Li, H.; Fan, C.; Lu, H.; Feng, C.; He, P.; Yang, X.; Xiang, C.; Zuo, J.; Tang, W. Protective Role of Berberine on Ulcerative Colitis through Modulating Enteric Glial Cells–Intestinal Epithelial Cells–Immune Cells Interactions. Acta Pharm. Sin. B 2020, 10, 447. [Google Scholar] [CrossRef]
- Tu, S.; Huang, Y.; Tian, H.; Xu, L.; Wang, X.; Huang, L.; Lei, X.; Xu, Z.; Liu, D. Berberine Enhances the Function of Intestinal Stem Cells in Healthy and Radiation-Injured Mice. Int. Immunopharmacol. 2024, 136, 112278. [Google Scholar] [CrossRef] [PubMed]
- Progatzky, F.; Pachnis, V. The Role of Enteric Glia in Intestinal Immunity. Curr. Opin. Immunol. 2022, 77, 102183. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.J.; Pothoulakis, C. Role of Neuropeptides in Inflammatory Bowel Disease. BASIC Sci. Rev. 2007, 13, 918–932. [Google Scholar] [CrossRef]
- López-Gómez, L.; Szymaszkiewicz, A.; Zielińska, M.; Abalo, R. Nutraceuticals and Enteric Glial Cells. Molecules 2021, 26, 3762. [Google Scholar] [CrossRef]
- Jing, W.; Dong, S.; Luo, X.; Liu, J.; Wei, B.; Du, W.; Yang, L.; Luo, H.; Wang, Y.; Wang, S.; et al. Berberine Improves Colitis by Triggering AhR Activation by Microbial Tryptophan Catabolites. Pharmacol. Res. 2021, 164, 105358. [Google Scholar] [CrossRef]
- Zou, K.; Li, Z.; Zhang, Y.; Zhang, H.Y.; Li, B.; Zhu, W.L.; Shi, J.Y.; Jia, Q.; Li, Y.M. Advances in the Study of Berberine and Its Derivatives: A Focus on Anti-Inflammatory and Anti-Tumor Effects in the Digestive System. Acta Pharmacol. Sin. 2017, 38, 157. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhao, L.; Yuan, X.; Li, Y.; Shi, J.; Zhang, H.; Zhao, Y.; Han, L.; Wang, H.; Yan, Y.; et al. Pre-Administration of Berberine Exerts Chemopreventive Effects in AOM/DSS-Induced Colitis-Associated Carcinogenesis Mice via Modulating Inflammation and Intestinal Microbiota. Nutrients 2022, 14, 726. [Google Scholar] [CrossRef]
- Zheng, C.; Wang, Y.; Xu, Y.; Zhou, L.; Hassan, S.; Xu, G.; Zou, X.; Zhang, M. Berberine Inhibits Dendritic Cells Differentiation in DSS-Induced Colitis by Promoting Bacteroides Fragilis. Int. Immunopharmacol. 2021, 101, 108329. [Google Scholar] [CrossRef]
- Li, C.; Cao, H.; Huan, Y.; Ji, W.; Liu, S.; Sun, S.; Liu, Q.; Lei, L.; Liu, M.; Gao, X.; et al. Berberine Combined with Stachyose Improves Glycometabolism and Gut Microbiota through Regulating Colonic MicroRNA and Gene Expression in Diabetic Rats. Life Sci. 2021, 284, 119928. [Google Scholar] [CrossRef]
- Li, M.; Zhou, W.; Dang, Y.; Li, C.; Ji, G.; Zhang, L. Berberine Compounds Improves Hyperglycemia via Microbiome Mediated Colonic TGR5-GLP Pathway in Db/Db Mice. Biomed. Pharmacother. 2020, 132, 110953. [Google Scholar] [CrossRef]
- Xu, X.; Gao, Z.; Yang, F.; Yang, Y.; Chen, L.; Han, L.; Zhao, N.; Xu, J.; Wang, X.; Ma, Y.; et al. Antidiabetic Effects of Gegen Qinlian Decoction via the Gut Microbiota Are Attributable to Its Key Ingredient Berberine. Genom. Proteom. Bioinform. 2020, 18, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Chen, H.; Yan, L.; Wang, W.; Wang, D. Berberine Alleviates Type 2 Diabetic Symptoms by Altering Gut Microbiota and Reducing Aromatic Amino Acids. Biomed. Pharmacother. 2020, 131, 110669. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J.H.; Yu, T.; Chen, Q.K. Effects of Berberine and Metformin on Intestinal Inflammation and Gut Microbiome Composition in Db/Db Mice. Biomed. Pharmacother. 2019, 118, 109131. [Google Scholar] [CrossRef]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Buda, A.; Romualdi, C.; Pohl, D.; Perini, B.; Lorenzon, G.; Marinelli, C.; D’Incà, R.; et al. Microbiota Changes Induced by Microencapsulated Sodium Butyrate in Patients with Inflammatory Bowel Disease. Neurogastroenterol. Motil. 2020, 32, 13–25. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-Derived Butyrate Dynamically Regulates Intestinal Homeostasis through Regulation of Actin-Associated Protein Synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Gao, Y.; Li, H.; Yang, H.; Su, J.; Huang, L. The Current Novel Therapeutic Regimens for Clostridium Difficile Infection (CDI) and the Potentials of Traditional Chinese Medicine in Treatment of CDI. Crit. Rev. Microbiol. 2019, 45, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Peng, G.; Liu, W.; Xu, H.; Su, J.R. Berberine Blocks the Relapse of Clostridium Difficile Infection in C57BL/6 Mice after Standard Vancomycin Treatment. Antimicrob. Agents Chemother. 2015, 59, 3726. [Google Scholar] [CrossRef]
- Feng, Y.; Li, Y.; Chen, C.; Lin, X.; Yang, Y.; Cai, H.; Lv, Z.; Cao, M.; Li, K.; Xu, J.; et al. Inhibiting Roles of Berberine in Gut Movement of Rodents Are Related to Activation of the Endogenous Opioid System. Phyther. Res. 2013, 27, 1564–1571. [Google Scholar] [CrossRef]
- Sun, H.; Wang, N.; Cang, Z.; Zhu, C.; Zhao, L.; Nie, X.; Cheng, J.; Xia, F.; Zhai, H.; Lu, Y. Modulation of Microbiota-Gut-Brain Axis by Berberine Resulting in Improved Metabolic Status in High-Fat Diet-Fed Rats. Obes. Facts 2017, 9, 365. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, J.; Zhu, S.; He, M.; Ma, S.; Jia, Q.; Sun, Q.; Song, L.; Wang, Y.; Duan, L. Berberine Ameliorates Ovariectomy-Induced Anxiety-like Behaviors by Enrichment in Equol Generating Gut Microbiota. Pharmacol. Res. 2021, 165, 105439. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zheng, J.; Hu, Y.; Hou, H.; Hao, S.; Liu, N.; Wang, Y. Amelioration of Intestinal Barrier Dysfunction by Berberine in the Treatment of Nonalcoholic Fatty Liver Disease in Rats. Pharmacogn. Mag. 2017, 13, 677. [Google Scholar] [CrossRef]
- Li, X.; Su, C.; Jiang, Z.; Yang, Y.; Zhang, Y.; Yang, M.; Zhang, X.; Du, Y.; Zhang, J.; Wang, L.; et al. Berberine Attenuates Choline-Induced Atherosclerosis by Inhibiting Trimethylamine and Trimethylamine-N-Oxide Production via Manipulating the Gut Microbiome. NPJ Biofilms Microbiomes 2021, 7, 36. [Google Scholar] [CrossRef]
- Shu, X.; Li, M.; Cao, Y.; Li, C.; Zhou, W.; Ji, G.; Zhang, L. Berberine Alleviates Non-Alcoholic Steatohepatitis Through Modulating Gut Microbiota Mediated Intestinal FXR Activation. Front. Pharmacol. 2021, 12, 750826. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, Q.; Ma, S.R.; Zhao, Z.X.; Pan, L.B.; Cong, L.; Han, P.; Peng, R.; Yu, H.; Lin, Y.; et al. Oral Berberine Improves Brain Dopa/Dopamine Levels to Ameliorate Parkinson’s Disease by Regulating Gut Microbiota. Signal Transduct. Target. Ther. 2021, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huang, J.; Li, T.; Zhang, S.; Wen, C.; Wang, L. Berberine Ameliorates AGVHD by Gut Microbiota Remodelling, TLR4 Signalling Suppression and Colonic Barrier Repairment for NLRP3 Inflammasome Inhibition. J. Cell. Mol. Med. 2022, 26, 1060. [Google Scholar] [CrossRef]
- Ming, J.; Yu, X.; Xu, X.; Wang, L.; Ding, C.; Wang, Z.; Xie, X.; Li, S.; Yang, W.; Luo, S.; et al. Effectiveness and Safety of Bifidobacterium and Berberine in Human Hyperglycemia and Their Regulatory Effect on the Gut Microbiota: A Multi-Center, Double-Blind, Randomized, Parallel-Controlled Study. Genome Med. 2021, 13, 125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Niu, H.; Zhang, W.; Mu, H.; Sun, C.; Duan, J. Synergy among Thymol, Eugenol, Berberine, Cinnamaldehyde and Streptomycin against Planktonic and Biofilm-Associated Food-Borne Pathogens. Lett. Appl. Microbiol. 2015, 60, 421–430. [Google Scholar] [CrossRef]
- Li, Q.; Wang, B.; Peng, S.; Wei, H.; Li, P.; Leng, Y.; Huang, X. Photodynamic Inactivation of Listeria monocytogenes Using a Natural Aggregation-Induced Emission Photosensitizer and Its Application in Salmon Preservation. LWT 2024, 193, 115762. [Google Scholar] [CrossRef]
- Kim, J.-W.; Mistry, B.; Shin, H.-S.; Kang, S.-S. Anti-Biofilm Activity of N-Mannich Bases of Berberine Linking Piperazine against Listeria Monocytogenes. Food Control 2021, 121, 107668. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, B.; Liu, X.; Liu, W.; Kang, W.; Zhou, J.; Zeng, J. Facile Synthesis of Tetrahydroprotoberberine and Protoberberine Alkaloids from Protopines and Study on Their Antibacterial Activities. Nat. Prod. Res. 2014, 28, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Li, M.; Muhammad, I.; Ma, X.; Chang, Y.; Li, R.; Li, C.; He, J.; Liu, F. Combination of Berberine and Ciprofloxacin Reduces Multi-Resistant Salmonella Strain Biofilm Formation by Depressing MRNA Expressions of LuxS, RpoE, and OmpR. J. Vet. Sci. 2018, 19, 808. [Google Scholar] [CrossRef]
- Jeyakkumar, P.; Zhang, L.; Avula, S.R.; Zhou, C.-H. Design, Synthesis and Biological Evaluation of Berberine-Benzimidazole Hybrids as New Type of Potentially DNA-Targeting Antimicrobial Agents. Eur. J. Med. Chem. 2016, 122, 205–215. [Google Scholar] [CrossRef]
- Aswathanarayan, J.B.; Vittal, R.R. Inhibition of Biofilm Formation and Quorum Sensing Mediated Phenotypes by Berberine in Pseudomonas Aeruginosa and Salmonella typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef]
- Mehta, J.; Rolta, R.; Dev, K. Role of Medicinal Plants from North Western Himalayas as an Efflux Pump Inhibitor against MDR AcrAB-TolC Salmonella enterica Serovar Typhimurium: In Vitro and In Silico Studies. J. Ethnopharmacol. 2022, 282, 114589. [Google Scholar] [CrossRef]
- Mehta, J.; Jandaik, S.U.; Urmila, S. Evaluation of phytochemicals and synergistic interaction between plant extracts and antibiotics for efflux pump inhibitory activity against Salmonella enterica serovar typhimurium strains. Int. J. Pharm. Pharm. Sci. 2016, 8, 217. [Google Scholar] [CrossRef]
- Xu, C.; Wang, F.; Huang, F.; Yang, M.; He, D.; Deng, L. Targeting Effect of Berberine on Type I Fimbriae of Salmonella typhimurium and Its Effective Inhibition of Biofilm. Appl. Microbiol. Biotechnol. 2021, 105, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Naz, F.; Kumar, M.; Koley, T.; Sharma, P.; Haque, M.A.; Kapil, A.; Kumar, M.; Kaur, P.; Ethayathulla, A.S. Screening of Plant-Based Natural Compounds as an Inhibitor of FtsZ from Salmonella typhi Using the Computational, Biochemical and in Vitro Cell-Based Studies. Int. J. Biol. Macromol. 2022, 219, 428–437. [Google Scholar] [CrossRef]
- Kudera, T.; Doskocil, I.; Salmonova, H.; Petrtyl, M.; Skrivanova, E.; Kokoska, L. In Vitro Selective Growth-Inhibitory Activities of Phytochemicals, Synthetic Phytochemical Analogs, and Antibiotics against Diarrheagenic/Probiotic Bacteria and Cancer/Normal Intestinal Cells. Pharmaceuticals 2020, 13, 233. [Google Scholar] [CrossRef]
- Haque, M.A.; Singh, M.; Tripathi, M.K.; Ethayathulla, A.S.; Kaur, P. Identification of Natural Small Molecule Modulators of MurB from Salmonella enterica Serovar Typhi Ty2 Strain Using Computational and Biophysical Approaches. Proteins Struct. Funct. Bioinforma. 2023, 91, 363–379. [Google Scholar] [CrossRef]
- Cui, X.; Liu, X.; Ma, X.; Li, S.; Zhang, J.; Han, R.; Yi, K.; Liu, J.; Pan, Y.; He, D.; et al. Restoring Colistin Sensitivity in Colistin-Resistant Salmonella and Escherichia coli: Combinatorial Use of Berberine and EDTA with Colistin. mSphere 2024, 9, e0018224. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.Y.; Chai, T.-T.; Chow, Y.-L. Evaluation of the Antibacterial Potency of Phytochemicals and Their Effects on Host Xenobiotic Response Using Caenorhabditis elegans. Rev. Bras. Farmacogn. 2024, 34, 1276–1286. [Google Scholar] [CrossRef]
- Saumya, D.; Kumar, D.M.; Anamika. Antibacterial and Antifungal Potential of Berberine Isolated from Stem of Berberis aristata DC. Int. J. DRUG Deliv. Technol. 2022, 13, 454–460. [Google Scholar] [CrossRef]
- Fan, Q.; Wu, J.; Xi, B.; Li, C.; Wang, X.; Li, H. A Screening Model of Antibacterial Agents Based on Escherichia coli Cell-Division Protein. Appl. Sci. 2023, 13, 4493. [Google Scholar] [CrossRef]
- Yan, D.; Li, J.; Xiong, Y.; Zhang, C.; Luo, J.; Han, Y.; Wang, R.; Jin, C.; Qian, H.; Li, J.; et al. Promotion of Quality Standard of Herbal Medicine by Constituent Removing and Adding. Sci. Rep. 2014, 4, 3668. [Google Scholar] [CrossRef]
- Fukamachi, H.; Matsumoto, C.; Omiya, Y.; Arimoto, T.; Morisaki, H.; Kataoka, H.; Kadena, M.; Funatsu, T.; Fukutake, M.; Kase, Y.; et al. Effects of Hangeshashinto on Growth of Oral Microorganisms. Evid. -Based Complement. Altern. Med. 2015, 2015, 512947. [Google Scholar] [CrossRef]
- Kumar, G.S.; Hazra, S. Sanguinarine, a Promising Anticancer Therapeutic: Photochemical and Nucleic Acid Binding Properties. RSC Adv. 2014, 4, 56518–56531. [Google Scholar] [CrossRef]
- Ahsan, H.; Reagan-Shaw, S.; Breur, J.; Ahmad, N. Sanguinarine Induces Apoptosis of Human Pancreatic Carcinoma AsPC-1 and BxPC-3 Cells via Modulations in Bcl-2 Family Proteins. Cancer Lett. 2007, 249, 198–208. [Google Scholar] [CrossRef]
- Niu, X.; Fan, T.; Li, W.; Xing, W.; Huang, H. The Anti-Inflammatory Effects of Sanguinarine and Its Modulation of Inflammatory Mediators from Peritoneal Macrophages. Eur. J. Pharmacol. 2012, 689, 262–269. [Google Scholar] [CrossRef]
- Danielewski, M.; Zielińska, S.; Merwid-Ląd, A.; Szandruk-Bender, M.; Słupski, W.; Włodarczyk, M.; Sozański, T.; Ziółkowski, P.; Szeląg, A.; Nowak, B. Sanguinarine–Chelerythrine from Coptis chinensis Offers Analgesic and Anti-Inflammatory Effects Without Gastrotoxicity. Pharmaceutics 2025, 17, 323. [Google Scholar] [CrossRef]
- Obiang-Obounou, B.W.; Kang, O.H.; Choi, J.G.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Shin, D.W.; Kim, K.W.; Park, C.B.; Kim, Y.G.; et al. The Mechanism of Action of Sanguinarine against Methicillin-Resistant Staphylococcus Aureus. J. Toxicol. Sci. 2011, 36, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Miao, F.; Yao, Y.; Cao, F.J.; Yang, R.; Ma, Y.N.; Qin, B.F.; Zhou, L. In Vitro Antifungal Activity of Sanguinarine and Chelerythrine Derivatives against Phytopathogenic Fungi. Molecules 2012, 17, 13026–13035. [Google Scholar] [CrossRef] [PubMed]
- Achkar, I.W.; Mraiche, F.; Mohammad, R.M.; Uddin, S. Anticancer Potential of Sanguinarine for Various Human Malignancies. Future Med. Chem. 2017, 9, 933–950. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.L.; Shi, Y.N.; Cao, Y.L.; Tan, X.; Zeng, Y.L.; Luo, S.T.; Li, Y.M.; Qin, L.; Xia, B.H.; Fu, R.G.; et al. Sanguinarine Protects against Indomethacin-Induced Small Intestine Injury in Rats by Regulating the Nrf2/NF-ΚB Pathways. Front. Pharmacol. 2022, 13, 960140. [Google Scholar] [CrossRef]
- Li, X.; Wu, X.; Wang, Q.; Xu, W.; Zhao, Q.; Xu, N.; Hu, X.; Ye, Z.; Yu, S.; Liu, J.; et al. Sanguinarine Ameliorates DSS Induced Ulcerative Colitis by Inhibiting NLRP3 Inflammasome Activation and Modulating Intestinal Microbiota in C57BL/6 Mice. Phytomedicine 2022, 104, 154394, Erratum in Phytomedicine 2022, 106, 154394. [Google Scholar] [CrossRef]
- Qu, Q.; Huang, X.; Zhu, Z.; Wang, J.; Zhao, M.; Cui, W.; Zheng, Y.; Liu, Y.; Chen, X.; Zhang, Z.; et al. Targeting Membrane Integrity and Imidazoleglycerol-Phosphate Dehydratase: Sanguinarine Multifaceted Approach against Staphylococcus Aureus Biofilms. Phytomedicine 2025, 138, 156428. [Google Scholar] [CrossRef]
- Gu, J.; Zhao, L.; Chen, Y.Z.; Guo, Y.X.; Sun, Y.; Guo, Q.; Duan, G.X.; Li, C.; Tang, Z.B.; Zhang, Z. Preventive Effect of Sanguinarine on Intestinal Injury in Mice Exposed to Whole Abdominal Irradiation. Biomed. Pharmacother 2022, 46, 112496. [Google Scholar] [CrossRef]
- Zhao, M.A.; Chu, J.; Feng, S.; Guo, C.; Xue, B.; He, K.; Li, L. Immunological mechanisms of inflammatory diseases caused by gut microbiota dysbiosis: A review. Biomed. Pharmacother. 2023, 164, 114985. [Google Scholar] [CrossRef]
- Jian, Y.; Zhang, D.; Liu, M.; Wang, Y.; Xu, Z.X. The Impact of Gut Microbiota on Radiation-Induced Enteritis. Front. Cell. Infect. Microbiol. 2021, 11, 586392. [Google Scholar] [CrossRef]
- Osei-Owusu, H.; Rondevaldova, J.; Houdkova, M.; Kudera, T.; Needham, T.; Mascellani, A.; Kokoska, L. Evaluation of In Vitro Synergistic Effects of Tetracycline with Alkaloid-Related Compounds against Diarrhoeic Bacteria. Int. J. Mol. Sci. 2024, 25, 6038. [Google Scholar] [CrossRef]
- Osei-Owusu, H.; Kudera, T.; Strakova, M.; Rondevaldova, J.; Skrivanova, E.; Novy, P.; Kokoska, L. In Vitro Selective Combinatory Effect of Ciprofloxacin with Nitroxoline, Sanguinarine, and Zinc Pyrithione against Diarrhea-Causing and Gut Beneficial Bacteria. Microbiol. Spectr. 2022, 10, e0106322. [Google Scholar] [CrossRef]
- Zhu, L.; Qu, S.; Wang, X.; Huang, H.; Li, H.; Zuo, L. Novel 2,3-Dihydro-1H-Imidazo[2,1-a]Isoquinolin-4-Ium Salts: Synthesis, Antitumor and Antibacterial Activity. Acta Pol. Pharm.-Drug Res. 2022, 79, 89–96. [Google Scholar] [CrossRef]
- Gobato, R.; Gobato, A.; Fedrigo, D.F.G. Allocryptopine, Berberine, Chelerythrine, Copsitine, Dihydrosanguinarine, Protopine and Sanguinarine. Molecular Geometry of the Main Alkaloids Found in the Seeds of Argemone Mexicana Linn. Parana J. Ofsci. Educ. 2015, 1, 8–16. [Google Scholar]
- Tolkachev, O.N.; Savina, A.A.; Sheichenko, V.I.; Proskudina, V.V. On the mechanism of thermolysis of the benzo[c]phenanthridine alkaloid chelerythrine. Pharm. Chem. J. 1999, 33, 37–38. [Google Scholar] [CrossRef]
- Klejdus, B.; Lojková, L.; Kosina, P.; Ulrichová, J.; Šimánek, V.; Kubáň, V. Liquid Chromatographic/Electrospray Mass Spectrometric Determination (LC/ESI-MS) of Chelerythrine and Dihydrochelerythrine in near-Critical CO2 Extracts from Real and Spiked Plasma Samples. Talanta 2007, 72, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Pan, Y.; Zhang, G.; Wu, Y.; Zhong, W.; Chu, C.; Qian, Y.; Zhu, G. Chelerythrine Inhibits Human Hepatocellular Carcinoma Metastasis in Vitro. Biol. Pharm. Bull. 2018, 41, 36–46. [Google Scholar] [CrossRef]
- Assaf, A.M.; Haddadin, R.N.; Aldouri, N.A.; Alabbassi, R.; Mashallah, S.; Mohammad, M.; Bustanji, Y. Anti-Cancer, Anti-Inflammatory and Anti-Microbial Activities of Plant Extracts Used against Hematological Tumors in Traditional Medicine of Jordan. J. Ethnopharmacol. 2013, 145, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Li, W.F.; Hao, D.J.; Fan, T.; Huang, H.M.; Yao, H.; Niu, X.F. Protective Effect of Chelerythrine against Ethanol-Induced Gastric Ulcer in Mice. Chem. Biol. Interact. 2014, 208, 18–27. [Google Scholar] [CrossRef]
- Qian, W.; Sun, Z.; Fu, Y.; Yang, M.; Wang, T.; Li, Y. Efficacy of Chelerythrine against Dual-Species Biofilms of Staphylococcus Aureus and Staphylococcus Lugdunensis. 3 Biotech. 2020, 10, 427. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Cheng, H.; Li, Y.; Zhou, G. Chelerythrine, a Major Ingredient Isolated from Macleaya cordata (Willd.) R. Br. (Papaveraceae), Inhibits Fluconazole-Resistant Candida albicans Biofilms. J. Herb. Med. 2023, 42, 100752. [Google Scholar] [CrossRef]
- Oshima, T.; Pavlick, K.P.; Laroux, F.S.; Verma, S.K.; Jordan, P.; Grisham, M.B.; Williams, L.; Alexander, J.S. Regulation and Distribution of MAdCAM-1 in Endothelial Cells in Vitro. Am. J. Physiol.-Cell Physiol. 2001, 281, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Epple, H.J.; Mankertz, J.; Ignatius, R.; Liesenfeld, O.; Fromm, M.; Zeitz, M.; Chakraborty, T.; Schulzke, J.D. Aeromonas Hydrophila Beta-Hemolysin Induces Active Chloride Secretion in Colon Epithelial Cells (HT-29/B6). Infect. Immun. 2004, 72, 4848. [Google Scholar] [CrossRef] [PubMed]
- Agbor, T.A.; Demma, Z.C.; Mumy, K.L.; Bien, J.D.; McCormick, B.A. The ERM Protein, Ezrin, Regulates Neutrophil Transmigration by Modulating the Apical Localization of MRP2 in Response to the SipA Effector Protein during Salmonella typhimurium Infection. Cell Microbiol. 2011, 12, 2007–2021. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, F.; Li, H.; Liu, J.; Jiang, Y.; Ren, F.; Huang, L.; Yuan, X.; Li, Y.; Yang, W.; et al. The Combination of Macleaya Extract and Glucose Oxidase Improves the Growth Performance, Antioxidant Capacity, Immune Function and Cecal Microbiota of Piglets. Front. Vet. Sci. 2023, 10, 2–11. [Google Scholar] [CrossRef]
- Lei, F.; Liu, X.; Huang, H.; Fu, S.; Zou, K.; Zhang, S.; Zhou, L.; Zeng, J.; Liu, H.; Jiang, L.; et al. The Macleaya cordata Symbiont: Revealing the Effects of Plant Niches and Alkaloids on the Bacterial Community. Front. Microbiol. 2021, 12, 681210. [Google Scholar] [CrossRef]
- Quinteros, J.A.; Scott, P.C.; Wilson, T.B.; Anwar, A.M.; Scott, T.; Muralidharan, C.; Van, T.T.H.; Moore, R.J. Isoquinoline Alkaloids Induce Partial Protection of Laying Hens from the Impact of Campylobacter Hepaticus (Spotty Liver Disease) Challenge. Poult. Sci. 2021, 100, 101423. [Google Scholar] [CrossRef] [PubMed]
- Mickdam, E.; Khiaosa-ard, R.; Metzler-Zebeli, B.U.; Klevenhusen, F.; Chizzola, R.; Zebeli, Q. Rumen Microbial Abundance and Fermentation Profile during Severe Subacute Ruminal Acidosis and Its Modulation by Plant Derived Alkaloids in Vitro. Anaerobe 2016, 39, 4–13. [Google Scholar] [CrossRef]
- Li, S.; Wang, N.; Tan, H.; Chueng, F.; Zhang, Z.; Yuen, M.; Feng, Y. Modulation of Gut Microbiota Mediates Berberine-induced Expansion of Immuno-suppressive Cells to against Alcoholic Liver Disease. Clin. Transl. Med. 2020, 10, e112. [Google Scholar] [CrossRef]
- Hu, N.X.; Chen, M.; Liu, Y.S.; Shi, Q.; Yang, B.; Zhang, H.C.; Cheng, P.; Tang, Q.; Liu, Z.Y.; Zeng, J.G. Pharmacokinetics of Sanguinarine, Chelerythrine, and Their Metabolites in Broiler Chickens Following Oral and Intravenous Administration. J. Vet. Pharmacol. Ther. 2019, 42, 197–206. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Yao, H.; Mu, Q.; Zhao, G.; Li, Y.; Hu, H.; Niu, X. Pharmacokinetic and Anti-Inflammatory Effects of Sanguinarine Solid Lipid Nanoparticles. Inflammation 2014, 37, 632–638. [Google Scholar] [CrossRef]
- Rosen, J.; Landriscina, A.; Adler, B.L.; Krauz, A.; Doerner, J.; Navati, M.; Musaev, T.; Gravekamp, C.; Nosanchuk, J.; Friedman, A.J. Characterization and Assessment of Nanoencapsulated Sanguinarine Chloride as a Potential Treatment for Melanoma. J. Drugs Dermatol. 2015, 14, 453–458. [Google Scholar] [PubMed]
- Liu, S.; Tao, Y.; Wu, S.; Lin, J.; Fu, S.; Lu, J.; Zhang, J.; Fu, B.; Zhang, E.; Xu, J.; et al. Sanguinarine Chloride Induces Ferroptosis by Regulating ROS/BACH1/HMOX1 Signaling Pathway in Prostate Cancer. Chin. Med. 2024, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Lin, S.; Chen, K.; Yin, S.; Peng, H.; Cai, N.; Ma, W.; Zhou, S.; Huang, Y. Natural Product Library Screens Identify Sanguinarine Chloride as a Potent Inhibitor of Telomerase Expression and Activity. Cells 2022, 11, 1485. [Google Scholar] [CrossRef] [PubMed]
- Walterová, D.; Ulrichová, J.; Válka, I.; Vicar, J.; Vavrecková, C.; Táborská, E.; Harjrader, R.J.; Meyer, D.L.; Cerná, H.; Simánek, V. Benzo[c]Phenanthridine Alkaloids Sanguinarine and Chelerythrine: Biological Activities and Dental Care Applications. Acta Univ. Palacki. Olomuc. Fac. Med. 1995, 139, 7–16. [Google Scholar]
- Croaker, A.; King, G.J.; Pyne, J.H.; Anoopkumar-Dukie, S.; Liu, L. Sanguinaria Canadensis: Traditional Medicine, Phytochemical Composition, Biological Activities and Current Uses. Int. J. Mol. Sci. 2016, 17, 1414. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, J.; Fan, Y.; Long, H.; Liang, M.; Chen, Q.; Wang, Z.; Wu, C.; Wang, Y. Macrophage-Targeted Berberine-Loaded β-Glucan Nanoparticles Enhance the Treatment of Ulcerative Colitis. Int. J. Nanomed. 2022, 17, 5303–5314. [Google Scholar] [CrossRef]
- Feng, X.; Xie, Q.; Xu, H.; Zhang, T.; Li, X.; Tian, Y.; Lan, H.; Kong, L.; Zhang, Z. Yeast Microcapsule Mediated Natural Products Delivery for Treating Ulcerative Colitis through Anti-Inflammatory and Regulation of Macrophage Polarization. ACS Appl. Mater. Interfaces 2022, 14, 31085–31098. [Google Scholar] [CrossRef]
- Andima, M.; Boese, A.; Paul, P.; Koch, M.; Loretz, B.; Lehr, C.-M. Targeting Intracellular Bacteria with Dual Drug-Loaded Lactoferrin Nanoparticles. ACS Infect. Dis. 2024, 10, 1696–1710. [Google Scholar] [CrossRef]


 ) and increases the level of anti-inflammatory cytokines (green square
) and increases the level of anti-inflammatory cytokines (green square  ) [181,187,189]. (C) SAN strengthens the intestinal barrier function through improving the expression of tight junction proteins, especially ZO-1 [186].
) and increases the level of anti-inflammatory cytokines (green square ) [181,187,189]. (C) SAN strengthens the intestinal barrier function through improving the expression of tight junction proteins, especially ZO-1 [186].
) [181,187,189]. (C) SAN strengthens the intestinal barrier function through improving the expression of tight junction proteins, especially ZO-1 [186].
) and increases the level of anti-inflammatory cytokines (green square ) [181,187,189]. (C) SAN strengthens the intestinal barrier function through improving the expression of tight junction proteins, especially ZO-1 [186].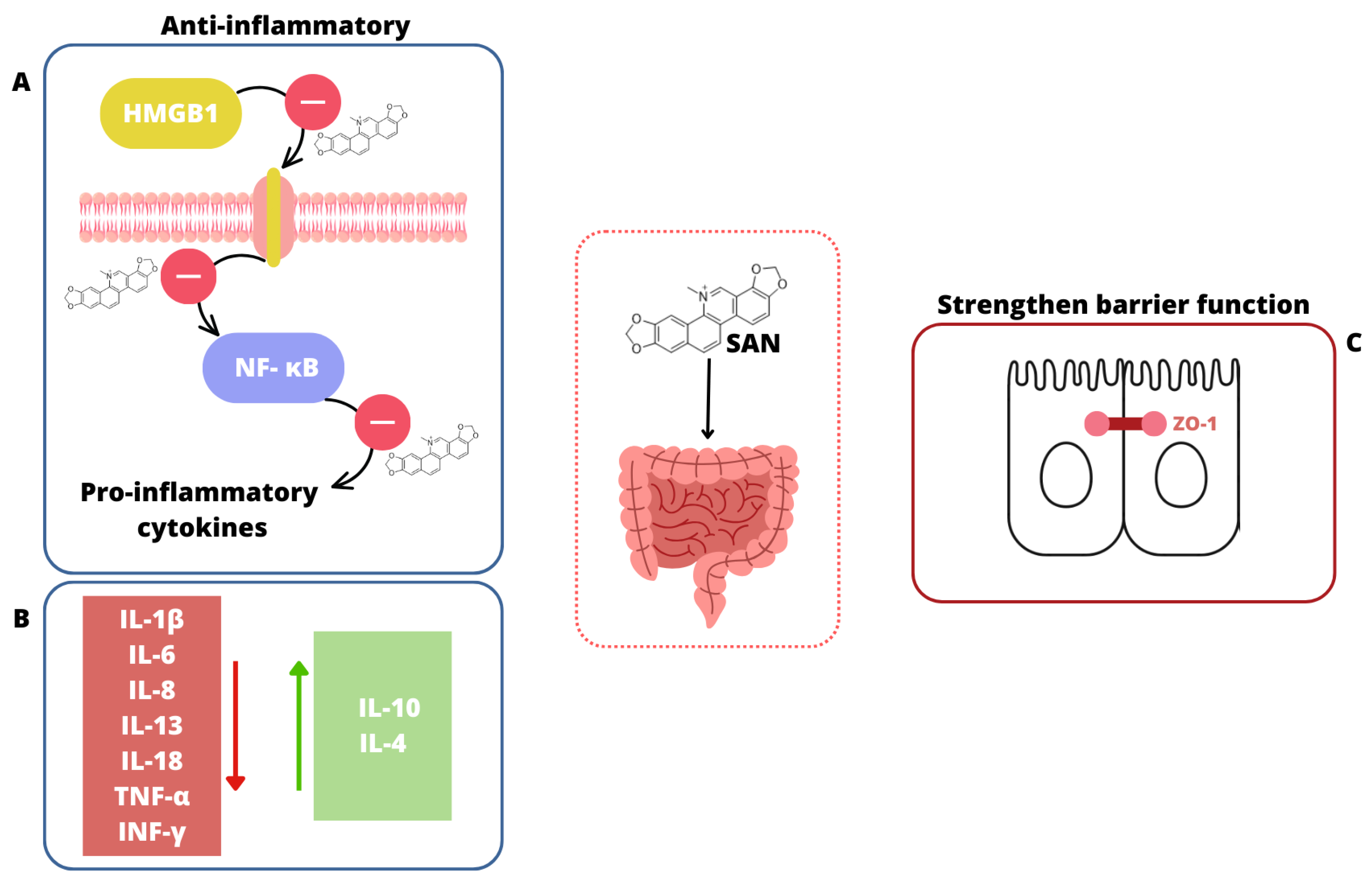

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Berberine | Chelerythrine | Sanguinarine | |
|---|---|---|---|
| Microbiome Protein Composition | Enrichment of proteins from specific bacteria: Verrucomicrobia, Proteobacteria 2, the Bacteroidetes phylum, the Akkermansia genus, and the Verrucomicrobia phylum 1 | Enrichment of proteins from the Bacteroides genus and the Akkermansia genus | |
| Activation of Bacterial Defense Mechanisms | Leads to the upregulation of proteins associated with microbial defense and stress responses. This indicates that BBR induces a protective or stress response in the gut microbiome. | Activates protective bacterial mechanisms. This is evidenced by the increase in proteins related to microbial defense and stress responses. These include proteins associated with the cell wall/membrane/envelope biogenesis and signal transduction mechanisms. | Triggers bacterial protective mechanisms, as indicated by the increased abundance of proteins involved in microbial defense and stress responses. This includes proteins related to cell wall/membrane/envelope biogenesis, signal transduction mechanisms, and other stress-related functions. |
| Berberine | |||
|---|---|---|---|
| Antibiotic/Chemotherapeutic | Type of Interaction | Pathogen Name | References |
| cyprofloxacin | synergistic | S. enterica subsp. enterica serovar Gallinarum | [164] |
| colistin | MDR S. enterica | [173] | |
| streptomycin | additive | L. monocytogenes CMCC 54004 | [160] |
| S. enterica subsp. enterica serovar Typhimurium SL1344 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duda-Madej, A.; Viscardi, S.; Stecko, J.; Szymańska, N.; Topola, E.; Pacyga, K.; Szandruk-Bender, M. Can Nature Overcome Invasive Gastrointestinal Infections? Int. J. Mol. Sci. 2025, 26, 5795. https://doi.org/10.3390/ijms26125795
Duda-Madej A, Viscardi S, Stecko J, Szymańska N, Topola E, Pacyga K, Szandruk-Bender M. Can Nature Overcome Invasive Gastrointestinal Infections? International Journal of Molecular Sciences. 2025; 26(12):5795. https://doi.org/10.3390/ijms26125795
Chicago/Turabian StyleDuda-Madej, Anna, Szymon Viscardi, Jakub Stecko, Natalia Szymańska, Ewa Topola, Katarzyna Pacyga, and Marta Szandruk-Bender. 2025. "Can Nature Overcome Invasive Gastrointestinal Infections?" International Journal of Molecular Sciences 26, no. 12: 5795. https://doi.org/10.3390/ijms26125795
APA StyleDuda-Madej, A., Viscardi, S., Stecko, J., Szymańska, N., Topola, E., Pacyga, K., & Szandruk-Bender, M. (2025). Can Nature Overcome Invasive Gastrointestinal Infections? International Journal of Molecular Sciences, 26(12), 5795. https://doi.org/10.3390/ijms26125795